

Exosomes: A Promising Cell-Free Therapeutic Tool for Treating Cutaneous Nerve Injuries and Promoting Wound Healing

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Cutaneous Nerve Injury
2.1. Causes of Cutaneous Nerve Injury
2.1.1. External Factors
2.1.2. Endogenous Factors
3. The Role of Nerves in the Regeneration of Skin Trauma Regeneration and Scar Healing
3.1. Nerves May Promote the Healing of Skin Wounds
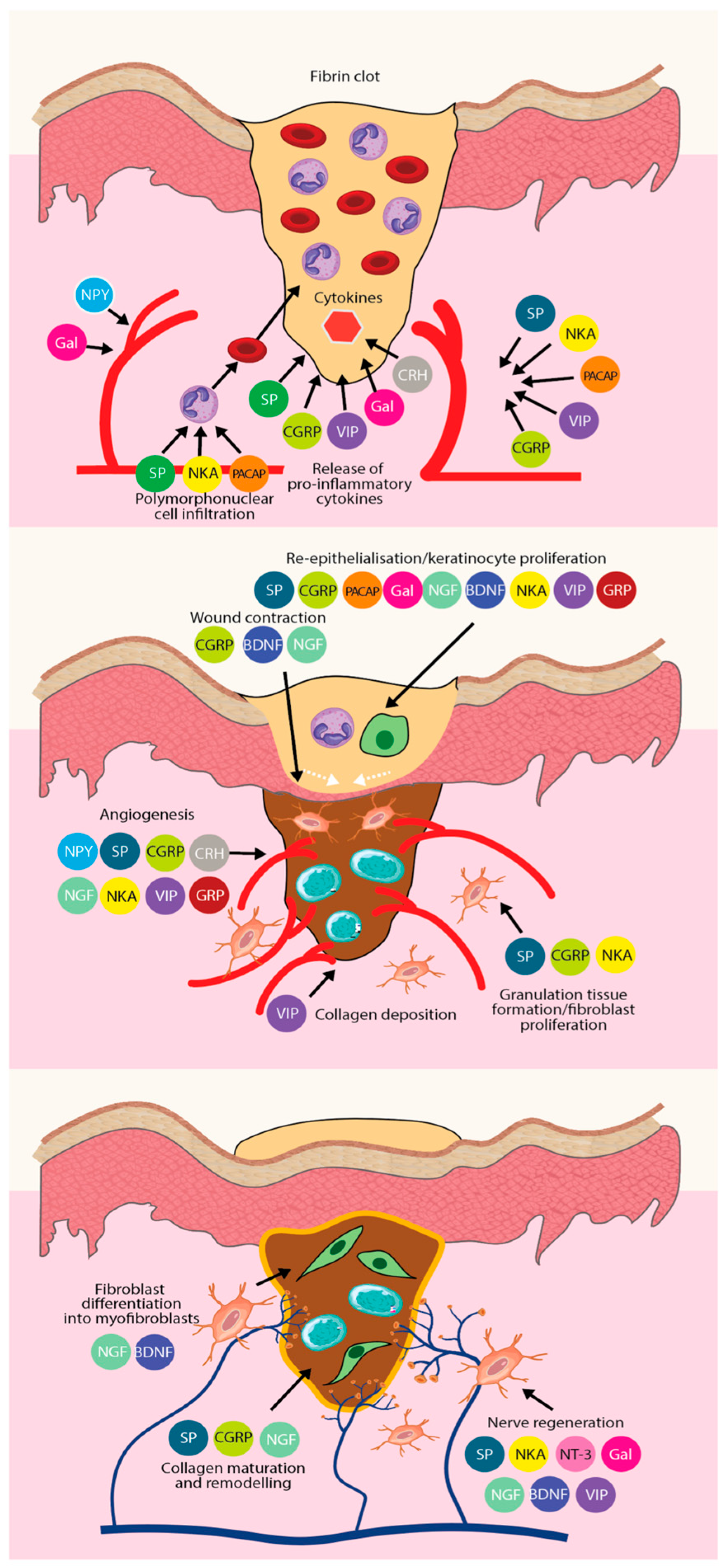
3.2. Neuropeptides Promote Wound Healing
4. The Role of Exosomes in the Regulation of Cutaneous Nerve Injury
4.1. Regulatory Role of Exosomes in Skin Damage
4.2. Regulation of Exosomes in Peripheral Nerve Injury
4.3. Exosome Modulation in Nerve Injury Pain
4.4. Synergistic Modulation of Exosomes in the Treatment of Skin Lesions and Nerve Damage
5. MSC-Exo Applications and Challenges
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, R.; Geyer, S.; Weninger, W.; Guimberteau, J.C.; Wong, J.K. The dynamic anatomy and patterning of skin. Exp. Dermatol. 2016, 25, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Markell, K.W.; Renz, E.M.; White, C.E.; Albrecht, M.E.; Blackbourne, L.H.; Park, M.S.; Barillo, D.A.; Chung, K.K.; Kozar, R.A.; Minei, J.P.; et al. Abdominal complications after severe burns. J. Am. Coll. Surg. 2009, 208, 940–947; discussion 947–949. [Google Scholar] [CrossRef] [PubMed]
- Stanton, E.; McMullen, K.; Won, P.; Schneider, J.C.; Ryan, C.; Carrougher, G.; Kowalske, K.; Yenikomshian, H.A. Neuropathic Pain After Burn Injury: A Severe and Common Problem in Recovery. Ann. Surg. 2024, 279, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Xu, Y.; Ahmad, M.A.; Javed, R.; Hagiwara, H.; Tian, X. Exosomes as a Promising Therapeutic Strategy for Peripheral Nerve Injury. Curr. Neuropharmacol. 2021, 19, 2141–2151. [Google Scholar] [CrossRef]
- Baranoski, G.V.G.; Alencar, P.; Van Leeuwen, S.R.; Chen, T.F. Tanning-Elicited Variations in the Ultraviolet Absorption Spectra of the Cutaneous Tissues: Skin Photobiology and Photomedicine Implications. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2021, 2021, 4262–4267. [Google Scholar] [CrossRef]
- Hercher, D.; Nguyen, M.Q.; Dworak, H. Extracellular vesicles and their role in peripheral nerve regeneration. Exp. Neurol. 2022, 350, 113968. [Google Scholar] [CrossRef]
- Namini, M.S.; Daneshimehr, F.; Beheshtizadeh, N.; Mansouri, V.; Ai, J.; Jahromi, H.K.; Ebrahimi-Barough, S. Cell-free therapy based on extracellular vesicles: A promising therapeutic strategy for peripheral nerve injury. Stem Cell Res. Ther. 2023, 14, 254. [Google Scholar] [CrossRef]
- Granstein, R.D.; Wagner, J.A.; Stohl, L.L.; Ding, W. Calcitonin gene-related peptide: Key regulator of cutaneous immunity. Acta Physiol. 2015, 213, 586–594. [Google Scholar] [CrossRef]
- Legat, F.J.; Jaiani, L.T.; Wolf, P.; Wang, M.; Lang, R.; Abraham, T.; Solomon, A.R.; Armstrong, C.A.; Glass, J.D.; Ansel, J.C. The role of calcitonin gene-related peptide in cutaneous immunosuppression induced by repeated subinflammatory ultraviolet irradiation exposure. Exp. Dermatol. 2004, 13, 242–250. [Google Scholar] [CrossRef]
- Wallengren, J.; Sundler, F. Phototherapy reduces the number of epidermal and CGRP-positive dermal nerve fibres. Acta Derm. Venereol. 2004, 84, 111–115. [Google Scholar] [CrossRef]
- Richner, M.; Ulrichsen, M.; Elmegaard, S.L.; Dieu, R.; Pallesen, L.T.; Vaegter, C.B. Peripheral nerve injury modulates neurotrophin signaling in the peripheral and central nervous system. Mol. Neurobiol. 2014, 50, 945–970. [Google Scholar] [CrossRef]
- Chen, W.P.; Chang, Y.C.; Hsieh, S.T. Trophic interactions between sensory nerves and their targets. J. Biomed. Sci. 1999, 6, 79–85. [Google Scholar] [CrossRef]
- Fonseca, B.L.; dos Santos, B.C.; Martins, P.; Bonorino, C.; Corte, T.W.F.; da Silva, V.D.; Bauer, M.E. Neuroprotective effects of a new skin care formulation following ultraviolet exposure. Cell Prolif. 2012, 45, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, L.F.T.; Anami, E.H.T.; Zampar, E.F.; Tanita, M.T.; Cardoso, L.T.Q.; Grion, C.M.C. Epidemiology and outcome analysis of burn patients admitted to an Intensive Care Unit in a University Hospital. Burns 2016, 42, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Saeman, M.R.; Hodgman, E.I.; Burris, A.; Wolf, S.E.; Arnoldo, B.D.; Kowalske, K.J.; Phelan, H.A. Epidemiology and outcomes of pediatric burns over 35 years at Parkland Hospital. Burns 2016, 42, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Carrougher, G.J.; Bamer, A.M.; Mason, S.; Stewart, B.T.; Gibran, N.S. Defining numerical cut points for mild, moderate, and severe pain in adult burn survivors: A northwest regional burn model system investigation. Burns 2023, 49, 310–316. [Google Scholar] [CrossRef]
- Zoghoul Alsmadi, N.; Deister, C.; Agrawal, N.; Tran, L.; Zhukauskas, R.; Neubauer Fischer, D.; Mercer, D. Characterization of Nerve Damage After an Injury to the Adjacent Soft Tissue: A Pilot Animal Study. Tissue Eng. Part C Methods 2023, 29, 547–557. [Google Scholar] [CrossRef]
- Rowe, G.; Allahham, A.; Edgar, D.W.; Rurak, B.K.; Fear, M.W.; Wood, F.M.; Vallence, A.M. Functional Brain Changes Following Burn Injury: A Narrative Review. Neurorehabil. Neural Repair 2024, 38, 62–72. [Google Scholar] [CrossRef]
- Allahham, A.; Rowe, G.; Stevenson, A.; Fear, M.W.; Vallence, A.M.; Wood, F.M. The impact of burn injury on the central nervous system. Burn. Trauma 2024, 12, tkad037. [Google Scholar] [CrossRef]
- Li, R.; Li, D.H.; Zhang, H.Y.; Wang, J.; Li, X.K.; Xiao, J. Growth factors-based therapeutic strategies and their underlying signaling mechanisms for peripheral nerve regeneration. Acta Pharmacol. Sin. 2020, 41, 1289–1300. [Google Scholar] [CrossRef]
- Walter, M.A.; Kurouglu, R.; Caulfield, J.B.; Vasconez, L.O.; Thompson, J.A. Enhanced peripheral nerve regeneration by acidic fibroblast growth factor. Lymphokine Cytokine Res. 1993, 12, 135–141. [Google Scholar] [PubMed]
- Caissie, R.; Gingras, M.; Champigny, M.-F.; Berthod, F. In vivo enhancement of sensory perception recovery in a tissue-engineered skin enriched with laminin. Biomaterials 2006, 27, 2988–2993. [Google Scholar] [CrossRef]
- Sorrell, J.M.; Caplan, A.I. Topical delivery of mesenchymal stem cells and their function in wounds. Stem Cell Res. Ther. 2010, 1, 30. [Google Scholar] [CrossRef] [PubMed]
- Sanford, A.; Gamelli, R.L. Lightning and thermal injuries. Handb. Clin. Neurol. 2014, 120, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Hellerich, U.; Pollak, S. Airplane crash. Traumatologic findings in cases of extreme body disintegration. Am. J. Forensic Med. Pathol. 1995, 16, 320–324. [Google Scholar] [CrossRef]
- Gitkind, A.I.; Zhao, P.; Oh-Park, M.Y.; Fast, A. Median palmar cutaneous nerve injury in a volleyball player. Am. J. Phys. Med. Rehabil. 2009, 88, 272–274. [Google Scholar] [CrossRef]
- Rayan, G.M. Archery-related injuries of the hand, forearm, and elbow. South Med. J. 1992, 85, 961–964. [Google Scholar] [CrossRef]
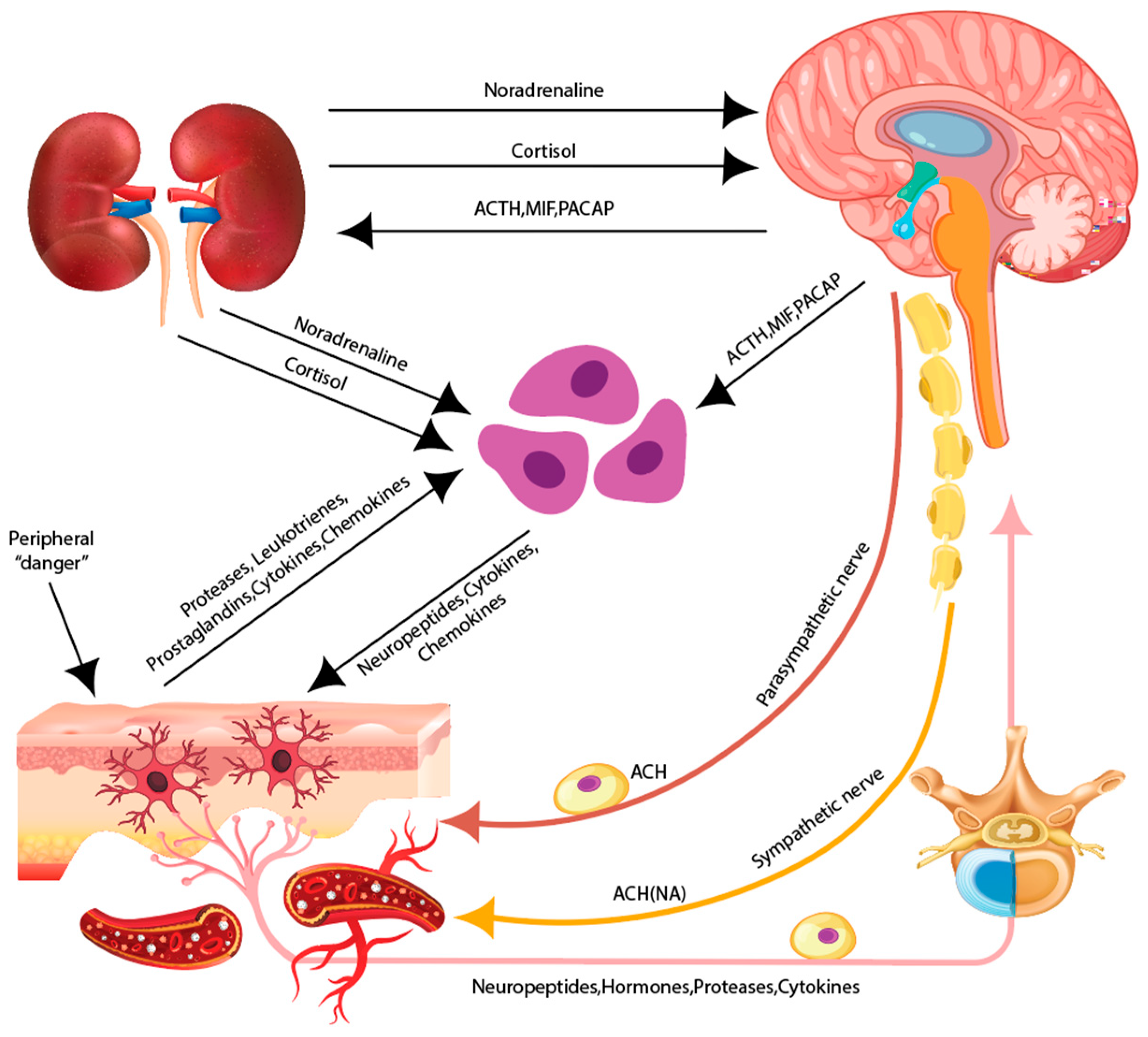
- Zmijewski, M.A.; Slominski, A.T. Neuroendocrinology of the skin: An overview and selective analysis. Dermatoendocrinol 2011, 3, 3–10. [Google Scholar] [CrossRef]
- Jin, R.; Luo, L.; Zheng, J. The Trinity of Skin: Skin Homeostasis as a Neuro-Endocrine-Immune Organ. Life 2022, 12, 725. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, 1–115. [Google Scholar] [CrossRef]
- Bove, D.; Lupoli, A.; Caccavale, S.; Piccolo, V.; Ruocco, E. Dermatological and immunological conditions due to nerve lesions. Funct. Neurol. 2013, 28, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Dimitrijević, M.; Stanojević, S. The intriguing mission of neuropeptide Y in the immune system. Amino Acids 2013, 45, 41–53. [Google Scholar] [CrossRef]
- Rodrigues, B.T.; Vangaveti, V.N.; Malabu, U.H. Prevalence and Risk Factors for Diabetic Lower Limb Amputation: A Clinic-Based Case Control Study. J. Diabetes Res. 2016, 2016, 5941957. [Google Scholar] [CrossRef] [PubMed]
- Jaul, E.; Barron, J.; Rosenzweig, J.P.; Menczel, J. An overview of co-morbidities and the development of pressure ulcers among older adults. BMC Geriatr. 2018, 18, 305. [Google Scholar] [CrossRef]
- Santesson, P.; Lins, P.E.; Kalani, M.; Adamson, U.; Lelic, I.; von Wendt, G.; Fagrell, B.; Jörneskog, G. Skin microvascular function in patients with type 1 diabetes: An observational study from the onset of diabetes. Diabetes Vasc. Dis. Res. 2017, 14, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Khaodhiar, L.; Dinh, T.; Schomacker, K.T.; Panasyuk, S.V.; Freeman, J.E.; Lew, R.; Vo, T.; Panasyuk, A.A.; Lima, C.; Giurini, J.M.; et al. The use of medical hyperspectral technology to evaluate microcirculatory changes in diabetic foot ulcers and to predict clinical outcomes. Diabetes Care 2007, 30, 903–910. [Google Scholar] [CrossRef]
- Vinik, A.I.; Erbas, T.; Stansberry, K.B.; Pittenger, G.L. Small fiber neuropathy and neurovascular disturbances in diabetes mellitus. Exp. Clin. Endocrinol. Diabetes 2001, 109 (Suppl. S2), S451–S473. [Google Scholar] [CrossRef]
- Richards, A.M.; Floyd, D.C.; Terenghi, G.; McGrouther, D.A. Cellular changes in denervated tissue during wound healing in a rat model. Br. J. Dermatol. 1999, 140, 1093–1099. [Google Scholar] [CrossRef]
- Ashrafi, M.; Baguneid, M.; Bayat, A. The Role of Neuromediators and Innervation in Cutaneous Wound Healing. Acta Derm. Venereol. 2016, 96, 587–594. [Google Scholar] [CrossRef]
- Ivanov, E.; Akhmetshina, M.; Erdiakov, A.; Gavrilova, S. Sympathetic System in Wound Healing: Multistage Control in Normal and Diabetic Skin. Int. J. Mol. Sci. 2023, 24, 2045. [Google Scholar] [CrossRef]
- Fukai, T.; Takeda, A.; Uchinuma, E. Wound healing in denervated rat skin. Wound Repair Regen. 2005, 13, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Stelnicki, E.J.; Doolabh, V.; Lee, S.; Levis, C.; Baumann, F.G.; Longaker, M.T.; Mackinnon, S. Nerve dependency in scarless fetal wound healing. Plast. Reconstr. Surg. 2000, 105, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Miao, X.-H.; Liu, T. More than Scratching the Surface: Recent Progress in Brain Mechanisms Underlying Itch and Scratch. Neurosci. Bull. 2020, 36, 85–88. [Google Scholar] [CrossRef]
- Cevikbas, F.; Lerner, E.A. Physiology and Pathophysiology of Itch. Physiol. Rev. 2020, 100, 945–982. [Google Scholar] [CrossRef]
- Sutaria, N.; Adawi, W.; Goldberg, R.; Roh, Y.S.; Choi, J.; Kwatra, S.G. Itch: Pathogenesis and treatment. J. Am. Acad. Dermatol. 2022, 86, 17–34. [Google Scholar] [CrossRef]
- Peng, L.; Yu, Q.; Zhang, J.; Mi, X.; Lin, W.; Qin, Y.; He, Y.; Guo, J.; Xiao, M.; Chen, M. Cupping for neurodermatitis: A protocol of systematic review and meta-analysis. Medicine 2020, 99, e22586. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Vander Does, A.; Mohsin, N.; Yosipovitch, G. Lichen Simplex Chronicus Itch: An Update. Acta Derm. Venereol. 2022, 102, adv00796. [Google Scholar] [CrossRef]
- Altunay, I.K. Neurodermatitis and emotions. Br. J. Dermatol. 2021, 185, E75–E76. [Google Scholar]
- Horch, K.W.; Tuckett, R.P.; Burgess, P.R. A key to the classification of cutaneous mechanoreceptors. J. Investig. Dermatol. 1977, 69, 75–82. [Google Scholar] [CrossRef]
- Shumway, N.K.; Cole, E.; Fernandez, K.H. Neurocutaneous disease: Neurocutaneous dysesthesias. J. Am. Acad. Dermatol. 2016, 74, 215–228. [Google Scholar] [CrossRef]
- Nedelec, B.; Hou, Q.; Sohbi, I.; Choinière, M.; Beauregard, G.; Dykes, R.W. Sensory perception and neuroanatomical structures in normal and grafted skin of burn survivors. Burns 2005, 31, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Dehran, M. Leprosy: A precipitating factor for complex regional pain syndrome. Minerva Anestesiol. 2010, 76, 758–760. [Google Scholar] [PubMed]
- Sehgal, V.N.; Sardana, K.; Dogra, S. Management of complications following leprosy: An evolving scenario. J. Dermatol. Treat. 2007, 18, 366–374. [Google Scholar] [CrossRef]
- Li, C.; Liu, S.Y.; Pi, W.; Zhang, P.X. Cortical plasticity and nerve regeneration after peripheral nerve injury. Neural Regen. Res. 2021, 16, 1518–1523. [Google Scholar] [CrossRef]
- Martínez-Marcos, A.; Sañudo, J.R. Cranial Nerves: Morphology and Clinical Relevance. Anat. Rec. 2019, 302, 555–557. [Google Scholar] [CrossRef]
- Johnson, R.W. Herpes zoster and postherpetic neuralgia. Expert Rev. Vaccines 2010, 9, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Castronovo, C.; Nikkels, A.F. Chronic herpes zoster duplex bilateralis. Acta Derm. Venereol. 2012, 92, 148–151. [Google Scholar] [CrossRef]
- Ruocco, V.; Sangiuliano, S.; Brunetti, G.; Ruocco, E. Beyond zoster: Sensory and immune changes in zoster-affected dermatomes: A review*. Acta Derm. Venereol. 2012, 92, 378–382. [Google Scholar] [CrossRef]
- Murthy, N.K.; Sharma, M.; Spinner, R.J. Primary peripheral nerve tumors associated with nerve-territory herpes zoster. Acta Neurochir. 2020, 162, 1147–1151. [Google Scholar] [CrossRef]
- Jabbour, S.A. Cutaneous manifestations of endocrine disorders: A guide for dermatologists. Am. J. Clin. Dermatol. 2003, 4, 315–331. [Google Scholar] [CrossRef]
- Abu Tahir, M.; Pramod, K.; Ansari, S.H.; Ali, J. Current remedies for vitiligo. Autoimmun. Rev. 2010, 9, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Al’Abadie, M.S.; Senior, H.J.; Bleehen, S.S.; Gawkrodger, D.J. Neuropeptide and neuronal marker studies in vitiligo. Br. J. Dermatol. 1994, 131, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Hristakieva, E.; Lazarova, R.; Lazarov, N.; Stanimirović, A.; Shani, J. Markers for vitiligo related neuropeptides in human skin nerve fibers. Acta Med. Croat. 2000, 54, 53–57. [Google Scholar]
- Yu, R.; Huang, Y.; Zhang, X.; Zhou, Y. Potential role of neurogenic inflammatory factors in the pathogenesis of vitiligo. J. Cutan. Med. Surg. 2012, 16, 230–244. [Google Scholar] [CrossRef]
- Lazarova, R.; Hristakieva, E.; Lazarov, N.; Shani, J. Vitiligo-related neuropeptides in nerve fibers of the skin. Arch. Physiol. Biochem. 2000, 108, 262–267. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Kou, K.; Nakamura, F.; Aihara, M.; Chen, H.; Seto, K.; Komori-Yamaguchi, J.; Kambara, T.; Nagashima, Y.; Goshima, Y.; Ikezawa, Z. Decreased expression of semaphorin-3A, a neurite-collapsing factor, is associated with itch in psoriatic skin. Acta Derm. Venereol. 2012, 92, 521–528. [Google Scholar] [CrossRef]
- Chen, S.-Q.; Chen, X.-Y.; Cui, Y.-Z.; Yan, B.-X.; Zhou, Y.; Wang, Z.-Y.; Xu, F.; Huang, Y.-Z.; Zheng, Y.-X.; Man, X.-Y. Cutaneous nerve fibers participate in the progression of psoriasis by linking epidermal keratinocytes and immunocytes. Cell. Mol. Life Sci. 2022, 79, 267. [Google Scholar] [CrossRef]
- Kyogoku, C.; Tsuchiya, N. A compass that points to lupus: Genetic studies on type I interferon pathway. Genes Immun. 2007, 8, 445–455. [Google Scholar] [CrossRef]
- Fong, S.Y.; Raja, J.; Wong, K.T.; Goh, K.J. Systemic lupus erythematosus may have an early effect on peripheral nerve function in patients without clinical or electrophysiological neuropathy: Comparison with age- and gender-matched controls. Rheumatol. Int. 2021, 41, 355–360. [Google Scholar] [CrossRef]
- Tekatas, A.; Tekatas, D.D.; Solmaz, V.; Karaca, T.; Pamuk, O.N. Small fiber neuropathy and related factors in patients with systemic lupus erythematosus; the results of cutaneous silent period and skin biopsy. Adv. Rheumatol. 2020, 60, 31. [Google Scholar] [CrossRef] [PubMed]
- Tavee, J.; Zhou, L. Small fiber neuropathy: A burning problem. Clevel. Clin. J. Med. 2009, 76, 297–305. [Google Scholar] [CrossRef]
- Wang, M.; Peng, Y.; Li, H.; Zhang, X. From monogenic lupus to TLR7/MyD88-targeted therapy. Innovation 2022, 3, 100299. [Google Scholar] [CrossRef]
- Maddison, B.; Parsons, A.; Sangueza, O.; Sheehan, D.J.; Yosipovitch, G. Retrospective study of intraepidermal nerve fiber distribution in biopsies of patients with nummular eczema. Am. J. Dermatopathol. 2011, 33, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Wilkin, J.; Dahl, M.; Detmar, M.; Drake, L.; Feinstein, A.; Odom, R.; Powell, F. Standard classification of rosacea: Report of the National Rosacea Society Expert Committee on the Classification and Staging of Rosacea. J. Am. Acad. Dermatol. 2002, 46, 584–587. [Google Scholar] [CrossRef]
- Gallo, R.L.; Granstein, R.D.; Kang, S.; Mannis, M.; Steinhoff, M.; Tan, J.; Thiboutot, D. Standard classification and pathophysiology of rosacea: The 2017 update by the National Rosacea Society Expert Committee. J. Am. Acad. Dermatol. 2018, 78, 148–155. [Google Scholar] [CrossRef]
- Tan, J.; Steinhoff, M.; Berg, M.; Del Rosso, J.; Layton, A.; Leyden, J.; Schauber, J.; Schaller, M.; Cribier, B.; Thiboutot, D.; et al. Shortcomings in rosacea diagnosis and classification. Br. J. Dermatol. 2017, 176, 197–199. [Google Scholar] [CrossRef]
- Powell, F.C. Clinical practice. Rosacea. N. Engl. J. Med. 2005, 352, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Schwab, V.D.; Sulk, M.; Seeliger, S.; Nowak, P.; Aubert, J.; Mess, C.; Rivier, M.; Carlavan, I.; Rossio, P.; Metze, D.; et al. Neurovascular and neuroimmune aspects in the pathophysiology of rosacea. J. Investig. Dermatol. Symp. Proc. 2011, 15, 53–62. [Google Scholar] [CrossRef]
- Muto, Y.; Wang, Z.; Vanderberghe, M.; Two, A.; Gallo, R.L.; Di Nardo, A. Mast cells are key mediators of cathelicidin-initiated skin inflammation in rosacea. J. Investig. Dermatol. 2014, 134, 2728–2736. [Google Scholar] [CrossRef]
- Roosterman, D.; Goerge, T.; Schneider, S.W.; Bunnett, N.W.; Steinhoff, M. Neuronal control of skin function: The skin as a neuroimmunoendocrine organ. Physiol. Rev. 2006, 86, 1309–1379. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.J.; Terenghi, G.; Hazari, A.; Wiberg, M. Nerve fibre and sensory end organ density in the epidermis and papillary dermis of the human hand. Br. J. Plast. Surg. 2005, 58, 774–779. [Google Scholar] [CrossRef]
- Vetrugno, R.; Liguori, R.; Cortelli, P.; Montagna, P. Sympathetic skin response: Basic mechanisms and clinical applications. Clin. Auton. Res. 2003, 13, 256–270. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, L.; Nabzdyk, C.; Andersen, N.D.; LoGerfo, F.W.; Veves, A. Inflammation and neuropeptides: The connection in diabetic wound healing. Expert Rev. Mol. Med. 2009, 11, e2. [Google Scholar] [CrossRef]
- Chéret, J.; Lebonvallet, N.; Buhé, V.; Carre, J.L.; Misery, L.; Le Gall-Ianotto, C. Influence of sensory neuropeptides on human cutaneous wound healing process. J. Dermatol. Sci. 2014, 74, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Hogg, P.J.; McLachlan, E.M. Blood vessels and nerves: Together or not? Lancet 2002, 360, 1714. [Google Scholar] [CrossRef]
- Fujiwara, T.; Kubo, T.; Kanazawa, S.; Shingaki, K.; Taniguchi, M.; Matsuzaki, S.; Gurtner, G.C.; Tohyama, M.; Hosokawa, K. Direct contact of fibroblasts with neuronal processes promotes differentiation to myofibroblasts and induces contraction of collagen matrix in vitro. Wound Repair Regen. 2013, 21, 588–594. [Google Scholar] [CrossRef]
- Romana-Souza, B.; Porto, L.C.; Monte-Alto-Costa, A. Cutaneous wound healing of chronically stressed mice is improved through catecholamines blockade. Exp. Dermatol. 2010, 19, 821–829. [Google Scholar] [CrossRef]
- Lotti, T.; Bianchi, B.; Panconesi, E. Neuropeptides and skin disorders. The new frontiers of neuro-endocrine-cutaneous immunology. Int. J. Dermatol. 1999, 38, 673–675. [Google Scholar] [CrossRef]
- Bevan, S.; Szolcsányi, J. Sensory neuron-specific actions of capsaicin: Mechanisms and applications. Trends Pharmacol. Sci. 1990, 11, 330–333. [Google Scholar] [CrossRef]
- Dray, A. Mechanism of action of capsaicin-like molecules on sensory neurons. Life Sci. 1992, 51, 1759–1765. [Google Scholar] [CrossRef] [PubMed]
- Baluk, P. Neurogenic inflammation in skin and airways. J. Investig. Dermatol. Symp. Proc. 1997, 2, 76–81. [Google Scholar] [CrossRef]
- Gürgen, S.G.; Sayın, O.; Cetin, F.; Tuç Yücel, A. Transcutaneous electrical nerve stimulation (TENS) accelerates cutaneous wound healing and inhibits pro-inflammatory cytokines. Inflammation 2014, 37, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Chéret, J.; Lebonvallet, N.; Carré, J.L.; Misery, L.; Le Gall-Ianotto, C. Role of neuropeptides, neurotrophins, and neurohormones in skin wound healing. Wound Repair Regen. 2013, 21, 772–788. [Google Scholar] [CrossRef]
- Zhao, B.; Zhang, Y.; Han, S.; Zhang, W.; Zhou, Q.; Guan, H.; Liu, J.; Shi, J.; Su, L.; Hu, D. Exosomes derived from human amniotic epithelial cells accelerate wound healing and inhibit scar formation. J. Mol. Histol. 2017, 48, 121–132. [Google Scholar] [CrossRef]
- Liu, J.; Yan, Z.; Yang, F.; Huang, Y.; Yu, Y.; Zhou, L.; Sun, Z.; Cui, D.; Yan, Y. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Accelerate Cutaneous Wound Healing by Enhancing Angiogenesis through Delivering Angiopoietin-2. Stem Cell Rev. Rep. 2021, 17, 305–317. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, X.; Hao, H.; Xu, H.; Shu, J.; Hou, Q.; Wang, M. Exosomes Derived From Umbilical Cord Mesenchymal Stem Cells Treat Cutaneous Nerve Damage and Promote Wound Healing. Front. Cell. Neurosci. 2022, 16, 913009. [Google Scholar] [CrossRef] [PubMed]
- Fawzy El-Sayed, K.M.; Dörfer, C.E. Gingival Mesenchymal Stem/Progenitor Cells: A Unique Tissue Engineering Gem. Stem Cells Int. 2016, 2016, 7154327. [Google Scholar] [CrossRef]
- Shi, Q.; Qian, Z.; Liu, D.; Sun, J.; Wang, X.; Liu, H.; Xu, J.; Guo, X. GMSC-Derived Exosomes Combined with a Chitosan/Silk Hydrogel Sponge Accelerates Wound Healing in a Diabetic Rat Skin Defect Model. Front. Physiol. 2017, 8, 904. [Google Scholar] [CrossRef]
- Zhang, H.; Shao, Z.; Zhu, Y.; Shi, L.; Li, Z.; Hou, R.; Zhang, C.; Yao, D. Toll-Like Receptor 4 (TLR4) Expression Affects Schwann Cell Behavior in vitro. Sci. Rep. 2018, 8, 11179. [Google Scholar] [CrossRef]
- Jiang, L.; Jones, S.; Jia, X. Stem Cell Transplantation for Peripheral Nerve Regeneration: Current Options and Opportunities. Int. J. Mol. Sci. 2017, 18, 94. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Verrilli, M.A.; Picou, F.; Court, F.A. Schwann cell-derived exosomes enhance axonal regeneration in the peripheral nervous system. Glia 2013, 61, 1795–1806. [Google Scholar] [CrossRef]
- Lopez-Verrilli, M.A.; Caviedes, A.; Cabrera, A.; Sandoval, S.; Wyneken, U.; Khoury, M. Mesenchymal stem cell-derived exosomes from different sources selectively promote neuritic outgrowth. Neuroscience 2016, 320, 129–139. [Google Scholar] [CrossRef]
- Wei, J.-j.; Chen, Y.-f.; Xue, C.-l.; Ma, B.-t.; Shen, Y.-m.; Guan, J.; Bao, X.-j.; Wu, H.; Han, Q.; Wang, R.-z.; et al. Protection of Nerve Injury with Exosome Extracted from Mesenchymal Stem Cell. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2016, 38, 33–36. [Google Scholar] [CrossRef]
- Mao, Q.; Nguyen, P.D.; Shanti, R.M.; Shi, S.; Shakoori, P.; Zhang, Q.; Le, A.D. Gingiva-Derived Mesenchymal Stem Cell-Extracellular Vesicles Activate Schwann Cell Repair Phenotype and Promote Nerve Regeneration. Tissue Eng. Part A 2019, 25, 887–900. [Google Scholar] [CrossRef]
- Dong, R.; Liu, Y.; Yang, Y.; Wang, H.; Xu, Y.; Zhang, Z. MSC-Derived Exosomes-Based Therapy for Peripheral Nerve Injury: A Novel Therapeutic Strategy. Biomed. Res. Int. 2019, 2019, 6458237. [Google Scholar] [CrossRef] [PubMed]
- Aswad, H.; Jalabert, A.; Rome, S. Depleting extracellular vesicles from fetal bovine serum alters proliferation and differentiation of skeletal muscle cells in vitro. BMC Biotechnol. 2016, 16, 32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiang, Y.; Huang, Q.; Wu, Z.; Pu, H.; Xu, Z.; Li, B.; Lu, X.; Yang, X.; Qin, J.; et al. Exosomes derived from adipose-derived stem cells overexpressing glyoxalase-1 protect endothelial cells and enhance angiogenesis in type 2 diabetic mice with limb ischemia. Stem Cell Res. Ther. 2021, 12, 403. [Google Scholar] [CrossRef]
- Didiot, M.C.; Hall, L.M.; Coles, A.H.; Haraszti, R.A.; Godinho, B.M.; Chase, K.; Sapp, E.; Ly, S.; Alterman, J.F.; Hassler, M.R.; et al. Exosome-mediated Delivery of Hydrophobically Modified siRNA for Huntingtin mRNA Silencing. Mol. Ther. 2016, 24, 1836–1847. [Google Scholar] [CrossRef]
- Moattari, M.; Kouchesfehani, H.M.; Kaka, G.; Sadraie, S.H.; Naghdi, M. Evaluation of nerve growth factor (NGF) treated mesenchymal stem cells for recovery in neurotmesis model of peripheral nerve injury. J. Cranio-Maxillofac. Surg. 2018, 46, 898–904. [Google Scholar] [CrossRef]
- Bucan, V.; Vaslaitis, D.; Peck, C.T.; Strauß, S.; Vogt, P.M.; Radtke, C. Effect of Exosomes from Rat Adipose-Derived Mesenchymal Stem Cells on Neurite Outgrowth and Sciatic Nerve Regeneration After Crush Injury. Mol. Neurobiol. 2019, 56, 1812–1824. [Google Scholar] [CrossRef]
- Sharma, P.; Mesci, P.; Carromeu, C.; McClatchy, D.R.; Schiapparelli, L.; Yates, J.R., 3rd; Muotri, A.R.; Cline, H.T. Exosomes regulate neurogenesis and circuit assembly. Proc. Natl. Acad. Sci. USA 2019, 116, 16086–16094. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, R.; Tilley, D.M.; Vogel, L.; Benyamin, R. The role of glia and the immune system in the development and maintenance of neuropathic pain. Pain Pract. 2010, 10, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-M.; Gibbs, K.M.; Davila, J.; Campbell, N.; Sung, S.; Todorova, T.I.; Otsuka, S.; Sabaawy, H.E.; Hart, R.P.; Schachner, M. MicroRNA miR-133b is essential for functional recovery after spinal cord injury in adult zebrafish. Eur. J. Neurosci. 2011, 33, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Shiue, S.-J.; Rau, R.-H.; Shiue, H.-S.; Hung, Y.-W.; Li, Z.-X.; Yang, K.D.; Cheng, J.-K. Mesenchymal stem cell exosomes as a cell-free therapy for nerve injury-induced pain in rats. Pain 2019, 160, 210–223. [Google Scholar] [CrossRef]
- Schratt, G.M.; Nigh, E.A.; Chen, W.G.; Hu, L.; Greenberg, M.E. BDNF regulates the translation of a select group of mRNAs by a mammalian target of rapamycin-phosphatidylinositol 3-kinase-dependent pathway during neuronal development. J. Neurosci. 2004, 24, 7366–7377. [Google Scholar] [CrossRef]
- Gao, Y.-Y.; Hong, X.-Y.; Wang, H.-J. Role of Nectin-1/c-Src Signaling in the Analgesic Effect of GDNF on a Rat Model of Chronic Constrictive Injury. J. Mol. Neurosci. 2016, 60, 258–266. [Google Scholar] [CrossRef]
- Yu, X.; Fu, X.; Yang, J.; Chen, L.; Leng, F.; Yang, Z.; Yu, C. Glucose/ROS cascade-responsive ceria nanozymes for diabetic wound healing. Mater. Today Bio 2022, 15, 100308. [Google Scholar] [CrossRef]
- Liang, Y.; Duan, L.; Lu, J.; Xia, J. Engineering exosomes for targeted drug delivery. Theranostics 2021, 11, 3183–3195. [Google Scholar] [CrossRef]
- Ha, D.; Yang, N.; Nadithe, V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: Current perspectives and future challenges. Acta Pharm. Sin. B 2016, 6, 287–296. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: From biogenesis to uptake and intracellular signalling. Cell Commun. Signal 2021, 19, 47. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Wang, X.; Zhao, H.; Deng, Y.; Zeng, W.; Yang, K.; Chen, H.; Yan, Q.; Li, C.; Wu, J.; et al. The effectiveness of cell-derived exosome therapy for diabetic wound: A systematic review and meta-analysis. Ageing Res. Rev. 2023, 85, 101858. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Li, C.; Szalad, A.; Wang, L.; Pan, W.; Zhang, R.; Chopp, M.; Zhang, Z.G.; Liu, X.S. Mesenchymal stromal cell-derived exosomes ameliorate peripheral neuropathy in a mouse model of diabetes. Diabetologia 2020, 63, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhao, X.; Zhong, Y.; Yang, P.; Gao, P.; Wu, X.; Wang, X.; An, W. Exosomal ncRNAs: The pivotal players in diabetic wound healing. Front. Immunol. 2022, 13, 1005307. [Google Scholar] [CrossRef]
- Lou, R.; Chen, J.; Zhou, F.; Wang, C.; Leung, C.H.; Lin, L. Exosome-cargoed microRNAs: Potential therapeutic molecules for diabetic wound healing. Drug Discov. Today 2022, 27, 103323. [Google Scholar] [CrossRef]
- Birklein, F.; Schlereth, T. Complex regional pain syndrome-significant progress in understanding. Pain 2015, 156 (Suppl. S1), S94–S103. [Google Scholar] [CrossRef]
- Bruehl, S. Complex regional pain syndrome. Br. Med. J. 2015, 351, h2730. [Google Scholar] [CrossRef]
- Krämer, H.H.; Eberle, T.; Üçeyler, N.; Wagner, I.; Klonschinsky, T.; Müller, L.P.; Sommer, C.; Birklein, F. TNF-α in CRPS and ‘normal’ trauma–significant differences between tissue and serum. Pain 2011, 152, 285–290. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Wilczynska, A.; Bushell, M. The complexity of miRNA-mediated repression. Cell Death Differ. 2015, 22, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Zhang, L.; Liang, C.; Liu, B.; Pan, X.; Wang, Y.; Zhang, Y.; Zhang, Y.; Xie, W.; Yan, B.; et al. Stem Cell-Derived Exosomes Prevent Aging-Induced Cardiac Dysfunction through a Novel Exosome/lncRNA MALAT1/NF-κB/TNF-α Signaling Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 9739258. [Google Scholar] [CrossRef] [PubMed]
- El-Derany, M.O.; Noureldein, M.H. Bone marrow mesenchymal stem cells and their derived exosomes resolve doxorubicin-induced chemobrain: Critical role of their miRNA cargo. Stem Cell Res. Ther. 2021, 12, 322. [Google Scholar] [CrossRef]
- Mack, M.R.; Kim, B.S. The Itch-Scratch Cycle: A Neuroimmune Perspective. Trends Immunol. 2018, 39, 980–991. [Google Scholar] [CrossRef]
- Trier, A.M.; Ver Heul, A.M.; Fredman, A.; Le, V.; Wang, Z.; Auyeung, K.; Meixiong, J.; Lovato, P.; Holtzman, M.J.; Wang, F.; et al. IL-33 potentiates histaminergic itch. J. Allergy Clin. Immunol. 2024, 153, 852–859.e853. [Google Scholar] [CrossRef]
- Cevikbas, F.; Wang, X.; Akiyama, T.; Kempkes, C.; Savinko, T.; Antal, A.; Kukova, G.; Buhl, T.; Ikoma, A.; Buddenkotte, J.; et al. A sensory neuron-expressed IL-31 receptor mediates T helper cell-dependent itch: Involvement of TRPV1 and TRPA1. J. Allergy Clin. Immunol. 2014, 133, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.A.; Kwon, J.; Kim, H.J.; Woo, S.Y. Mesenchymal stem cell exosomes differentially regulate gene expression of mast cells. Biochem. Biophys. Res. Commun. 2024, 696, 149517. [Google Scholar] [CrossRef]
- Xie, Y.; Yu, L.; Cheng, Z.; Peng, Y.; Cao, Z.; Chen, B.; Duan, Y.; Wang, Y. SHED-derived exosomes promote LPS-induced wound healing with less itching by stimulating macrophage autophagy. J. Nanobiotechnol. 2022, 20, 239. [Google Scholar] [CrossRef]
- Kim, C.; Kim, Y.; Lim, J.Y.; Kim, M.; Zheng, H.; Kim, M.; Hwang, S.W. Pamoic acid-induced peripheral GPR35 activation improves pruritus and dermatitis. Br. J. Pharmacol. 2023, 180, 3059–3070. [Google Scholar] [CrossRef]
- Nattkemper, L.A.; Lipman, Z.M.; Ingrasci, G.; Maldonado, C.; Garces, J.C.; Loayza, E.; Yosipovitch, G. Neuroimmune Mediators of Pruritus in Hispanic Scalp Psoriatic Itch. Acta Derm. Venereol. 2023, 103, adv4463. [Google Scholar] [CrossRef]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Rohde, E.; Pachler, K.; Gimona, M. Manufacturing and characterization of extracellular vesicles from umbilical cord-derived mesenchymal stromal cells for clinical testing. Cytotherapy 2019, 21, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.H.; Pan, N.F.; Lin, S.S.; Qiu, Z.Y.; Liang, P.; Wang, J.; Zhang, Z.; Pan, Y.C. Exosomes from mmu_circ_0001052-modified adipose-derived stem cells promote angiogenesis of DFU via miR-106a-5p and FGF4/p38MAPK pathway. Stem Cell Res. Ther. 2022, 13, 336. [Google Scholar] [CrossRef]
- Jian, X.; Han, J.; Liu, X.; Deng, Y.; Gao, S.; Xiao, S.; Zhang, Y.; Jian, S.; Huang, Z.; Hou, Y.; et al. Exosome-carried miR-1248 from adipose-derived stem cells improves angiogenesis in diabetes-associated wounds. Int. J. Biol. Macromol. 2025, 297, 139822. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, Z.; Li, S.; Xie, X.; Qin, L.; Zhang, Q.; Yang, Y.; Wang, T.; Zhang, Y. Exosomes: Compositions, biogenesis, and mechanisms in diabetic wound healing. J. Nanobiotechnol. 2024, 22, 398. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, H.; Xue, B.; Liu, L.; Tang, Y.; Wang, Z.; Yao, K. MSC-derived exosomal circMYO9B accelerates diabetic wound healing by promoting angiogenesis through the hnRNPU/CBL/KDM1A/VEGFA axis. Commun. Biol. 2024, 7, 1700. [Google Scholar] [CrossRef] [PubMed]
- Paicius, R.; White, Z.S.; Smith, C.; Lightner, A.L.; Ransom, J.T.; Lee, D.W.; Speare, S. Safety and Efficacy of Intravenous ExoFlo in the Treatment of Complex Regional Pain Syndrome. Pain Physician 2023, 26, E851–E857. [Google Scholar] [CrossRef]
- Chen, L.Y.; Kao, T.W.; Chen, C.C.; Niaz, N.; Lee, H.L.; Chen, Y.H.; Kuo, C.C.; Shen, Y.A. Frontier Review of the Molecular Mechanisms and Current Approaches of Stem Cell-Derived Exosomes. Cells 2023, 12, 2018. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, Y.; Luo, R.; Zhao, L.; Chen, D.; Cao, L.; Jin, Z.; Li, K.; Wang, M. Exosomes: A Promising Cell-Free Therapeutic Tool for Treating Cutaneous Nerve Injuries and Promoting Wound Healing. Int. J. Mol. Sci. 2025, 26, 5323. https://doi.org/10.3390/ijms26115323
Mu Y, Luo R, Zhao L, Chen D, Cao L, Jin Z, Li K, Wang M. Exosomes: A Promising Cell-Free Therapeutic Tool for Treating Cutaneous Nerve Injuries and Promoting Wound Healing. International Journal of Molecular Sciences. 2025; 26(11):5323. https://doi.org/10.3390/ijms26115323
Chicago/Turabian StyleMu, Yujie, Ruting Luo, Le Zhao, Danting Chen, Lixin Cao, Zhenkai Jin, Kun Li, and Min Wang. 2025. "Exosomes: A Promising Cell-Free Therapeutic Tool for Treating Cutaneous Nerve Injuries and Promoting Wound Healing" International Journal of Molecular Sciences 26, no. 11: 5323. https://doi.org/10.3390/ijms26115323
APA StyleMu, Y., Luo, R., Zhao, L., Chen, D., Cao, L., Jin, Z., Li, K., & Wang, M. (2025). Exosomes: A Promising Cell-Free Therapeutic Tool for Treating Cutaneous Nerve Injuries and Promoting Wound Healing. International Journal of Molecular Sciences, 26(11), 5323. https://doi.org/10.3390/ijms26115323