
Uridine, a Therapeutic Nucleoside, Exacerbates Alcoholic Liver Disease via SRC Kinase Activation: A Network Toxicology and Molecular Dynamics Perspective


Abstract
1. Introduction
2. Results
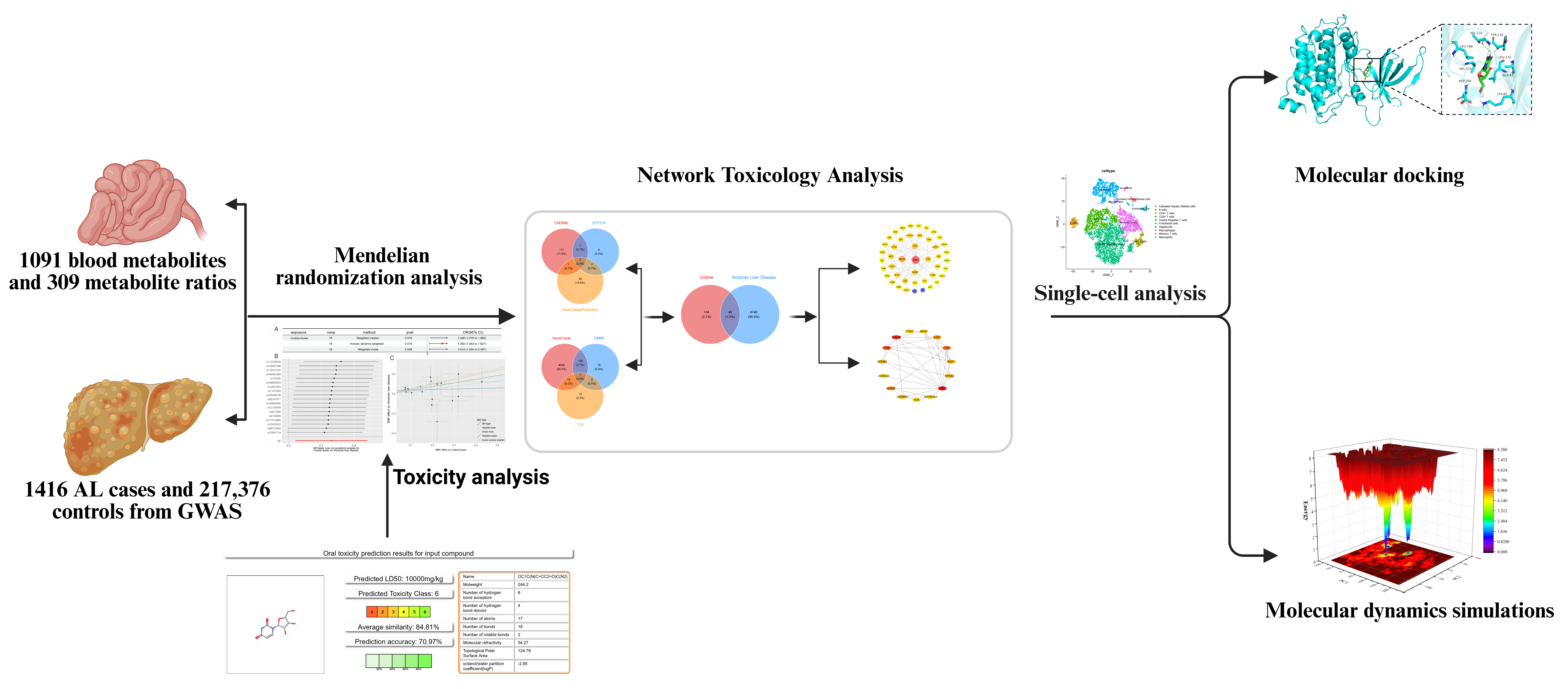
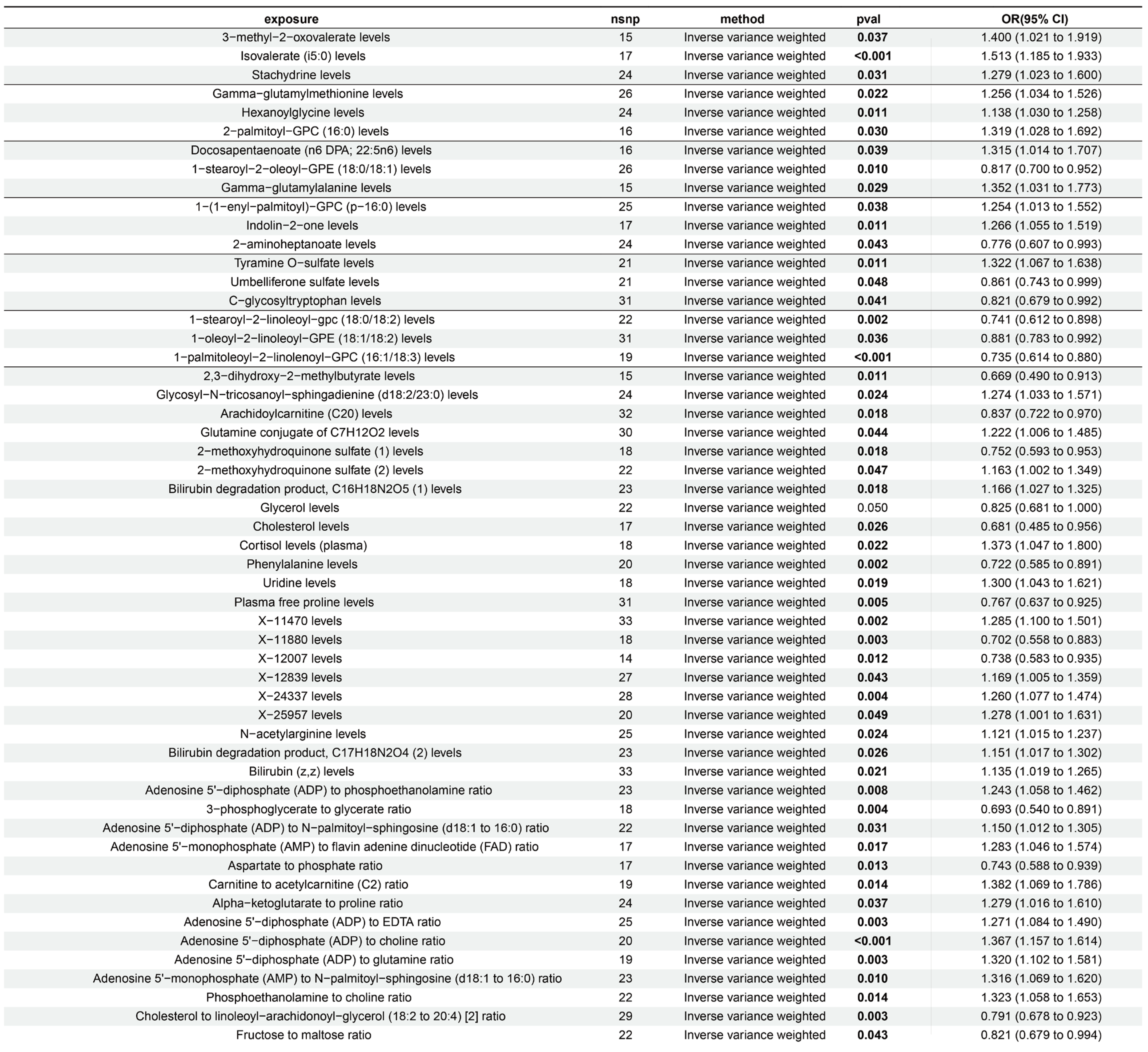
2.1. Screening of 1400 Exposure Variables
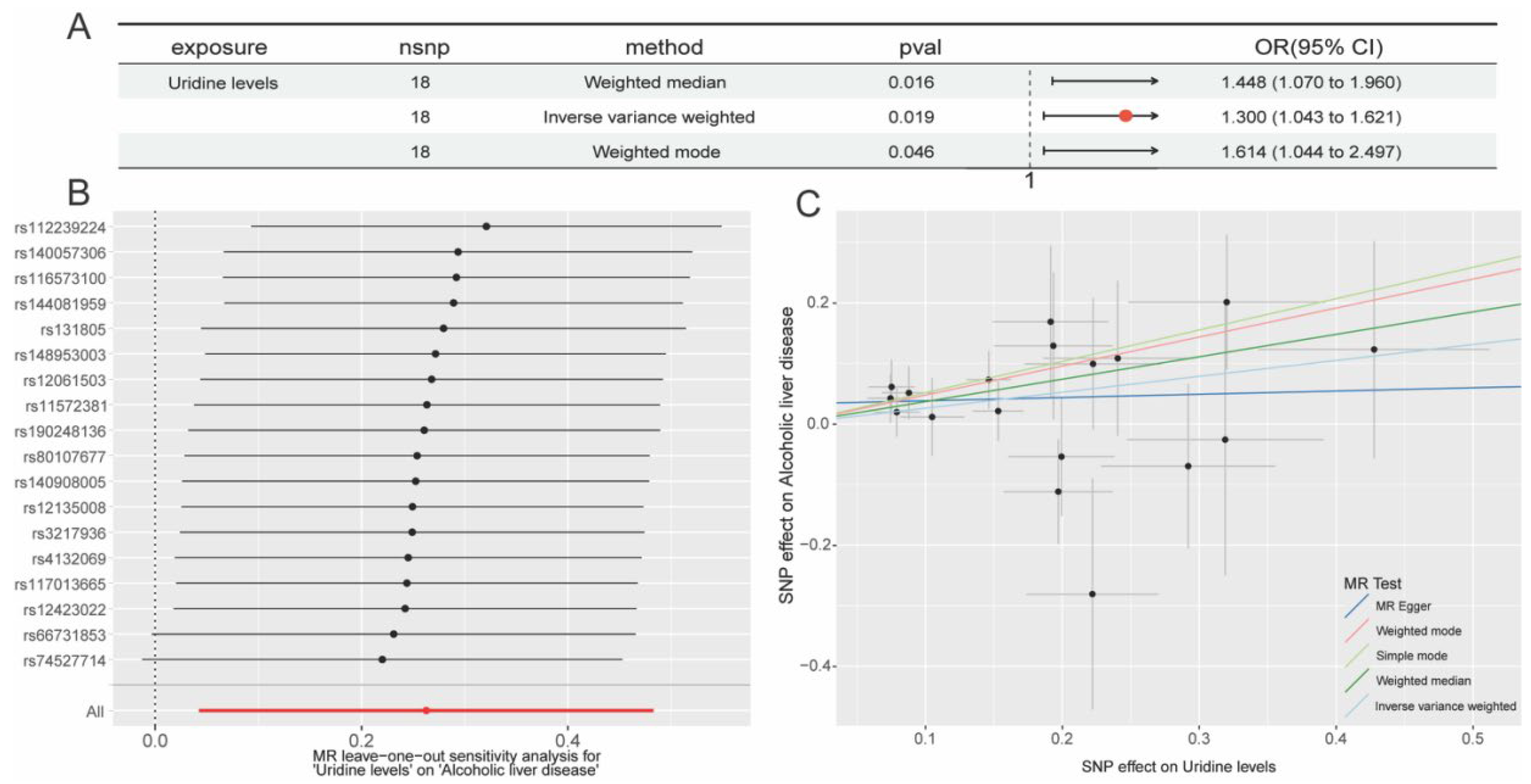
2.2. Further Screening of 33 Exposure Variables
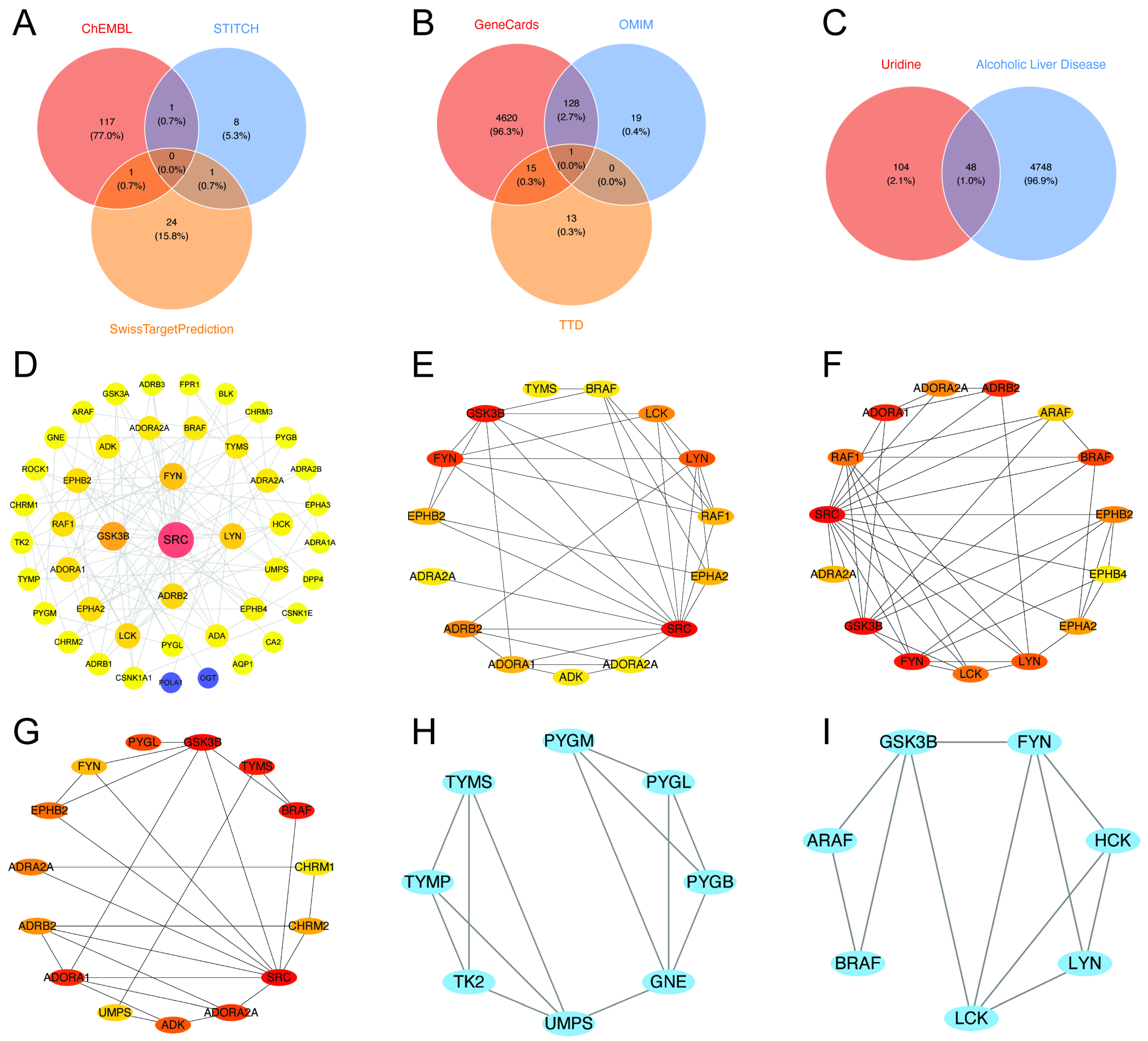
2.3. In-Depth Exploration of Uridine’s Potential Target Identification in ALD
2.4. Construction of the PPI Network and Identification of Hub Gene Connectivity
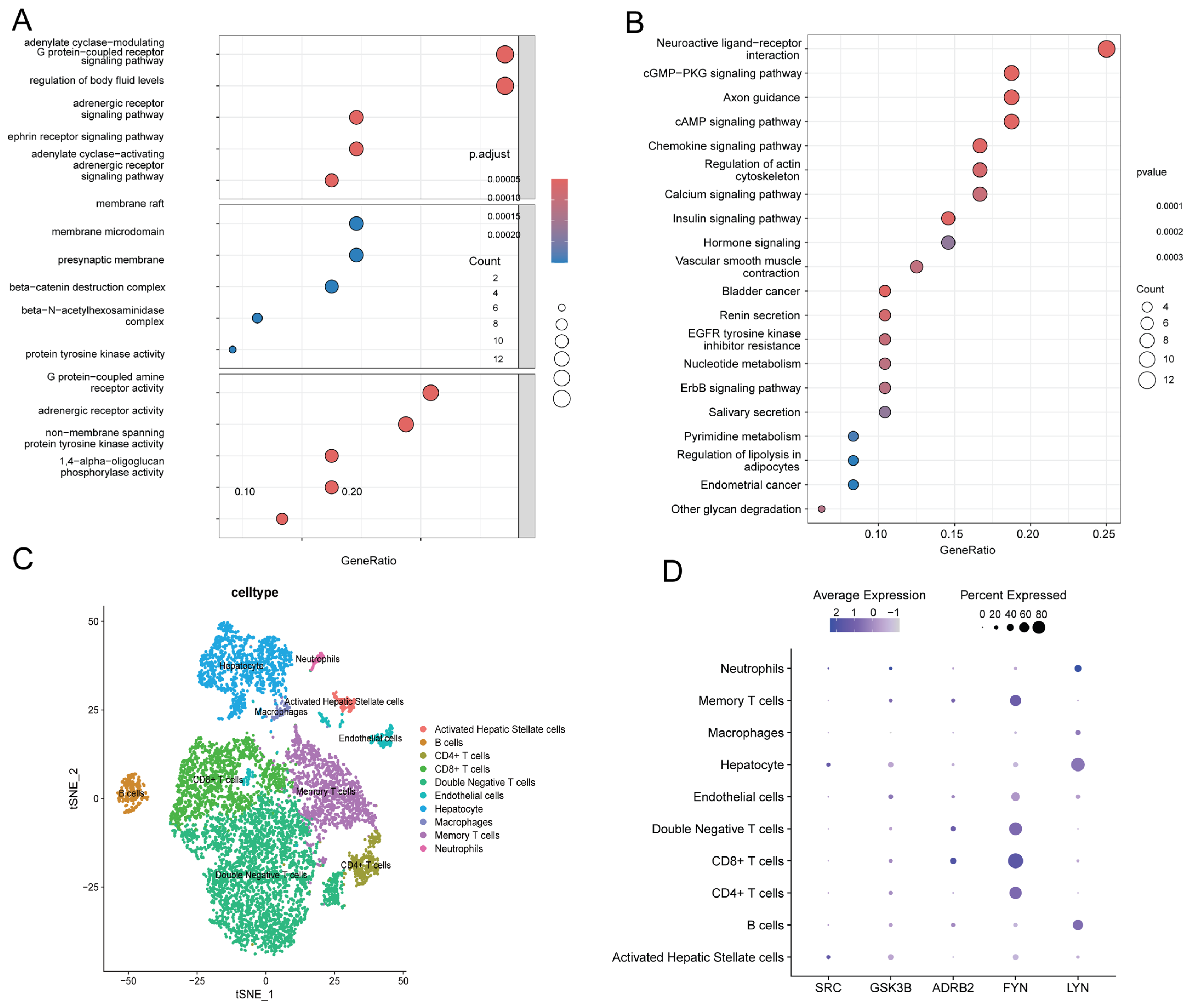
2.5. KEGG and GO Enrichment Analyses
2.6. Single-Cell Profiling Reveals Immune–Fibrotic Dysregulation in ALD
2.7. Molecular Docking Analysis of Uridine with Core Target Proteins Associated with ALD
2.8. Molecular Dynamics Simulation Analysis
3. Discussion
3.1. Mendelian Randomization Analysis
3.2. Network Toxicology and Molecular Docking: Analysis of Multi-Target Synergistic Effects
3.3. Single-Cell Transcriptomics: Cellular Heterogeneity and Microenvironmental Localization of Target Genes
3.4. MD: Analyzing Binding Stability and Allosteric Regulation from Atomic Motion
3.5. Innovations and Limitations of This Study
4. Materials and Methods
4.1. Causal Effect Analysis of 1091 Blood Metabolites, 309 Metabolite Ratios, and ALD
4.2. Screening of Exposure Factors from the Results of Section 4.1
4.3. Acquisition of Uridine Targets
4.4. Collection of AD-Related Targets
4.5. Construction of Protein–Protein Interaction Network and Selection of Hub Targets
4.6. Functional and Pathway Enrichment Analysis of Target Genes
4.7. Single-Cell RNA Analysis of Hub Genes in ALD
4.8. Molecular Docking Interactions Between Hub Targets and Uridine
4.9. Molecular Dynamics Simulation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AKT | Protein kinase B |
| ALD | Alcoholic liver disease |
| AMPK | AMP-activated protein kinase |
| ADRB2 | Adrenergic receptor beta 2 |
| AMBER | AMBER force field |
| cGMP-PKG | Cyclic GMP-dependent protein kinase G |
| CDA | Cedazuridine |
| CNS | Central nervous system |
| CHARMM | CHARMM force field |
| CIs | Confidence intervals |
| CytoHubba | Cytoscape plugin for hub gene identification |
| DFG | Asp-Phe-Gly motif |
| DisGeNET | Disease Gene Network |
| EDTA | Ethylenediaminetetraacetic acid |
| FAD | Flavin adenine dinucleotide |
| FEL | Free energy landscape |
| FYN | Fyn kinase |
| GEO | Gene Expression Omnibus |
| GPCR | G protein-coupled receptor |
| GSK3B | Glycogen synthase kinase 3 beta |
| GWAS | Genome-wide association study |
| IVW | Inverse variance weighted |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LD | Linkage disequilibrium |
| LYN | Lyn kinase |
| MD | Molecular dynamics |
| MR | Mendelian randomization |
| MR-Egger | MR-Egger regression |
| MCODE | Molecular complex detection |
| OR | Odds ratio |
| PDB | Protein Data Bank |
| PPI | Protein–protein interaction |
| PC | Principal component |
| RESP | RESPA multi-time step algorithm |
| ROS | Reactive oxygen species |
| SESN2 | Sestrin 2 |
| SPC | Simple Point Charge water model |
| SRC | Src kinase |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| THBS1 | Thrombospondin 1 |
| TIM-3 | T cell immunoglobulin and mucin domain-containing molecule 3 |
| TTD | Therapeutic Target Database |
| UPP1 | Uridine Pyrophosphatase 1 |
| UTP | Uridine Triphosphate |
| UDP-GlCNAC | Uridine Diphosphate-N-acetylglucosamine |
| VOM(bpy)2Cl | Vanadium complex [VO(bpy)2Cl] |
| PD-1 | Programmed cell death protein 1 |
| Rg | Radius of gyration |
| SASA | Solvent-accessible surface area |
| t-SNE | t-Distributed Stochastic Neighbor Embedding |
| RMSD | Root Mean Square Deviation |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| id.Exposure | Trait | nsnp | Method | pval | OR(95% CI) |
|---|---|---|---|---|---|
| GCST90199631 | 3-methyl-2-oxovalerate levels | 15 | Inverse variance weighted | 0.037 | 1.400 (1.021 to 1.919) |
| GCST90199662 | Isovalerate (i5:0) levels | 17 | Inverse variance weighted | <0.001 | 1.513 (1.185 to 1.933) |
| GCST90199738 | Stachydrine levels | 24 | Inverse variance weighted | 0.031 | 1.279 (1.023 to 1.600) |
| GCST90199751 | Gamma-glutamylmethionine levels | 26 | Inverse variance weighted | 0.022 | 1.256 (1.034 to 1.526) |
| GCST90199789 | 2-palmitoyl-GPC (16:0) levels | 16 | Inverse variance weighted | 0.03 | 1.319 (1.028 to 1.692) |
| GCST90199796 | Docosapentaenoate (n6 DPA; 22:5n6) levels | 16 | Inverse variance weighted | 0.039 | 1.315 (1.014 to 1.707) |
| GCST90199815 | 1-ribosyl-imidazoleacetate levels | 15 | Inverse variance weighted | 0.029 | 1.352 (1.031 to 1.773) |
| GCST90199899 | 1-(1-enyl-palmitoyl)-GPC (p-16:0) levels | 25 | Inverse variance weighted | 0.038 | 1.254 (1.013 to 1.552) |
| GCST90199944 | Indolin-2-one levels | 17 | Inverse variance weighted | 0.011 | 1.266 (1.055 to 1.519) |
| GCST90199988 | Tyramine O-sulfate levels | 21 | Inverse variance weighted | 0.011 | 1.322 (1.067 to 1.638) |
| GCST90200114 | Glycosyl-N-tricosanoyl-sphingadienine (d18:2/23:0) levels | 24 | Inverse variance weighted | 0.024 | 1.274 (1.033 to 1.571) |
| GCST90200167 | Glutamine conjugate of C7H12O2 levels | 30 | Inverse variance weighted | 0.044 | 1.222 (1.006 to 1.485) |
| GCST90200273 | 2-methoxyhydroquinone sulfate (2) levels | 22 | Inverse variance weighted | 0.047 | 1.163 (1.002 to 1.349) |
| GCST90200277 | Bilirubin degradation product, C16H18N2O5 (1) levels | 23 | Inverse variance weighted | 0.018 | 1.166 (1.027 to 1.325) |
| GCST90200378 | Cortisol levels (plasma) | 18 | Inverse variance weighted | 0.022 | 1.373 (1.047 to 1.800) |
| GCST90200407 | Uridine levels | 18 | Inverse variance weighted | 0.019 | 1.300 (1.043 to 1.621) |
| GCST90200470 | X-11470 levels | 33 | Inverse variance weighted | 0.002 | 1.285 (1.100 to 1.501) |
| GCST90200518 | X-12839 levels | 27 | Inverse variance weighted | 0.043 | 1.169 (1.005 to 1.359) |
| GCST90200629 | X-24337 levels | 28 | Inverse variance weighted | 0.004 | 1.260 (1.077 to 1.474) |
| GCST90200660 | X-25957 levels | 20 | Inverse variance weighted | 0.049 | 1.278 (1.001 to 1.631) |
| GCST90200679 | N-acetylarginine levels | 25 | Inverse variance weighted | 0.024 | 1.121 (1.015 to 1.237) |
| GCST90200702 | Bilirubin degradation product, C17H18N2O4 (2) levels | 23 | Inverse variance weighted | 0.026 | 1.151 (1.017 to 1.302) |
| GCST90200704 | Bilirubin (z,z) levels | 33 | Inverse variance weighted | 0.021 | 1.135 (1.019 to 1.265) |
| GCST90200716 | Adenosine 5′-diphosphate (ADP) to phosphoethanolamine ratio | 23 | Inverse variance weighted | 0.008 | 1.243 (1.058 to 1.462) |
| GCST90200823 | Adenosine 5′-diphosphate (ADP) to N-palmitoyl-sphingosine (d18:1 to 16:0) ratio | 22 | Inverse variance weighted | 0.031 | 1.150 (1.012 to 1.305) |
| GCST90200843 | Adenosine 5′-monophosphate (AMP) to flavin adenine dinucleotide (FAD) ratio | 17 | Inverse variance weighted | 0.017 | 1.283 (1.046 to 1.574) |
| GCST90200930 | Carnitine to acetylcarnitine (C2) ratio | 19 | Inverse variance weighted | 0.014 | 1.382 (1.069 to 1.786) |
| GCST90200933 | Alpha-ketoglutarate to proline ratio | 24 | Inverse variance weighted | 0.037 | 1.279 (1.016 to 1.610) |
| GCST90200940 | Adenosine 5′-diphosphate (ADP) to EDTA ratio | 25 | Inverse variance weighted | 0.003 | 1.271 (1.084 to 1.490) |
| GCST90200958 | Adenosine 5′-diphosphate (ADP) to choline ratio | 20 | Inverse variance weighted | <0.001 | 1.367 (1.157 to 1.614) |
| GCST90200962 | Adenosine 5′-diphosphate (ADP) to glutamine ratio | 19 | Inverse variance weighted | 0.003 | 1.320 (1.102 to 1.581) |
| GCST90200969 | Adenosine 5′-monophosphate (AMP) to N-palmitoyl-sphingosine (d18:1 to 16:0) ratio | 23 | Inverse variance weighted | 0.01 | 1.316 (1.069 to 1.620) |
| GCST90200980 | Phosphoethanolamine to choline ratio | 22 | Inverse variance weighted | 0.014 | 1.323 (1.058 to 1.653) |
| Target Protein | Mode | Binding Energy (kcal/mol) | RMSD (l.b.) | RMSD (u.b.) |
|---|---|---|---|---|
| GSK3β | 1 | −6.2 | 0 | 0 |
| GSK3β | 2 | −5.9 | 10.48 | 11.417 |
| GSK3β | 3 | −5.7 | 10.587 | 12.213 |
| GSK3β | 4 | −5.7 | 11.35 | 12.751 |
| GSK3β | 5 | −5.6 | 2.094 | 2.739 |
| GSK3β | 6 | −5.6 | 11.883 | 13.019 |
| GSK3β | 7 | −5.6 | 10.464 | 12.336 |
| GSK3β | 8 | −5.5 | 11.958 | 13.951 |
| GSK3β | 9 | −5.5 | 10.536 | 12.283 |
| FYN | 1 | −5.8 | 0 | 0 |
| FYN | 2 | −5.5 | 4.445 | 8.273 |
| FYN | 3 | −5.3 | 20.255 | 21.76 |
| FYN | 4 | −5.3 | 19.685 | 21.447 |
| FYN | 5 | −5.2 | 12.991 | 14.232 |
| FYN | 6 | −5.1 | 16.306 | 18.021 |
| FYN | 7 | −5.1 | 19.551 | 20.597 |
| FYN | 8 | −5 | 15.97 | 17.596 |
| FYN | 9 | −5 | 20.298 | 21.485 |
| LYN | 1 | −4.9 | 0 | 0 |
| LYN | 2 | −4.6 | 2.393 | 5.332 |
| LYN | 3 | −4.6 | 1.791 | 5.382 |
| LYN | 4 | −4.5 | 2.532 | 5.797 |
| LYN | 5 | −4.4 | 2.914 | 4.5 |
| LYN | 6 | −4.2 | 2.204 | 3.657 |
| LYN | 7 | −4 | 2.433 | 4.414 |
| LYN | 8 | −4 | 18.351 | 19.299 |
| LYN | 9 | −4 | 2.281 | 5.179 |
| SRC | 1 | −5.6 | 0 | 0 |
| SRC | 2 | −5.2 | 2.119 | 2.344 |
| SRC | 3 | −4.7 | 2.099 | 4.939 |
| SRC | 4 | −4.7 | 2.351 | 4.883 |
| SRC | 5 | −4.7 | 10.497 | 12.017 |
| SRC | 6 | −4.7 | 2.225 | 5.673 |
| SRC | 7 | −4.6 | 11.765 | 13.288 |
| SRC | 8 | −4.6 | 11.315 | 12.514 |
| SRC | 9 | −4.5 | 2.48 | 3.411 |
| ADRB2 | 1 | −7.1 | 0 | 0 |
| ADRB2 | 2 | −6.9 | 6.448 | 8.318 |
| ADRB2 | 3 | −6.8 | 5.79 | 7.32 |
| ADRB2 | 4 | −6.8 | 5.218 | 7.241 |
| ADRB2 | 5 | −6.8 | 5.791 | 7.899 |
| ADRB2 | 6 | −6.7 | 6.054 | 7.859 |
| ADRB2 | 7 | −6.6 | 2.066 | 3.398 |
| ADRB2 | 8 | −6.6 | 5.869 | 8.013 |
| ADRB2 | 9 | −6.5 | 2.246 | 5.281 |
References
- Roerecke, M.; Vafaei, A.; Hasan, O.S.M.; Chrystoja, B.R.; Cruz, M.; Lee, R.; Neuman, M.G.; Rehm, J. Alcohol Consumption and Risk of Liver Cirrhosis: A Systematic Review and Meta-Analysis. Am. J. Gastroenterol. 2019, 114, 1574–1586. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E.; Guo, G.L. Understanding Environmental Contaminants’ Direct Effects on Non-alcoholic Fatty Liver Disease Progression. Curr. Environ. Health Rep. 2019, 6, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.Q. Heavy Metals and Pesticides Toxicity in Agricultural Soil and Plants: Ecological Risks and Human Health Implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhou, H.; Yang, S.; He, C. Combining mendelian randomization analysis and network toxicology strategy to identify causality and underlying mechanisms of environmental pollutants with glioblastoma: A study of Methyl-4-hydroxybenzoate. Ecotoxicol. Environ. Saf. 2024, 287, 117311. [Google Scholar] [CrossRef]
- Qu, T.; Sun, Q.; Tan, B.; Wei, H.; Qiu, X.; Xu, X.; Gao, H.; Zhang, S. Integration of network toxicology and transcriptomics reveals the novel neurotoxic mechanisms of 2, 2′, 4, 4′-tetrabromodiphenyl ether. J. Hazard. Mater. 2024, 486, 136999. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, Y.; Deng, Y.; Gao, L. Uridine and its role in metabolic diseases, tumors, and neurodegenerative diseases. Front. Physiol. 2024, 15, 1360891. [Google Scholar] [CrossRef]
- Venediktova, N.; Solomadin, I.; Nikiforova, A.; Belosludtsev, K.N.; Mironova, G. Effects of the Long-Term Administration of Uridine on the Functioning of Rat Liver Mitochondria in Hyperthyroidism. Int. J. Mol. Sci. 2023, 24, 16730. [Google Scholar] [CrossRef]
- Levenfors, J.J.; Bjerketorp, J.; Guss, B.; Nord, C.; Cao, S.; Hughes, D.; Broberg, A.; Oberg, B. Repurposing zidovudine and 5-fluoro-2′-deoxyuridine as antibiotic drugs made possible by synergy with both trimethoprim and the mitochondrial toxicity-reducing agent uridine. J. Antimicrob. Chemother. 2025, 80, 509–517. [Google Scholar] [CrossRef]
- Wright, N.J.; Zhang, F.; Suo, Y.; Kong, L.; Yin, Y.; Fedor, J.G.; Sharma, K.; Borgnia, M.J.; Im, W.; Lee, S.Y. Antiviral drug recognition and elevator-type transport motions of CNT3. Nat. Chem. Biol. 2024, 20, 1144–1153. [Google Scholar] [CrossRef]
- Miao, Y.S.; Wang, J.Y.; Zhuang, R.R.; Huo, X.K.; Yi, Z.C.; Sun, X.N.; Yu, Z.L.; Tian, X.G.; Ning, J.; Feng, L.; et al. A high-affinity fluorescent probe for human uridine-disphosphate glucuronosyltransferase 1A9 function monitoring under environmental pollutant exposure. J. Hazard. Mater. 2024, 465, 133439. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, S.; Xie, C.; Fang, J. Uridine Metabolism and Its Role in Glucose, Lipid, and Amino Acid Homeostasis. Biomed Res. Int. 2020, 2020, 7091718. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lu, T.; Pettersson-Kymmer, U.; Stewart, I.D.; Butler-Laporte, G.; Nakanishi, T.; Cerani, A.; Liang, K.Y.H.; Yoshiji, S.; Willett, J.D.S.; et al. Genomic atlas of the plasma metabolome prioritizes metabolites implicated in human diseases. Nat. Genet. 2023, 55, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, D.; Zhang, L.; Xiao, Y.; Wu, Q.; Wang, Y.; Chen, J.; Zhan, Q. Src heterodimerically activates Lyn or Fyn to serve as targets for the diagnosis and treatment of esophageal squamous cell carcinoma. Sci. China Life Sci. 2023, 66, 1245–1263. [Google Scholar] [CrossRef] [PubMed]
- Shimpi, A.A.; Williams, E.D.; Ling, L.; Tamir, T.; White, F.M.; Fischbach, C. Phosphoproteomic Changes Induced by Cell-Derived Matrix and Their Effect on Tumor Cell Migration and Cytoskeleton Remodeling. ACS Biomater. Sci. Eng. 2023, 9, 6835–6848. [Google Scholar] [CrossRef]
- Vom Stein, A.F.; Rebollido-Rios, R.; Lukas, A.; Koch, M.; von Lom, A.; Reinartz, S.; Bachurski, D.; Rose, F.; Bozek, K.; Abdallah, A.T.; et al. LYN kinase programs stromal fibroblasts to facilitate leukemic survival via regulation of c-JUN and THBS1. Nat. Commun. 2023, 14, 1330. [Google Scholar] [CrossRef]
- Khoury, M.; Guo, Q.; Furuta, K.; Correia, C.; Meroueh, C.; Kim Lee, H.S.; Warasnhe, K.; Valenzuela-Perez, L.; Mazar, A.P.; Kim, I.; et al. Glycogen synthase kinase 3 activity enhances liver inflammation in MASH. JHEP Rep. 2024, 6, 101073. [Google Scholar] [CrossRef]
- Gao, T.; Wang, J.; Xiao, M.; Wang, J.; Wang, S.; Tang, Y.; Zhang, J.; Lu, G.; Guo, H.; Guo, Y.; et al. SESN2-Mediated AKT/GSK-3β/NRF2 Activation to Ameliorate Adriamycin Cardiotoxicity in High-Fat Diet-Induced Obese Mice. Antioxid. Redox Signal 2024, 40, 598–615. [Google Scholar] [CrossRef]
- Bagnato, A.; Rosano, L. New Routes in GPCR/β-Arrestin-Driven Signaling in Cancer Progression and Metastasis. Front. Pharmacol. 2019, 10, 114. [Google Scholar] [CrossRef]
- Zi, L.; Ma, W.; Zhang, L.; Qiao, B.; Qiu, Z.; Xu, J.; Zhang, J.; Ye, Y.; Yang, Y.; Dong, K.; et al. Uridine Inhibits Hepatocellular Carcinoma Cell Development by Inducing Ferroptosis. J. Clin. Med. 2023, 12, 3552. [Google Scholar] [CrossRef]
- Uspalenko, N.I.; Mosentsov, A.A.; Khmil, N.V.; Pavlik, L.L.; Belosludtseva, N.V.; Khunderyakova, N.V.; Shigaeva, M.I.; Medvedeva, V.P.; Malkov, A.E.; Kitchigina, V.F.; et al. Uridine as a Regulator of Functional and Ultrastructural Changes in the Brain of Rats in a Model of 6-OHDA-Induced Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 14304. [Google Scholar] [CrossRef]
- Zhou, N.N.; Wang, T.; Lin, Y.X.; Xu, R.; Wu, H.X.; Ding, F.F.; Qiao, F.; Du, Z.Y.; Zhang, M.L. Uridine alleviates high-carbohydrate diet-induced metabolic syndromes by activating sirt1/AMPK signaling pathway and promoting glycogen synthesis in Nile tilapia (Oreochromis niloticus). Anim. Nutr. 2023, 14, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.T.; Wood, D.M.; Dargan, P.I. Review of the fluoropyrimidine antidote uridine triacetate. Br. J. Clin. Pharmacol. 2025, 91, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Jacob, A.; Sekkath Veedu, J.; Selene, I.; Raj, R.; Kannan, L.; Patel, R. Case report: Uridine triacetate in the management of delayed onset 5-fluorouracil toxicity: A case report and review of literature. Front. Pharmacol. 2022, 13, 977734. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; Bhatti, M.R.; Kellenberger, A.; Sun, W.; Modica, S.; Horing, M.; Liebisch, G.; Krieger, J.P.; Wolfrum, C.; Challa, T.D. A low-carbohydrate diet induces hepatic insulin resistance and metabolic associated fatty liver disease in mice. Mol. Metab. 2023, 69, 101675. [Google Scholar] [CrossRef]
- Sheel, A.; Bae, J.; Asada, A.; Otterson, G.A.; Baliga, R.R.; Koenig, K.L. Reversible cardiomyopathy in a patient with chronic myelomonocytic leukemia treated with decitabine/cedazuridine: A case report. Cardiooncology 2023, 9, 4. [Google Scholar] [CrossRef]
- Davey Smith, G.; Hemani, G. Mendelian randomization: Genetic anchors for causal inference in epidemiological studies. Hum. Mol. Genet. 2014, 23, R89–R98. [Google Scholar] [CrossRef]
- Gao, H.; Li, J.; Ma, Q.; Zhang, Q.; Li, M.; Hu, X. Causal Associations of Environmental Pollution and Cardiovascular Disease: A Two-Sample Mendelian Randomization Study. Glob. Heart 2024, 19, 52. [Google Scholar] [CrossRef]
- Skinner, O.S.; Blanco-Fernandez, J.; Goodman, R.P.; Kawakami, A.; Shen, H.; Kemeny, L.V.; Joesch-Cohen, L.; Rees, M.G.; Roth, J.A.; Fisher, D.E.; et al. Salvage of ribose from uridine or RNA supports glycolysis in nutrient-limited conditions. Nat. Metab. 2023, 5, 765–776. [Google Scholar] [CrossRef]
- Lane, A.N.; Fan, T.W. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef]
- Strefeler, A.; Blanco-Fernandez, J.; Jourdain, A.A. Nucleosides are overlooked fuels in central carbon metabolism. Trends Endocrinol. Metab. 2024, 35, 290–299. [Google Scholar] [CrossRef]
- Sheth, V.G.; Sharma, N.; Kabeer, S.W.; Tikoo, K. Lactobacillus rhamnosus supplementation ameliorates high fat diet-induced epigenetic alterations and prevents its intergenerational inheritance. Life Sci. 2022, 311, 121151. [Google Scholar] [CrossRef] [PubMed]
- Sol Dourdin, T.; Guyomard, K.; Rabiller, M.; Houssais, N.; Cormier, A.; Le Monier, P.; Sussarellu, R.; Riviere, G. Ancestors’ Gift: Parental Early Exposure to the Environmentally Realistic Pesticide Mixture Drives Offspring Phenotype in a Larger Extent Than Direct Exposure in the Pacific Oyster, Crassostrea gigas. Environ. Sci. Technol. 2024, 58, 1865–1876. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, J.; Chen, Z.; Wu, W.; Lu, L.; Cheng, Y.; Li, S.; Chen, L.; Tan, X.; Yang, L.; et al. DNA methylation reprogramming mediates transgenerational diabetogenic effect induced by early-life p,p’-DDE exposure. Chemosphere 2024, 349, 140907. [Google Scholar] [CrossRef] [PubMed]
- Klibaner-Schiff, E.; Simonin, E.M.; Akdis, C.A.; Cheong, A.; Johnson, M.M.; Karagas, M.R.; Kirsh, S.; Kline, O.; Mazumdar, M.; Oken, E.; et al. Environmental exposures influence multigenerational epigenetic transmission. Clin. Epigenet. 2024, 16, 145. [Google Scholar] [CrossRef]
- Baccarelli, A.; Bollati, V. Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef]
- Zhu, L.; Yuhan, J.; Huang, K.; He, X.; Liang, Z.; Xu, W. Multidimensional analysis of the epigenetic alterations in toxicities induced by mycotoxins. Food Chem. Toxicol. 2021, 153, 112251. [Google Scholar] [CrossRef]
- Langie, S.A.; Koppen, G.; Desaulniers, D.; Al-Mulla, F.; Al-Temaimi, R.; Amedei, A.; Azqueta, A.; Bisson, W.H.; Brown, D.G.; Brunborg, G.; et al. Causes of genome instability: The effect of low dose chemical exposures in modern society. Carcinogenesis 2015, 36 (Suppl. S1), S61–S88. [Google Scholar] [CrossRef]
- Liu, B.; Zeng, Q.; Chen, H.; Liao, J.; Bai, Y.; Han, Q.; Qiao, N.; Wang, S.; Mehmood, K.; Hussain, R.; et al. The hepatotoxicity of altrazine exposure in mice involves the intestinal microbiota. Chemosphere 2021, 272, 129572. [Google Scholar] [CrossRef]
- Liao, J.; Liu, Y.; Yi, J.; Li, Y.; Li, Q.; Li, Y.; Shang, P.; Guo, J.; Hu, L.; Pan, J.; et al. Gut microbiota disturbance exaggerates battery wastewater-induced hepatotoxicity through a gut-liver axis. Sci. Total Environ. 2022, 809, 152188. [Google Scholar] [CrossRef]
- Gieger, C.; Geistlinger, L.; Altmaier, E.; Hrabe de Angelis, M.; Kronenberg, F.; Meitinger, T.; Mewes, H.W.; Wichmann, H.E.; Weinberger, K.M.; Adamski, J.; et al. Genetics meets metabolomics: A genome-wide association study of metabolite profiles in human serum. PLoS Genet. 2008, 4, e1000282. [Google Scholar] [CrossRef]
- Bian, Q.; Wang, Y.J.; Shi, Q. Progress in research on focal adhesion kinase signal transduction and cross-talk. Zhong Xi Yi Jie He Xue Bao 2005, 3, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Dobie, R.; Wilson-Kanamori, J.R.; Dora, E.F.; Henderson, B.E.P.; Luu, N.T.; Portman, J.R.; Matchett, K.P.; Brice, M.; Marwick, J.A.; et al. Resolving the fibrotic niche of human liver cirrhosis at single-cell level. Nature 2019, 575, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, X.; Chu, Y.; Li, C.; Li, X.; Meng, X.; Fang, Y.; No, K.T.; Mao, J.; Zeng, X. Exploring the Conformational Ensembles of Protein-Protein Complex with Transformer-Based Generative Model. J. Chem. Theory Comput. 2024, 20, 4469–4480. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, S.; Kanagalingam, G.; Fleckenstein, F.; Froescher, D.; Hasse, H.; Stephan, S. Extension of the MolMod Database to Transferable Force Fields. J. Chem. Inf. Model. 2023, 63, 7148–7158. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Qin, Z.; Kang, Y.; Hong, X.; Hou, T. Quasiclassical Trajectory Simulation as a Protocol to Build Locally Accurate Machine Learning Potentials. J. Chem. Inf. Model. 2023, 63, 1133–1142. [Google Scholar] [CrossRef]
- Xu, X.; Li, J. Deciphering Dynamics of the Cl + SiH4→H + SiH3Cl Reaction on a Machine Learning Made Globally Accurate Full-Dimensional Potential Energy Surface. J. Phys. Chem. A 2022, 126, 6456–6466. [Google Scholar] [CrossRef]
- Herringer, N.S.M.; Dasetty, S.; Gandhi, D.; Lee, J.; Ferguson, A.L. Permutationally Invariant Networks for Enhanced Sampling (PINES): Discovery of Multimolecular and Solvent-Inclusive Collective Variables. J. Chem. Theory Comput. 2024, 20, 178–198. [Google Scholar] [CrossRef]
- Fang, W.; Zhu, Y.C.; Cheng, Y.; Hao, Y.P.; Richardson, J.O. Robust Gaussian Process Regression Method for Efficient Tunneling Pathway Optimization: Application to Surface Processes. J. Chem. Theory Comput. 2024, 20, 3766–3778. [Google Scholar] [CrossRef]
- Toopradab, B.; Xie, W.; Duan, L.; Hengphasatporn, K.; Harada, R.; Sinsulpsiri, S.; Shigeta, Y.; Shi, L.; Maitarad, P.; Rungrotmongkol, T. Machine learning-based QSAR and LB-PaCS-MD guided design of SARS-CoV-2 main protease inhibitors. Bioorg. Med. Chem. Lett. 2024, 110, 129852. [Google Scholar] [CrossRef]
- Khanal, P.; Zargari, F.; Dey, Y.N.; Nikfarjam, Z. Olanzapine manipulates neuroactive signals and may onset metabolic disturbances. J. Biomol. Struct. Dyn. 2024, 42, 6613–6627. [Google Scholar] [CrossRef]
- Santos, T.M.R.; Tavares, C.A.; da Cunha, E.F.F.; Ramalho, T.C. Vanadium complex as a potential modulator of the autophagic mechanism through proteins PI3K and ULK1: Development, validation and biological implications of a specific force field for [VO(bpy)(2)Cl]. J. Biomol. Struct. Dyn. 2024, 42, 9118–9132. [Google Scholar] [CrossRef] [PubMed]
- Kurki, M.; Nesterenko, A.M.; Alsaker, N.E.; Ferreira, T.M.; Kyllonen, S.; Poso, A.; Bartos, P.; Miettinen, M.S. Solid-State NMR Validation of OPLS4: Structure of PC-Lipid Bilayers and Its Modulation by Dehydration. J. Phys. Chem. B 2024, 128, 12483–12492. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ji, H.; Li, Z.; Luan, Y.; Zhu, C.; Li, D.; Gao, Y.; Yan, Z. Gut microbiota causally impacts adrenal function: A two-sample mendelian randomization study. Sci. Rep. 2024, 14, 23338. [Google Scholar] [CrossRef] [PubMed]
- Genomes Project, C.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Banerjee, P.; Kemmler, E.; Dunkel, M.; Preissner, R. ProTox 3.0: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2024, 52, W513–W520. [Google Scholar] [CrossRef]
- Zhao, X.; Xiu, J.; Yang, H.; Han, W.; Jin, Y. Network Pharmacology and Bioinformatics Study of Six Medicinal Food Homologous Plants Against Colorectal Cancer. Int. J. Mol. Sci. 2025, 26, 930. [Google Scholar] [CrossRef]
- Khan, A.; Mohammad, T.; Shamsi, A.; Hussain, A.; Alajmi, M.F.; Husain, S.A.; Iqbal, M.A.; Hassan, M.I. Identification of plant-based hexokinase 2 inhibitors: Combined molecular docking and dynamics simulation studies. J. Biomol. Struct. Dyn. 2022, 40, 10319–10331. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Wang, Z.; Wang, J.; Xu, S.; Zhang, T. Uridine, a Therapeutic Nucleoside, Exacerbates Alcoholic Liver Disease via SRC Kinase Activation: A Network Toxicology and Molecular Dynamics Perspective. Int. J. Mol. Sci. 2025, 26, 5473. https://doi.org/10.3390/ijms26125473
Liu Z, Wang Z, Wang J, Xu S, Zhang T. Uridine, a Therapeutic Nucleoside, Exacerbates Alcoholic Liver Disease via SRC Kinase Activation: A Network Toxicology and Molecular Dynamics Perspective. International Journal of Molecular Sciences. 2025; 26(12):5473. https://doi.org/10.3390/ijms26125473
Chicago/Turabian StyleLiu, Zhenyu, Zhihao Wang, Jie Wang, Shiquan Xu, and Tong Zhang. 2025. "Uridine, a Therapeutic Nucleoside, Exacerbates Alcoholic Liver Disease via SRC Kinase Activation: A Network Toxicology and Molecular Dynamics Perspective" International Journal of Molecular Sciences 26, no. 12: 5473. https://doi.org/10.3390/ijms26125473
APA StyleLiu, Z., Wang, Z., Wang, J., Xu, S., & Zhang, T. (2025). Uridine, a Therapeutic Nucleoside, Exacerbates Alcoholic Liver Disease via SRC Kinase Activation: A Network Toxicology and Molecular Dynamics Perspective. International Journal of Molecular Sciences, 26(12), 5473. https://doi.org/10.3390/ijms26125473