
DMPC-Based Liposomal Vesicles for Encapsulation and Controlled Release of NMN and Matrigel in Sarcopenia Therapy

 ,
,  ,
,  ,
,  ,
,  , ,
, , 
Abstract
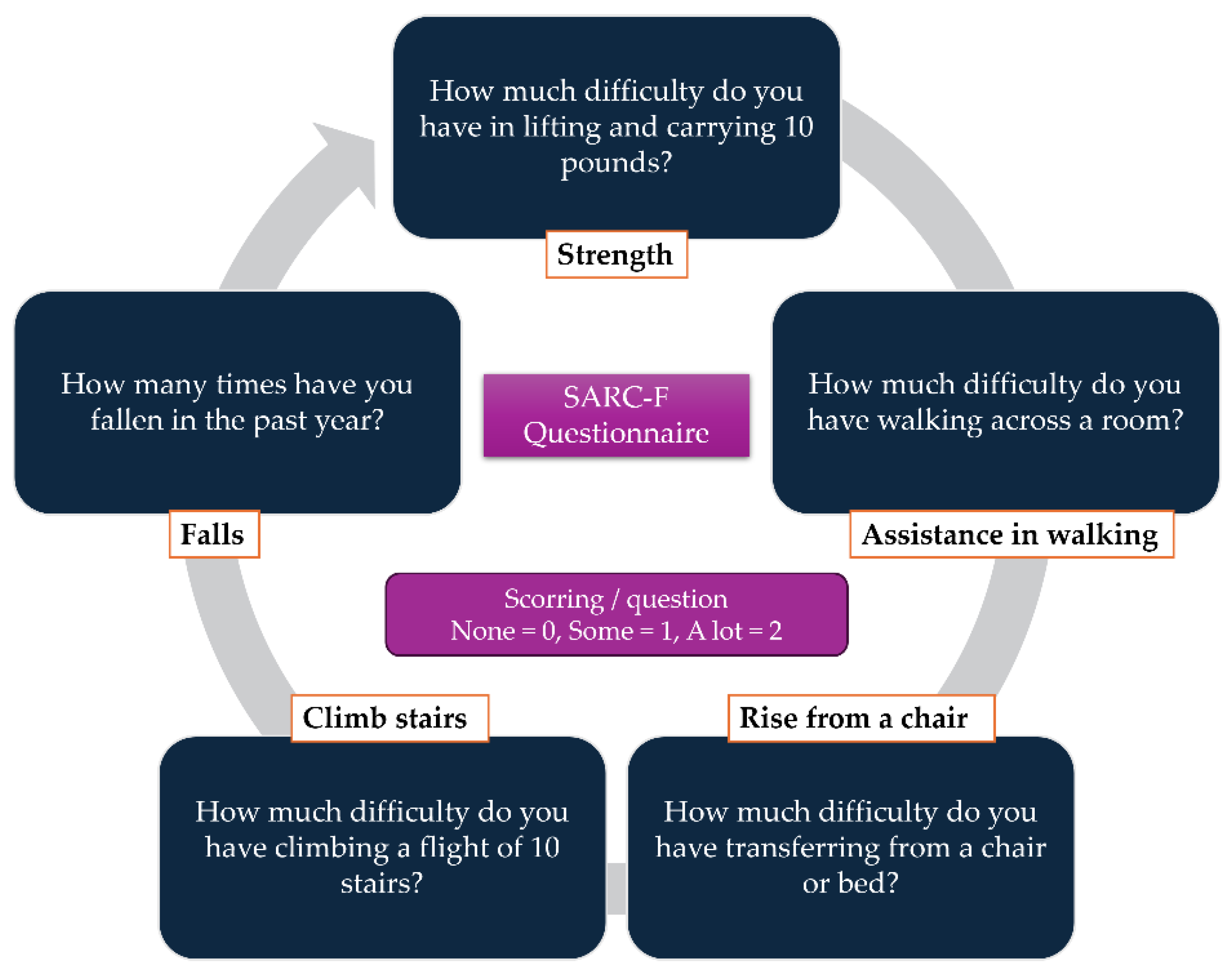
1. Introduction
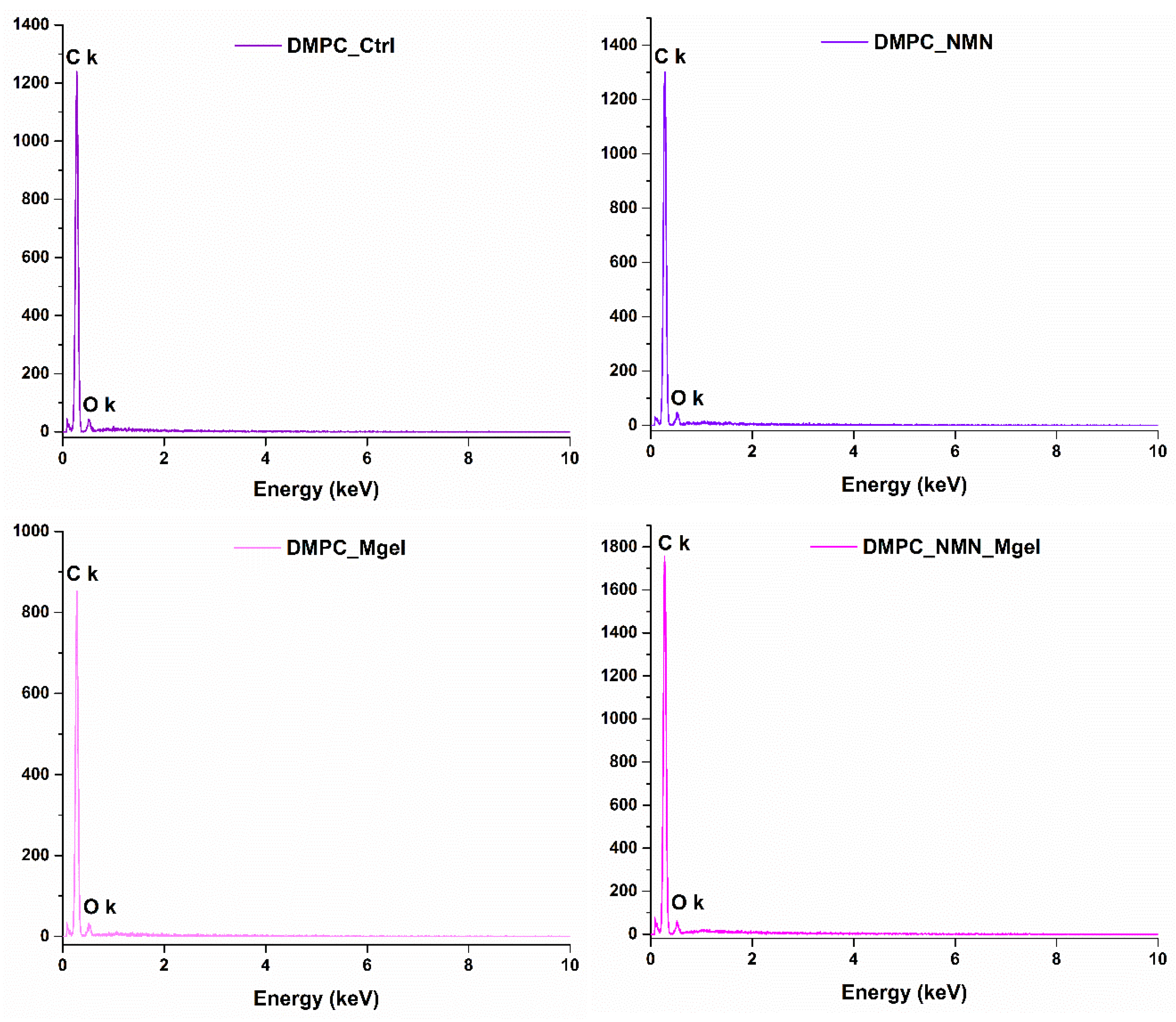
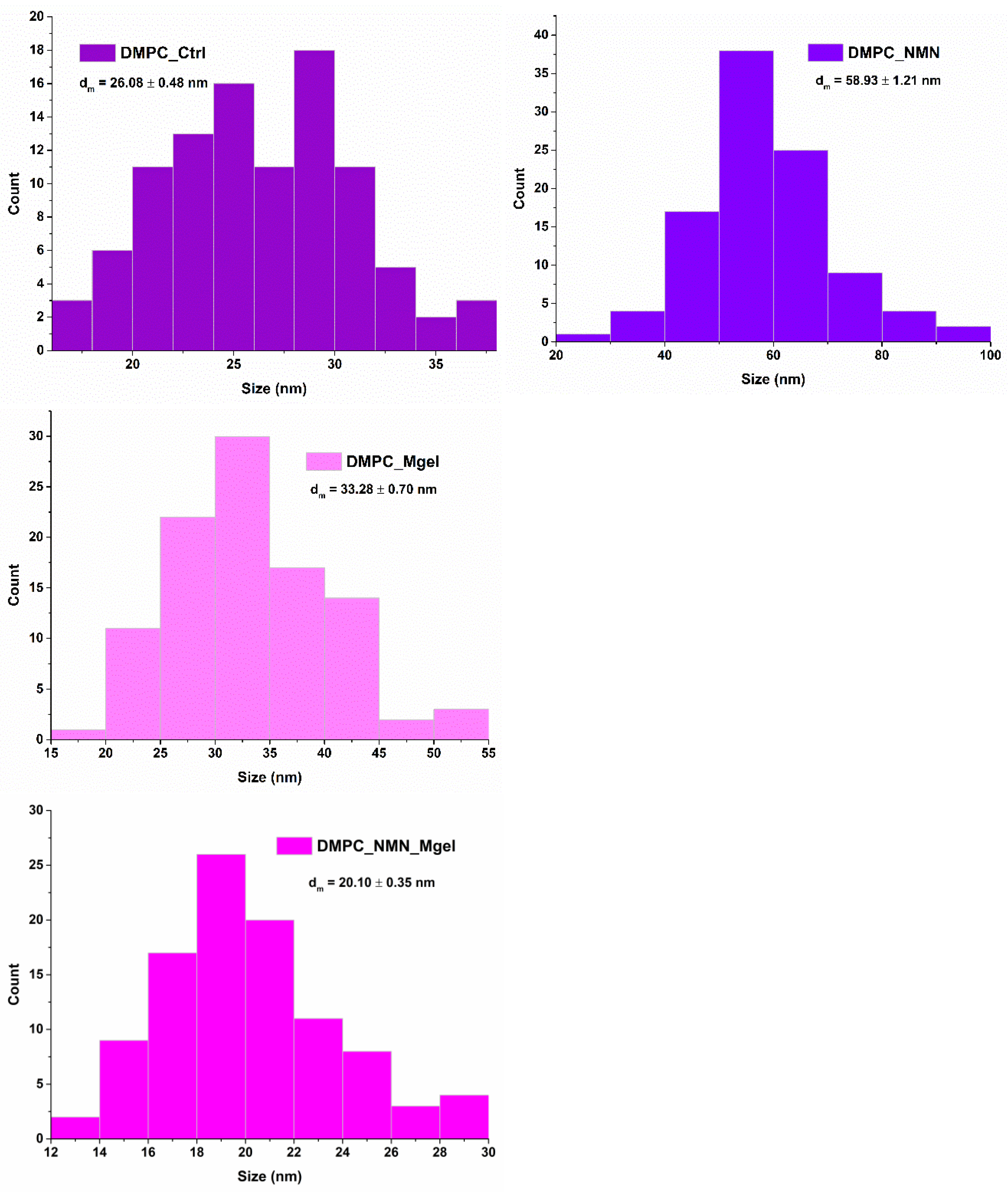
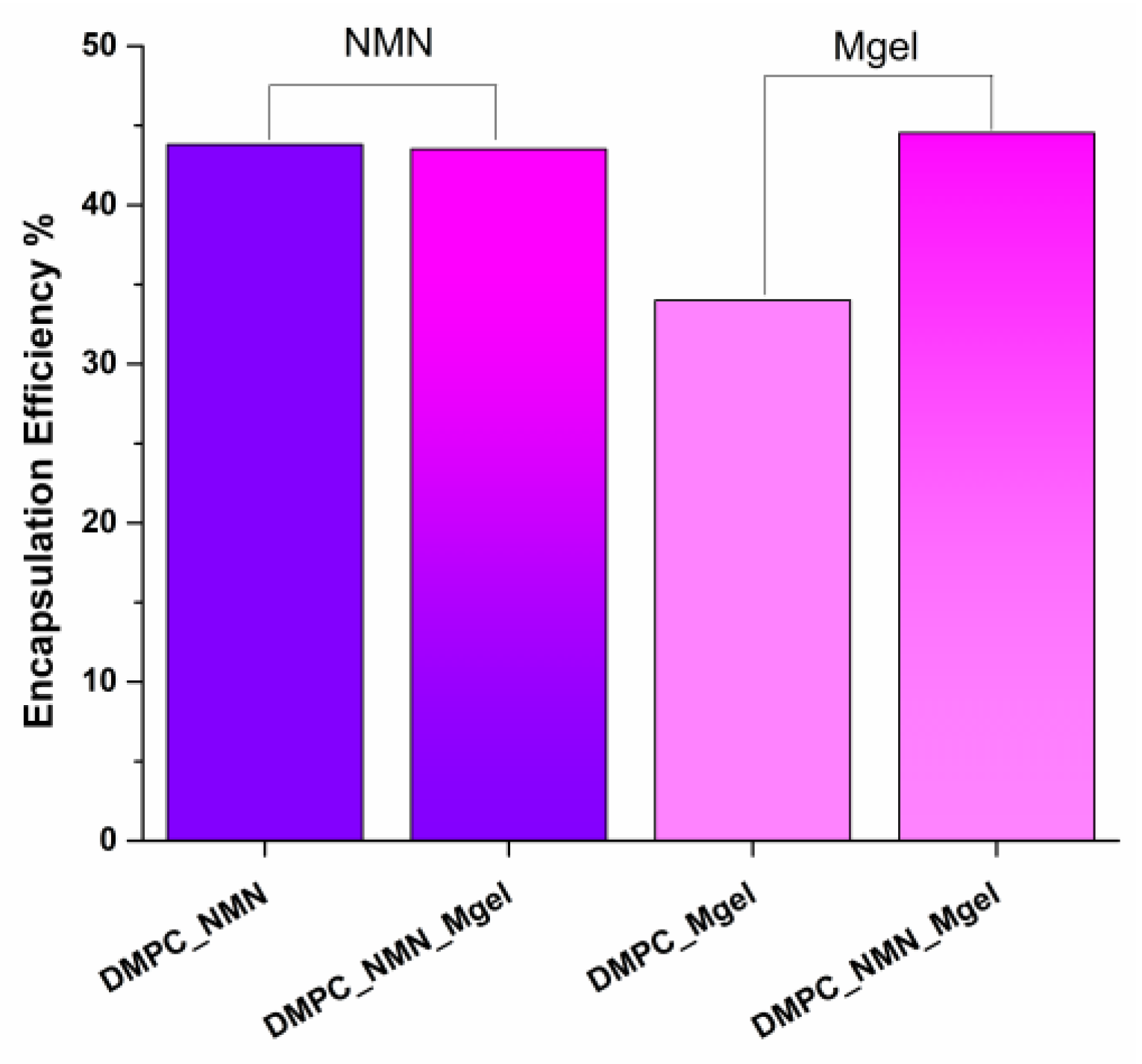
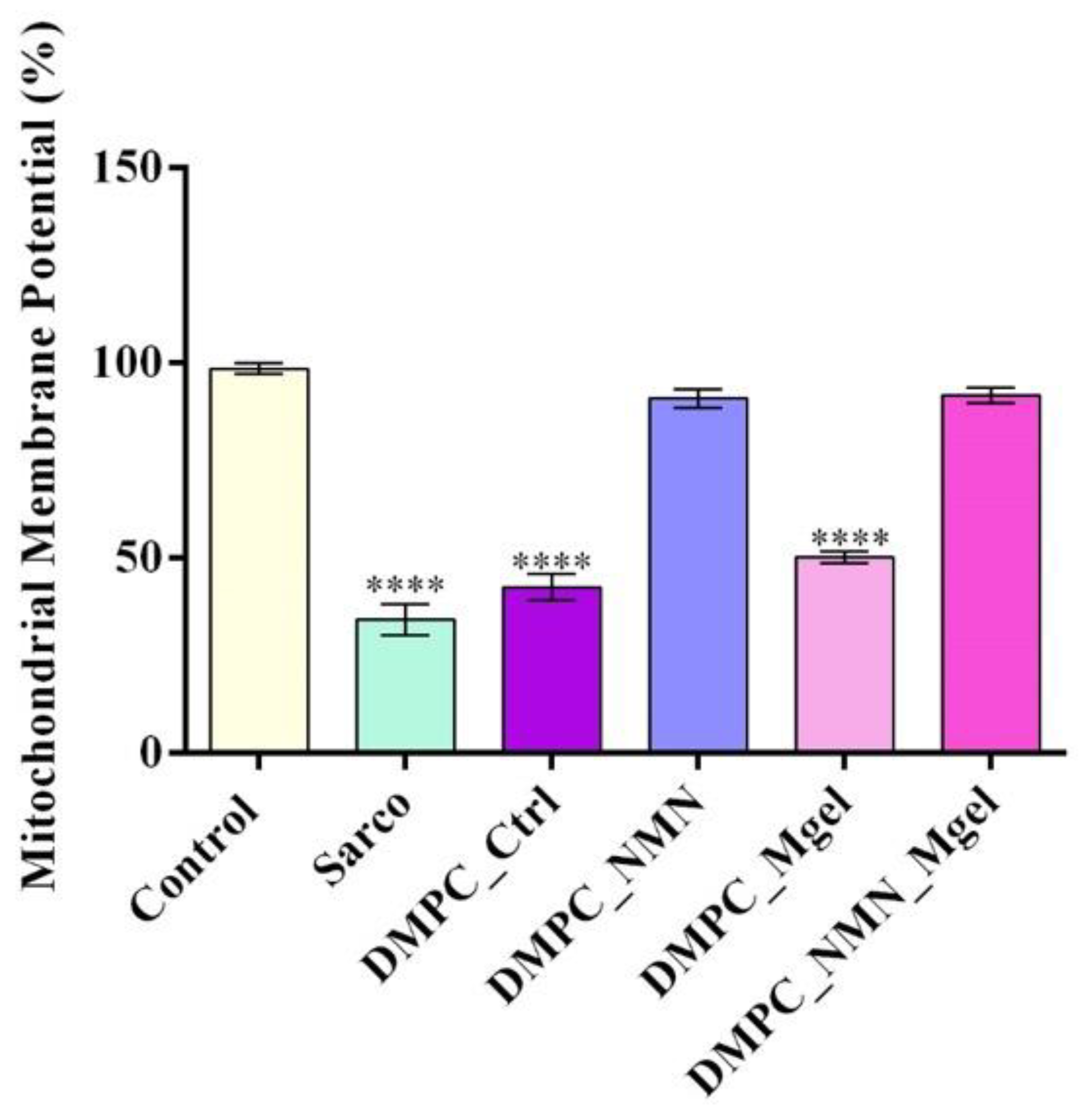
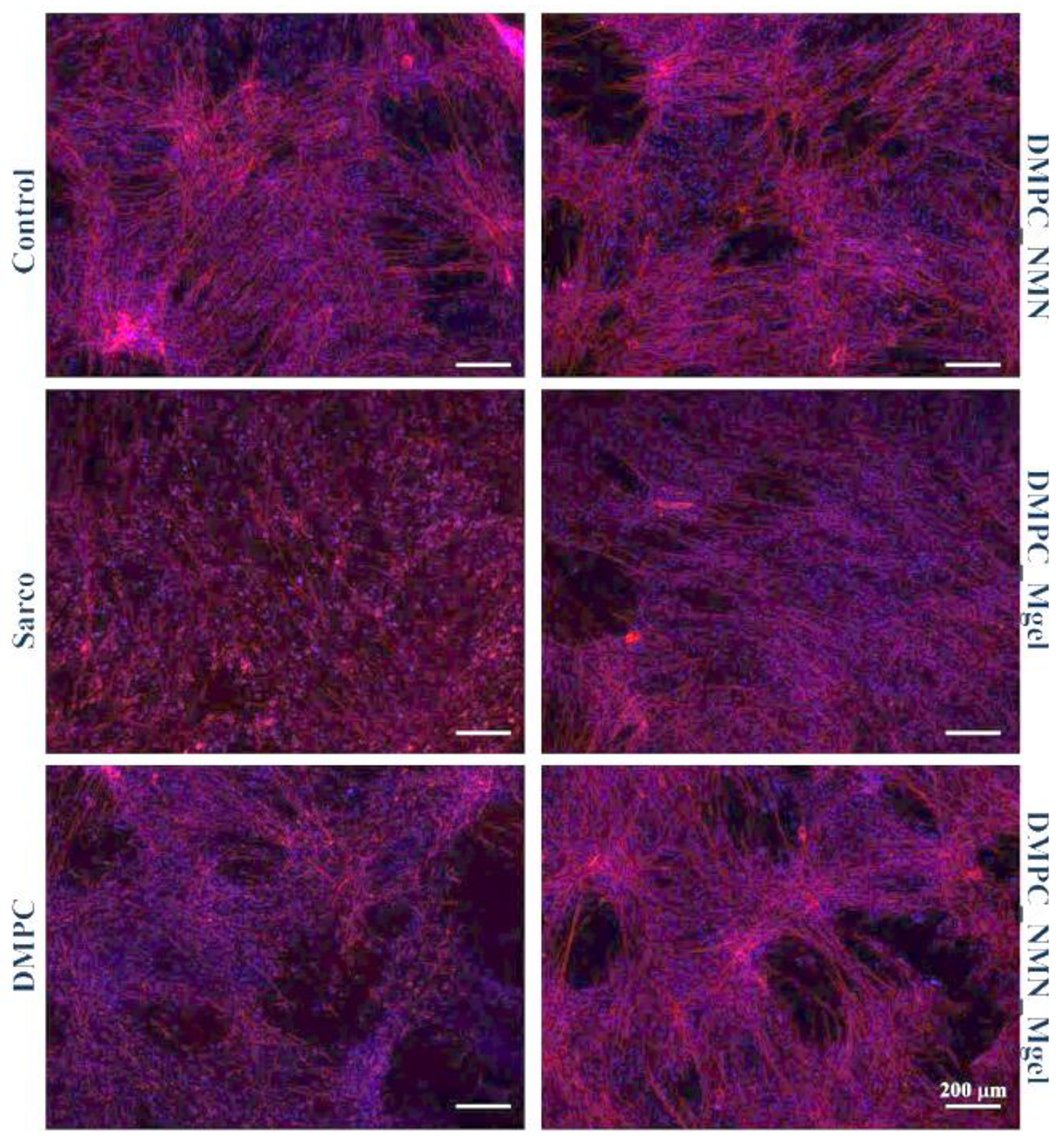
2. Results
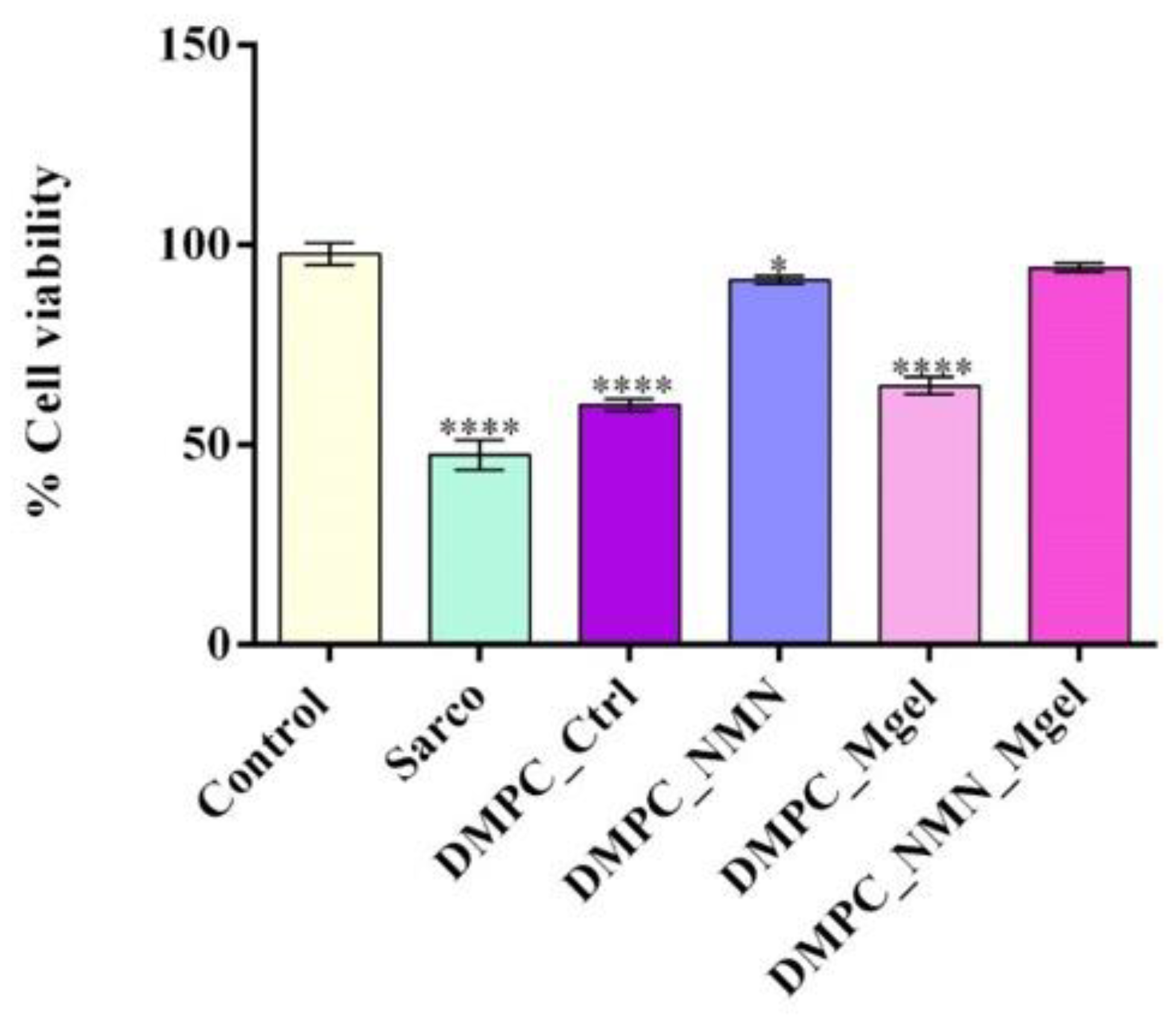
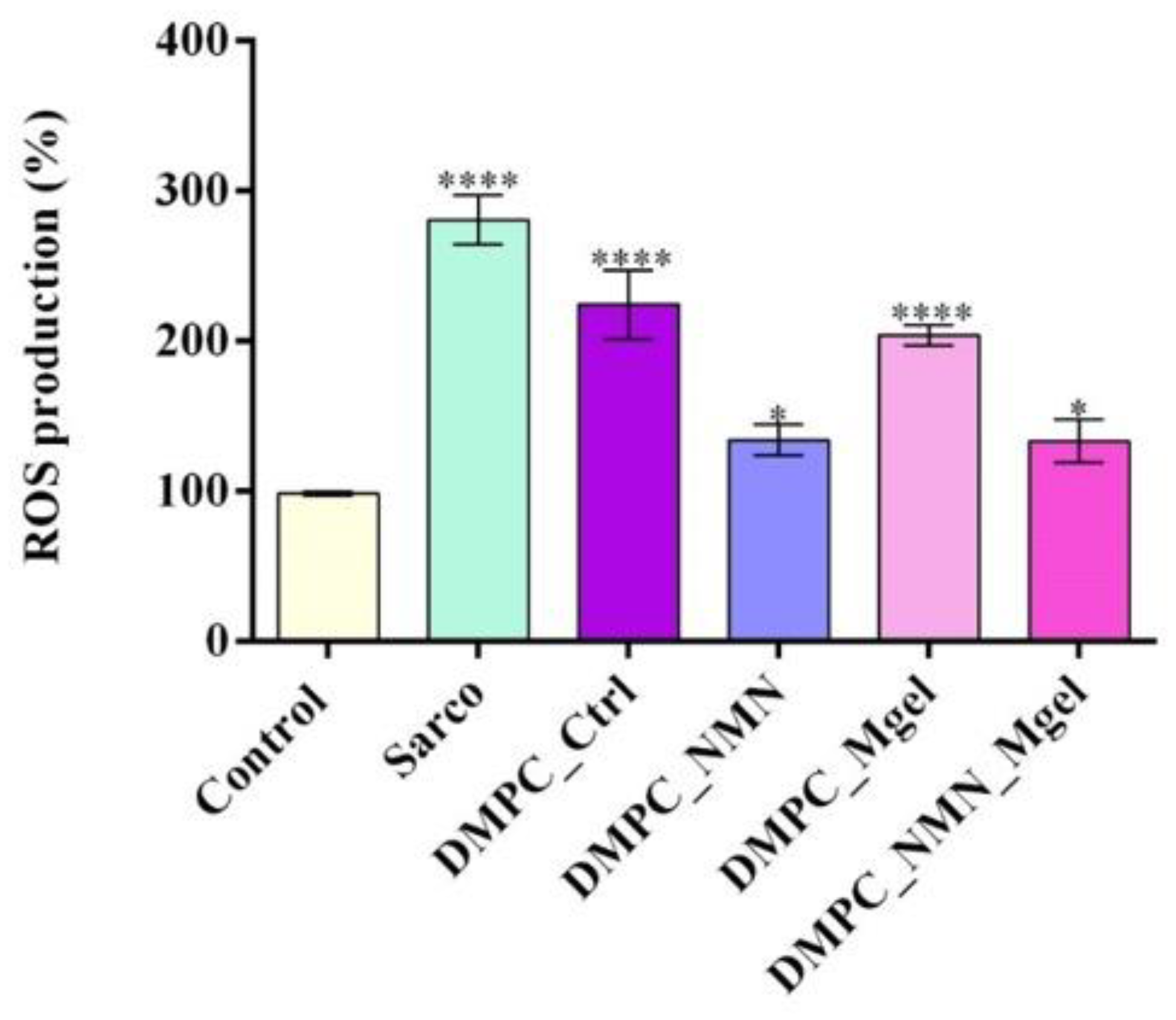
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
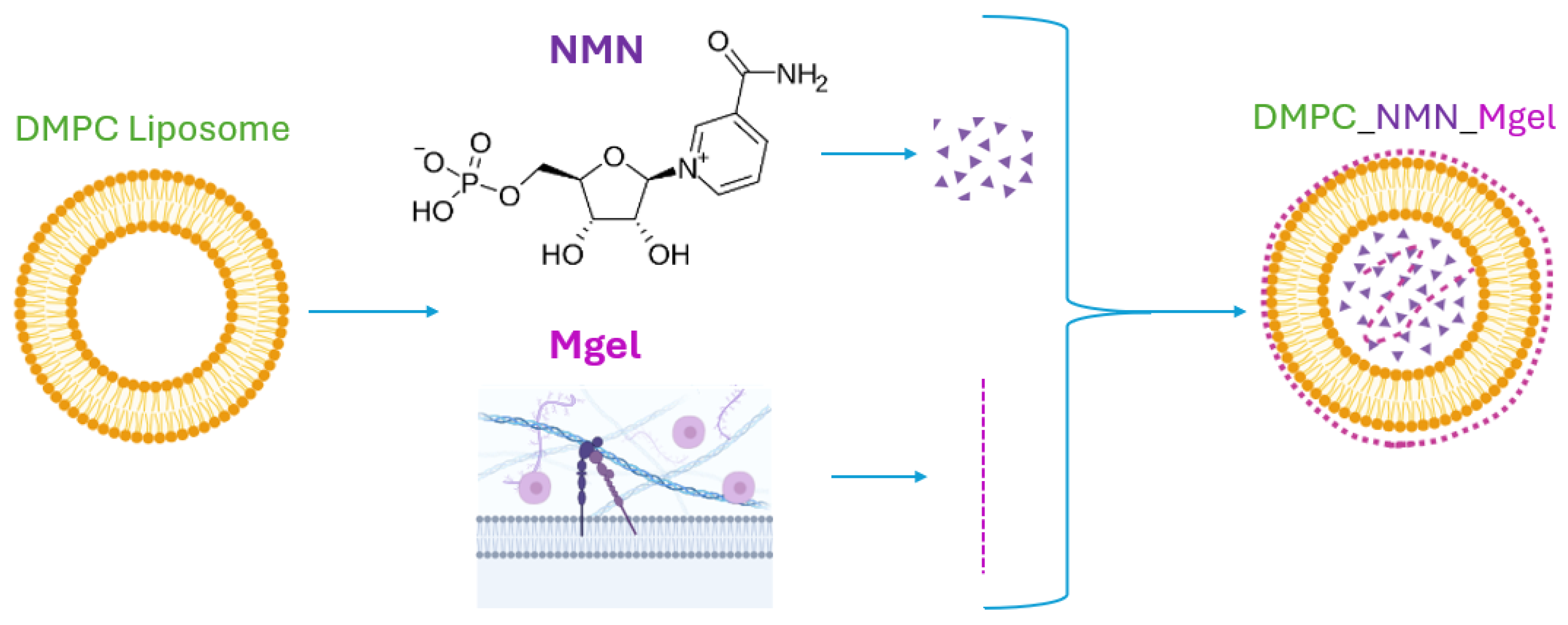
4.2.1. Synthesis of DMPC-Based Lipid Vesicles Encapsulate NMN and Matrigel
4.2.2. Control Sample (DMPC_Ctrl)
4.2.3. NMN-Loaded Vesicles (DMPC_NMN)
4.2.4. Matrigel-Loaded Vesicles (DMPC_Mgel)
4.2.5. Dual-Encapsulated Vesicles (DMPC_NMN_Mgel)
4.3. Analysis Methods
4.3.1. Dynamic Light Scattering (DLS)
4.3.2. Scanning Electron Microscopy (SEM)
4.3.3. Transmission Electron Microscopy (TEM)
4.3.4. Encapsulation Efficiency (EE%)
4.3.5. Drug Release %
4.4. In Vitro Biological Investigations
4.4.1. Cell Culture Model
- A.
- Control group—differentiation medium was refreshed without additional treatment;
- B.
- Sarcopenia model group—myotubes exposed to 100 μM H2O2 to induce a sarcopenia-like phenotype;
- C.
- DMPC group—myotubes treated with 100 μM H2O2 + DMPC vesicles;
- D.
- DMPC NMN group—myotubes treated with 100 μM H2O2 + DMPC vesicles loaded with NMN;
- E.
- DMPC Matrigel group—myotubes treated with 100 μM H2O2 + DMPC vesicles loaded with Matrigel;
- F.
- DMPC NMN + Matrigel group—myotubes treated with 100 μM H2O2 + DMPC vesicles loaded with NMN and Matrigel.
4.4.2. Cell Viability Evaluation
4.4.3. Detection of Intracellular Reactive Oxygen Species (ROS)
4.4.4. Nitric Oxide (NO) Quantification
4.4.5. Mitochondrial Membrane Potential (MMP) Evaluation
4.4.6. Analysis of Cytoskeletal Integrity
4.4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ispoglou, T.; Wilson, O.; McCullough, D.; Aldrich, L.; Ferentinos, P.; Lyall, G.; Stavropoulos-Kalinoglou, A.; Duckworth, L.; Brown, M.A.; Sutton, L.; et al. A Narrative Review of Non-Pharmacological Strategies for Managing Sarcopenia in Older Adults with Cardiovascular and Metabolic Diseases. Biology 2023, 12, 892. [Google Scholar] [CrossRef]
- Petermann-Rocha, F.; Balntzi, V.; Gray, S.R.; Lara, J.; Ho, F.K.; Pell, J.P.; Celis-Morales, C. Global prevalence of sarcopenia and severe sarcopenia: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle 2022, 13, 86–99. [Google Scholar] [CrossRef]
- Bauer, J.; Morley, J.E.; Schols, A.; Ferrucci, L.; Cruz-Jentoft, A.J.; Dent, E.; Baracos, V.E.; Crawford, J.A.; Doehner, W.; Heymsfield, S.B.; et al. Sarcopenia: A Time for Action. An SCWD Position Paper. J. Cachexia Sarcopenia Muscle 2019, 10, 956–961. [Google Scholar] [CrossRef]
- Tournadre, A.; Vial, G.; Capel, F.; Soubrier, M.; Boirie, Y. Sarcopenia. Jt. Bone Spine 2019, 86, 309–314. [Google Scholar] [CrossRef]
- Albano, D.; Messina, C.; Vitale, J.; Sconfienza, L.M. Imaging of sarcopenia: Old evidence and new insights. Eur. Radiol. 2020, 30, 2199–2208. [Google Scholar] [CrossRef]
- Xie, W.Q.; He, M.; Yu, D.J.; Wu, Y.X.; Wang, X.H.; Lv, S.; Xiao, W.F.; Li, Y.S. Mouse models of sarcopenia: Classification and evaluation. J. Cachexia Sarcopenia Muscle 2021, 12, 538–554. [Google Scholar] [CrossRef]
- Al Saedi, A.; Debruin, D.A.; Hayes, A.; Hamrick, M. Lipid metabolism in sarcopenia. Bone 2022, 164, 116539. [Google Scholar] [CrossRef]
- Keller, K. Sarcopenia. Wien. Med. Wochenschr. 2019, 169, 157–172. [Google Scholar] [CrossRef]
- Cho, M.-R.; Lee, S.; Song, S.-K. A Review of Sarcopenia Pathophysiology, Diagnosis, Treatment and Future Direction. J. Korean Med. Sci. 2022, 37, e146. [Google Scholar] [CrossRef]
- Zhang, X.; Li, H.; He, M.; Wang, J.; Wu, Y.; Li, Y. Immune system and sarcopenia: Presented relationship and future perspective. Exp. Gerontol. 2022, 164, 111823. [Google Scholar] [CrossRef]
- Chen, B.; Shan, T. The role of satellite and other functional cell types in muscle repair and regeneration. J. Muscle Res. Cell Motil. 2019, 40, 1–8. [Google Scholar] [CrossRef]
- Hunter, G.R.; Singh, H.; Carter, S.J.; Bryan, D.R.; Fisher, G. Sarcopenia and Its Implications for Metabolic Health. J. Obes. 2019, 2019, 8031705. [Google Scholar] [CrossRef]
- Kim, J.W.; Kim, R.; Choi, H.; Lee, S.-J.; Bae, G.-U. Understanding of sarcopenia: From definition to therapeutic strategies. Arch. Pharmacal Res. 2021, 44, 876–889. [Google Scholar] [CrossRef]
- Bilski, J.; Pierzchalski, P.; Szczepanik, M.; Bonior, J.; Zoladz, J.A. Multifactorial Mechanism of Sarcopenia and Sarcopenic Obesity. Role of Physical Exercise, Microbiota and Myokines. Cells 2022, 11, 160. [Google Scholar] [CrossRef]
- Pascual-Fernández, J.; Fernández-Montero, A.; Córdova-Martínez, A.; Pastor, D.; Martínez-Rodríguez, A.; Roche, E. Sarcopenia: Molecular Pathways and Potential Targets for Intervention. Int. J. Mol. Sci. 2020, 21, 8844. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, Y.; Wang, J. Protein Nutritional Support: The Classical and Potential New Mechanisms in the Prevention and Therapy of Sarcopenia. J. Agric. Food Chem. 2020, 68, 4098–4108. [Google Scholar] [CrossRef]
- Aslam, M.A.; Ma, E.B.; Huh, J.Y. Pathophysiology of sarcopenia: Genetic factors and their interplay with environmental factors. Metab.-Clin. Exp. 2023, 149, 155711. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhou, X.; Zhu, A.; Xiong, S.; Xie, J.; Bai, Z. Advances in exercise to alleviate sarcopenia in older adults by improving mitochondrial dysfunction. Front. Physiol. 2023, 14, 1196426. [Google Scholar] [CrossRef]
- Xu, X.; Wen, Z. The mediating role of inflammaging between mitochondrial dysfunction and sarcopenia in aging: A review. Am. J. Clin. Exp. Immunol. 2023, 12, 109–126. [Google Scholar]
- Romanello, V. The Interplay between Mitochondrial Morphology and Myomitokines in Aging Sarcopenia. Int. J. Mol. Sci. 2021, 22, 91. [Google Scholar] [CrossRef]
- Jackson, M.J. Reactive oxygen species in sarcopenia: Should we focus on excess oxidative damage or defective redox signalling? Mol. Asp. Med. 2016, 50, 33–40. [Google Scholar] [CrossRef]
- Xu, H.; Brown, J.L.; Bhaskaran, S.; Van Remmen, H. Reactive oxygen species in the pathogenesis of sarcopenia. Free Radic. Biol. Med. 2025, 227, 446–458. [Google Scholar] [CrossRef]
- Yoo, S.-Z.; No, M.-H.; Heo, J.-W.; Park, D.-H.; Kang, J.-H.; Kim, S.H.; Kwak, H.-B. Role of exercise in age-related sarcopenia. J. Exerc. Rehabil. 2018, 14, 551–558. [Google Scholar] [CrossRef]
- Vergara-Fernandez, O.; Trejo-Avila, M.; Salgado-Nesme, N. Sarcopenia in patients with colorectal cancer: A comprehensive review. World J. Clin. Cases 2020, 8, 1188. [Google Scholar] [CrossRef]
- Su, M.; Qiu, F.; Li, Y.; Che, T.; Li, N.; Zhang, S. Mechanisms of the NAD+ salvage pathway in enhancing skeletal muscle function. Front. Cell Dev. Biol. 2024, 12, 1464815. [Google Scholar] [CrossRef]
- Ioniță-Radu, F.; Nicolau, I.-N.; Petrache, O.-G.; Groșeanu, M.-L.; Bojincă, V.-C.; Negru, M.-M.; Bucurică, S.; Anghel, D. Correlation between Trabecular Bone Score and Homocysteine Level in Rheumatoid Arthritis Patients on Anti-TNF Inhibitors. Life 2024, 14, 463. [Google Scholar] [CrossRef]
- Wagner, S.; Manickam, R.; Brotto, M.; Tipparaju, S.M. NAD+ centric mechanisms and molecular determinants of skeletal muscle disease and aging. Mol. Cell. Biochem. 2022, 477, 1829–1848. [Google Scholar] [CrossRef]
- Membrez, M.; Migliavacca, E.; Christen, S.; Yaku, K.; Trieu, J.; Lee, A.K.; Morandini, F.; Giner, M.P.; Stiner, J.; Makarov, M.V.; et al. Trigonelline is an NAD+ precursor that improves muscle function during ageing and is reduced in human sarcopenia. Nat. Metab. 2024, 6, 433–447. [Google Scholar] [CrossRef]
- Covarrubias, A.J.; Perrone, R.; Grozio, A.; Verdin, E. NAD+ metabolism and its roles in cellular processes during ageing. Nat. Rev. Mol. Cell Biol. 2021, 22, 119–141. [Google Scholar] [CrossRef]
- Janssens, G.E.; Grevendonk, L.; Perez, R.Z.; Schomakers, B.V.; de Vogel-van den Bosch, J.; Geurts, J.M.W.; van Weeghel, M.; Schrauwen, P.; Houtkooper, R.H.; Hoeks, J. Healthy aging and muscle function are positively associated with NAD+ abundance in humans. Nat. Aging 2022, 2, 254–263. [Google Scholar] [CrossRef]
- Ackermans, L.L.G.C.; Rabou, J.; Basrai, M.; Schweinlin, A.; Bischoff, S.C.; Cussenot, O.; Cancel-Tassin, G.; Renken, R.J.; Gómez, E.; Sánchez-González, P.; et al. Screening, diagnosis and monitoring of sarcopenia: When to use which tool? Clin. Nutr. ESPEN 2022, 48, 36–44. [Google Scholar] [CrossRef]
- Messina, C.; Maffi, G.; Vitale, J.A.; Ulivieri, F.M.; Guglielmi, G.; Sconfienza, L.M. Diagnostic imaging of osteoporosis and sarcopenia: A narrative review. Quant. Imaging Med. Surg. 2018, 8, 86–99. [Google Scholar] [CrossRef]
- Shen, Y.; Shi, Q.; Nong, K.; Li, S.; Yue, J.; Huang, J.; Dong, B.; Beauchamp, M.; Hao, Q. Exercise for sarcopenia in older people: A systematic review and network meta-analysis. J. Cachexia Sarcopenia Muscle 2023, 14, 1199–1211. [Google Scholar] [CrossRef]
- Rooks, D.; Roubenoff, R. Development of Pharmacotherapies for the Treatment of Sarcopenia. J. Frailty Aging 2019, 8, 120–130. [Google Scholar] [CrossRef]
- Coen, P.M.; Musci, R.V.; Hinkley, J.M.; Miller, B.F. Mitochondria as a Target for Mitigating Sarcopenia. Front. Physiol. 2019, 9, 1883. [Google Scholar] [CrossRef]
- Delama, A.; Teixeira, M.I.; Dorati, R.; Genta, I.; Conti, B.; Lamprou, D.A. Microfluidic encapsulation method to produce stable liposomes containing iohexol. J. Drug Deliv. Sci. Technol. 2019, 54, 101340. [Google Scholar] [CrossRef]
- Luiz, H.; Oliveira Pinho, J.; Gaspar, M.M. Advancing Medicine with Lipid-Based Nanosystems—The Successful Case of Liposomes. Biomedicines 2023, 11, 435. [Google Scholar] [CrossRef]
- Guimarães, D.; Cavaco-Paulo, A.; Nogueira, E. Design of liposomes as drug delivery system for therapeutic applications. Int. J. Pharm. 2021, 601, 120571. [Google Scholar] [CrossRef]
- Almeida, B.; Nag, O.K.; Rogers, K.E.; Delehanty, J.B. Recent Progress in Bioconjugation Strategies for Liposome-Mediated Drug Delivery. Molecules 2020, 25, 5672. [Google Scholar] [CrossRef]
- González-Cela-Casamayor, M.A.; López-Cano, J.J.; Bravo-Osuna, I.; Andrés-Guerrero, V.; Vicario-de-la-Torre, M.; Guzmán-Navarro, M.; Benítez-del-Castillo, J.M.; Herrero-Vanrell, R.; Molina-Martínez, I.T. Novel Osmoprotective DOPC-DMPC Liposomes Loaded with Antihypertensive Drugs as Potential Strategy for Glaucoma Treatment. Pharmaceutics 2022, 14, 1405. [Google Scholar] [CrossRef]
- Sivagnanam, S.; Das, K.; Pan, I.; Stewart, A.; Barik, A.; Maity, B.; Das, P. Engineered triphenylphosphonium-based, mitochondrial-targeted liposomal drug delivery system facilitates cancer cell killing actions of chemotherapeutics. RSC Chem. Biol. 2024, 5, 236–248. [Google Scholar] [CrossRef]
- Ferreira, M.; Pinto, S.N.; Aires-da-Silva, F.; Bettencourt, A.; Aguiar, S.I.; Gaspar, M.M. Liposomes as a Nanoplatform to Improve the Delivery of Antibiotics into Staphylococcus aureus Biofilms. Pharmaceutics 2021, 13, 321. [Google Scholar] [CrossRef]
- Rahman, M.M.; Wang, J.; Wang, G.; Su, Z.; Li, Y.; Chen, Y.; Meng, J.; Yao, Y.; Wang, L.; Wilkens, S.; et al. Chimeric nanobody-decorated liposomes by self-assembly. Nat. Nanotechnol. 2024, 19, 818–824. [Google Scholar] [CrossRef]
- Igarashi, M.; Nakagawa-Nagahama, Y.; Miura, M.; Kashiwabara, K.; Yaku, K.; Sawada, M.; Sekine, R.; Fukamizu, Y.; Sato, T.; Sakurai, T.; et al. Chronic nicotinamide mononucleotide supplementation elevates blood nicotinamide adenine dinucleotide levels and alters muscle function in healthy older men. NPJ Aging 2022, 8, 5. [Google Scholar] [CrossRef]
- Liang, R.; Xiang, Q.; Dai, M.; Lin, T.; Xie, D.; Song, Q.; Liu, Y.; Yue, J. Identification of nicotinamide N-methyltransferase as a promising therapeutic target for sarcopenia. Aging Cell 2024, 23, e14236. [Google Scholar] [CrossRef]
- Wen, J.; Syed, B.; Kim, S.; Shehabat, M.; Ansari, U.; Razick, D.I.; Akhtar, M.; Pai, D. Improved Physical Performance Parameters in Patients Taking Nicotinamide Mononucleotide (NMN): A Systematic Review of Randomized Control Trials. Cureus 2024, 16, e65961. [Google Scholar] [CrossRef]
- Alave Reyes-Furrer, A.; De Andrade, S.; Bachmann, D.; Jeker, H.; Steinmann, M.; Accart, N.; Dunbar, A.; Rausch, M.; Bono, E.; Rimann, M.; et al. Matrigel 3D bioprinting of contractile human skeletal muscle models recapitulating exercise and pharmacological responses. Commun. Biol. 2021, 4, 1183. [Google Scholar] [CrossRef]
- De Stefano, P.; Briatico-Vangosa, F.; Bianchi, E.; Pellegata, A.F.; Hartung de Hartungen, A.; Corti, P.; Dubini, G. Bioprinting of Matrigel Scaffolds for Cancer Research. Polymers 2021, 13, 2026. [Google Scholar] [CrossRef]
- Wang, J.; Chu, R.; Ni, N.; Nan, G. The effect of Matrigel as scaffold material for neural stem cell transplantation for treating spinal cord injury. Sci. Rep. 2020, 10, 2576. [Google Scholar] [CrossRef]
- Fu, T.; Liang, P.; Song, J.; Wang, J.; Zhou, P.; Tang, Y.; Li, J.; Huang, E. Matrigel Scaffolding Enhances BMP9-induced Bone Formation in Dental Follicle Stem/Precursor Cells. Int. J. Med. Sci. 2019, 16, 567–575. [Google Scholar] [CrossRef]
- Kelly, C.; Jefferies, C.; Cryan, S.A. Targeted liposomal drug delivery to monocytes and macrophages. J. Drug Deliv. 2011, 2011, 727241. [Google Scholar] [CrossRef]
- Takano, S.; Aramaki, Y.; Tsuchiya, S. Physicochemical properties of liposomes affecting apoptosis induced by cationic liposomes in macrophages. Pharm. Res. 2003, 20, 962–968. [Google Scholar] [CrossRef]
- Ren, H.; He, Y.; Liang, J.; Cheng, Z.; Zhang, M.; Zhu, Y.; Hong, C.; Qin, J.; Xu, X.; Wang, J. Role of Liposome Size, Surface Charge, and PEGylation on Rheumatoid Arthritis Targeting Therapy. ACS Appl. Mater. Interfaces 2019, 11, 20304–20315. [Google Scholar] [CrossRef]
- Epstein-Barash, H.; Gutman, D.; Markovsky, E.; Mishan-Eisenberg, G.; Koroukhov, N.; Szebeni, J.; Golomb, G. Physicochemical parameters affecting liposomal bisphosphonates bioactivity for restenosis therapy: Internalization, cell inhibition, activation of cytokines and complement, and mechanism of cell death. J. Control. Release 2010, 146, 182–195. [Google Scholar] [CrossRef]
- Demonbreun, A.R.; Quattrocelli, M.; Barefield, D.Y.; Allen, M.V.; Swanson, K.E.; McNally, E.M. An actin-dependent annexin complex mediates plasma membrane repair in muscle. J. Cell Biol. 2016, 213, 705–718. [Google Scholar] [CrossRef]
- Lombardo, D.; Kiselev, M.A. Methods of Liposomes Preparation: Formation and Control Factors of Versatile Nanocarriers for Biomedical and Nanomedicine Application. Pharmaceutics 2022, 14, 543. [Google Scholar] [CrossRef]
- Putri, D.; Dwiastuti, R.; Hadimartono, M.; Nugroho, A. Optimization of Mixing Temperature and Sonication Duration in Liposome Preparation. J. Pharm. Sci. Community 2017, 14, 79–85. [Google Scholar] [CrossRef]
- de Freitas, C.F.; Calori, I.R.; Tessaro, A.L.; Caetano, W.; Hioka, N. Rapid formation of Small Unilamellar Vesicles (SUV) through low-frequency sonication: An innovative approach. Colloids Surf. B Biointerfaces 2019, 181, 837–844. [Google Scholar] [CrossRef]
- Nowroozi, F.; Almasi, A.; Javidi, J.; Haeri, A.; Dadashzadeh, S. Effect of Surfactant Type, Cholesterol Content and Various Downsizing Methods on the Particle Size of Niosomes. Iran. J. Pharm. Res. 2018, 17, 1–11. [Google Scholar]
- Zylberberg, C.; Matosevic, S. Pharmaceutical liposomal drug delivery: A review of new delivery systems and a look at the regulatory landscape. Drug Deliv. 2016, 23, 3319–3329. [Google Scholar] [CrossRef]
- Kapoor, M.; Lee, S.L.; Tyner, K.M. Liposomal Drug Product Development and Quality: Current US Experience and Perspective. AAPS J. 2017, 19, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Solomon, D.; Gupta, N.; Mulla, N.S.; Shukla, S.; Guerrero, Y.A.; Gupta, V. Role of In Vitro Release Methods in Liposomal Formulation Development: Challenges and Regulatory Perspective. AAPS J. 2017, 19, 1669–1681. [Google Scholar] [CrossRef] [PubMed]
- Borys, N.; Dewhirst, M.W. Drug development of lyso-thermosensitive liposomal doxorubicin: Combining hyperthermia and thermosensitive drug delivery. Adv. Drug Deliv. Rev. 2021, 178, 113985. [Google Scholar] [CrossRef] [PubMed]
- Haemmerich, D.; Motamarry, A. Thermosensitive Liposomes for Image-Guided Drug Delivery. Adv. Cancer Res. 2018, 139, 121–146. [Google Scholar] [CrossRef]
- Brummelhuis, I.S.G.; Simons, M.; Lindner, L.H.; Kort, S.; de Jong, S.; Hossann, M.; Witjes, J.A.; Oosterwijk, E. DPPG2-based thermosensitive liposomes as drug delivery system for effective muscle-invasive bladder cancer treatment in vivo. Int. J. Hyperth. 2021, 38, 1415–1424. [Google Scholar] [CrossRef]
- McGorm, H.; Roberts, L.A.; Coombes, J.S.; Peake, J.M. Turning Up the Heat: An Evaluation of the Evidence for Heating to Promote Exercise Recovery, Muscle Rehabilitation and Adaptation. Sports Med. 2018, 48, 1311–1328. [Google Scholar] [CrossRef]
- Zanoli, G.; Albarova-Corral, I.; Ancona, M.; Grattagliano, I.; Hotfiel, T.; Iolascon, G.; Krüger, K.; Rodríguez Maruri, G. Current Indications and Future Direction in Heat Therapy for Musculoskeletal Pain: A Narrative Review. Muscles 2024, 3, 212–223. [Google Scholar] [CrossRef]
- Behera, D.; Shetty, A. Effectiveness of heat and cold therapy in muscle spasm: A review. Int. J. Phys. Educ. Sports Health 2023, 10, 196–200. [Google Scholar] [CrossRef]
- Marchini, A.; Gelain, F. Synthetic scaffolds for 3D cell cultures and organoids: Applications in regenerative medicine. Crit. Rev. Biotechnol. 2022, 42, 468–486. [Google Scholar] [CrossRef]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D Cell Culture Systems: Tumor Application, Advantages, and Disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef]
- Dohi, K.; Manabe, Y.; Fujii, N.L.; Furuichi, Y. Achieving myoblast engraftment into intact skeletal muscle via extracellular matrix. Front. Cell Dev. Biol. 2025, 12, 1502332. [Google Scholar] [CrossRef] [PubMed]
- Mankhong, S.; Kim, S.; Moon, S.; Kwak, H.B.; Park, D.H.; Kang, J.H. Experimental Models of Sarcopenia: Bridging Molecular Mechanism and Therapeutic Strategy. Cells 2020, 9, 1385. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Orita, S.; Inage, K.; Yamauchi, K.; Abe, K.; Inoue, M.; Norimoto, M.; Umimura, T.; Eguchi, Y.; Fujimoto, K.; et al. Skeletal Muscle Cell Oxidative Stress as a Possible Therapeutic Target in a Denervation-Induced Experimental Sarcopenic Model. Spine 2019, 44, E446–E455. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, N.; He, L.; Xu, M.; Li, Y. 5’-Cytimidine Monophosphate Ameliorates H2O2-Induced Muscular Atrophy in C2C12 Myotubes by Activating IRS-1/Akt/S6K Pathway. Antioxidants 2024, 13, 249. [Google Scholar] [CrossRef]
- Grefte, S.; Vullinghs, S.; Kuijpers-Jagtman, A.M.; Torensma, R.; Von den Hoff, J.W. Matrigel, but not collagen I, maintains the differentiation capacity of muscle derived cells in vitro. Biomed. Mater. 2012, 7, 055004. [Google Scholar] [CrossRef]
- Grefte, S.; Adjobo-Hermans, M.J.W.; Versteeg, E.M.M.; Koopman, W.J.H.; Daamen, W.F. Impaired primary mouse myotube formation on crosslinked type I collagen films is enhanced by laminin and entactin. Acta Biomater. 2016, 30, 265–276. [Google Scholar] [CrossRef]
- Sims, C.A.; Guan, Y.; Mukherjee, S.; Singh, K.; Botolin, P.; Davila, A., Jr.; Baur, J.A. Nicotinamide mononucleotide preserves mitochondrial function and increases survival in hemorrhagic shock. JCI Insight 2018, 3, e120182. [Google Scholar] [CrossRef]
- Migliavacca, E.; Tay, S.K.H.; Patel, H.P.; Sonntag, T.; Civiletto, G.; McFarlane, C.; Forrester, T.; Barton, S.J.; Leow, M.K.; Antoun, E.; et al. Mitochondrial oxidative capacity and NAD(+) biosynthesis are reduced in human sarcopenia across ethnicities. Nat. Commun. 2019, 10, 5808. [Google Scholar] [CrossRef]
- Long, A.N.; Owens, K.; Schlappal, A.E.; Kristian, T.; Fishman, P.S.; Schuh, R.A. Effect of nicotinamide mononucleotide on brain mitochondrial respiratory deficits in an Alzheimer’s disease-relevant murine model. BMC Neurol. 2015, 15, 19. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
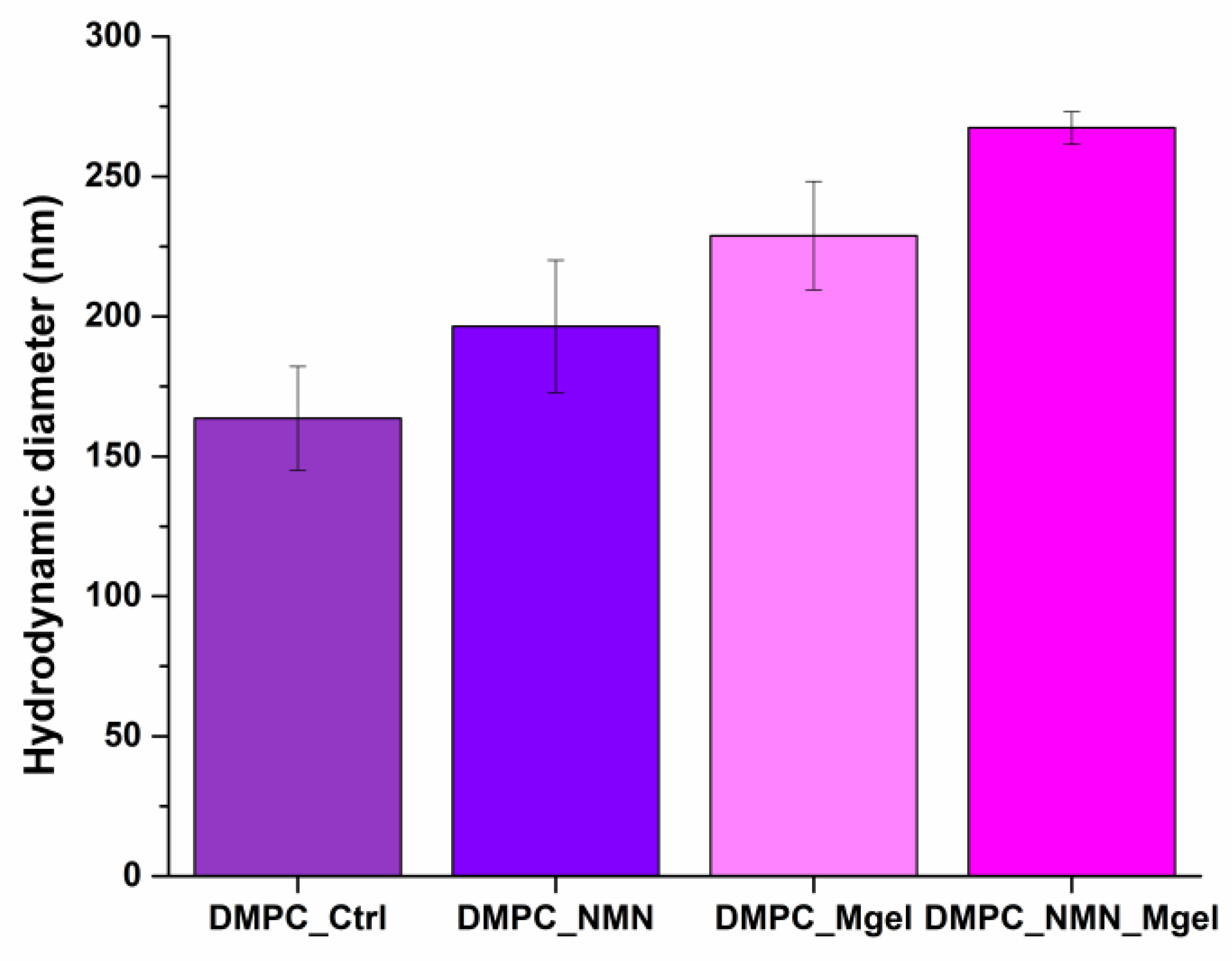
| DMPC_Ctrl | DMPC_NMN | DMPC_Mgel | DMPC_NMN_Mgel | |
|---|---|---|---|---|
| Hydrodynamic diameter (nm) | 163.6 | 196.4 | 228.8 | 267.4 |
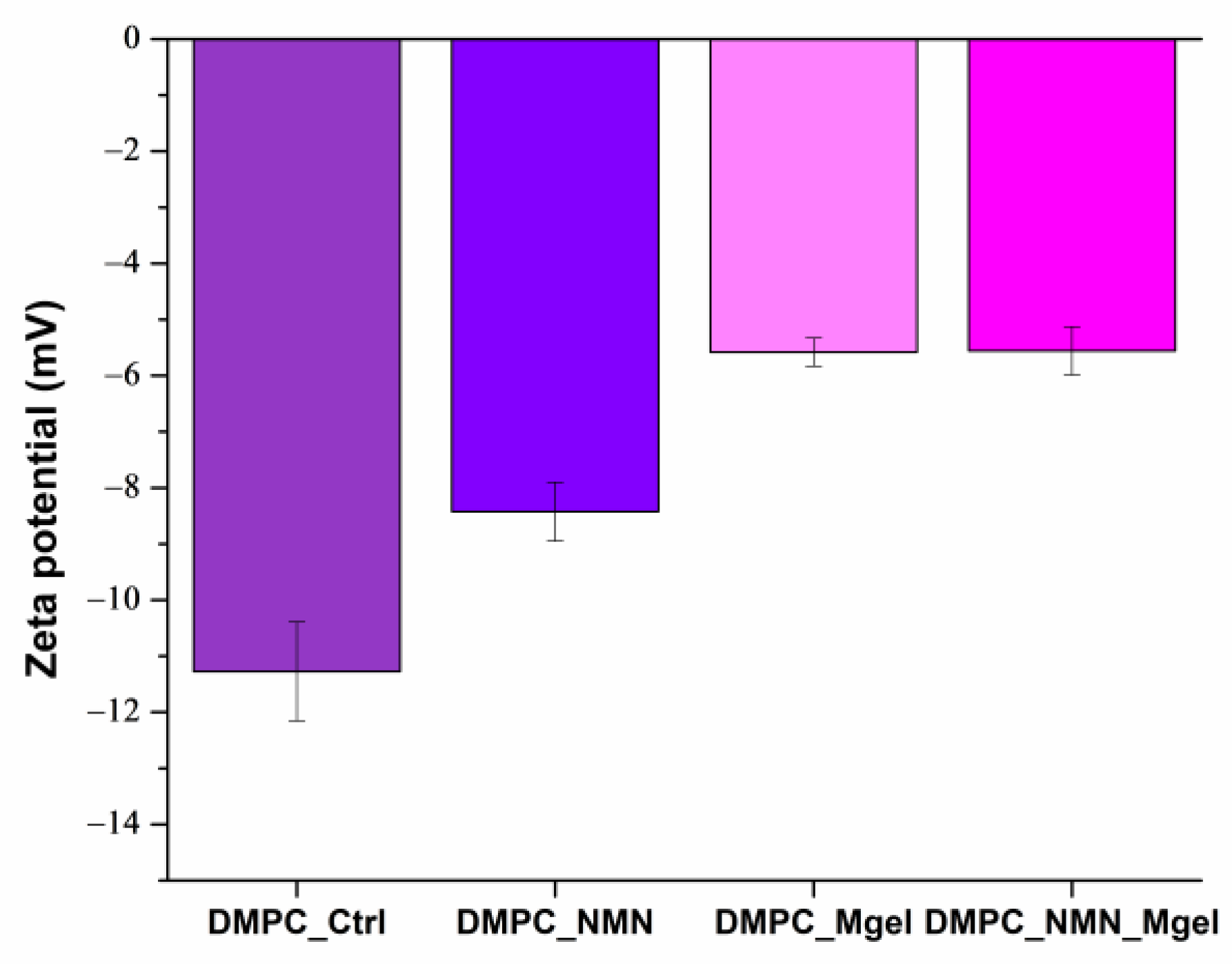
| DMPC_Ctrl | DMPC_NMN | DMPC_Mgel | DMPC_NMN_Mgel | |
|---|---|---|---|---|
| Zeta potential (mV) | −11.27 | −8.42 | −5.58 | −5.55 |
| DMPC_Ctrl | DMPC_NMN | DMPC_Mgel | DMPC_NMN_Mgel | PDI Range | |
|---|---|---|---|---|---|
| PDI | 0.081 | 0.081 | 0.081 | 0.081 | <0.1 (Monodisperse) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Najm, A.; Bîrcă, A.C.; Niculescu, A.-G.; Alberts, A.; Grumezescu, A.M.; Gălățeanu, B.; Vasile, B.Ș.; Beuran, M.; Gaspar, B.S.; Turculeț, C.Ș.; et al. DMPC-Based Liposomal Vesicles for Encapsulation and Controlled Release of NMN and Matrigel in Sarcopenia Therapy. Int. J. Mol. Sci. 2025, 26, 5594. https://doi.org/10.3390/ijms26125594
Najm A, Bîrcă AC, Niculescu A-G, Alberts A, Grumezescu AM, Gălățeanu B, Vasile BȘ, Beuran M, Gaspar BS, Turculeț CȘ, et al. DMPC-Based Liposomal Vesicles for Encapsulation and Controlled Release of NMN and Matrigel in Sarcopenia Therapy. International Journal of Molecular Sciences. 2025; 26(12):5594. https://doi.org/10.3390/ijms26125594
Chicago/Turabian StyleNajm, Alfred, Alexandra Cătălina Bîrcă, Adelina-Gabriela Niculescu, Adina Alberts, Alexandru Mihai Grumezescu, Bianca Gălățeanu, Bogdan Ștefan Vasile, Mircea Beuran, Bogdan Severus Gaspar, Claudiu Ștefan Turculeț, and et al. 2025. "DMPC-Based Liposomal Vesicles for Encapsulation and Controlled Release of NMN and Matrigel in Sarcopenia Therapy" International Journal of Molecular Sciences 26, no. 12: 5594. https://doi.org/10.3390/ijms26125594
APA StyleNajm, A., Bîrcă, A. C., Niculescu, A.-G., Alberts, A., Grumezescu, A. M., Gălățeanu, B., Vasile, B. Ș., Beuran, M., Gaspar, B. S., Turculeț, C. Ș., & Hudiță, A. (2025). DMPC-Based Liposomal Vesicles for Encapsulation and Controlled Release of NMN and Matrigel in Sarcopenia Therapy. International Journal of Molecular Sciences, 26(12), 5594. https://doi.org/10.3390/ijms26125594