
Targeting Lineage-Specific Functions of NR4A1 for Cancer Immunotherapy
 , ,
, ,  , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Cell Types
2.1. T Cells
2.1.1. T Cell Function in the TME and Its Regulation
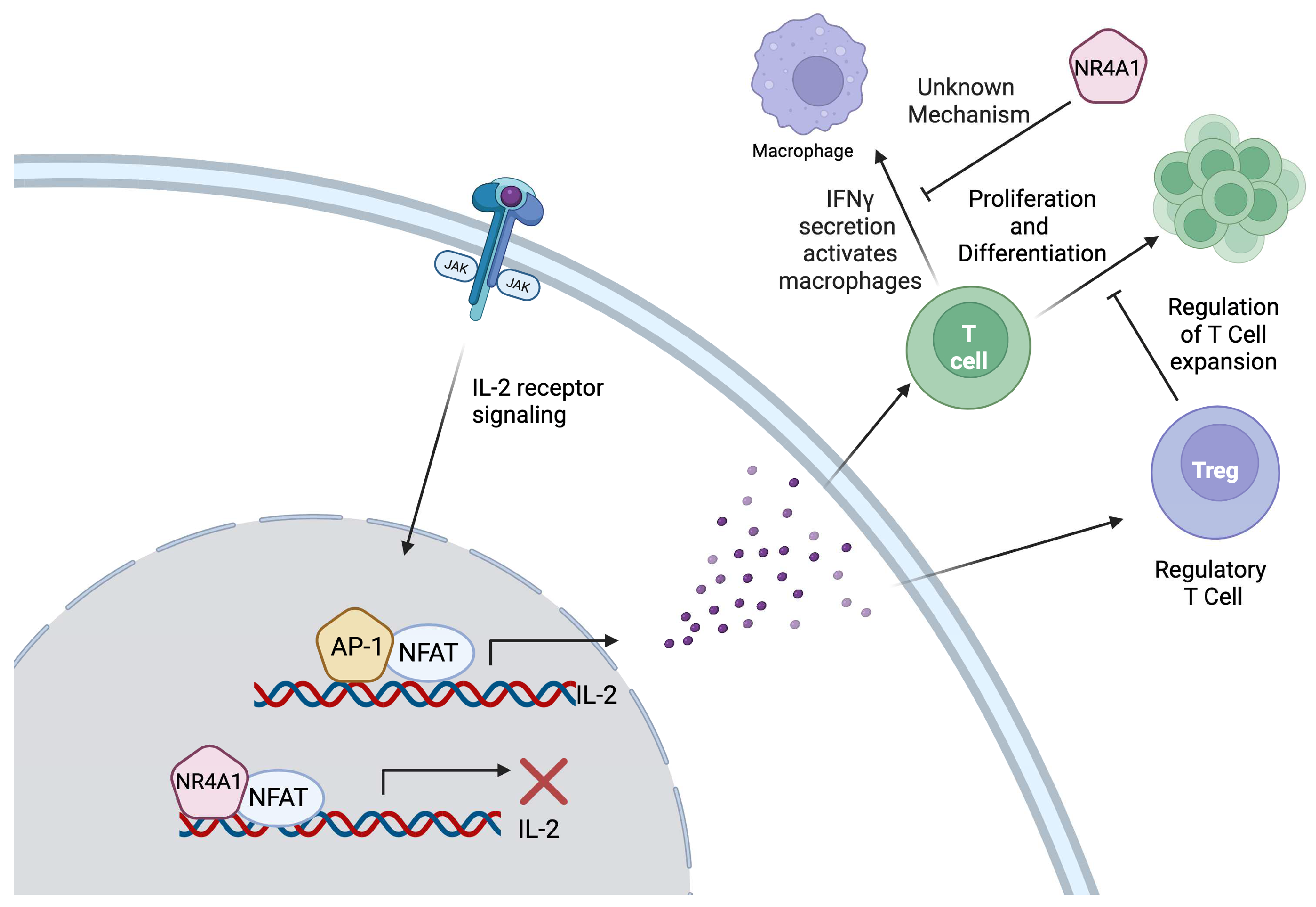
2.1.2. Role of NR4A1 in T Cell Biology
2.2. Monocytes
2.2.1. Monocyte Types and Functions in TME
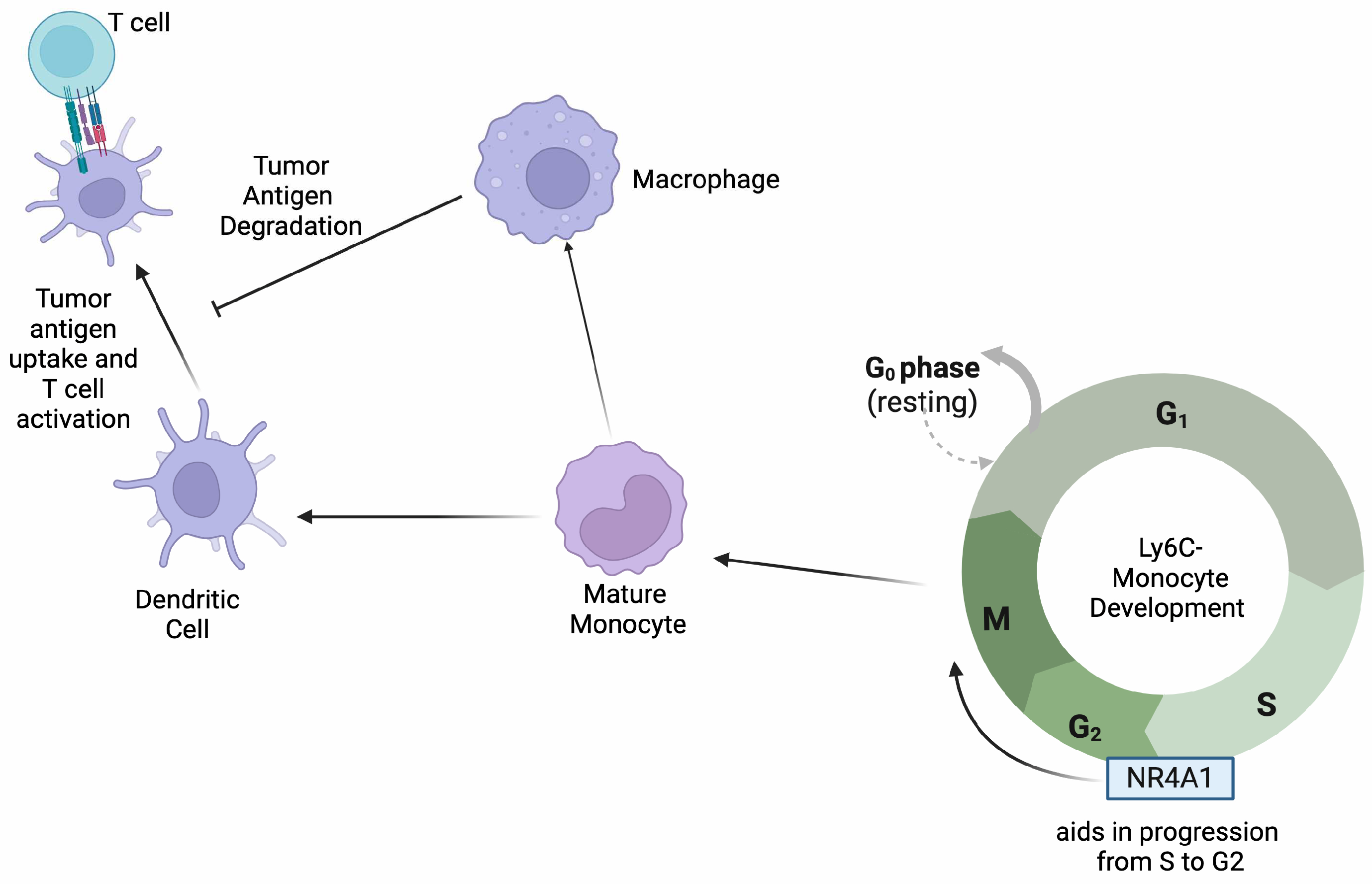
2.2.2. Role of NR4A1 in Monocytes
2.3. Natural Killer Cells
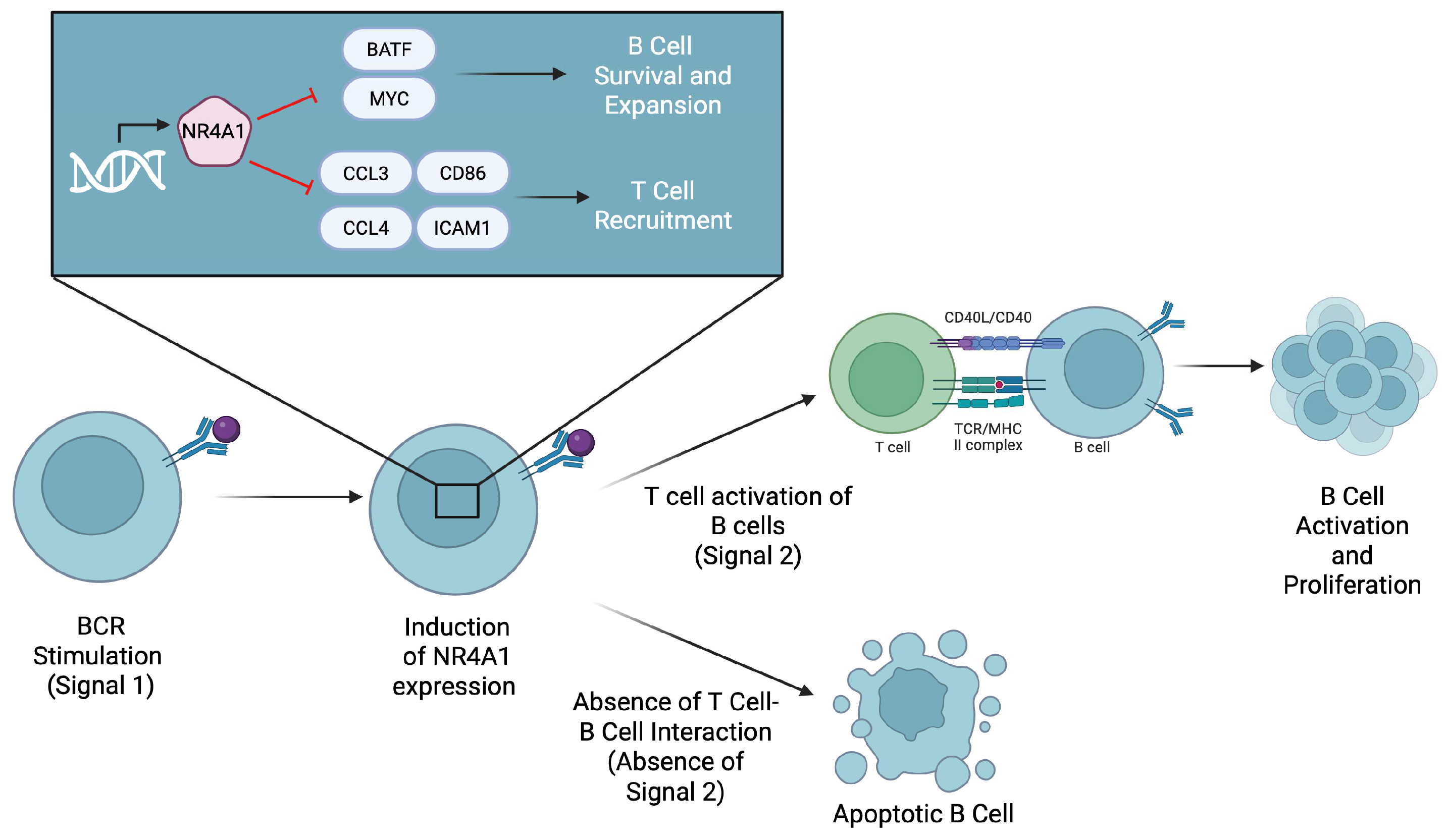
2.4. B Cells
2.5. Macrophages and Neutrophils
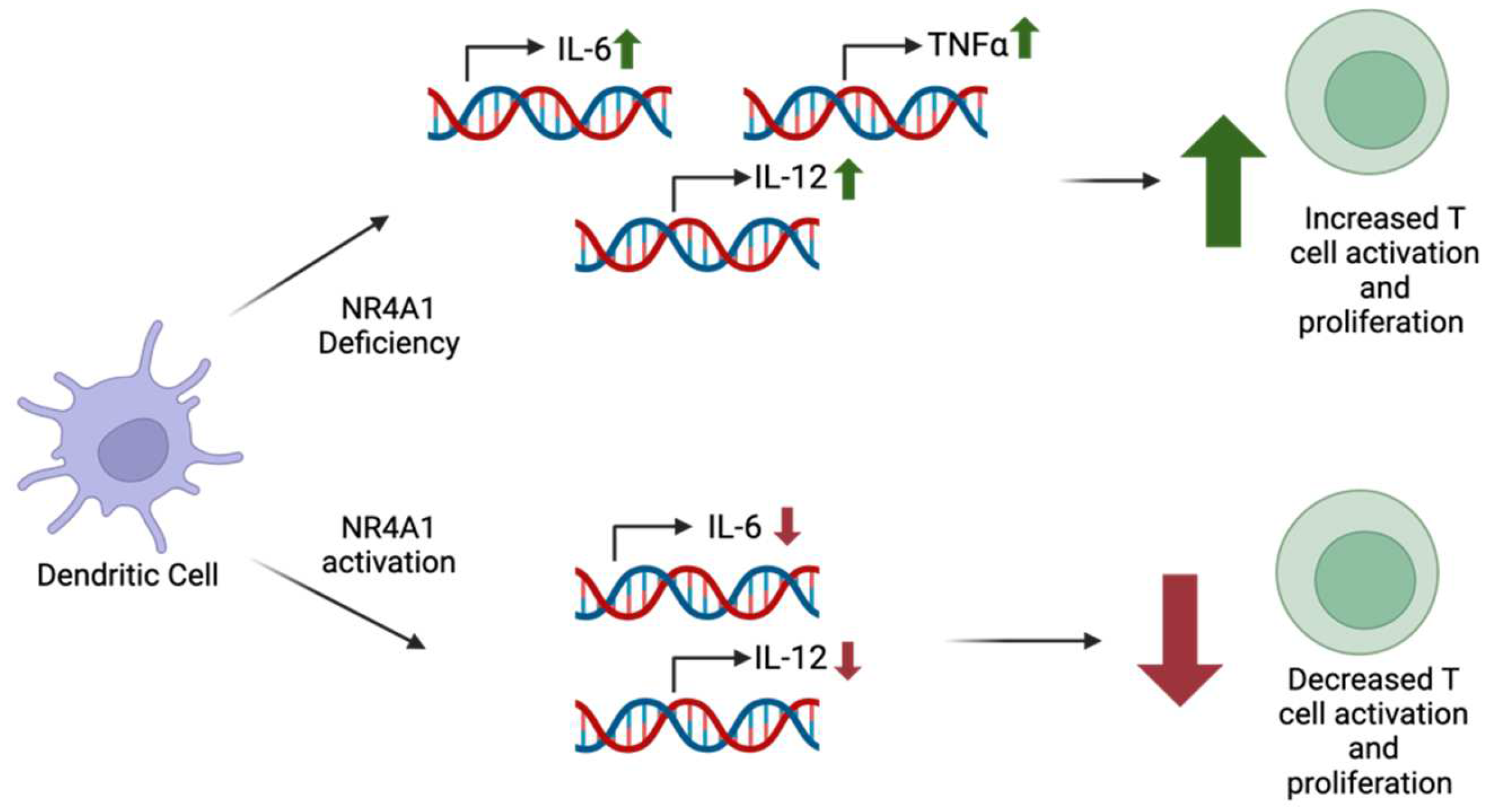
2.6. Dendritic Cells
2.7. Potential Therapeutic Interventions
2.7.1. Small-Molecule Inhibitors of NR4A1
2.7.2. Direct NR4A1 Degradation via PROTAC Therapy
2.7.3. NR4A1 Agonists
3. Conclusions, Perspectives, and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TME | Tumor microenvironment |
| NR4A1 | Nuclear receptor 4A1 |
| COAD | Colon adenocarcinoma |
| PD-1 | Programmed death-1 |
| PD-L1 | Programmed death-ligand 1 |
| IFN-γ | Interferon gamma |
| Tregs | Regulatory T cells |
| IL-2 | Interleukin-2 |
| AP-1 | Activator protein-1 |
| NF-κB | Nuclear factor kappa-B |
| NFAT | Nuclear factor of activated T cell |
| IL-2Rα | IL-2 receptor alpha |
| CCR2 | C-C chemokine receptor type 2 |
| CX3CR1 | C-X3-C motif chemokine receptor 1 |
| C/EBP-β | CCAAT enhancer binding protein beta |
| KLF | Krüppel-like factors |
| NK | Natural killer |
| PROTAC | Proteolysis targeting chimera |
| TNF | Tumor necrosis factor |
| TRAIL | Tumor necrosis factor-related apoptosis-inducing ligand |
| PUMA | p53 upregulated modulator of apoptosis |
| CCL3 | C-C motif chemokine ligand |
| ICAM1 | Intracellular adhesion molecule 1 |
| DCs | Dendritic cells |
| DIM-C-pPhOH | 1,1-bis(3′-indolyl)-1-(p-hydroxyphenyl) methane |
| POI | Protein of interest |
References
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef]
- Mayer, S.; Milo, T.; Isaacson, A.; Halperin, C.; Miyara, S.; Stein, Y.; Lior, C.; Pevsner-Fischer, M.; Tzahor, E.; Mayo, A.; et al. The tumor microenvironment shows a hierarchy of cell-cell interactions dominated by fibroblasts. Nat. Commun. 2023, 14, 5810. [Google Scholar] [CrossRef]
- Lu, C.; Liu, Y.; Ali, N.M.; Zhang, B.; Cui, X. The role of innate immune cells in the tumor microenvironment and research progress in anti-tumor therapy. Front. Immunol. 2023, 13, 1039260. [Google Scholar] [CrossRef]
- Sadeghi, M.; Dehnavi, S.; Sharifat, M.; Amiri, A.M.; Khodadadi, A. Innate immune cells: Key players of orchestra in modulating tumor microenvironment (TME). Heliyon 2024, 10, e27480. [Google Scholar] [CrossRef]
- Hinshaw, D.C.; Shevde, L.A. The tumor microenvironment innately modulates cancer progression. Cancer Res. 2019, 79, 4557–4566. [Google Scholar] [CrossRef]
- Inaba, K.; Metlay, J.P.; Crowley, M.T.; Steinman, R.M. Dendritic cells pulsed with protein antigens in vitro can prime antigen-specific, MHC-restricted T cells in situ. J. Exp. Med. 1990, 172, 631–640. [Google Scholar] [CrossRef]
- Dubois, B.; Bridon, J.-M.; Fayette, J.; Barthélémy, C.; Banchereau, J.; Caux, C.; Brière, F. Dendritic cells directly modulate B cell growth and differentiation. J. Leukoc. Biol. 1999, 66, 224–230. [Google Scholar] [CrossRef]
- Deng, S.; Chen, B.; Huo, J.; Liu, X. Therapeutic potential of NR4A1 in cancer: Focus on metabolism. Front. Oncol. 2022, 12, 972984. [Google Scholar] [CrossRef]
- Tel-Karthaus, N.; Kers-Rebel, E.D.; Looman, M.W.; Ichinose, H.; de Vries, C.J.; Ansems, M. Nuclear receptor nur77 deficiency alters dendritic cell function. Front. Immunol. 2018, 9, 1797. [Google Scholar] [CrossRef]
- Hanna, R.N.; Carlin, L.M.; Hubbeling, H.G.; Nackiewicz, D.; Green, A.M.; Punt, J.A.; Geissmann, F.; Hedrick, C.C. The transcription factor NR4A1 (Nur77) controls bone marrow differentiation and the survival of Ly6C-monocytes. Nat. Immunol. 2011, 12, 778–785. [Google Scholar] [CrossRef]
- Hanna, R.N.; Cekic, C.; Sag, D.; Tacke, R.; Thomas, G.D.; Nowyhed, H.; Herrley, E.; Rasquinha, N.; McArdle, S.; Wu, R.; et al. Patrolling monocytes control tumor metastasis to the lung. Science 2015, 350, 985–990. [Google Scholar] [CrossRef]
- Thomas, G.D.; Hanna, R.N.; Vasudevan, N.T.; Hamers, A.A.; Romanoski, C.E.; McArdle, S.; Ross, K.D.; Blatchley, A.; Yoakum, D.; Hamilton, B.A.; et al. Deleting an Nr4a1 Super-Enhancer Subdomain Ablates Ly6Clow Monocytes while Preserving Macrophage Gene Function. Immunity 2016, 45, 975–987. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Z.; Li, M.; Chen, Z.; Tang, W.; Cheng, X. NR4A1 as a potential therapeutic target in colon adenocarcinoma: A computational analysis of immune infiltration and drug response. Front. Genet. 2023, 14, 1181320. [Google Scholar] [CrossRef]
- Allen, L.-A.H. NR4A proteins and neutrophil lifespan. Blood 2017, 130, 958–959. [Google Scholar] [CrossRef]
- Tacke, R.; Hilgendorf, I.; Garner, H.; Waterborg, C.; Park, K.; Nowyhed, H.; Hanna, R.N.; Wu, R.; Swirski, F.K.; Geissmann, F.; et al. The transcription factor NR4A1 is essential for the development of a novel macrophage subset in the thymus. Sci. Rep. 2015, 5, 10055. [Google Scholar] [CrossRef]
- Tang, F.; Li, J.; Qi, L.; Liu, D.; Bo, Y.; Qin, S.; Miao, Y.; Yu, K.; Hou, W.; Li, J.; et al. A pan-cancer single-cell panorama of human natural killer cells. Cell 2023, 186, 4235–4251. [Google Scholar] [CrossRef]
- Moreno-Nieves, U.Y.; Tay, J.K.; Saumyaa, S.; Horowitz, N.B.; Shin, J.H.; Mohammad, I.A.; Luca, B.; Mundy, D.C.; Gulati, G.S.; Bedi, N.; et al. Landscape of innate lymphoid cells in human head and neck cancer reveals divergent NK cell states in the tumor microenvironment. Proc. Natl. Acad. Sci. USA 2021, 118, e2101169118. [Google Scholar] [CrossRef]
- Wang, R.; Bao, W.; Pal, M.; Liu, Y.; Yazdanbakhsh, K.; Zhong, H. Intermediate monocytes induced by IFN-γ inhibit cancer metastasis by promoting NK cell activation through FOXO1 and interleukin-27. J. Immunother. Cancer 2022, 10, e003539. [Google Scholar] [CrossRef]
- Yu, W.; He, J.; Wang, F.; He, Q.; Shi, Y.; Tao, X.; Sun, B. NR4A1 mediates NK-cell dysfunction in hepatocellular carcinoma via the IFN-γ/p-STAT1/IRF1 pathway. Immunology 2023, 169, 69–82. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Lu, H.; Li, J.; Yan, X.; Xiao, M.; Hao, J.; Alekseev, A.; Khong, H.; Chen, T.; et al. Genome-wide analysis identifies NR4A1 as a key mediator of T cell dysfunction. Nature 2019, 567, 525–529. [Google Scholar] [CrossRef]
- Chen, J.; López-Moyado, I.F.; Seo, H.; Lio, C.-W.J.; Hempleman, L.J.; Sekiya, T.; Yoshimura, A.; Scott-Browne, J.P.; Rao, A. NR4A transcription factors limit CAR T cell function in solid tumours. Nature 2019, 567, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Mohankumar, K.; Wright, G.; Kumaravel, S.; Shrestha, R.; Zhang, L.; Abdelrahim, M.; Chapkin, R.S.; Safe, S. Bis-indole-derived NR4A1 antagonists inhibit colon tumor and splenic growth and T-cell exhaustion. Cancer Immunol. Immunother. 2023, 72, 3985–3999. [Google Scholar] [CrossRef] [PubMed]
- Sharonov, G.V.; Serebrovskaya, E.O.; Yuzhakova, D.V.; Britanova, O.V.; Chudakov, D.M. B cells, plasma cells and antibody repertoires in the tumour microenvironment. Nat. Rev. Immunol. 2020, 20, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, A.J.A.; Rinner, B.; Wenzl, K.; Pichler, M.; Troppan, K.; Steinbauer, E.; Schwarzenbacher, D.; Reitter, S.; Feichtinger, J.; Tierling, S.; et al. NR4A1-mediated apoptosis suppresses lymphomagenesis and is associated with a favorable cancer-specific survival in patients with aggressive B-cell lymphomas. Blood 2014, 123, 2367–2377. [Google Scholar] [CrossRef]
- Tan, C.; Hiwa, R.; Mueller, J.L.; Vykunta, V.; Hibiya, K.; Noviski, M.; Huizar, J.; Brooks, J.F.; Garcia, J.; Heyn, C.; et al. NR4A nuclear receptors restrain B cell responses to antigen when second signals are absent or limiting. Nat. Immunol. 2020, 21, 1267–1279. [Google Scholar] [CrossRef]
- Wang, L.; Xiao, Y.; Luo, Y.; Master, R.P.; Mo, J.; Kim, M.-C.; Liu, Y.; Maharjan, C.K.; Patel, U.M.; De, U.; et al. PROTAC-mediated NR4A1 degradation as a novel strategy for cancer immunotherapy. J. Exp. Med. 2024, 221, e20231519. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, Y.; Zhu, B. T-cell exhaustion in the tumor microenvironment. Cell Death Dis. 2015, 6, e1792. [Google Scholar] [CrossRef]
- Whelan, M.C.; Casey, G.; MacConmara, M.; Lederer, J.A.; Soden, D.; Collins, J.K.; Tangney, M.; O’Sullivan, G.C. Effective immunotherapy of weakly immunogenic solid tumours using a combined immunogene therapy and regulatory T-cell inactivation. Cancer Gene Ther. 2010, 17, 501–511. [Google Scholar] [CrossRef]
- Peralta, R.M.; Xie, B.; Lontos, K.; Nieves-Rosado, H.; Spahr, K.; Joshi, S.; Ford, B.R.; Quann, K.; Frisch, A.T.; Dean, V.; et al. Dysfunction of exhausted T cells is enforced by MCT11-mediated lactate metabolism. Nat. Immunol. 2024, 25, 2297–2307. [Google Scholar] [CrossRef]
- Yao, H.; Lan, J.; Li, C.; Shi, H.; Brosseau, J.-P.; Wang, H.; Lu, H.; Fang, C.; Zhang, Y.; Liang, L.; et al. Inhibiting PD-L1 palmitoylation enhances T-cell immune responses against tumours. Nat. Biomed. Eng. 2019, 3, 306–317. [Google Scholar] [CrossRef]
- Tang, H.; Liang, Y.; Anders, R.A.; Taube, J.M.; Qiu, X.; Mulgaonkar, A.; Liu, X.; Harrington, S.M.; Guo, J.; Xin, Y.; et al. PD-L1 on host cells is essential for PD-L1 blockade-mediated tumor regression. J. Clin. Investig. 2018, 128, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Diskin, B.; Adam, S.; Cassini, M.F.; Sanchez, G.; Liria, M.; Aykut, B.; Buttar, C.; Li, E.; Sundberg, B.; Salas, R.D.; et al. PD-L1 engagement on T cells promotes self-tolerance and suppression of neighboring macrophages and effector T cells in cancer. Nat. Immunol. 2020, 21, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jiang, P.; Wei, S.; Xu, X.; Wang, J. Regulatory T cells in tumor microenvironment: New mechanisms, potential therapeutic strategies and future prospects. Mol. Cancer 2020, 19, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Oberle, N.; Krammer, P.H. Molecular mechanisms of treg-mediated T cell suppression. Front. Immunol. 2012, 3, 51. [Google Scholar] [CrossRef]
- Malek, T.R. The biology of interleukin-2. Annu. Rev. Immunol. 2008, 26, 453–479. [Google Scholar] [CrossRef]
- Ross, S.H.; Cantrell, D.A. Signaling and Function of Interleukin-2 in T Lymphocytes. Annu. Rev. Immunol. 2018, 36, 411–433. [Google Scholar] [CrossRef]
- Chinen, T.; Kannan, A.K.; Levine, A.G.; Fan, X.; Klein, U.; Zheng, Y.; Gasteiger, G.; Feng, Y.; Fontenot, J.D.; Rudensky, A.Y. An essential role for the IL-2 receptor in Treg cell function. Nat. Immunol. 2016, 17, 1322–1333. [Google Scholar] [CrossRef]
- Suzuki, H.; Kündig, T.M.; Furlonger, C.; Wakeham, A.; Timms, E.; Matsuyama, T.; Schmits, R.; Simard, J.J.L.; Ohashi, P.S.; Griesser, H.; et al. Deregulated T cell activation and autoimmunity in mice lacking interleukin-2 receptor beta. Science 1995, 268, 1472–1476. [Google Scholar] [CrossRef]
- Willerford, D.M.; Chen, J.; Ferry, J.A.; Davidson, L.; Ma, A.; Alt, F.W. Interleukin-2 receptor alpha chain regulates the size and content of the peripheral lymphoid compartment. Immunity 1995, 3, 521–530. [Google Scholar] [CrossRef]
- Alspach, E.; Lussier, D.M.; Schreiber, R.D. Interferon γ and its important roles in promoting and inhibiting spontaneous and therapeutic cancer immunity. Cold Spring Harb. Perspect. Biol. 2019, 11, a028480. [Google Scholar] [CrossRef]
- Srirat, T.; Hayakawa, T.; Mise-Omata, S.; Nakagawara, K.; Ando, M.; Shichino, S.; Ito, M.; Yoshimura, A. NR4a1/2 deletion promotes accumulation of TCF1+ stem-like precursors of exhausted CD8+ T cells in the tumor microenvironment. Cell Rep. 2024, 43, 113898. [Google Scholar] [CrossRef] [PubMed]
- Karki, K.; Wright, G.A.; Mohankumar, K.; Jin, U.-H.; Zhang, X.-H.; Safe, S. A Bis-Indole-Derived NR4A1 Antagonist Induces PD-L1 Degradation and Enhances Antitumor Immunity. Cancer Res. 2020, 80, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Y.; Xia, H.; Chen, Y.H. Monocytes in tumorigenesis and tumor immunotherapy. Cells 2023, 12, 1673. [Google Scholar] [CrossRef] [PubMed]
- Ziegler-Heitbrock, L.; Ancuta, P.; Crowe, S.; Dalod, M.; Grau, V.; Hart, D.N.; Leenen, P.J.M.; Liu, Y.-J.; MacPherson, G.; Randolph, G.J.; et al. Nomenclature of monocytes and dendritic cells in blood. Blood 2010, 116, e74–e80. [Google Scholar] [CrossRef]
- Passlick, B.; Flieger, D.; Ziegler-Heitbrock, H. Identification and characterization of a novel monocyte subpopulation in human peripheral blood. Blood 1989, 74, 2527–2534. [Google Scholar] [CrossRef]
- Zawada, A.M.; Rogacev, K.S.; Rotter, B.; Winter, P.; Marell, R.R.; Fliser, D.; Heine, G.H. SuperSAGE evidence for CD14++CD16+ monocytes as a third monocyte subset. Blood 2011, 118, e50–e61. [Google Scholar] [CrossRef]
- Qian, B.-Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef]
- Movahedi, K.; Laoui, D.; Gysemans, C.; Baeten, M.; Stangé, G.; Van den Bossche, J.; Mack, M.; Pipeleers, D.; Veld, P.I.; De Baetselier, P.; et al. Different tumor microenvironments contain functionally distinct subsets of macrophages derived from Ly6C(high) monocytes. Cancer Res. 2010, 70, 5728–5739. [Google Scholar] [CrossRef]
- Shibuya, T.; Kamiyama, A.; Sawada, H.; Kikuchi, K.; Maruyama, M.; Sawado, R.; Ikeda, N.; Asano, K.; Kurotaki, D.; Tamura, T.; et al. Immunoregulatory monocyte subset promotes metastasis associated with therapeutic intervention for primary tumor. Front. Immunol. 2021, 12, 663115. [Google Scholar] [CrossRef]
- Geissmann, F.; Jung, S.; Littman, D.R. Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity 2003, 19, 71–82. [Google Scholar] [CrossRef]
- Auffray, C.; Fogg, D.; Garfa, M.; Elain, G.; Join-Lambert, O.; Kayal, S.; Sarnacki, S.; Cumano, A.; Lauvau, G.; Geissmann, F. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 2007, 317, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Carlin, L.M.; Stamatiades, E.G.; Auffray, C.; Hanna, R.N.; Glover, L.; Vizcay-Barrena, G.; Hedrick, C.C.; Cook, H.T.; Diebold, S.; Geissmann, F. Nr4a1-dependent Ly6C(low) monocytes monitor endothelial cells and orchestrate their disposal. Cell 2013, 153, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Palframan, R.T.; Jung, S.; Cheng, G.; Weninger, W.; Luo, Y.; Dorf, M.; Littman, D.R.; Rollins, B.J.; Zweerink, H.; Rot, A.; et al. Inflammatory chemokine transport and presentation in HEV: A remote control mechanism for monocyte recruitment to lymph nodes in inflamed tissues. J. Exp. Med. 2001, 194, 1361–1374. [Google Scholar] [CrossRef] [PubMed]
- Cassetta, L.; Pollard, J.W. Cancer immunosurveillance: Role of patrolling monocytes. Cell Res. 2015, 26, 3–4. [Google Scholar] [CrossRef]
- Melaiu, O.; Lucarini, V.; Cifaldi, L.; Fruci, D. Influence of the tumor microenvironment on NK cell function in solid tumors. Front. Immunol. 2020, 10, 3038. [Google Scholar] [CrossRef]
- Zhang, E.; Ding, C.; Li, S.; Zhou, X.; Aikemu, B.; Fan, X.; Sun, J.; Zheng, M.; Yang, X. Roles and mechanisms of tumour-infiltrating B cells in human cancer: A new force in immunotherapy. Biomark. Res. 2023, 11, 28. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Boutilier, A.J.; Elsawa, S.F. Macrophage polarization states in the tumor microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef]
- Kainulainen, K.; Takabe, P.; Heikkinen, S.; Aaltonen, N.; de la Motte, C.; Rauhala, L.; Durst, F.C.; Oikari, S.; Hukkanen, T.; Rahunen, E.; et al. M1 Macrophages Induce Protumor Inflammation in Melanoma Cells through TNFR-NF-κB Signaling. J. Investig. Dermatol. 2022, 142, 3041–3051. [Google Scholar] [CrossRef]
- Ma, Y.; Shurin, G.V.; Peiyuan, Z.; Shurin, M.R. Dendritic cells in the cancer microenvironment. J. Cancer 2013, 4, 36–44. [Google Scholar] [CrossRef]
- Del Prete, A.; Salvi, V.; Soriani, A.; Laffranchi, M.; Sozio, F.; Bosisio, D.; Sozzani, S. Dendritic cell subsets in cancer immunity and tumor antigen sensing. Cell. Mol. Immunol. 2023, 20, 432–447. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.W.; Kim, K.-D.; Lee, H.K. The role of dendritic cells in tumor microenvironments and their uses as therapeutic targets. BMB Rep. 2021, 54, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.L.; Murray, T.F.P.Z.; Beckham, K.S.H.; Douce, G.; Mowat, A.M. Signal regulatory protein alpha (SIRPα) regulates the homeostasis of CD103(+) CD11b(+) DCs in the intestinal lamina propria. Eur. J. Immunol. 2014, 44, 3658–3668. [Google Scholar] [CrossRef]
- Zheng, J.; Wang, M.; Pang, L.; Wang, S.; Kong, Y.; Zhu, X.; Zhou, X.; Wang, X.; Chen, C.; Ning, H.; et al. Identification of a novel DEC-205 binding peptide to develop dendritic cell-targeting nanovaccine for cancer immunotherapy. J. Control. Release 2024, 373, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Carelock, M.E.; Master, R.P.; Kim, M.-C.; Jin, Z.; Wang, L.; Maharjan, C.K.; Hua, N.; De, U.; Kolb, R.; Xiao, Y.; et al. Targeting intracellular proteins with cell type-specific functions for cancer immunotherapy. Life Med. 2023, 2, lnad019. [Google Scholar] [CrossRef]
- Hedrick, E.; Li, X.; Cheng, Y.; Lacey, A.; Mohankumar, K.; Zarei, M.; Safe, S. Potent inhibition of breast cancer by bis-indole-derived nuclear receptor 4A1 (NR4A1) antagonists. Breast Cancer Res. Treat. 2019, 177, 29–40. [Google Scholar] [CrossRef]
- Shrestha, R.; Mohankumar, K.; Martin, G.; Hailemariam, A.; Lee, S.-O.; Jin, U.-H.; Burghardt, R.; Safe, S. Flavonoids kaempferol and quercetin are nuclear receptor 4A1 (NR4A1, Nur77) ligands and inhibit rhabdomyosarcoma cell and tumor growth. J. Exp. Clin. Cancer Res. 2021, 40, 1–17. [Google Scholar] [CrossRef]
- Liu, J.; Wang, G.-H.; Duan, Y.-H.; Dai, Y.; Bao, Y.; Hu, M.; Zhou, Y.-Q.; Li, M.; Jiang, F.; Zhou, H.; et al. Modulation of the Nur77-Bcl-2 apoptotic pathway by p38α MAPK. Oncotarget 2017, 8, 69731–69745. [Google Scholar] [CrossRef]
- Hu, M.; Luo, Q.; Alitongbieke, G.; Chong, S.; Xu, C.; Xie, L.; Chen, X.; Zhang, D.; Zhou, Y.; Wang, Z.; et al. Celastrol-Induced Nur77 Interaction with TRAF2 Alleviates Inflammation by Promoting Mitochondrial Ubiquitination and Autophagy. Mol. Cell 2017, 66, 141–153. [Google Scholar] [CrossRef]
- Wang, C.; Dai, S.; Zhao, X.; Zhang, Y.; Gong, L.; Fu, K.; Ma, C.; Peng, C.; Li, Y. Celastrol as an emerging anticancer agent: Current status, challenges and therapeutic strategies. Biomed. Pharmacother. 2023, 163, 114882. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, Y.; Luo, W.; Chen, S.; Lin, F.; Zhang, X.; Fan, S.; Shen, X.; Wang, Y.; Liang, G. Celastrol induces ROS-mediated apoptosis via directly targeting peroxiredoxin-2 in gastric cancer cells. Theranostics 2020, 10, 10290–10308. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, M.; Yang, Y.; Du, C.; Zhou, H.; Liu, C.; Chen, Y.; Fan, L.; Ma, H.; Gong, Y.; et al. An overview of PROTACs: A promising drug discovery paradigm. Mol. Biomed. 2022, 3, 46. [Google Scholar] [CrossRef]
- Zhan, Y.; Du, X.; Chen, H.; Liu, J.; Zhao, B.; Huang, D.; Li, G.; Xu, Q.; Zhang, M.; Weimer, B.C.; et al. Cytosporone B is an agonist for nuclear orphan receptor Nur77. Nat. Chem. Biol. 2008, 4, 548–556. [Google Scholar] [CrossRef]
- Mullican, S.E.; Zhang, S.; Konopleva, M.; Ruvolo, V.; Andreeff, M.; Milbrandt, J.; Conneely, O.M. Abrogation of nuclear receptors Nr4a3 and Nr4a1 leads to development of acute myeloid leukemia. Nat. Med. 2007, 13, 730–735. [Google Scholar] [CrossRef]
- Boudreaux, S.P.; Ramirez-Herrick, A.M.; Duren, R.P.; Conneely, O.M. Genome-wide profiling reveals transcriptional repression of MYC as a core component of NR4A tumor suppression in acute myeloid leukemia. Oncogenesis 2012, 1, e19. [Google Scholar] [CrossRef]
- Boudreaux, S.P.; Duren, R.P.; Call, S.G.; Nguyen, L.; Freire, P.R.; Narayanan, P.; Redell, M.S.; Conneely, O.M. Drug targeting of NR4A nuclear receptors for treatment of acute myeloid leukemia. Leukemia 2018, 33, 52–63. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Types | Tumor Types | Effects of NR4A1 | Mechanism of Action | Summary |
|---|---|---|---|---|
| Monocytes [10,11,12] | Melanoma and lung | Inhibition of NR4A1 leads to monocytes being trapped in the S/G2 phase of cell cycle, decreasing proliferation and activation. | Decreased NR4A1 is correlated with CX3CR1, JunB, and C/EBP-β, which are essential in monocyte differentiation and proliferation. | Since the depletion of NR4A1 in monocytes leads to decreased proliferation and differentiation, an agonist that can promote its effects within this cell type can aid in combating tumor progression. |
| Neutrophils [13,14] | Colon adenocarcinomas (COAD) | High NR4A1 expression leads to better prognosis in COAD. | Predictive models identify NR4A1 as a major player in tumor survival. | Depending on NR4A1 expression levels, different therapeutic drugs can be utilized for treatment. Higher levels of NR4A1 expression are associated with better outcomes. More research is needed to determine the impact, if any, of NR4A1 in neutrophil function. |
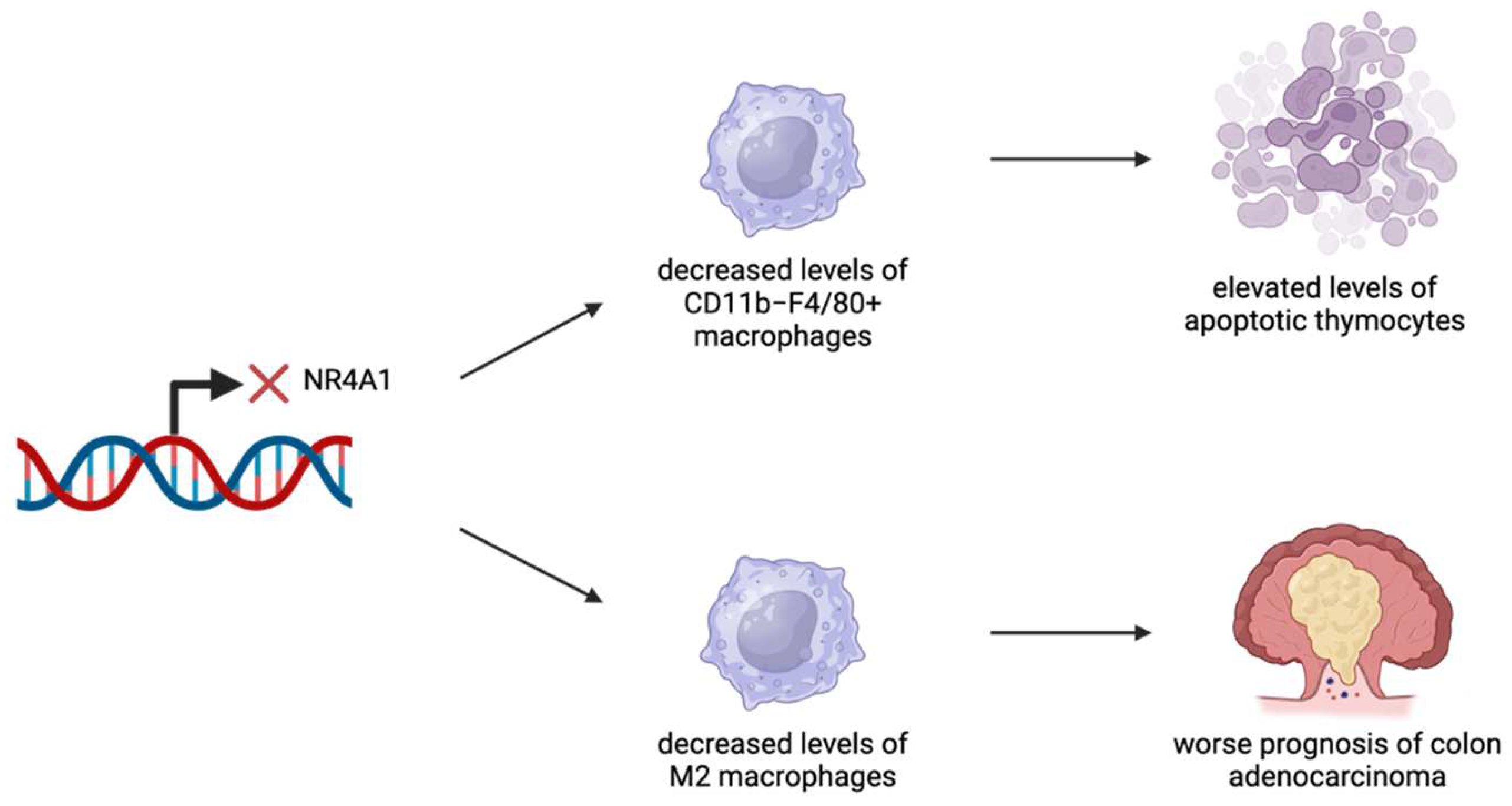
| Macrophages [13,15] | Apoptotic thymocytes, COADs | Inadequate levels of macrophages lead to difficulties in degrading apoptotic thymocytes. High NR4A1 expression leads to better prognosis in COAD. | Lack of NR4A1 expression leads to a decrease in macrophage production. Predictive models identify NR4A1 as a major player in tumor survival. | NR4A1 expression is vital for macrophages to successfully degrade apoptotic thymocytes. Depending on NR4A1 expression levels, different therapeutic drugs can be utilized for treatment. Higher levels of NR4A1 expression are associated with better outcomes. |
| NK Cells [16,17,18,19] | Head and neck squamous carcinomas, lung cancer, hepatocellular carcinoma | Overexpression can lead to tumor progression and increased metastasis. | Overexpression of NR4A1 is associated with decreased production of granzyme and perforin and inhibition of IFN-γ pathway. | Tumor-associated NK cells show an overexpression of NR4A1, which correlates with increased tumor progression and poor prognosis. |
| Dendritic Cells (DCs) [9] | Acute lymphoblastic leukemia | Activation of NR4A1 leads to decreased inflammatory responses. | Through the regulation of NF-κB pathway, IL-6 and TNFα production is decreased. | NR4A1 expression in DCs can vary depending on immune stimuli, but deficiency has been associated with elevated inflammatory responses. More research is needed to determine specific pathways of mechanisms in different DC subsets. |
| T cells [20,21,22] | Melanoma, colon, lung, and other solid tumors | Inhibition of T cell development and differentiation. | Blocks IL-2 transcription by competing with AP-1 binding and decreasing IFN-γ production. Contributes to reduced effector cytokine production and increased expression of T cell exhaustion genes. | Since NR4A1 exerts an inhibitory effect on T-cells through the decreased production of IL-2 and IFN-γ, a treatment targeting NR4A1 in these cells can suppress tumor progression. |
| B cells [23,24,25,26] | B cell lymphoma and melanoma | In B cell lymphoma, overexpression of NR4A1 leads to increased apoptosis and decreased tumor progression. In melanoma studies, degradation of NR4A1 led to increased proliferation of tumor-infiltrating B cells and decreased tumor progression. | In B cell lymphoma, NR4A1 leads to the increased expression of the apoptotic genes TRAIL, Bim, and Puma. In melanoma studies, NR4A1 limits the ability of B cells to be activated without co-stimulatory responses. | In B cell lymphoma, a drug agonist for NR4A1 can lead to increased tumor apoptosis, leading to better clinical results. In melanoma, a drug that inhibits NR4A1 can lead to increased tumor-infiltrating B cells and decreased tumor progression. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleberg, J.; Nataraj, A.; Xiao, Y.; Podder, B.R.; Jin, Z.; Tithi, T.I.; Zheng, G.; Smalley, K.S.M.; Moser, E.K.; Safe, S.; et al. Targeting Lineage-Specific Functions of NR4A1 for Cancer Immunotherapy. Int. J. Mol. Sci. 2025, 26, 5266. https://doi.org/10.3390/ijms26115266
Kleberg J, Nataraj A, Xiao Y, Podder BR, Jin Z, Tithi TI, Zheng G, Smalley KSM, Moser EK, Safe S, et al. Targeting Lineage-Specific Functions of NR4A1 for Cancer Immunotherapy. International Journal of Molecular Sciences. 2025; 26(11):5266. https://doi.org/10.3390/ijms26115266
Chicago/Turabian StyleKleberg, Jeremy, Akhila Nataraj, Yufeng Xiao, Bristy R. Podder, Zeng Jin, Tanzia Islam Tithi, Guangrong Zheng, Keiran S. M. Smalley, Emily K. Moser, Stephen Safe, and et al. 2025. "Targeting Lineage-Specific Functions of NR4A1 for Cancer Immunotherapy" International Journal of Molecular Sciences 26, no. 11: 5266. https://doi.org/10.3390/ijms26115266
APA StyleKleberg, J., Nataraj, A., Xiao, Y., Podder, B. R., Jin, Z., Tithi, T. I., Zheng, G., Smalley, K. S. M., Moser, E. K., Safe, S., Maharjan, C. K., Kolb, R., & Zhang, W. (2025). Targeting Lineage-Specific Functions of NR4A1 for Cancer Immunotherapy. International Journal of Molecular Sciences, 26(11), 5266. https://doi.org/10.3390/ijms26115266