
Plant–Fungi Mutualism, Alternative Splicing, and Defense Responses: Balancing Symbiosis and Immunity



Abstract
1. Introduction
2. Classification of Symbiotic Fungi and Their Ecological Functions
2.1. Diversity of Symbiotic Fungi
2.2. Effects of Symbiotic Fungi on Plant Growth and Development
3. Role of Alternative Splicing in Plant–Fungi Symbiosis
3.1. Effects of Alternative Splicing on Plants
3.2. The Complexity of Alternative Splicing Regulation in Fungi
3.3. Involvement of Alternative Splicing in Fungi Interactions
4. Molecular Mechanisms Associated with Fungus Symbiosis via Alternative Splicing
4.1. Molecular Mechanism of the Host’s Regulatory Factors
4.2. Responsive Factors from Commensal Fungi
5. Small Molecules and Their Target Proteins That Can Modulate Plant–Fungus Interactions
6. Summary and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal Rewards Stabilize Cooperation in the Mycorrhizal Symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Delaux, P.M.; Schornack, S. Plant evolution driven by interactions with symbiotic and pathogenic microbes. Science 2021, 371, eaba6605. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.M.; Karanja, J.; Bello, S.K. Piriformospora indica colonization reprograms plants to improved P-uptake, enhanced crop performance, and biotic/abiotic stress tolerance. Physiol. Mol. Plant Pathol. 2019, 106, 232–237. [Google Scholar] [CrossRef]
- Vahabi, K.; Sherameti, I.; Bakshi, M.; Mrozinska, A.; Ludwig, A.; Oelmuller, R. Microarray analyses during early and later stages of the Arabidopsis/Piriformospora indica interaction. Genom. Data 2015, 6, 16–18. [Google Scholar] [CrossRef]
- Yamato, M.; Yagita, M.; Kusakabe, R.; Shimabukuro, K.; Yamana, K.; Suetsugu, K. Impact of mycoheterotrophy on the growth of Gentiana zollingeri (Gentianaceae), as suggested by size variation, morphology, and 13C abundance of flowering shoots. J. Plant Res. 2023, 136, 853–863. [Google Scholar] [CrossRef]
- Wang, W.; Shi, J.; Xie, Q.; Jiang, Y.; Yu, N.; Wang, E. Nutrient Exchange and Regulation in Arbuscular Mycorrhizal Symbiosis. Mol. Plant 2017, 10, 1147–1158. [Google Scholar] [CrossRef]
- Chen, S.Y.; Li, C.; Jia, X.B.; Lai, S.J. Sequence and Evolutionary Features for the Alternatively Spliced Exons of Eukaryotic Genes. Int. J. Mol. Sci. 2019, 20, 3834. [Google Scholar] [CrossRef]
- Wright, C.J.; Smith, C.W.J.; Jiggins, C.D. Alternative splicing as a source of phenotypic diversity. Nat. Rev. Genet. 2022, 23, 697–710. [Google Scholar] [CrossRef]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef]
- Wen, J.J.; Qin, Z.; Sun, L.; Zhang, Y.M.; Wang, D.L.; Peng, H.R.; Yao, Y.Y.; Hu, Z.R.; Ni, Z.F.; Sun, Q.X.; et al. Alternative splicing of TaHSFA6e modulates heat shock protein-mediated translational regulation in response to heat stress in wheat. New Phytol. 2023, 239, 2235–2247. [Google Scholar] [CrossRef]
- Li, Z.X.; Tang, J.; Bassham, D.C.; Howell, S.H. Daily temperature cycles promote alternative splicing of RNAs encoding SR45a, a splicing regulator in maize. Plant Physiol. 2021, 186, 1318–1335. [Google Scholar] [CrossRef] [PubMed]
- Li, H.X.; Li, A.X.; Shen, W.; Ye, N.H.; Wang, G.Q.; Zhang, J.H. Global Survey of Alternative Splicing in Rice by Direct RNA Sequencing During Reproductive Development: Landscape and Genetic Regulation. Rice 2021, 14, 75. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.X.; Zhang, D.D.; Li, Z.W.; Liang, H.Z.; Deng, R.F.; Su, X.G.; Jiang, Y.M.; Duan, X.W. Alternative splicing of MaMYB16L regulates starch degradation in banana fruit during ripening. J. Integr. Plant Biol. 2021, 63, 1341–1352. [Google Scholar] [CrossRef]
- Zeng, Z.; Liu, Y.; Feng, X.Y.; Li, S.X.; Jiang, X.M.; Chen, J.Q.; Shao, Z.Q. The RNAome landscape of tomato during arbuscular mycorrhizal symbiosis reveals an evolving RNA layer symbiotic regulatory network. Plant Commun. 2023, 4, 100429. [Google Scholar] [CrossRef]
- Zorin, E.A.; Afonin, A.M.; Kulaeva, O.A.; Gribchenko, E.S.; Shtark, O.Y.; Zhukov, V.A. Transcriptome Analysis of Alternative Splicing Events Induced by Arbuscular Mycorrhizal Fungi (Rhizophagus irregularis) in Pea (Pisum sativum L.) Roots. Plants 2020, 9, 700. [Google Scholar] [CrossRef]
- Zhang, X.H.; Gao, H.M.; Liang, Y.Q.; Cao, Y.P. Full-length transcriptome analysis of asparagus roots reveals the molecular mechanism of salt tolerance induced by arbuscular mycorrhizal fungi. Environ. Exp. Bot. 2021, 185, 104402. [Google Scholar] [CrossRef]
- Wang, C.; Zhu, H.; Jin, L.P.; Chen, T.; Wang, L.X.; Kang, H.; Hong, Z.L.; Zhang, Z.M. Splice variants of the SIP1 transcripts play a role in nodule organogenesis in Lotus japonicus. Plant Mol. Biol. 2013, 82, 97–111. [Google Scholar] [CrossRef]
- Huisman, R.; Hontelez, J.; Mysore, K.S.; Wen, J.Q.; Bisseling, T.; Limpens, E. A symbiosis-dedicated SYNTAXIN OF PLANTS 13II isoform controls the formation of a stable host-microbe interface in symbiosis. New Phytol. 2016, 211, 1338–1351. [Google Scholar] [CrossRef]
- Liu, Y.N.; Liu, C.C.; Guo, R.; Tian, L.; Cheng, J.F.; Wu, Y.N.; Wang, D.; Wang, B. The Rice Qa-SNAREs in SYP13 Subfamily Are Involved in Regulating Arbuscular Mycorrhizal Symbiosis and Seed Fertility. Front. Plant Sci. 2022, 13, 898286. [Google Scholar] [CrossRef]
- Ruzicka, D.; Chamala, S.; Barrios-Masias, F.H.; Martin, F.; Smith, S.; Jackson, L.E.; Barbazuk, W.B.; Schachtman, D.P. Inside Arbuscular Mycorrhizal Roots-Molecular Probes to Understand the Symbiosis. Plant Genome 2013, 6, 13. [Google Scholar] [CrossRef]
- Gomez-Gallego, T.; Benabdellah, K.; Merlos, M.A.; Jimenez-Jimenez, A.M.; Alcon, C.; Berthomieu, P.; Ferrol, N. The Rhizophagus irregularis Genome Encodes Two CTR Copper Transporters That Mediate Cu Import Into the Cytosol and a CTR-Like Protein Likely Involved in Copper Tolerance. Front. Plant Sci. 2019, 10, 604. [Google Scholar] [CrossRef] [PubMed]
- Kloppholz, S.; Kuhn, H.; Requena, N. A Secreted Fungal Effector of Glomus intraradices Promotes Symbiotic Biotrophy. Curr. Biol. 2011, 21, 1204–1209. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef]
- Zheng, L.S.; Zhao, S.P.; Zhou, Y.F.; Yang, G.L.; Chen, A.; Li, X.X.; Wang, J.X.; Tian, J.; Liao, H.; Wang, X.R. The soybean sugar transporter GmSWEET6 participates in sucrose transport towards fungi during arbuscular mycorrhizal symbiosis. Plant Cell Environ. 2024, 47, 1041–1052. [Google Scholar] [CrossRef]
- Peterson, R.L.P.L.; Massicotte, H.B.; Melville, L.H. Mycorrhizas: Anatomy and Cell Biology; NRC Research Press: Ottawa, ON, Canada, 2004. [Google Scholar]
- Prieto, I.; Roldán, A.; Huygens, D.; del Mar Alguacil, M.; Navarro-Cano, J.A.; Querejeta, J.I. Species-specific roles of ectomycorrhizal fungi in facilitating interplant transfer of hydraulically redistributed water between Pinus halepensis saplings and seedlings. Plant Soil 2016, 406, 15–27. [Google Scholar] [CrossRef]
- Policelli, N.; Hoeksema, J.D.; Moyano, J.; Vilgalys, R.; Vivelo, S.; Bhatnagar, J.M. Global pine tree invasions are linked to invasive root symbionts. New Phytol. 2023, 237, 16–21. [Google Scholar] [CrossRef]
- Cao, D.C. “Smother love” of mycorrhiza: Hypoxic germination of orchids. Plant Physiol. 2023, 192, 2240–2242. [Google Scholar] [CrossRef]
- Chen, X.G.; Wu, Y.H.; Li, N.Q.; Gao, J.Y. What role does the seed coat play during symbiotic seed germination in orchids: An experimental approach with Dendrobium officinale. BMC Plant Biol. 2022, 22, 375. [Google Scholar] [CrossRef]
- Gao, X.Z.; Wang, Y.; Deng, D.; Luo, Y.L.; Shao, S.C.; Luo, Y. Morphogenesis Changes in Protocorm Development during Symbiotic Seed Germination of Dendrobium chrysotoxum (Orchidaceae) with Its Mycobiont, Tulasnella sp. Horticulturae 2023, 9, 531. [Google Scholar] [CrossRef]
- Shao, S.C.; Wang, Q.X.; Beng, K.C.; Zhao, D.K.; Jacquemyn, H. Fungi isolated from host protocorms accelerate symbiotic seed germination in an endangered orchid species (Dendrobium chrysotoxum) from southern China. Mycorrhiza 2020, 30, 529–539. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.Y.; Chen, X.M.; Guo, S.X.; Lee, Y.I. Effect of different mycobionts on symbiotic germination and seedling growth of Dendrobium officinale, an important medicinal orchid. Bot. Stud. 2020, 61, 10. [Google Scholar] [CrossRef]
- Qin, X.Y.; Xu, J.; An, X.L.; Yang, J.; Wang, Y.; Dou, M.J.; Wang, M.G.; Huang, J.; Fu, Y.J. Insight of endophytic fungi promoting the growth and development of woody plants. Crit. Rev. Biotechnol. 2023, 22, 78–99. [Google Scholar] [CrossRef]
- Wu, Y.H.; Wang, H.; Liu, M.; Li, B.; Chen, X.; Ma, Y.T.; Yan, Z.Y. Effects of Native Arbuscular Mycorrhizae Isolated on Root Biomass and Secondary Metabolites of Salvia miltiorrhiza Bge. Front. Plant Sci. 2021, 12, 617892. [Google Scholar] [CrossRef]
- Chen, S.C.; Zhao, H.J.; Zou, C.C.; Li, Y.S.; Chen, Y.F.; Wang, Z.H.; Jiang, Y.; Liu, A.R.; Zhao, P.Y.; Wang, M.M.; et al. Combined Inoculation with Multiple Arbuscular Mycorrhizal Fungi Improves Growth, Nutrient Uptake and Photosynthesis in Cucumber Seedlings. Front. Microbiol. 2017, 8, 2516. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Nie, Y.Y.; Fan, X.N.; Wei, W.; Chen, H.; Xie, X.A.; Tang, M. A transcriptional activator from Rhizophagus irregularis regulates phosphate uptake and homeostasis in AM symbiosis during phosphorous starvation. Front. Microbiol. 2023, 13, 21. [Google Scholar] [CrossRef]
- Cao, M.A.; Liu, R.C.; Xiao, Z.Y.; Hashem, A.; Abd Allah, E.F.; Alsayed, M.F.; Harsonowati, W.; Wu, Q.S. Symbiotic Fungi Alter the Acquisition of Phosphorus in Camellia oleifera through Regulating Root Architecture, Plant Phosphate Transporter Gene Expressions and Soil Phosphatase Activities. J. Fungi 2022, 8, 800. [Google Scholar] [CrossRef]
- Morsy, M.; Cleckler, B.; Armuelles-Millican, H. Fungal Endophytes Promote Tomato Growth and Enhance Drought and Salt Tolerance. Plants 2020, 9, 877. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, W.W.; Qi, S.S.; Cheng, H.; Li, Q.; Ran, Q.; Dai, Z.C.; Du, D.L.; Egan, S.; Thomas, T. Arbuscular mycorrhizal fungi improve the growth and disease resistance of the invasive plant Wedelia trilobata. J. Appl. Microbiol. 2021, 130, 582–591. [Google Scholar] [CrossRef]
- Shafiei, F.; Shahidi-Noghabi, S.; Sedaghati, E. The impact of arbuscular mycorrhizal fungi on tomato plant resistance against Tuta absoluta (Meyrick) in greenhouse conditions. J. Asia-Pac. Entomol. 2022, 25, 8. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Mosaddeghi, M.R.; Hosseini, F.; Hajabbasi, M.A.; Sabzalian, M.R.; Sepehri, M. Epichloe spp. and Serendipita indica endophytic fungi: Functions in plant-soil relations. In Advances in Agronomy; Sparks, D.L., Ed.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2021; Volume 165, pp. 59–113. [Google Scholar]
- Kou, M.Z.; Bastías, D.A.; Christensen, M.J.; Zhong, R.; Nan, Z.B.; Zhang, X.X. The Plant Salicylic Acid Signalling Pathway Regulates the Infection of a Biotrophic Pathogen in Grasses Associated with an Epichloe Endophyte. J. Fungi 2021, 7, 633. [Google Scholar] [CrossRef]
- Bu, Y.Y.; Guo, P.; Ji, Y.L.; Zhang, S.T.; Yu, H.S.; Wang, Z.W. Effects of Epichloe sinica on Roegneria kamoji seedling physiology under PEG-6000 simulated drought stress. Symbiosis 2019, 77, 123–132. [Google Scholar] [CrossRef]
- Nagabhyru, P.; Dinkins, R.D.; Schardl, C.L. Transcriptome analysis of Epichloe strains in tall fescue in response to drought stress. Mycologia 2022, 114, 697–712. [Google Scholar] [CrossRef]
- Cibils-Stewart, X.; Powell, J.R.; Popay, A.J.; Lattanzi, F.A.; Hartley, S.E.; Johnson, S.N. Reciprocal Effects of Silicon Supply and Endophytes on Silicon Accumulation and Epichloe Colonization in Grasses. Front. Plant Sci. 2020, 11, 593198. [Google Scholar] [CrossRef]
- Jeong, S. SR Proteins: Binders, Regulators, and Connectors of RNA. Mol. Cells 2017, 40, 1–9. [Google Scholar] [CrossRef]
- Liu, Y.C.; Kim, D.; Choi, N.; Oh, J.; Ha, J.; Zhou, J.H.; Zheng, X.X.; Shen, H.H. hnRNP A1 Regulates Alternative Splicing of Tau Exon 10 by Targeting 3′ Splice Sites. Cells 2020, 9, 936. [Google Scholar] [CrossRef]
- Wilkinson, M.E.; Charenton, C.; Nagai, K. RNA Splicing by the Spliceosome. In Annual Review of Biochemistry; Kornberg, R.D., Ed.; Annual Reviews: Palo Alto, CA, USA, 2020; Volume 89, pp. 359–388. [Google Scholar]
- Dikaya, V.; El Arbi, N.; Rojas-Murcia, N.; Nardeli, S.M.; Goretti, D.; Schmid, M. Insights into the role of alternative splicing in plant temperature response. J. Exp. Bot. 2021, 72, 7384–7403. [Google Scholar] [CrossRef]
- Sybilska, E.; Daszkowska-Golec, A. Alternative splicing in ABA signaling during seed germination. Front. Plant Sci. 2023, 14, 1144990. [Google Scholar] [CrossRef]
- Xue, L.; Wang, E. Arbuscular mycorrhizal associations and the major regulators. Front. Agric. Sci. Eng. 2020, 7, 296–306. [Google Scholar] [CrossRef]
- Pan, H.R.; Oztas, O.; Zhang, X.W.; Wu, X.Y.; Stonoha, C.; Wang, E.; Wang, B.; Wang, D. A symbiotic SNARE protein generated by alternative termination of transcription. Nat. Plants 2016, 2, 15197. [Google Scholar] [CrossRef]
- Laloum, T.; Martin, G.; Duque, P. Alternative Splicing Control of Abiotic Stress Responses. Trends Plant Sci. 2018, 23, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Wang, W.; Li, H.; Li, H.; Yang, Y.; Zheng, H.; Tao, J. Anthocyanin accumulation in grape berry flesh is associated with an alternative splicing variant of VvMYBA1. Plant Physiol. Biochem. 2023, 195, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kufel, J.; Diachenko, N.; Golisz, A. Alternative splicing as a key player in the fine-tuning of the immunity response in Arabidopsis. Mol. Plant Pathol. 2022, 23, 1226–1238. [Google Scholar] [CrossRef]
- Shang, X.D.; Cao, Y.; Ma, L.G. Alternative Splicing in Plant Genes: A Means of Regulating the Environmental Fitness of Plants. Int. J. Mol. Sci. 2017, 18, 432. [Google Scholar] [CrossRef]
- Matsukura, S.; Mizoi, J.; Yoshida, T.; Todaka, D.; Ito, Y.; Maruyama, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Mol. Genet. Genom. 2010, 283, 185–196. [Google Scholar] [CrossRef]
- Shen, Y.F.; Wu, X.P.; Liu, D.M.; Song, S.J.; Liu, D.C.; Wang, H.Q. Cold-dependent alternative splicing of a Jumonji C domain-containing gene MtJMJC5 in Medicago truncatula. Biochem. Biophys. Res. Commun. 2016, 474, 271–276. [Google Scholar] [CrossRef]
- Kariman, K.; Barker, S.J.; Tibbett, M. Structural plasticity in root-fungal symbioses: Diverse interactions lead to improved plant fitness. PeerJ 2018, 6, e6030. [Google Scholar] [CrossRef]
- Fang, S.M.; Hou, X.; Qiu, K.H.; He, R.; Feng, X.S.; Liang, X.L. The occurrence and function of alternative splicing in fungi. Fungal Biol. Rev. 2020, 34, 178–188. [Google Scholar] [CrossRef]
- Dong, W.X.; Ding, J.L.; Gao, Y.; Peng, Y.J.; Feng, M.G.; Ying, S.H. Transcriptomic insights into the alternative splicing-mediated adaptation of the entomopathogenic fungus Beauveria bassiana to host niches: Autophagy-related gene 8 as an example. Environ. Microbiol. 2017, 19, 4126–4139. [Google Scholar] [CrossRef]
- Jia, Y.L.; Zhou, E.; Lee, S.; Bianco, T. Coevolutionary Dynamics of Rice Blast Resistance Gene Pi-ta and Magnaporthe oryzae Avirulence Gene AVR-Pita 1. Phytopathology 2016, 106, 676–683. [Google Scholar] [CrossRef]
- Rodriguez-Kessler, M.; Baeza-Montañez, L.; García-Pedrajas, M.D.; Tapia-Moreno, A.; Gold, S.; Jiménez-Bremont, J.F.; Ruiz-Herrera, J. Isolation of UmRrm75, a gene involved in dimorphism and virulence of Ustilago maydis. Microbiol. Res. 2012, 167, 270–282. [Google Scholar] [CrossRef] [PubMed]
- He, S.F.; Huang, K.; Li, B.G.; Lu, G.D.; Wang, A.R. Functional Analysis of a Salicylate Hydroxylase in Sclerotinia sclerotiorum. J. Fungi 2023, 9, 1169. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, K.V.; Tian, Z.P.; Wang, Y.F.; Smith, J.; Zylstra, G.; Huang, B.R.; Belanger, F.C. Functional characterization of salicylate hydroxylase from the fungal endophyte Epichloe festucae. Sci. Rep. 2015, 5, 10939. [Google Scholar] [CrossRef]
- De Bortoli, F.; Espinosa, S.; Zhao, R. DEAH-Box RNA Helicases in Pre-mRNA Splicing. Trends Biochem. Sci. 2021, 46, 225–238. [Google Scholar] [CrossRef]
- Hiruma, K.; Gerlach, N.; Sacristan, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramirez, D.; Bucher, M.; O'Connell, R.J.; et al. Root Endophyte Colletotrichum tofieldiae Confers Plant Fitness Benefits that Are Phosphate Status Dependent. Cell 2016, 165, 464–474. [Google Scholar] [CrossRef]
- Recchia, G.H.; Konzen, E.R.; Cassieri, F.; Caldas, D.G.G.; Tsai, S.M. Arbuscular Mycorrhizal Symbiosis Leads to Differential Regulation of Drought-Responsive Genes in Tissue-Specific Root Cells of Common Bean. Front. Microbiol. 2018, 9, 1339. [Google Scholar] [CrossRef]
- Liu, C.Y.; Zhang, F.; Zhang, D.J.; Srivastava, A.K.; Wu, Q.S.; Zou, Y.N. Mycorrhiza stimulates root-hair growth and IAA synthesis and transport in trifoliate orange under drought stress. Sci. Rep. 2018, 8, 1978. [Google Scholar] [CrossRef]
- Keymer, A.; Pimprikar, P.; Wewer, V.; Huber, C.; Brands, M.; Bucerius, S.L.; Delaux, P.M.; Klingl, V.; von Röpenack-Lahaye, E.; Wang, T.L.; et al. Lipid transfer from plants to arbuscular mycorrhiza fungi. eLife 2017, 6, e29107. [Google Scholar] [CrossRef]
- Luginbuehl, L.H.; Menard, G.N.; Kurup, S.; Van Erp, H.; Radhakrishnan, G.V.; Breakspear, A.; Oldroyd, G.E.D.; Eastmond, P.J. Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 2017, 356, 1175–1178. [Google Scholar] [CrossRef]
- Campos, C.; Carvalho, M.; Brígido, C.; Goss, M.J.; Nobre, T. Symbiosis Specificity of the Preceding Host Plant Can Dominate but Not Obliterate the Association Between Wheat and Its Arbuscular Mycorrhizal Fungal Partners. Front. Microbiol. 2018, 9, 2920. [Google Scholar] [CrossRef]
- Brotman, Y.; Lisec, J.; Méret, M.; Chet, I.; Willmitzer, L.; Viterbo, A. Transcript and metabolite analysis of the Trichoderma-induced systemic resistance response to Pseudomonas syringae in Arabidopsis thaliana. Microbiology 2012, 158, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-Z.; Li, Y.-K.; Chen, Y.-L.; Zhou, Y.; Sahu, S.K.; Liu, N.; Wu, H.; Shui, G.; Chen, Q.; Yao, N. Exploring the plant lipidome: Techniques, challenges, and prospects. Adv. Biotechnol. 2024, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, K.; Kobae, Y.; Toju, H. Beneficial associations between Brassicaceae plants and fungal endophytes under nutrient-limiting conditions: Evolutionary origins and host-symbiont molecular mechanisms. Curr. Opin. Plant Biol. 2018, 44, 145–154. [Google Scholar] [CrossRef]
- Fernández, I.; Cosme, M.; Stringlis, I.A.; Yu, K.; de Jonge, R.; van Wees, S.M.; Pozo, M.J.; Pieterse, C.M.J.; van der Heijden, M.G.A. Molecular dialogue between arbuscular mycorrhizal fungi and the nonhost plant Arabidopsis thaliana switches from initial detection to antagonism. New Phytol. 2019, 223, 867–881. [Google Scholar] [CrossRef]
- Zogli, P.; Libault, M. Plant response to biotic stress: Is there a common epigenetic response during plant-pathogenic and symbiotic interactions? Plant Sci. Int. J. Exp. Plant Biol. 2017, 263, 89–93. [Google Scholar] [CrossRef]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.; Huang, H.D.; Jin, H.L. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef]
- Cai, Q.; Qiao, L.L.; Wang, M.; He, B.Y.; Lin, F.M.; Palmquist, J.; Huang, S.N.D.; Jin, H.L. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef]
- Tian, W.J.; Ge, Y.Y.; Liu, X.Y.; Dou, G.M.; Ma, Y.C. Identification and characterization of Populus microRNAs in response to plant growth-promoting endophytic Streptomyces sp. SSD49. World J. Microbiol. Biotechnol. 2019, 35, 11. [Google Scholar] [CrossRef]
- Lauressergues, D.; Delaux, P.M.; Formey, D.; Lelandais-Brière, C.; Fort, S.; Cottaz, S.; Bécard, G.; Niebel, A.; Roux, C.; Combier, J.P. The microRNA miR171h modulates arbuscular mycorrhizal colonization of Medicago truncatula by targeting NSP2. Plant J. 2012, 72, 512–522. [Google Scholar] [CrossRef]
- Schaarschmidt, S.; Gresshoff, P.M.; Hause, B. Analyzing the soybean transcriptome during autoregulation of mycorrhization identifies the transcription factors GmNF-YA1a/b as positive regulators of arbuscular mycorrhization. Genome Biol. 2013, 14, R62. [Google Scholar] [CrossRef]
- Laloum, T.; De Mita, S.; Gamas, P.; Baudin, M.; Niebel, A. CCAAT-box binding transcription factors in plants: Y so many? Trends Plant Sci. 2013, 18, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Prasad Singh, P.; Srivastava, D.; Jaiswar, A.; Adholeya, A. Effector proteins of Rhizophagus proliferus: Conserved protein domains may play a role in host-specific interaction with different plant species. Braz. J. Microbiol. 2019, 50, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Betz, R.; Heidt, S.; Figueira-Galán, D.; Hartmann, M.; Langner, T.; Requena, N. Alternative splicing regulation in plants by SP7-like effectors from symbiotic arbuscular mycorrhizal fungi. Nat. Commun. 2024, 15, 21. [Google Scholar] [CrossRef]
- Tominaga, T.; Miura, C.; Takeda, N.; Kanno, Y.; Takemura, Y.; Seo, M.; Yamato, M.; Kaminaka, H. Gibberellin Promotes Fungal Entry and Colonization during Paris-Type Arbuscular Mycorrhizal Symbiosis in Eustoma grandiflorum. Plant Cell Physiol. 2020, 61, 565–575. [Google Scholar] [CrossRef]
- Martin-Rodriguez, J.A.; Ocampo, J.A.; Molinero-Rosales, N.; Tarkowska, D.; Ruiz-Rivero, O.; Garcia-Garrido, J.M. Role of gibberellins during arbuscular mycorrhizal formation in tomato: New insights revealed by endogenous quantification and genetic analysis of their metabolism in mycorrhizal roots. Physiol. Plant. 2015, 154, 66–81. [Google Scholar] [CrossRef]
- Foo, E.; Ross, J.J.; Jones, W.T.; Reid, J.B. Plant hormones in arbuscular mycorrhizal symbioses: An emerging role for gibberellins. Ann. Bot. 2013, 111, 769–779. [Google Scholar] [CrossRef]
- Takeda, N.; Handa, Y.; Tsuzuki, S.; Kojima, M.; Sakakibara, H.; Kawaguchi, M. Gibberellins Interfere with Symbiosis Signaling and Gene Expression and Alter Colonization by Arbuscular Mycorrhizal Fungi in Lotus japonicus. Plant Physiol. 2015, 167, 545–557. [Google Scholar] [CrossRef]
- Rai, M.; Agarkar, G. Plant-fungal interactions: What triggers the fungi to switch among lifestyles? Crit. Rev. Microbiol. 2016, 42, 428–438. [Google Scholar] [CrossRef]
- Vayssieres, A.; Pencik, A.; Felten, J.; Kohler, A.; Ljung, K.; Martin, F.; Legue, V. Development of the Poplar-Laccaria bicolor Ectomycorrhiza Modifies Root Auxin Metabolism, Signaling, and Response. Plant Physiol. 2015, 169, 890–902. [Google Scholar] [CrossRef]
- Medina, M.J.H.; Gagnon, H.; Piché, Y.; Ocampo, J.A.; Garrido, J.M.G.; Vierheilig, H. Root colonization by arbuscular mycorrhizal fungi is affected by the salicylic acid content of the plant. Plant Sci. 2003, 164, 993–998. [Google Scholar] [CrossRef]
- de Los Santos, R.T.; Vierheilig, H.; Ocampo, J.A.; Garrido, J.M.G. Altered pattern of arbuscular mycorrhizal formation in tomato ethylene mutants. Plant Signal. Behav. 2011, 6, 755–758. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, R.; Schaarschmidt, S.; Hause, B. Repeated leaf wounding alters the colonization of Medicago truncatula roots by beneficial and pathogenic microorganisms. Plant Cell Environ. 2012, 35, 1344–1357. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Medina, M.J.; Tamayo, M.I.; Vierheilig, H.; Ocampo, J.A.; Garcia-Garrido, J.M. The jasmonic acid signalling pathway restricts the development of the arbuscular mycorrhizal association in tomato. J. Plant Growth Regul. 2008, 27, 221–230. [Google Scholar] [CrossRef]
- Volpe, V.; Chialva, M.; Mazzarella, T.; Crosino, A.; Capitanio, S.; Costamagna, L.; Kohlen, W.; Genre, A. Long-lasting impact of chitooligosaccharide application on strigolactone biosynthesis and fungal accommodation promotes arbuscular mycorrhiza in Medicago truncatula. New Phytol. 2023, 237, 2316–2331. [Google Scholar] [CrossRef]
- Rozpadek, P.; Domka, A.M.; Nosek, M.; Wazny, R.; Jedrzejczyk, R.J.; Wiciarz, M.; Turnau, K. The Role of Strigolactone in the Cross-Talk Between Arabidopsis thaliana and the Endophytic Fungus Mucor sp. Front. Microbiol. 2018, 9, 441. [Google Scholar] [CrossRef]
- von Sivers, L.; Jaspar, H.; Johst, B.; Roese, M.; Bitterlich, M.; Franken, P.; Kuhn, C. Brassinosteroids Affect the Symbiosis Between the AM Fungus Rhizoglomus irregularis and Solanaceous Host Plants. Front. Plant Sci. 2019, 10, 571. [Google Scholar] [CrossRef]
- Mercy, L.; Lucic-Mercy, E.; Nogales, A.; Poghosyan, A.; Schneider, C.; Arnholdt-Schmitt, B. A Functional Approach towards Understanding the Role of the Mitochondrial Respiratory Chain in an Endomycorrhizal Symbiosis. Front. Plant Sci. 2017, 8, 417. [Google Scholar] [CrossRef]
- Tanaka, A.; Christensen, M.J.; Takemoto, D.; Park, P.; Scott, B. Reactive oxygen species play a role in regulating a fungus-perennial ryegrass mutualistic interaction. Plant Cell 2006, 18, 1052–1066. [Google Scholar] [CrossRef]
- Nagahashi, G.; Douds, D.D. The effects of hydroxy fatty acids on the hyphal branching of germinated spores of AM fungi. Fungal Biol. 2011, 115, 351–358. [Google Scholar] [CrossRef]
- Kameoka, H.; Gutjahr, C. Functions of Lipids in Development and Reproduction of Arbuscular Mycorrhizal Fungi. Plant Cell Physiol. 2022, 63, 1356–1365. [Google Scholar] [CrossRef]
- Sugiura, Y.; Akiyama, R.; Tanaka, S.; Yano, K.; Kameoka, H.; Marui, S.; Saito, M.; Kawaguchi, M.; Akiyama, K.; Saito, K. Myristate can be used as a carbon and energy source for the asymbiotic growth of arbuscular mycorrhizal fungi. Proc. Natl. Acad. Sci. USA 2020, 117, 25779–25788. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Hashimoto, K.; Kobayashi, Y.; Yano, K.; Maeda, T.; Kameoka, H.; Ezawa, T.; Saito, K.; Akiyama, K.; Kawaguchi, M. Asymbiotic mass production of the arbuscular mycorrhizal fungus Rhizophagus clarus. Commun. Biol. 2022, 5, 43. [Google Scholar] [CrossRef] [PubMed]
- Etemadi, M.; Gutjahr, C.; Couzigou, J.-M.; Zouine, M.; Lauressergues, D.; Timmers, A.; Audran, C.; Bouzayen, M.; Bécard, G.; Combier, J.-P. Auxin Perception Is Required for Arbuscule Development in Arbuscular Mycorrhizal Symbiosis. Plant Physiol. 2014, 166, 281–292. [Google Scholar] [CrossRef]
- Muller, L.M.; Harrison, M.J. Phytohormones, miRNAs, and peptide signals integrate plant phosphorus status with arbuscular mycorrhizal symbiosis. Curr. Opin. Plant Biol. 2019, 50, 132–139. [Google Scholar] [CrossRef]
- Hofferek, V.; Mendrinna, A.; Gaude, N.; Krajinski, F.; Devers, E.A. MiR171h restricts root symbioses and shows like its target NSP2 a complex transcriptional regulation in Medicago truncatula. BMC Plant Biol. 2014, 14, 199. [Google Scholar] [CrossRef]
- Couzigou, J.-M.; Lauressergues, D.; André, O.; Gutjahr, C.; Guillotin, B.; Bécard, G.; Combier, J.-P. Positive Gene Regulation by a Natural Protective miRNA Enables Arbuscular Mycorrhizal Symbiosis. Cell Host Microbe 2017, 21, 106–112. [Google Scholar] [CrossRef]
- Le Marquer, M.; Bécard, G.; Frei dit Frey, N. Arbuscular mycorrhizal fungi possess a CLAVATA3/embryo surrounding region-related gene that positively regulates symbiosis. New Phytol. 2019, 222, 1030–1042. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, S.; Li, P.L.; Yin, Y.L.; Niu, Q.L.; Yan, J.Q.; Huang, D.F. Plant buffering against the high-light stress-induced accumulation of CsGA2ox8 transcripts via alternative splicing to finely tune gibberellin levels and maintain hypocotyl elongation. Hortic. Res. 2021, 8, 10. [Google Scholar] [CrossRef]
- Miura, C.; Furui, Y.; Yamamoto, T.; Kanno, Y.; Honjo, M.; Yamaguchi, K.; Suetsugu, K.; Yagame, T.; Seo, M.; Shigenobu, S.; et al. Autoactivation of mycorrhizal symbiosis signaling through gibberellin deactivation in orchid seed germination. Plant Physiol. 2024, 194, 546–563. [Google Scholar] [CrossRef]
- Wang, Z.J.; Ji, H.T.; Yuan, B.J.; Wang, S.F.; Su, C.; Yao, B.; Zhao, H.T.; Li, X. ABA signalling is fine-tuned by antagonistic HAB1 variants. Nat. Commun. 2015, 6, 8138. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Kanwar, P.; Samtani, H.; Kaur, K.; Jha, S.K.; Pandey, G.K. Alternative Splicing of CIPK3 Results in Distinct Target Selection to Propagate ABA Signaling in Arabidopsis. Front. Plant Sci. 2017, 8, 1924. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. Abiotic Stresses Cause Differential Regulation of Alternative Splice Forms of GATA Transcription Factor in Rice. Front. Plant Sci. 2017, 8, 1944. [Google Scholar] [CrossRef] [PubMed]
- Daguerre, Y.; Basso, V.; Hartmann-Wittulski, S.; Schellenberger, R.; Meyer, L.; Bailly, J.; Kohler, A.; Plett, J.M.; Martin, F.; Veneault-Fourrey, C. The mutualism effector MiSSP7 of Laccaria bicolor alters the interactions between the poplar JAZ6 protein and its associated proteins. Sci. Rep. 2020, 10, 20362. [Google Scholar] [CrossRef]
- Takaoka, Y.; Suzuki, K.; Nozawa, A.; Takahashi, H.; Sawasaki, T.; Ueda, M. Protein-protein interactions between jasmonate-related master regulator MYC and transcriptional mediator MED25 depend on a short binding domain. J. Biol. Chem. 2022, 298, 10. [Google Scholar] [CrossRef]
- Ortu, G.; Balestrini, R.; Pereira, P.A.; Becker, J.D.; Küster, H.; Bonfante, P. Plant Genes Related to Gibberellin Biosynthesis and Signaling Are Differentially Regulated during the Early Stages of AM Fungal Interactions. Mol. Plant 2012, 5, 951–954. [Google Scholar] [CrossRef]
- Guillotin, B.; Etemadi, M.; Audran, C.; Bouzayen, M.; Bécard, G.; Combier, J.P. Sl-IAA27 regulates strigolactone biosynthesis and mycorrhization in tomato (var. MicroTom). New Phytol. 2017, 213, 1124–1132. [Google Scholar] [CrossRef]
- Liu, X.; Hu, Q.L.; Yan, J.J.; Sun, K.; Liang, Y.; Jia, M.R.; Meng, X.B.; Fang, S.; Wang, Y.Q.; Jing, Y.H.; et al. ζ-Carotene Isomerase Suppresses Tillering in Rice through the Coordinated Biosynthesis of Strigolactone and Abscisic Acid. Mol. Plant 2020, 13, 1784–1801. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, L.Y.; Ou, Q.J.; You, H.Y.; Wang, J.; Niu, J. Alternative First Exons Drive Enzymatic Activity Variation in Chalcone Synthase 3 of Dendrobium sinense. Forests 2023, 14, 1702. [Google Scholar] [CrossRef]
- Liu, J.; Miao, P.F.; Qin, W.Q.; Hu, W.; Wei, Z.Z.; Ding, W.S.; Zhang, H.; Wang, Z. A novel single nucleotide mutation of TFL1 alters the plant architecture of Gossypium arboreum through changing the pre-mRNA splicing. Plant Cell Rep. 2024, 43, 15. [Google Scholar] [CrossRef]
- Steinkellner, S.; Lendzemo, V.; Langer, I.; Schweiger, P.; Khaosaad, T.; Toussaint, J.-P.; Vierheilig, H. Flavonoids and Strigolactones in Root Exudates as Signals in Symbiotic and Pathogenic Plant-Fungus Interactions. Molecules 2007, 12, 1290–1306. [Google Scholar] [CrossRef]
- Arango, D.; Morohashi, K.; Yilmaz, A.; Kuramochi, K.; Parihar, A.; Brahimaj, B.; Grotewold, E.; Doseff, A.I. Molecular basis for the action of a dietary flavonoid revealed by the comprehensive identification of apigenin human targets. Proc. Natl. Acad. Sci. USA 2013, 110, E2153–E2162. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.L.; Du, J.E.; Li, C.L.; Li, H.Q.; Guo, H.Q.; Li, Z.Y. Kaempferol Can Reverse the 5-Fu Resistance of Colorectal Cancer Cells by Inhibiting PKM2-Mediated Glycolysis. Int. J. Mol. Sci. 2022, 23, 3544. [Google Scholar] [CrossRef] [PubMed]
- Sudhakaran, M.; Navarrete, T.G.; Mejia-Guerra, K.; Mukundi, E.; Eubank, T.D.; Grotewold, E.; Arango, D.; Doseff, A.I. Transcriptome reprogramming through alternative splicing triggered by apigenin drives cell death in triple-negative breast cancer. Cell Death Dis. 2023, 14, 10. [Google Scholar] [CrossRef]
- Ji, X.J.; Mao, X.; Hao, Q.T.; Liu, B.L.; Xue, J.A.; Li, R.Z. Splice Variants of the Castor WRI1 Gene Upregulate Fatty Acid and Oil Biosynthesis When Expressed in Tobacco Leaves. Int. J. Mol. Sci. 2018, 19, 146. [Google Scholar] [CrossRef]
- Rich, M.K.; Vigneron, N.; Libourel, C.; Keller, J.; Xue, L.; Hajheidari, M.; Radhakrishnan, G.V.; Le Ru, A.; Diop, S.I.; Potente, G.; et al. Lipid exchanges drove the evolution of mutualism during plant terrestrialization. Science 2021, 372, 864–868. [Google Scholar] [CrossRef]
- Filippovich, S.Y.; Bachurina, G.P. Nitric Oxide in Fungal Metabolism (Review). Appl. Biochem. Microbiol. 2021, 57, 694–705. [Google Scholar] [CrossRef]
- Sharina, I.G.; Martin, E. The Role of Reactive Oxygen and Nitrogen Species in the Expression and Splicing of Nitric Oxide Receptor. Antioxid. Redox Signal. 2017, 26, 122–136. [Google Scholar] [CrossRef]
- Taze, C.; Drakouli, S.; Samiotaki, M.; Panayotou, G.; Simos, G.; Georgatsou, E.; Mylonis, I. Short-term hypoxia triggers ROS and SAFB mediated nuclear matrix and mRNA splicing remodeling. Redox Biol. 2022, 58, 102545. [Google Scholar] [CrossRef]
- Zuo, D.D.; Sun, H.T.; Yang, L.; Zheng, M.L.; Zhang, J.; Guo, D.L. Hydrogen peroxide priming triggers splicing memory in grape berries. Plant Mol. Biol. 2024, 114, 129. [Google Scholar] [CrossRef]
- Hnilicová, J.; Hozeifi, S.; Dusková, E.; Icha, J.; Tománková, T.; Stanek, D. Histone Deacetylase Activity Modulates Alternative Splicing. PLoS ONE 2011, 6, e16727. [Google Scholar] [CrossRef]
- Maunakea, A.K.; Chepelev, I.; Cui, K.R.; Zhao, K.J. Intragenic DNA methylation modulates alternative splicing by recruiting MeCP2 to promote exon recognition. Cell Res. 2013, 23, 1256–1269. [Google Scholar] [CrossRef] [PubMed]
- Jabre, I.; Chaudhary, S.; Wilson, C.M.; Staiger, D.; Syed, N. Stochastic Variation in DNA Methylation Modulates Nucleosome Occupancy and Alternative Splicing in Arabidopsis thaliana. Plants 2022, 11, 1105. [Google Scholar] [CrossRef] [PubMed]
- Markus, M.A.; Marques, F.Z.; Morris, B.J. Resveratrol, by Modulating RNA Processing Factor Levels, Can Influence the Alternative Splicing of Pre-mRNAs. PLoS ONE 2011, 6, e28926. [Google Scholar] [CrossRef]
- Feng, D.R.; Cheng, Y.; Meng, Y.; Zou, L.P.; Huang, S.Z.; Xie, J.Y. Multiple effects of curcumin on promoting expression of the exon 7-containing SMN2 transcript. Genes Nutr. 2015, 10, 9. [Google Scholar] [CrossRef]
- Piombo, E.; Vetukuri, R.R.; Konakalla, N.C.; Kalyandurg, P.B.; Sundararajan, P.; Jensen, D.F.; Karlsson, M.; Dubey, M. RNA silencing is a key regulatory mechanism in the biocontrol fungus Clonostachys rosea-wheat interactions. BMC Biol. 2024, 22, 219. [Google Scholar] [CrossRef]
- Rigo, R.; Bazin, J.; Romero-Barrios, N.; Moison, M.; Lucero, L.; Christ, A.; Benhamed, M.; Blein, T.; Huguet, S.; Charon, C.; et al. The Arabidopsis lncRNA ASCO modulates the transcriptome through interaction with splicing factors. EMBO Rep. 2020, 21, e48977. [Google Scholar] [CrossRef]
- Liu, Y.; Do, S.; Huynh, H.; Li, J.-X.; Liu, Y.-G.; Du, Z.-Y.; Chen, M.-X. Importance of pre-mRNA splicing and its study tools in plants. Adv. Biotechnol. 2024, 2, 4. [Google Scholar] [CrossRef]
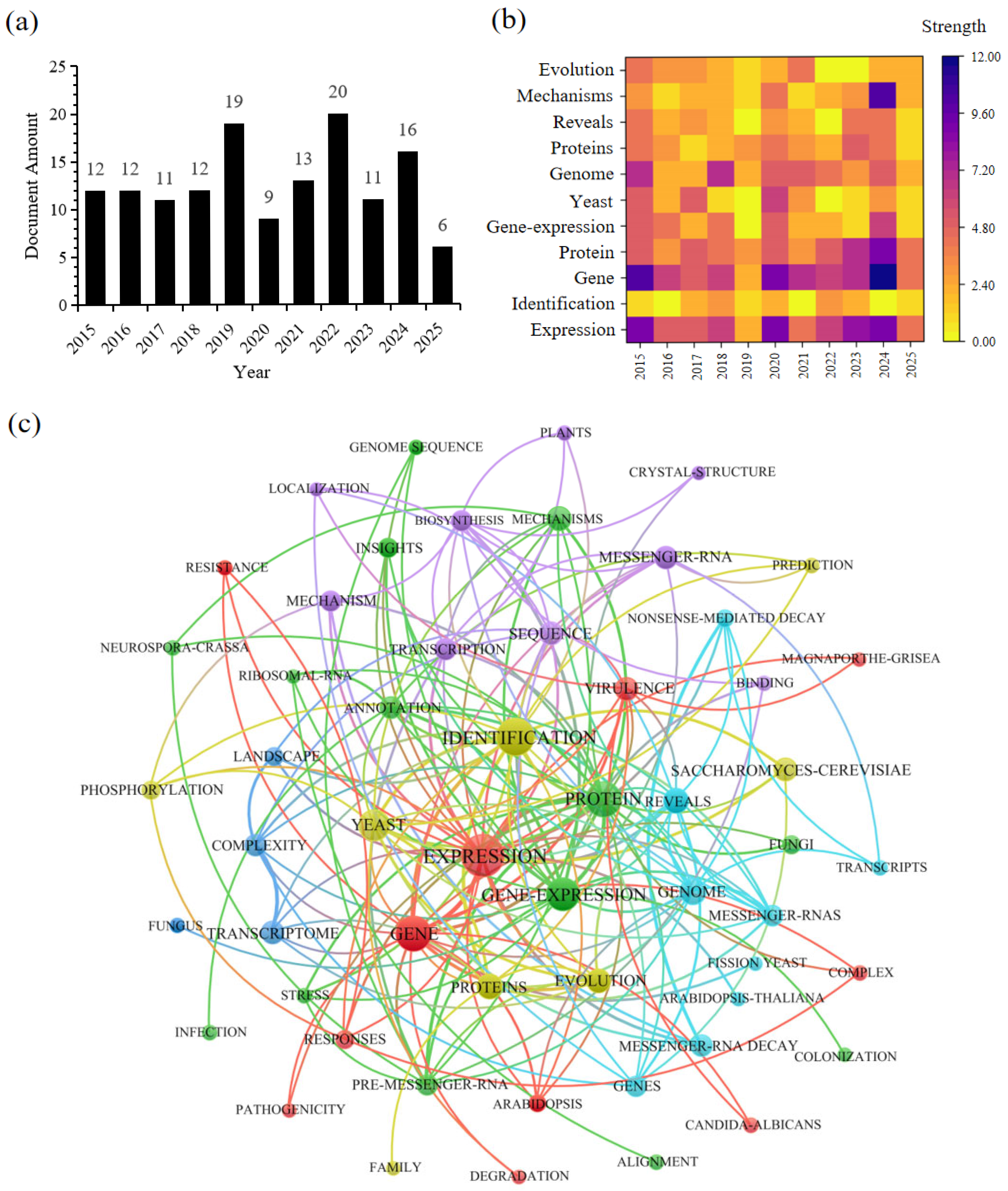

{kind=link}
{kind=link}
| Plant Species | Fungi Species | AS Detection | References |
|---|---|---|---|
| S. lycopersicum cv. | Rhizophagus irregularis | AS profiling; RNA-Seq; PacBio (Novogene (Beijing, China)); Illumina (New England Biolabs, Ipswich, MA, USA) | [14] |
| Pisum sativum L. | Rhizophagus irregularis | AS profiling; bioinformatics; DESeq2; SUPPA2; DRIMSeq; IsoformSwitchAnalyseR (http://bioconductor.org/packages/IsoformSwitchAnalyzeR, accessed on 20 March 2025) | [15] |
| Asparagus officinalis L. | Rhizophagus irregularis | AS talavista; DESeq2; Bio-Rad (Bio-Rad CFX96 real-time PCR detection system, Boulder, CO, USA) | [16] |
| Lotus japonicus | Glomus intraradices | AS profiling; gene expression profiling | [17] |
| Medicago truncatula Jemalong A17; O. sativa ssp. japonica | Rhizophagus irregularis | AS profiling; Bio-Rad; gene expression profiling | [18,19] |
| Solanum lycopersicum L. culteugenic RioGrande 76R | Glomus intraradices | AS profiling; GMAP | [20] |
| Daucus carota L. | Rhizophagus irregularis | AS profiling; Bio-Rad | [21] |
| Medicago truncatula L.; Nicotiana benthamiana | Glomus intraradices | AS profiling; gene expression analyses | [22] |
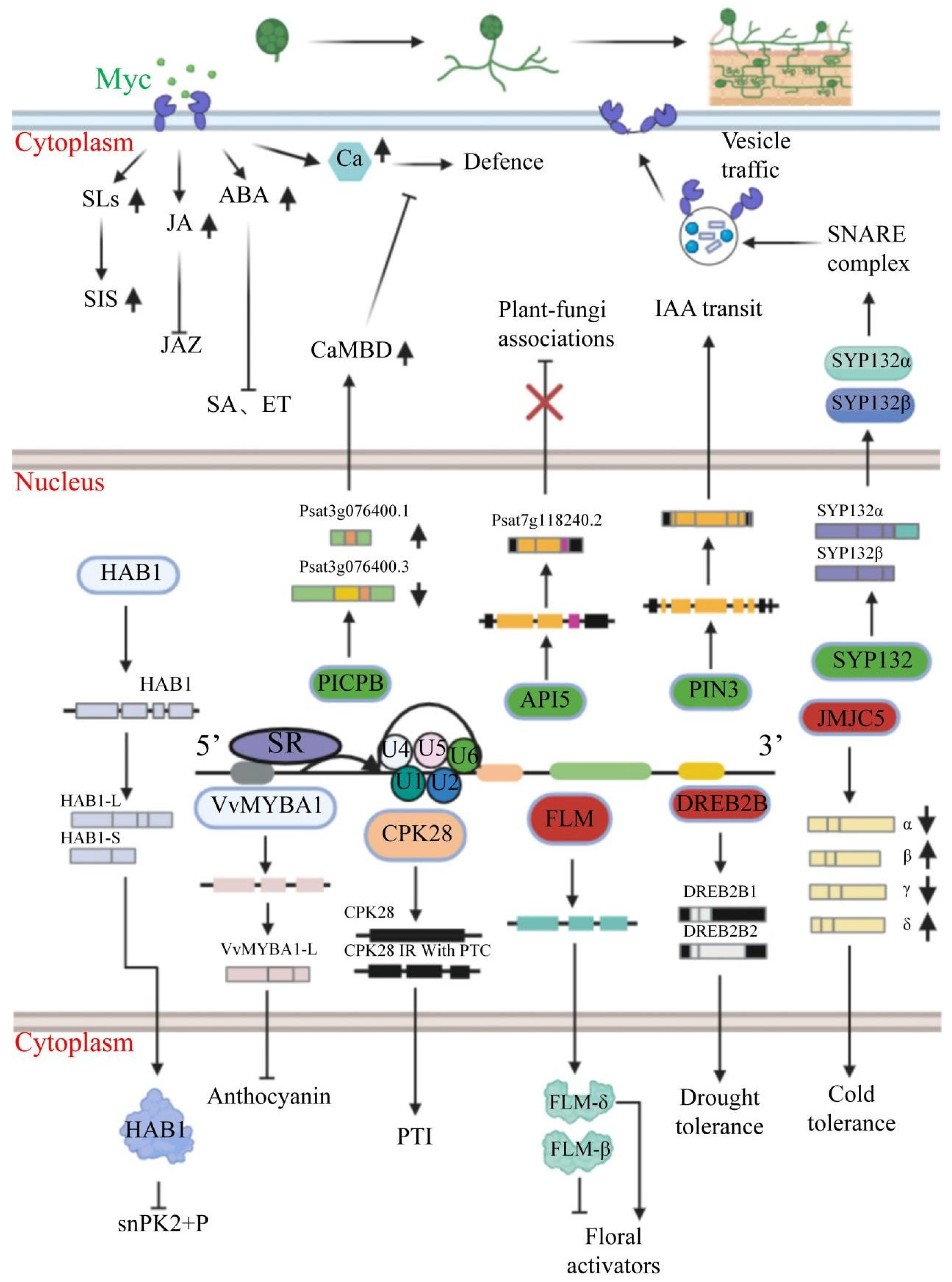
| Alternative Splicing Gene | Fungus Name | Host | Cotyledon Type | Mechanism of Modulation/Determinant | Function | References |
|---|---|---|---|---|---|---|
| Plant Effector | ||||||
| PICBP | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | AS of the PICPB gene produces transcript variants containing either two or three calmodulin-binding domains (CaMBDs); the two-CaMBD isoform exhibits upregulated expression during arbuscular mycorrhizal symbiosis. | Symbiotic signaling | [15] |
| AFC3 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | AFC3 is involved in the alternative splicing process and regulates AS by regulating the expression of splicing factors, and isoforms with PTC are increased in the mycorrhizal roots. | Symbiotic signaling | [15] |
| PIN3 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | AM symbiosis upregulates an alternatively spliced, truncated PIN3 variant that mediates phytohormone transport. | Symbiotic signaling | [15] |
| CAR11 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | In fungal symbiosis, CAR11 full-length transcripts increase, whereas truncated or NMD-targeted transcripts decrease. | Symbiotic signaling | [15] |
| ESP3 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | Longer-transcript isoforms containing HA2 or OB_NTP_bind regions are generated. | Symbiotic signaling | [15] |
| P450 | Rhizophagus irregularis | S. lycopersicum cv. | dicotyledon | P450 can produce three different isoforms through the AS process, but the expression of only one of these isoforms is upregulated in AMF colonization. | Symbiotic signaling | [14] |
| CRK25 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | AM fungi activate plant defence responses but interrupt the transduction of plant-defence-related signals by causing premature termination codons in the ORF region of CPK25 through the AS process, resulting in truncated isoforms, which are able to bind but not phosphorylate ligand signals, or degradation via NMD. | Immune regulation | [15] |
| API5 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | The AM fungi, after symbiosis with plants, produce isoforms that retain introns as well as premature stop codons in the ORF region by AS to repress the expression of the API5 gene. | Immune regulation | [15] |
| GLP2-1 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | Upregulation of isoform GLP2-1 expression occurs in response to salt stress. | Immune regulation | [16] |
| FTSH4 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | In AM symbiotic mycorrhizae, the Psat0ss1279g0480.1 isoform with more intronic fragments is upregulated, while the Psat0ss1279g0480.2 isoform is downregulated. | Immune regulation | [15] |
| NUS1 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | Isoforms are produced that retain introns as well as premature stop codons in the ORF. | Sugar transport | [15] |
| ENT3 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | During salt stress, ENT3 undergoes increased alternative splicing in mycorrhizal systems. | Substance transmembrane transport, immune regulation | [16] |
| NAC068 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | During salt stress, NAC068 undergoes enhanced alternative splicing in mycorrhizal systems. | Metabolic regulation, immune regulation, phytohormone signaling | [16] |
| AAP19-2 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | Salt stress promotes alternative splicing of the AAP19-2 gene in mycorrhizal plants. | Nutrient exchange, signaling, immune regulation | [16] |
| CBL3 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | Under saline stress, the CBL3 gene shows significantly increased alternative splicing during mycorrhizal symbiosis. | Regulation of symbiosis and immune signaling pathways | [16] |
| CML21 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | Alternative splicing of CML21 in mycorrhizae undergoes significant upregulation in response to salt stress. | Regulation of symbiosis and immune signaling pathways | [16] |
| CYP21-1 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | Upregulation of isoform CYP21-1 expression occurrs in response to salt stress. | Regulation of symbiosis and immune signaling pathways | [16] |
| ATGs | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | ATG can produce multiple isoforms via AS, and the expression of the isoforms ATG8I and ATG8C undergoes upregulation in response to salt stress. | Nutrient cycling, immune regulation, maintenance of symbiotic structural homeostasis | [16] |
| CER1 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | Alternative splicing of CER1 in mycorrhizae undergoes significant upregulation in response to salt stress. | Immune modulation, stability of symbiotic structures | [16] |
| CB5LP | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | Alternative splicing of CB5LP in mycorrhizae undergoes significant upregulation in response to salt stress. | Symbiotic signaling pathways, structural elements | [16] |
| SYP13Ⅱ | Rhizophagus irregularis-infected Allium schoenoprasum | Medicago truncatula Gaertn. | dicotyledon | SYP13II can produce two isoforms, SYP13IIα and SYP13IIβ, by alternative splicing. The SYP13IIα isoform is upregulated in AM-fungal-symbiotic plants and is involved in the transport of substances between plants and microorganisms. | Material transport, structural components | [18,19] |
| DEAH1 | Rhizophagus irregularis | Pisum sativum L. | dicotyledon | Conformational changes in the spliceosome during its catalytic cycle are orchestrated by core splicing factors. | Structural components | [15,67] |
| SF3B5 | Rhizophagus irregularis | Asparagus officinalis L. cv. | monocotyledon | A part of the U2 small nuclear ribonucleoprotein particle (snRNP), which participatesin alternative splicing, is formed. | Structural components | [16] |
| Fungus Effector | ||||||
| RiCTR3 | Rhizophagus irregularis | Daucus carota L. | dicotyledon | Mycorrhizal roots are highly expressed both RiCTR3 splice variants (RiCTR3A and RiCTR3B); the first-intron-lacking RiCTR3A isoform enhances copper tolerance under copper toxicity. | Copper ion equilibrium, reactive oxygen species (ROS) stress adaptation | [21] |
| SP7 | Glomus intraradices | Oryza sativa L. | monocotyledon | SP7 can form isoforms of different lengths, and the isoform corresponding to the longest 1.8-kb cDNA is the predominant form during the growth of plant–fungus interactions. | Immune evasion, developmental regulation, metabolic coordination | [22] |
| Treatment | Fungus Name | Host | Mechanism of Modulation/Determinant | References |
|---|---|---|---|---|
| GAs | Rhizophagus irregularis | E. grandiflorum cv. Pink Thumb | GA is involved in RAM1 expression through the CSSP signalling pathway and promotes AM fungal arbuscule formation. | [87] |
| Glomus irregularis | Solanum lycopersicum L. | GA treatment inhibits fungal infection and arbuscule development in Arum-type mycorrhizae. | [88,89,90] | |
| Paecilomyces formosus | Cucumis sativus L. | Cucumbers synthesized nonfunctional GAs that reduced salt stress impacts. | [91] | |
| IAA | Laccaria bicolor | Populus tremula × Populus alba | IAA helps ECM root development. | [92] |
| Paecilomyces formosus | Cucumis sativus L. | Enhanced IAA production in cucumber plants contributes to salt stress tolerance. | [91] | |
| SA | Glomus intraradices | Nicotiana tabacum L. | High-concentration SA treatment negatively affects fungal colonization. | [93] |
| Ethylene | Glomus clarum | Solanum lycopersicum L. | In epi plants, ethylene inhibits fungal colonization. | [94] |
| JA | Glomus intraradices | Medicago truncatula L. | JA biosynthesis promotes mycorrhization in G. intraradices. | [95] |
| Glomus intraradices | Solanum lycopersicum L. | The JA signalling pathway limits symbiotic interactions between AM fungi and plants. | [96] | |
| SLs | Funneliformis mosseae | Medicago truncatula cv. | Perception of strigolactones (SLs) by AM fungi leads to hyphal branching induction. | [97] |
| Mucor sp. | Arabidopsis thaliana L. | SL mediates symbiosis through two mechanisms: extracellular signaling to AM fungi and intrinsic regulation in plants. | [98] | |
| BRs | Rhizoglomus irregularis | Solanum lycopersicum L. | Mycorrhizal germination in plants is promoted through BR signalling. | [99] |
| ABA | R. irregulare | Solanum tuberosum L. | The stimulation of hyphal branching around spores by ABA suggests its positive effect on spore viability. | [100] |
| ROS | Epichloe festucae | Lolium perenne L. | NADPH oxidase (NoxA) inactivation mediates symbiotic transition by controlling fungal development in plant hosts. | [101] |
| 2-Hydroxytetradecanoic acid (2-OH-C14:0) | Gigaspora gigantea | Daucus carota L. | Mycelial elongation and branching are stimulated. | [102,103] |
| Myristic acid (C14:0) | R. irregularis; R. clarus HR1; G. margarita K-1 | Daucus carota L. | AMF are induced to form symbiotic spores. | [103,104,105] |
| miR393 | Rhizophagus irregularis DAOM197198 | Medicago truncatula Gaertn. | Impedes growth hormone signalling in AMF-containing host cells and affects tuft formation. | [106,107] |
| miR171h | Rhizophagus irregularis | Medicago truncatula Gaertn. | miR171h is involved in mediating the negative regulatory mechanism of NSP2 to combat AMF overcolonization. | [82,107,108] |
| miR171b | Rhizophagus irregularis | Medicago truncatula Gaertn. | LOM1 expression for mycorrhization is enhanced and protected. | [107,109] |
| RiCLE1 | R. irregularis DAOM 197198 | M. truncatula Gaertn.; Pisum sativum L. | Host root branching and AMF colonization are promoted. | [107,110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Muhammad Aslam, M.; Chen, M.; Das, D. Plant–Fungi Mutualism, Alternative Splicing, and Defense Responses: Balancing Symbiosis and Immunity. Int. J. Mol. Sci. 2025, 26, 5197. https://doi.org/10.3390/ijms26115197
Zhao X, Muhammad Aslam M, Chen M, Das D. Plant–Fungi Mutualism, Alternative Splicing, and Defense Responses: Balancing Symbiosis and Immunity. International Journal of Molecular Sciences. 2025; 26(11):5197. https://doi.org/10.3390/ijms26115197
Chicago/Turabian StyleZhao, Xiaoqiong, Mehtab Muhammad Aslam, Moxian Chen, and Debatosh Das. 2025. "Plant–Fungi Mutualism, Alternative Splicing, and Defense Responses: Balancing Symbiosis and Immunity" International Journal of Molecular Sciences 26, no. 11: 5197. https://doi.org/10.3390/ijms26115197
APA StyleZhao, X., Muhammad Aslam, M., Chen, M., & Das, D. (2025). Plant–Fungi Mutualism, Alternative Splicing, and Defense Responses: Balancing Symbiosis and Immunity. International Journal of Molecular Sciences, 26(11), 5197. https://doi.org/10.3390/ijms26115197