
HSD3B1 (c.1100C) Genotype Is Associated with Distinct Tumoral and Clinical Outcomes in Breast and Endometrial Cancers

 , , , , , , , , ,
, , , , , , , , , 
Abstract
1. Introduction
2. Results
2.1. HSD3B1 Genotypes and Demographics Data
2.2. HSD3B1 Variants Have Limited Impact on Survival Outcomes in BC and EC
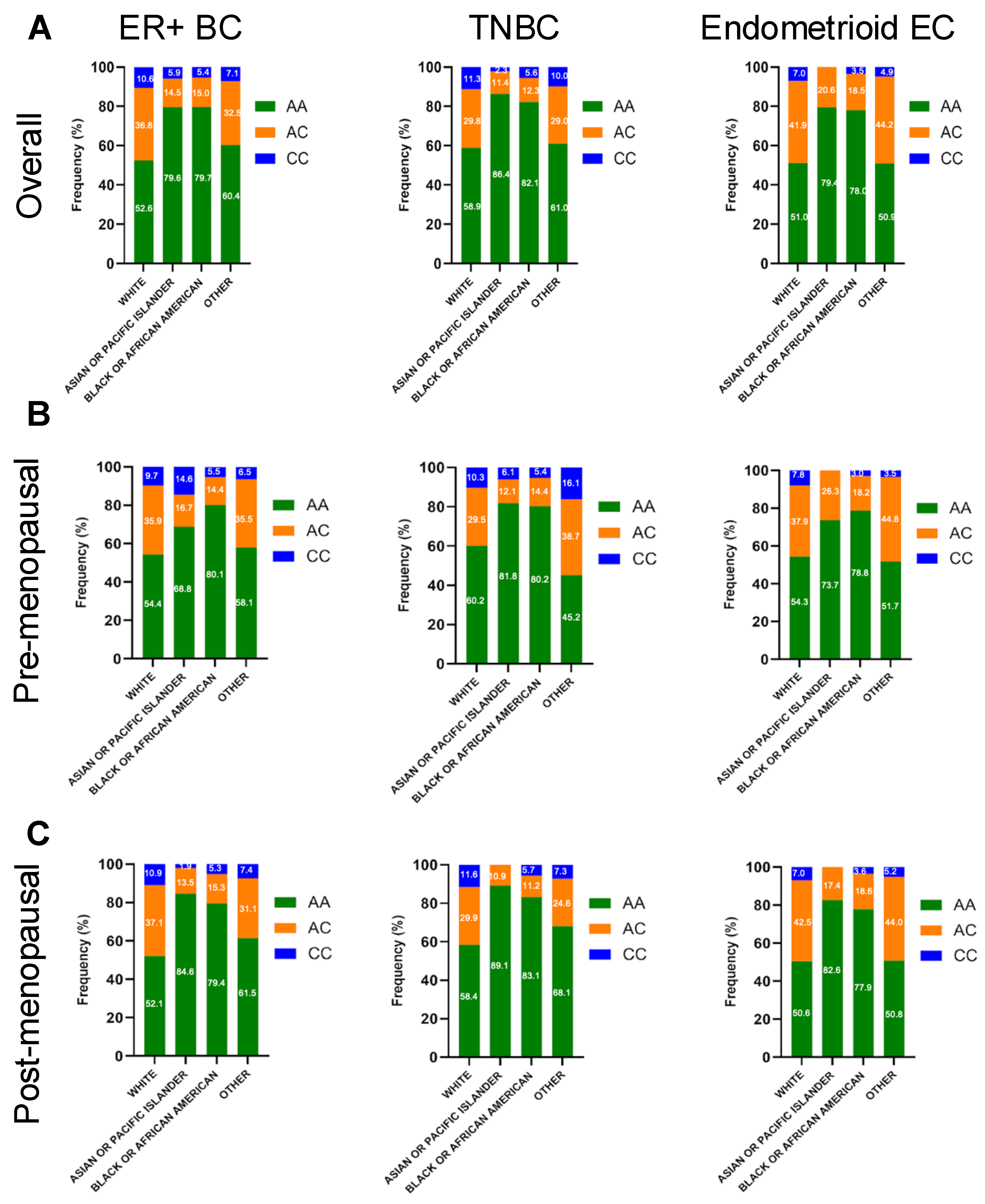
2.3. HSD3B1 Genotypes Exhibit Different Distributions by Race
2.4. HSD3B1 Variants Are Associated with Somatic Alterations in BC and EC
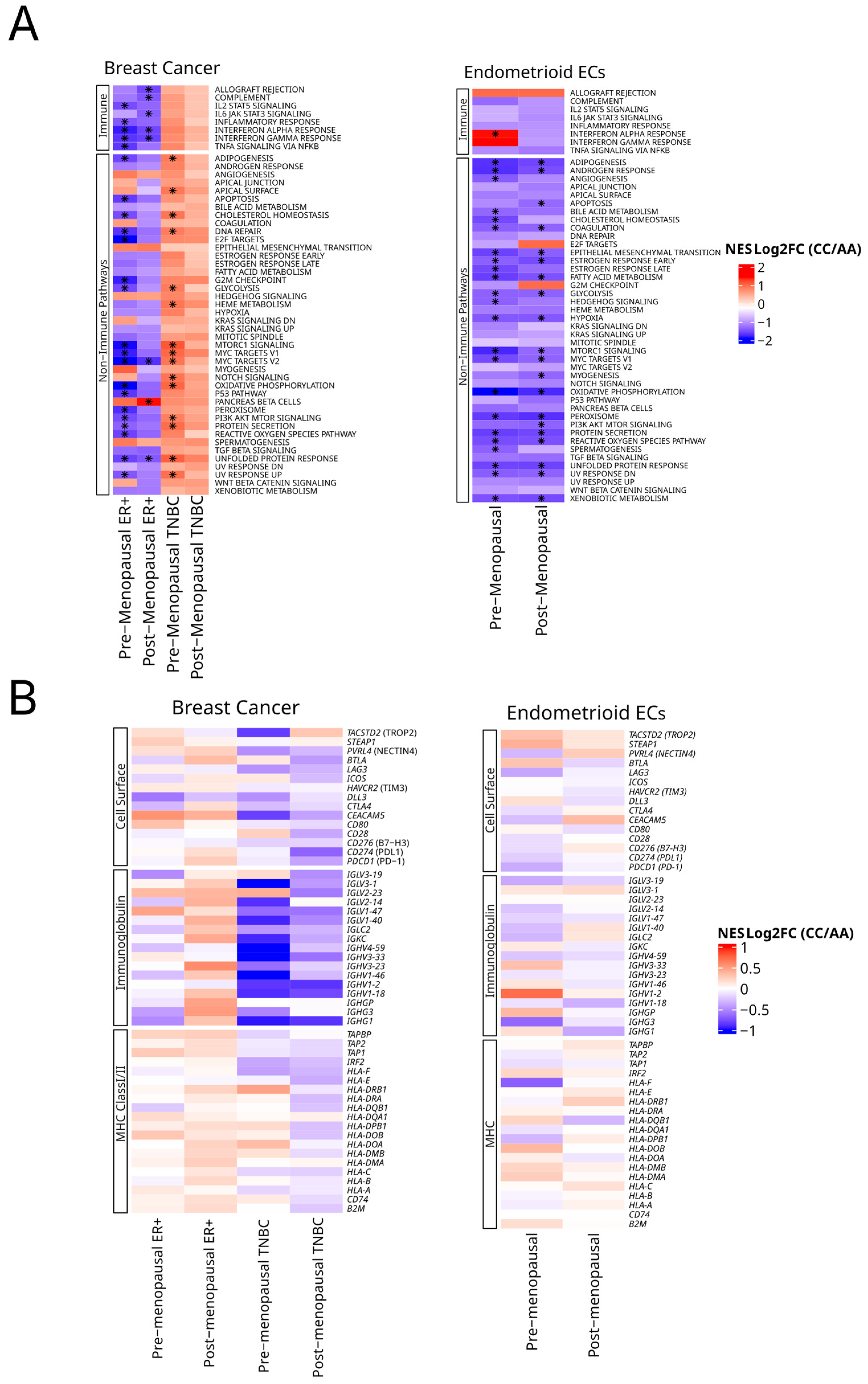
2.5. Differences in Gene Expression Across HSD3B1 Genotypes Is Observed in BC and EC
2.6. HSD3B1 Regulates Hallmark Signaling Pathways and Immune-Related Genes
3. Discussion
Limitations of the Study
4. Materials and Methods
4.1. Patient Specimens
4.2. Patient Cohort and HSD3B1 Genotype
4.3. Whole-Exome Sequencing and NGS
4.4. Whole-Transcriptome Sequencing
4.5. Immune Signatures
4.6. Statistical Analysis
4.7. TMB and MSI/dMMR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3βHSD1 | 3β-Hydroxysteroid Dehydrogenase Type 1 |
| AR | Androgen Receptor |
| BC | Breast Cancer |
| CAN | Copy Number Amplification |
| DHEA | Dehydroepiandrosterone |
| DHT | Dihydrotestosterone |
| EC | Endometrial Cancer |
| ER | Estrogen Receptor |
| GSEA | Gene Set Enrichment Analysis |
| HSD3B1 | Hydroxy-delta-5-steroid Dehydrogenase, 3 Beta-and Steroid Delta-Isomerase 1 |
| IHC | Immunohistochemistry |
| MMR | Mismatch Repair |
| MSI | Microsatellite Instability |
| OS | Overall Survival |
| PI | Pacific Islanders |
| PR | Progesterone Receptor |
| TMB | Tumor Mutational Burden |
| TNBC | Triple-negative Breast Cancer |
References
- Naelitz, B.D.; Sharifi, N. Through the Looking-Glass: Reevaluating DHEA Metabolism Through HSD3B1 Genetics. Trends Endocrinol. Metab. 2020, 31, 680–690. [Google Scholar] [CrossRef] [PubMed]
- Freitas, P.F.S.; Abdshah, A.; McKay, R.R.; Sharifi, N. HSD3B1, Prostate Cancer Mortality and Modifiable Outcomes. Nat. Rev. Urol. 2024, 22, 313–320. [Google Scholar] [CrossRef]
- Thomas, L.; Sharifi, N. Germline HSD3B1 Genetics and Prostate Cancer Outcomes. Urology 2020, 145, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-H.; Li, R.; Kuri, B.; Lotan, Y.; Roehrborn, C.G.; Liu, J.; Vessella, R.; Nelson, P.S.; Kapur, P.; Guo, X.; et al. A Gain-of-Function Mutation in DHT Synthesis in Castration-Resistant Prostate Cancer. Cell 2013, 154, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Hearn, J.W.D.; AbuAli, G.; Reichard, C.A.; Reddy, C.A.; Magi-Galluzzi, C.; Chang, K.-H.; Carlson, R.; Rangel, L.; Reagan, K.; Davis, B.J.; et al. HSD3B1 and Resistance to Androgen-Deprivation Therapy in Prostate Cancer: A Retrospective, Multicohort Study. Lancet Oncol. 2016, 17, 1435–1444. [Google Scholar] [CrossRef]
- McKay, R.R.; Nelson, T.J.; Pagadala, M.S.; Teerlink, C.C.; Gao, A.; Bryant, A.K.; Agiri, F.Y.; Guram, K.; Thompson, R.F.; Pridgen, K.M.; et al. Adrenal-Permissive Germline HSD3B1 Allele and Prostate Cancer Outcomes. JAMA Netw. Open 2024, 7, e242976. [Google Scholar] [CrossRef]
- Saha Roy, S.; Vadlamudi, R.K. Role of Estrogen Receptor Signaling in Breast Cancer Metastasis. Int. J. Breast Cancer 2012, 2012, 654698. [Google Scholar] [CrossRef]
- Flanagan, M.R.; Sharifi, N.; Gadi, V.K. ASO Author Reflections: Inheritance of Adrenal Permissive HSD3B1 Genotype Negatively Impacts Outcomes in Hormone Receptor-Positive Postmenopausal Breast Cancer. Ann. Surg. Oncol. 2022, 29, 7202–7203. [Google Scholar] [CrossRef]
- McManus, J.M.; Vargas, R.; Bazeley, P.S.; Schumacher, F.R.; Sharifi, N. Association Between Adrenal-Restrictive HSD3B1 Inheritance and Hormone-Independent Subtypes of Endometrial and Breast Cancer. JNCI Cancer Spectr. 2022, 6, pkac061. [Google Scholar] [CrossRef]
- Kellen, S.; Makovec, A.; Miller, C.D.; Nazari, S.S.; Elliott, A.; Deacon, A.; John, E.; Vobugari, N.; Agarwal, N.; McKay, R.R.; et al. The Influence of the Germline HSD3B1 Adrenal-Permissive Allele (c.1100 C) on the Somatic Alteration Landscape, the Transcriptome, and Immune Cell Infiltration in Prostate Cancer. Cancers 2025, 17, 1270. [Google Scholar] [CrossRef]
- Rs1047303 RefSNP Report-dbSNP-NCBI. Available online: https://www.ncbi.nlm.nih.gov/snp/rs1047303?horizontal_tab=true#frequency_tab (accessed on 23 April 2025).
- Li, J.; Shi, D.; Li, S.; Shi, X.; Liu, Y.; Zhang, Y.; Wang, G.; Zhang, C.; Xia, T.; Piao, H.; et al. KEAP1 Promotes Anti-Tumor Immunity by Inhibiting PD-L1 Expression in NSCLC. Cell Death Dis. 2024, 15, 175. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kang, J.; Jung, E.S.; Lee, A. High Expression of NRF2 and Low Expression of KEAP1 Predict Worse Survival in Patients With Operable Triple-Negative Breast Cancer. J. Breast Cancer 2023, 26, 461–478. [Google Scholar] [CrossRef]
- West, N.R.; Watson, P.H. S100A7 (Psoriasin) is Induced by the Proinflammatory Cytokines Oncostatin-M and Interleukin-6 in Human Breast Cancer. Oncogene 2010, 29, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, H.; Kang, Y.-W.; Kim, Y.; Park, M.-Y.; Song, J.-H.; Jo, Y.; Dao, T.; Ryu, D.; Lee, J.; et al. LY6D Is Crucial for Lipid Accumulation and Inflammation in Nonalcoholic Fatty Liver Disease. Exp. Mol. Med. 2023, 55, 1479–1491. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Lv, X.; Li, Y.; Li, M. Attention-Based GCN Integrates Multi-Omics Data for Breast Cancer Subtype Classification and Patient-Specific Gene Marker Identification. Brief. Funct. Genom. 2023, 22, 463–474. [Google Scholar] [CrossRef]
- Mustacchi, G.; Sormani, M.P.; Bruzzi, P.; Gennari, A.; Zanconati, F.; Bonifacio, D.; Monzoni, A.; Morandi, L. Identification and Validation of a New Set of Five Genes for Prediction of Risk in Early Breast Cancer. Int. J. Mol. Sci. 2013, 14, 9686–9702. [Google Scholar] [CrossRef]
- Mou, M.A.; Keya, N.A.; Islam, M.; Hossain, M.J.; Al Habib, M.S.; Alam, R.; Rana, S.; Samad, A.; Ahammad, F. Validation of CSN1S1 Transcriptional Expression, Promoter Methylation, and Prognostic Power in Breast Cancer Using Independent Datasets. Biochem. Biophys. Rep. 2020, 24, 100867. [Google Scholar] [CrossRef]
- Song, Y.; Li, Z.; Li, L.; Zhou, H.; Zeng, T.-T.; Jin, C.; Lin, J.-R.; Gao, S.; Li, Y.; Guan, X.-Y.; et al. SERPINA11 Inhibits Metastasis in Hepatocellular Carcinoma by Suppressing MEK/ERK Signaling Pathway. J. Hepatocell. Carcinoma 2021, 8, 759–771. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Flanagan, M.R.; Doody, D.R.; Voutsinas, J.; Wu, Q.; Banda, K.; Sharifi, N.; Li, C.I.; Gadi, V.K. Association of HSD3B1 Genotype and Clinical Outcomes in Postmenopausal Estrogen-Receptor-Positive Breast Cancer. Ann. Surg. Oncol. 2022, 29, 7194–7201. [Google Scholar] [CrossRef]
- Kruse, M.L.; Patel, M.; McManus, J.; Chung, Y.-M.; Li, X.; Wei, W.; Bazeley, P.S.; Nakamura, F.; Hardaway, A.; Downs, E.; et al. Adrenal-Permissive HSD3B1 Genetic Inheritance and Risk of Estrogen-Driven Postmenopausal Breast Cancer. JCI Insight 2021, 6, e150403. [Google Scholar] [CrossRef]
- University of Washington, Association of HSD3B1 Genotype with Response to Preoperative Letrozole Therapy Among Postmenopausal Women with Estrogen-Receptor Positive (ER+) HER2/Neu-Negative (HER2-) Invasive Carcinomas of the Breast; clinicaltrials.gov. 2025. Available online: https://clinicaltrials.gov/study/NCT05183828?term=hsd3b1&cond=breast%20cancer&rank=1 (accessed on 12 June 2025).
- Gabay, M.; Li, Y.; Felsher, D.W. MYC Activation Is a Hallmark of Cancer Initiation and Maintenance. Cold Spring Harb. Perspect. Med. 2014, 4, a014241. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Hou, Z.; Huang, S.; Mei, Z.; Chen, L.; Guo, J.; Gao, Y.; Zhuang, Q.; Zhang, X.; Tan, Q.; Yang, T.; et al. Inhibiting 3βHSD1 to Eliminate the Oncogenic Effects of Progesterone in Prostate Cancer. Cell Rep. Med. 2022, 3, 100561. [Google Scholar] [CrossRef]
- Chesner, L.N.; Polesso, F.; Graff, J.N.; Hawley, J.E.; Smith, A.K.; Lundberg, A.; Das, R.; Shenoy, T.; Sjöström, M.; Zhao, F.; et al. Androgen Receptor Inhibition Increases MHC Class I Expression and Improves Immune Response in Prostate Cancer. Cancer Discov. 2025, 15, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Polesso, F.; Wang, C.; Sehrawat, A.; Hawkins, R.M.; Murray, S.E.; Thomas, G.V.; Caruso, B.; Thompson, R.F.; Wood, M.A.; et al. Androgen Receptor Activity in T Cells Limits Checkpoint Blockade Efficacy. Nature 2022, 606, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Yang, J.C.; Chen, B.; Nip, C.; Van Dyke, J.E.; Zhang, X.; Chen, H.-W.; Evans, C.P.; Murphy, W.J.; Liu, C. Androgen Receptor Blockade Resistance with Enzalutamide in Prostate Cancer Results in Immunosuppressive Alterations in the Tumor Immune Microenvironment. J. Immunother. Cancer 2023, 11, e006581. [Google Scholar] [CrossRef]
- Poitout, V.; Robertson, R.P. Glucolipotoxicity: Fuel Excess and Beta-Cell Dysfunction. Endocr. Rev. 2008, 29, 351–366. [Google Scholar] [CrossRef]
- Faustin, B.; Lartigue, L.; Bruey, J.-M.; Luciano, F.; Sergienko, E.; Bailly-Maitre, B.; Volkmann, N.; Hanein, D.; Rouiller, I.; Reed, J.C. Reconstituted NALP1 Inflammasome Reveals Two-Step Mechanism of Caspase-1 Activation. Mol. Cell 2007, 25, 713–724. [Google Scholar] [CrossRef]
- Li, Z.; Wei, H.; Li, S.; Wu, P.; Mao, X. The Role of Progesterone Receptors in Breast Cancer. Drug Des. Dev. Ther. 2022, 16, 305–314. [Google Scholar] [CrossRef]
- Rothlin, C.V.; Ghosh, S. Lifting the Innate Immune Barriers to Antitumor Immunity. J. Immunother. Cancer 2020, 8, e000695. [Google Scholar] [CrossRef] [PubMed]
- Weiss, L.K.; Burkman, R.T.; Cushing-Haugen, K.L.; Voigt, L.F.; Simon, M.S.; Daling, J.R.; Norman, S.A.; Bernstein, L.; Ursin, G.; Marchbanks, P.A.; et al. Hormone Replacement Therapy Regimens and Breast Cancer Risk. Obstet. Gynecol. 2002, 100, 1148. [Google Scholar] [PubMed]
- Chang, C.-H.; Lee, Y.-C.; Huang, C.-W.; Lu, Y.-M. Is Tamoxifen Good Enough for the Asian Population in ER+ HER2- Post-Menopausal Women with Early Breast Cancer? A Nationwide Population-Based Cohort Study. PLoS ONE 2024, 19, e0313120. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.C.; Blanchard, Z.; Maurer, K.A.; Gertz, J. Estrogen Signaling in Endometrial Cancer: A Key Oncogenic Pathway with Several Open Questions. Horm. Cancer 2019, 10, 51–63. [Google Scholar] [CrossRef]
- Finotello, F.; Mayer, C.; Plattner, C.; Laschober, G.; Rieder, D.; Hackl, H.; Krogsdam, A.; Loncova, Z.; Posch, W.; Wilflingseder, D.; et al. Molecular and Pharmacological Modulators of the Tumor Immune Contexture Revealed by Deconvolution of RNA-Seq Data. Genome Med. 2019, 11, 34. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Characteristics, N (%) | Adrenal Restrictive (AA), N (%) | Heterozygous (AC), N (%) | Adrenal Permissive (CC), N (%) | Total | FDR |
|---|---|---|---|---|---|
| Patients | |||||
| Genotype | 3220 (63.32) | 1415 (27.83) | 450 (8.85) | 5085 (100) | |
| Subtype Data | |||||
| ER+/HER2- | 1980 (59.41) | 1040 (31.20) | 313 (9.39) | 3333 (100) | |
| TNBC | 1240 (70.78) | 375 (21.40) | 137 (7.82) | 1752 (100) | |
| Inferred Menopausal Status (assumed based on age at molecular profiling) | |||||
| Premenopausal (≤55 years) | 1040 (63.88) | 444 (27.27) | 144 (8.85) | 1628 (100) | |
| Postmenopausal (>55 years) | 2180 (63.06) | 971 (28.09) | 306 (8.85) | 3457 (100) | |
| Menopausal Subtype Data | |||||
| Premenopausal ER+ | 560 (60.54) | 279 (30.16) | 86 (9.30) | 925 (100) | |
| Premenopausal TNBC | 480 (68.28) | 165 (23.47) | 58 (8.25) | 703 (100) | |
| Postmenopausal ER+ | 1420 (58.97) | 761 (31.60) | 227 (9.43) | 2408 (100) | |
| Postmenopausal TNBC | 760 (72.45) | 210(20.02) | 79 (7.53) | 1049 (100) | |
| Race Data (Self-Identified) | |||||
| White | 1428 (54.82) | 890 (34.17) | 287 (11.02) | 2605 (100) | 3.50 × 10−36 |
| Black Or African American | 753 (81.32) | 123 (13.28) | 50 (5.40) | 926 (100) | 6.80 × 10−35 |
| Asian Or Pacific Islander | 142 (80.23) | 24 (13.56) | 11 (6.21) | 177 (100) | 1.60 × 10−5 |
| Other | 144 (60.50) | 75 (31.51) | 19 (7.98) | 238 (100) | 4.20 × 10−1 |
| Unknown | 284 (64.84) | 124 (28.31) | 30 (6.85) | 438 (100) | 3.04 × 10−1 |
| Tumor Mutational Burden (TMB) and Microsatellite Instability | |||||
| TMB High | 880 (56.63) | 600 (38.61) | 74 (4.76) | 1554 (100) | 1.00 |
| dMMR/MSI-H High | 771 (56.86) | 520 (38.35) | 65 (4.79) | 1356 (100) | 1.00 |
| Patient Characteristics, N (%) | Adrenal Restrictive (AA), N (%) | Heterozygous (AC), N (%) | Adrenal Permissive (CC), N (%) | Total | q-Values (FDR) |
|---|---|---|---|---|---|
| Patients | |||||
| Genotype | 3499 (60.63) | 1935 (33.53) | 337 (5.84) | 5771 (100.00) | |
| Subtype Data | |||||
| Carcinosarcoma | 114 (71.70) | 32 (20.13) | 13 (8.18) | 159 (100) | |
| Clear Cell | 153 (62.20) | 76 (30.89) | 17 (6.91) | 246 (100) | |
| Endometrioid | 1876 (56.80) | 1230 (37.24) | 197 (5.96) | 3303 (100) | |
| Serous | 1270 (65.13) | 571 (29.28) | 109 (5.59) | 1950 (100) | |
| Inferred Menopausal Status (assumed based on age at molecular profiling) | |||||
| Premenopausal (≤55 years) | 485 (62.91) | 242 (31.39) | 44 (5.71) | 771 (100) | |
| Postmenopausal (>55 years) | 3014 (60.28) | 1693 (33.86) | 293 (5.86) | 5000 (100) | |
| Menopausal Subtype Data | |||||
| Premenopausal Endometrioid | 399 (60.92) | 216 (32.98) | 40 (6.11) | 655 (100) | |
| Postmenopausal Endometrioid | 1477 (55.78) | 1014 (38.29) | 157 (5.93) | 2648 (100) | |
| Premenopausal Other Subtypes | 86 (74.14) | 26 (22.41) | 4 (3.45) | 116 (100) | |
| Postmenopausal Other Subtypes | 1537 (65.35) | 679 (28.87) | 136 (5.78) | 2352 (100) | |
| ER-Positive Prevalence by Subtype | |||||
| Premenopausal Endometrioid | 345 (60.85) | 185 (32.63) | 37 (6.52) | 567 (100) | 1.00 |
| Postmenopausal Endometrioid | 1270 (55.12) | 892 (38.72) | 142 (6.16) | 2304 (100) | 1.00 |
| Premenopausal Other Subtypes | 42 (72.41) | 14 (24.14) | 2 (3.45) | 58 (100) | 9.5 × 10−1 |
| Postmenopausal Other Subtypes | 849 (65.36) | 387 (29.79) | 63 (4.85) | 1299 (100) | 9.0 x 10−1 |
| Race Data (Self-Identified) | |||||
| White | 1597 (52.67) | 1205 (39.74) | 230 (7.59) | 3032 (100) | 1.80 × 10−1 |
| Black or African American | 882 (79.89) | 199 (18.03) | 22 (2.08) | 1104 (100) | 1.13 × 10−37 |
| Asian or Pacific Islander | 146 (78.49) | 38 (20.43) | 2 (1.08) | 186 (100) | 3.17 × 10−18 |
| Other | 127 (53.14) | 93 (38.91) | 19 (7.95) | 239 (100) | 2.88 × 10−6 |
| Unknown | 238 (56.53) | 159 (37.77) | 24 (5.70) | 421 (100) | 5.92 × 10−2 |
| Tumor Mutational Burden (TMB) and Microsatellite Instability | |||||
| TMB High | 880 (56.63) | 600 (38.61) | 74 (4.76) | 1554 (100) | 1.00 |
| dMMR/MSI-H High | 771 (56.86) | 520 (38.35) | 65 (4.79) | 1356 (100) | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vobugari, N.; Makovec, A.; Kellen, S.; Nazari, S.S.; Elliott, A.; Schmeck, D.; Deacon, A.; von Dohlen, G.; John, E.; Barata, P.C.; et al. HSD3B1 (c.1100C) Genotype Is Associated with Distinct Tumoral and Clinical Outcomes in Breast and Endometrial Cancers. Int. J. Mol. Sci. 2025, 26, 5720. https://doi.org/10.3390/ijms26125720
Vobugari N, Makovec A, Kellen S, Nazari SS, Elliott A, Schmeck D, Deacon A, von Dohlen G, John E, Barata PC, et al. HSD3B1 (c.1100C) Genotype Is Associated with Distinct Tumoral and Clinical Outcomes in Breast and Endometrial Cancers. International Journal of Molecular Sciences. 2025; 26(12):5720. https://doi.org/10.3390/ijms26125720
Chicago/Turabian StyleVobugari, Nikitha, Allison Makovec, Samuel Kellen, Shayan S. Nazari, Andrew Elliott, Devin Schmeck, Aiden Deacon, Gabriella von Dohlen, Emily John, Pedro C. Barata, and et al. 2025. "HSD3B1 (c.1100C) Genotype Is Associated with Distinct Tumoral and Clinical Outcomes in Breast and Endometrial Cancers" International Journal of Molecular Sciences 26, no. 12: 5720. https://doi.org/10.3390/ijms26125720
APA StyleVobugari, N., Makovec, A., Kellen, S., Nazari, S. S., Elliott, A., Schmeck, D., Deacon, A., von Dohlen, G., John, E., Barata, P. C., Agarwal, N., Geller, M. A., Erickson, B. K., Sledge, G., Ostrander, J. H., McKay, R. R., Ryan, C. J., Sharifi, N., Antonarakis, E. S., & Hwang, J. (2025). HSD3B1 (c.1100C) Genotype Is Associated with Distinct Tumoral and Clinical Outcomes in Breast and Endometrial Cancers. International Journal of Molecular Sciences, 26(12), 5720. https://doi.org/10.3390/ijms26125720