
BACE1 Inhibition Protects Against Type 2 Diabetes Mellitus by Restoring Insulin Receptor in Mice

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
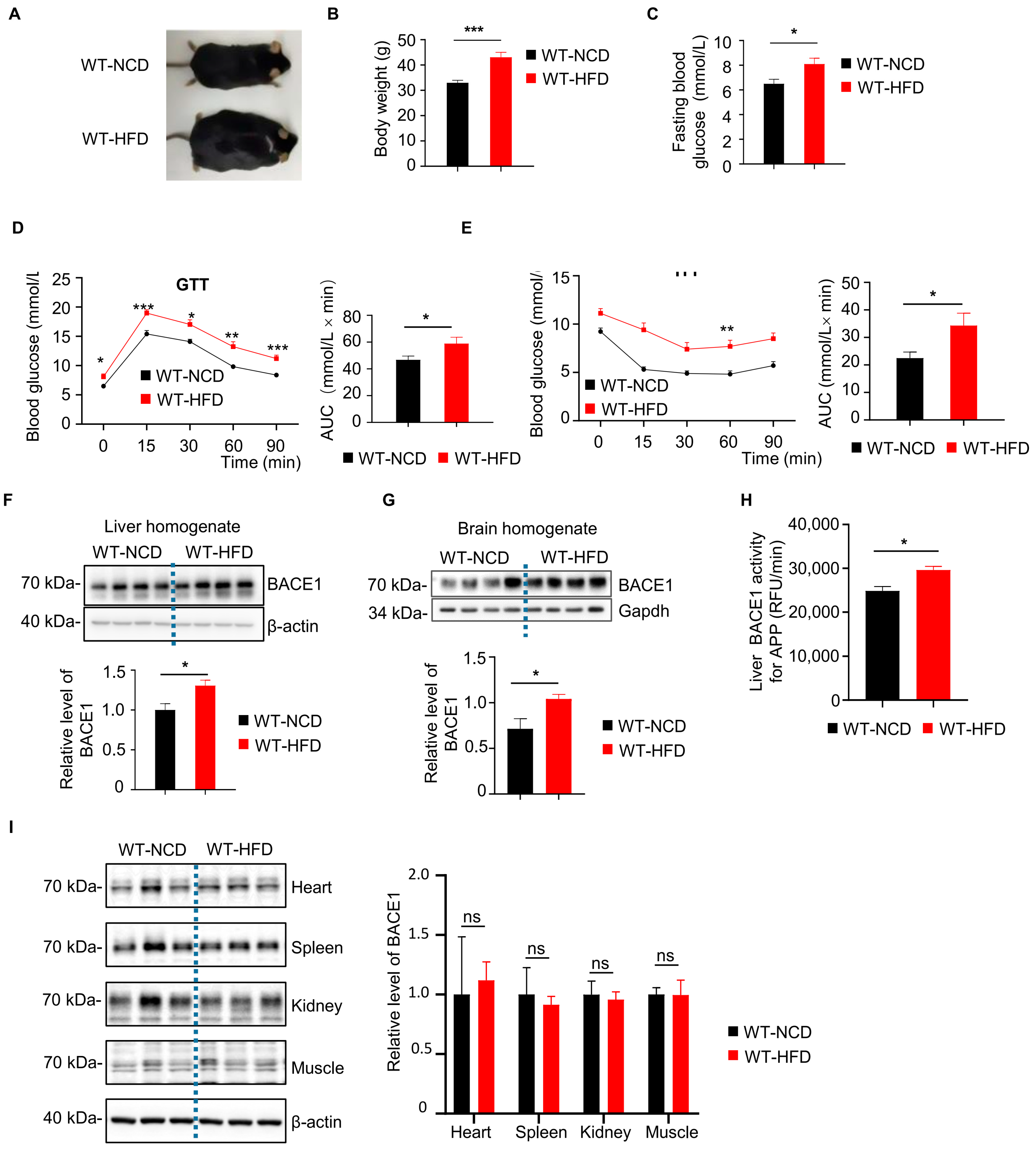
2.1. Elevated BACE1 Protein Levels and Enzyme Activity in Liver and Brain Form WT-HFD Mice
2.2. BACE1 Overexpression Caused Reduced Liver InsR Level and Diabetic Phenotypes in HUBC Mice
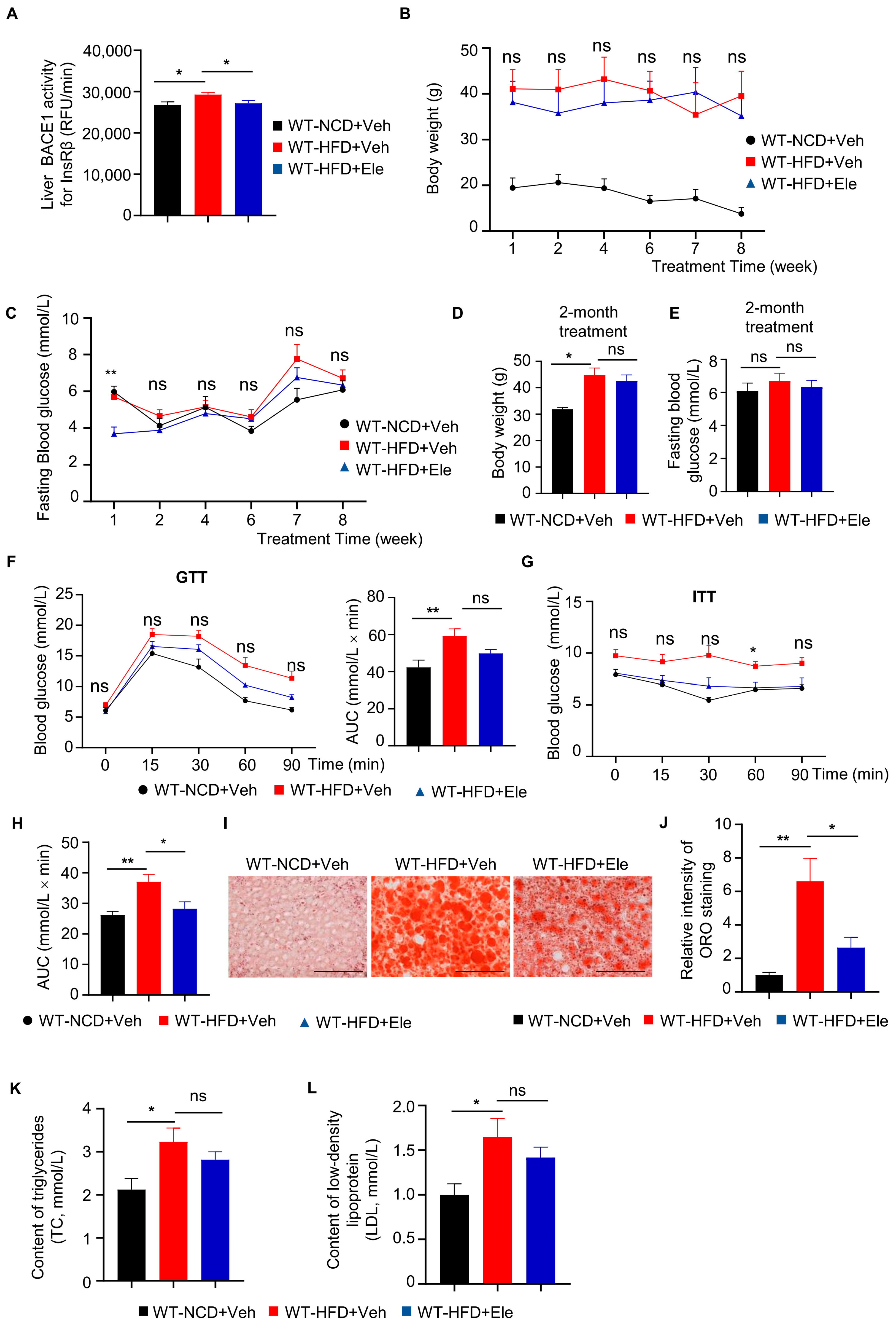
2.3. Elenbecestat Treatment Improved Glucose Tolerance and Insulin Sensitivity in HFD Mice
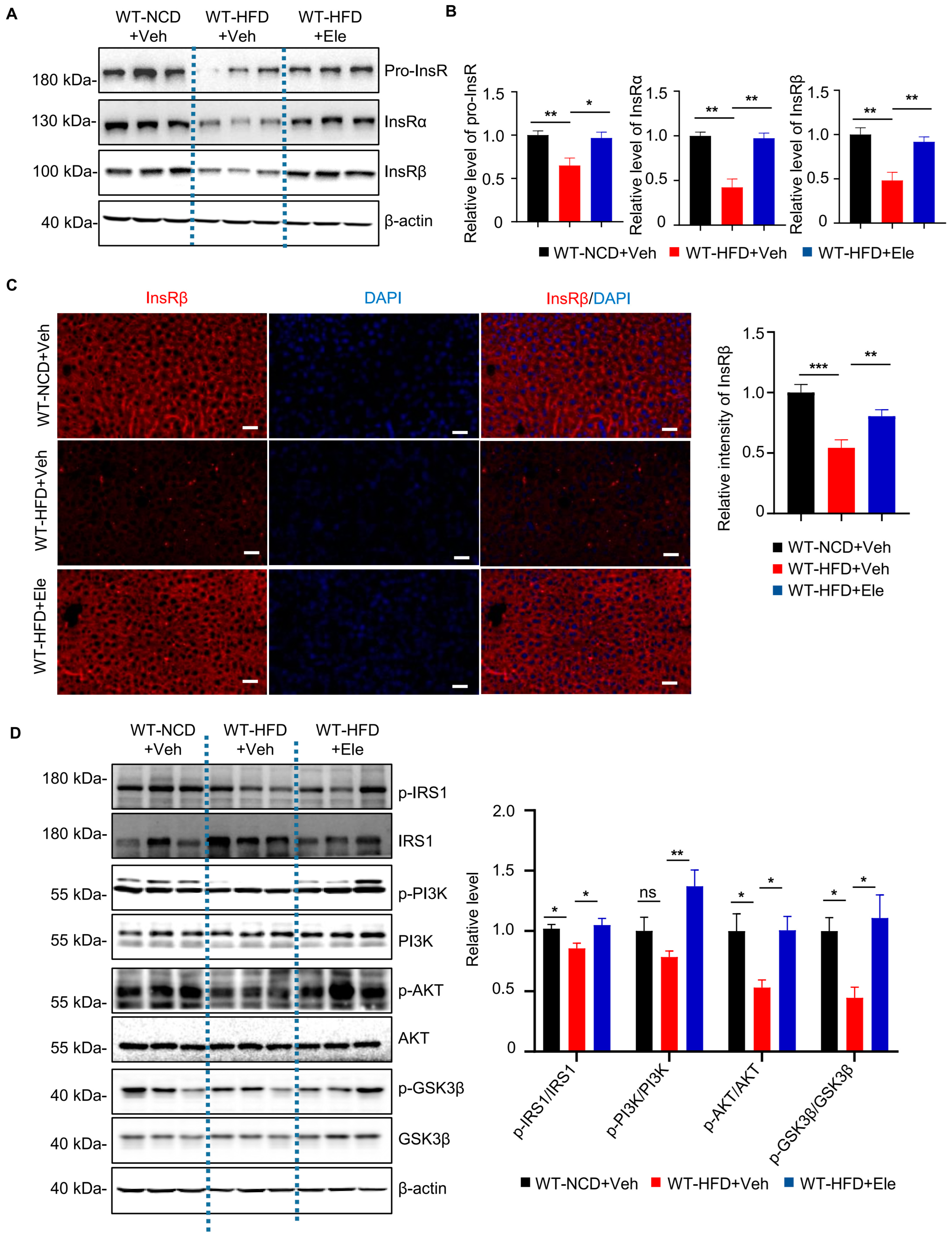
2.4. Elenbecestat Treatment Restored Hepatic Insulin Receptor Levels and Its Downstream Pathway in HFD Mice
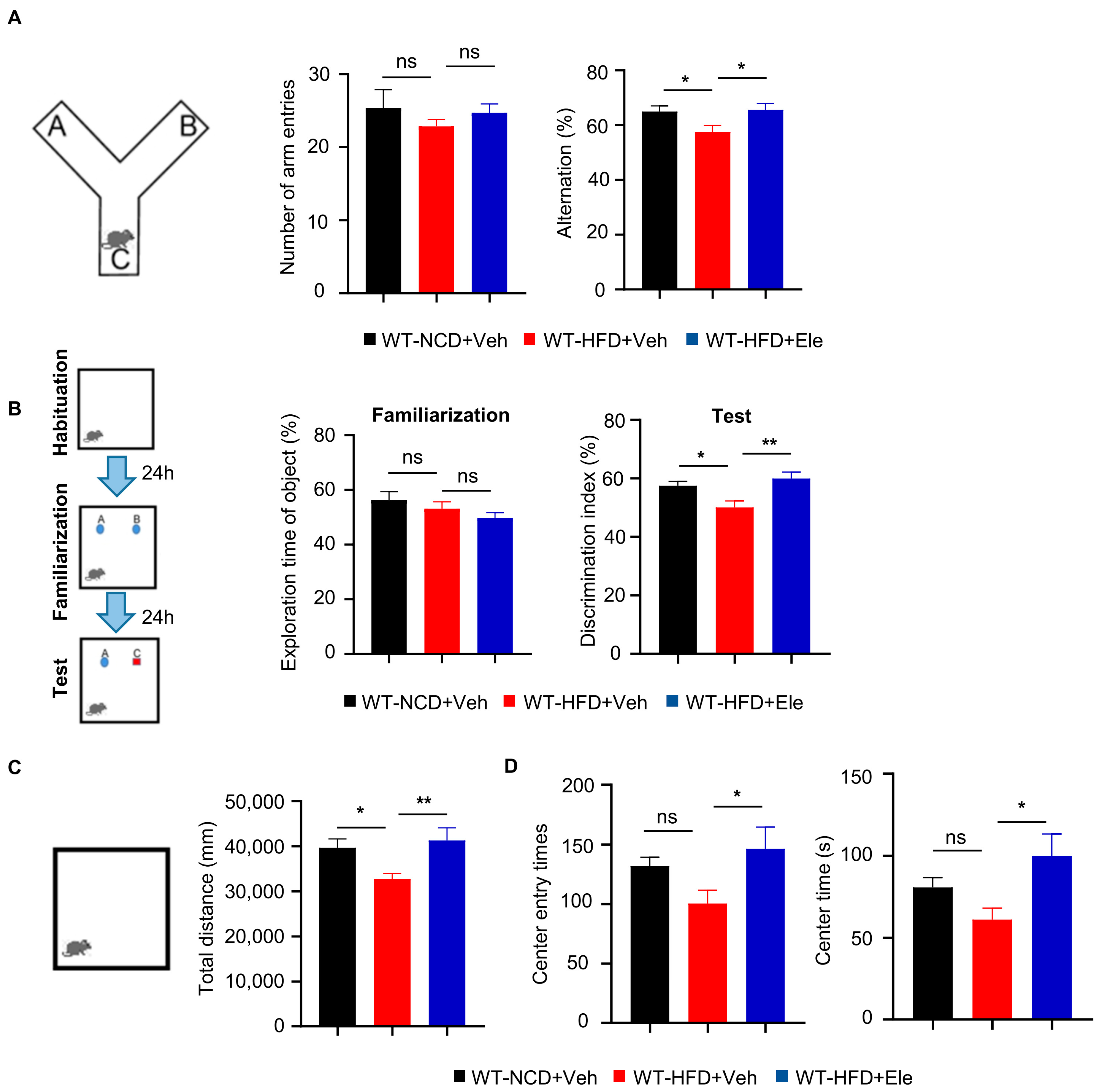
2.5. Elenbecestat Treatment Ameliorated Cognitive Impairment and Anxiety Behavior in HFD Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. High-Fat Diet Treatments
4.3. Drug Treatments
4.4. Metabolic Measurements
4.5. Western Blotting
4.6. Oil Red O (ORO) Staining
4.7. BACE1 Enzyme Activity Assay
4.8. Immunofluorescence Staining
4.9. Serum Biochemical Analysis
4.10. Behavior Tests
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BACE1 | Β-secretase 1 |
| AD | Alzheimer’s disease |
| T2DM | Type 2 diabetes mellitus |
| HFD | High-fat diet |
| HUBC | Systemic overexpression of bace1 |
| InsR | Insulin receptor |
| App | Amyloid precursor protein |
| WT | Wild-type |
| Ele | Elenbecestat |
| NCD | Normal chow diet |
| GTT | Glucose tolerance test |
| ITT | Insulin tolerance test |
| ORO | Oil Red O |
| BCA | Bicinchonic acid |
| NGS | Normal goat serum |
| LDL | Low-density lipoprotein |
| TG | Triglycerides |
| AUC | Area under the curve |
| WAT | White adipose tissue |
| ns | No significant differences |
| HDL | High-density lipoprotein |
References
- GBD 2021 Diabetes Collaborators. Global, regional, and national burden of diabetes from 1990 to 2021, with projections of prevalence to 2050: A systematic analysis for the Global Burden of Disease Study 2021. Lancet 2023, 402, 203–234. [Google Scholar] [CrossRef] [PubMed]
- Salunkhe, V.A.; Veluthakal, R.; Kahn, S.E.; Thurmond, D.C. Novel approaches to restore beta cell function in prediabetes and type 2 diabetes. Diabetologia 2018, 61, 1895–1901. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Kuroko, M.; Sekine, S.; Matsui, S.; Kikuchi, O.; Susanti, V.Y.; Kobayashi, M.; Tanaka, Y.; Yuasa, T.; Kitamura, T. Overexpression of insulin receptor partially improves obese and diabetic phenotypes in db/db mice. Endocr. J. 2015, 62, 787–796. [Google Scholar] [CrossRef]
- Muniroh, L.; Mahmudah; Solfaine, R. Effect of Tithonia diversifolia Leaf Extract on Leptin, Adiponectin, and Insulin Receptor Levels in Diabetic Rats. Prev. Nutr. Food Sci. 2022, 27, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef]
- Yang, Q.; Vijayakumar, A.; Kahn, B.B. Metabolites as regulators of insulin sensitivity and metabolism. Nat. Rev. Mol. Cell Biol. 2018, 19, 654–672. [Google Scholar] [CrossRef]
- Hopkins, B.D.; Goncalves, M.D.; Cantley, L.C. Insulin-PI3K signalling: An evolutionarily insulated metabolic driver of cancer. Nat. Rev. Endocrinol. 2020, 16, 276–283. [Google Scholar] [CrossRef]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef]
- Batista, T.M.; Haider, N.; Kahn, C.R. Defining the underlying defect in insulin action in type 2 diabetes. Diabetologia 2021, 64, 994–1006. [Google Scholar] [CrossRef]
- Saltiel, A.R. Insulin signaling in health and disease. J. Clin. Investig. 2021, 131, e142241. [Google Scholar] [CrossRef]
- Kolterman, O.G.; Gray, R.S.; Griffin, J.; Burstein, P.; Insel, J.; Scarlett, J.A.; Olefsky, J.M. Receptor and postreceptor defects contribute to the insulin resistance in noninsulin-dependent diabetes mellitus. J. Clin. Investig. 1981, 68, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Brierley, G.V.; Webber, H.; Rasijeff, E.; Grocott, S.; Siddle, K.; Semple, R.K. Anti-Insulin Receptor Antibodies Improve Hyperglycemia in a Mouse Model of Human Insulin Receptoropathy. Diabetes 2020, 69, 2481–2489. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, H.; Palyha, O.; Mu, J. Restoration of insulin receptor improves diabetic phenotype in T2DM mice. JCI Insight 2019, 4, e124945. [Google Scholar] [CrossRef]
- Brüning, J.C.; Michael, M.; Winnay, J.N.; Hayashi, T.; Hörsch, D.; Accili, D.; Goodyear, L.J.; Kahn, C. A muscle-specific insulin receptor knockout exhibits features of the metabolic syndrome of NIDDM without altering glucose tolerance. Mol. Cell 1998, 2, 559–569. [Google Scholar] [CrossRef]
- Vassar, R. BACE1: The beta-secretase enzyme in Alzheimer’s disease. J. Mol. Neurosci. 2004, 23, 105–114. [Google Scholar] [CrossRef]
- Zhao, W.Q.; Townsend, M. Insulin resistance and amyloidogenesis as common molecular foundation for type 2 diabetes and Alzheimer’s disease. Biochim. Biophys. Acta 2009, 1792, 482–496. [Google Scholar] [CrossRef]
- Meakin, P.J.; Mezzapesa, A.; Benabou, E.; Haas, M.E.; Bonardo, B.; Grino, M.; Brunel, J.-M.; Desbois-Mouthon, C.; Biddinger, S.B.; Govers, R.; et al. The beta secretase BACE1 regulates the expression of insulin receptor in the liver. Nat. Commun. 2018, 9, 1306. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Liu, Y.; Zhang, M.; Chen, Z.; Zhang, W.; Ge, Y.; Kang, D.; Gao, F.; Shen, Y. Increased β-site APP cleaving enzyme 1-mediated insulin receptor cleavage in type 2 diabetes mellitus with cognitive impairment. Alzheimers Dement. 2021, 17, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Plucińska, K.; Dekeryte, R.; Koss, D.; Shearer, K.; Mody, N.; Whitfield, P.D.; Doherty, M.K.; Mingarelli, M.; Welch, A.; Riedel, G.; et al. Neuronal human BACE1 knockin induces systemic diabetes in mice. Diabetologia 2016, 59, 1513–1523. [Google Scholar] [CrossRef]
- Hamilton, D.L.; Findlay, J.A.; Montagut, G.; Meakin, P.J.; Bestow, D.; Jalicy, S.M.; Ashford, M.L.J. Altered amyloid precursor protein processing regulates glucose uptake and oxidation in cultured rodent myotubes. Diabetologia 2014, 57, 1684–1692. [Google Scholar] [CrossRef]
- Meakin, P.J.; Harper, A.J.; Hamilton, D.L.; Gallagher, J.; McNeilly, A.D.; Burgess, L.A.; Vaanholt, L.M.; Bannon, K.A.; Latcham, J.; Hussain, I.; et al. Reduction in BACE1 decreases body weight, protects against diet-induced obesity and enhances insulin sensitivity in mice. Biochem. J. 2012, 441, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://beta.clinicaltrials.gov/study/NCT02322021 (accessed on 17 April 2025).
- Available online: https://www.janssen.com/neuroscience/update-janssens-bace-inhibitor-program (accessed on 17 April 2025).
- En Li Cho, E.; Ang, C.Z.; Quek, J.; Fu, C.E.; Lim, L.K.E.; Heng, Z.E.Q.; Tan, D.J.H.; Lim, W.H.; Yong, J.N.; Zeng, R.; et al. Global prevalence of non-alcoholic fatty liver disease in type 2 diabetes mellitus: An updated systematic review and meta-analysis. Gut 2023, 72, 2138–2148. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, J.M.; Baptista, F.I.; Macedo, M.P.; Ambrósio, A.F. Inside the Diabetic Brain: Role of Different Players Involved in Cognitive Decline. ACS Chem. Neurosci. 2016, 7, 131–142. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, N.-Q.; Yan, F.; Jin, H.; Zhou, S.-Y.; Shi, J.-S.; Jin, F. Diabetes mellitus and Alzheimer’s disease: GSK-3β as a potential link. Behav. Brain Res. 2018, 339, 57–65. [Google Scholar] [CrossRef]
- Dutta, B.J.; Singh, S.; Seksaria, S.; Das Gupta, G.; Singh, A. Inside the diabetic brain: Insulin resistance and molecular mechanism associated with cognitive impairment and its possible therapeutic strategies. Pharmacol. Res. 2022, 182, 106358. [Google Scholar] [CrossRef]
- Ogrodnik, M.; Zhu, Y.; Langhi, L.G.; Tchkonia, T.; Krüger, P.; Fielder, E.; Victorelli, S.; Ruswhandi, R.A.; Giorgadze, N.; Pirtskhalava, T.; et al. Obesity-Induced Cellular Senescence Drives Anxiety and Impairs Neurogenesis. Cell Metab. 2019, 29, 1061–1077.e8. [Google Scholar] [CrossRef]
- Moussa-Pacha, N.M.; Abdin, S.M.; Omar, H.A.; Alniss, H.; Al-Tel, T.H. BACE1 inhibitors: Current status and future directions in treating Alzheimer’s disease. Med. Res. Rev. 2020, 40, 339–384. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Gómez, N.; Valdes-Gonzalez, M.; Garrido-Suárez, B.B.; Garrido, G. Pharmacological Inventions for Alzheimer Treatment in the United States of America: A Revision Patent from 2010–2020. J. Prev. Alzheimers Dis. 2023, 10, 50–68. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gao, F.; Yang, X.; Lin, T.; Li, Z.; Wang, Q.; Yao, Y.; Li, L.; Ding, X.; Shi, K.; et al. Endothelial BACE1 Impairs Cerebral Small Vessels via Tight Junctions and eNOS. Circ. Res. 2022, 130, 1321–1341. [Google Scholar] [CrossRef]
- Meakin, P.J.; Coull, B.M.; Tuharska, Z.; McCaffery, C.; Akoumianakis, I.; Antoniades, C.; Brown, J.; Griffin, K.J.; Platt, F.; Ozber, C.H.; et al. Elevated circulating amyloid concentrations in obesity and diabetes promote vascular dysfunction. J. Clin. Investig. 2020, 130, 4104–4117. [Google Scholar] [CrossRef]
- Arnold, S.E.; Arvanitakis, Z.; Macauley-Rambach, S.L.; Koenig, A.M.; Wang, H.-Y.; Ahima, R.S.; Craft, S.; Gandy, S.; Buettner, C.; Stoeckel, L.E.; et al. Brain insulin resistance in type 2 diabetes and Alzheimer disease: Concepts and conundrums. Nat. Rev. Neurol. 2018, 14, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Sung, P.-S.; Li, Y.-R.; Huang, W.-K.; Lee, T.-W.; Huang, C.-C.; Lee, T.-H.; Chen, T.-H.; Wei, Y.-C. Telmisartan use and risk of dementia in type 2 diabetes patients with hypertension: A population-based cohort study. PLoS Med. 2021, 18, e1003707. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, M.; Bourassa, P.; Tremblay, C.; Caron, V.; Sugère, C.; Emond, V.; A Bennett, D.; Calon, F. Cerebrovascular insulin receptors are defective in Alzheimer’s disease. Brain 2023, 146, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Heni, M.; Kullmann, S.; Preissl, H.; Fritsche, A.; Häring, H.-U. Impaired insulin action in the human brain: Causes and metabolic consequences. Nat. Rev. Endocrinol. 2015, 11, 701–711. [Google Scholar] [CrossRef]
- Talbot, K.; Wang, H.-Y.; Kazi, H.; Han, L.-Y.; Bakshi, K.P.; Stucky, A.; Fuino, R.L.; Kawaguchi, K.R.; Samoyedny, A.J.; Wilson, R.S.; et al. Demonstrated brain insulin resistance in Alzheimer’s disease patients is associated with IGF-1 resistance, IRS-1 dysregulation, and cognitive decline. J. Clin. Investig. 2012, 122, 1316–1338. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, T.; Liang, T.; Shen, Y.; Gao, F. BACE1 Inhibition Protects Against Type 2 Diabetes Mellitus by Restoring Insulin Receptor in Mice. Int. J. Mol. Sci. 2025, 26, 5100. https://doi.org/10.3390/ijms26115100
Lin T, Liang T, Shen Y, Gao F. BACE1 Inhibition Protects Against Type 2 Diabetes Mellitus by Restoring Insulin Receptor in Mice. International Journal of Molecular Sciences. 2025; 26(11):5100. https://doi.org/10.3390/ijms26115100
Chicago/Turabian StyleLin, Tingting, Ting Liang, Yong Shen, and Feng Gao. 2025. "BACE1 Inhibition Protects Against Type 2 Diabetes Mellitus by Restoring Insulin Receptor in Mice" International Journal of Molecular Sciences 26, no. 11: 5100. https://doi.org/10.3390/ijms26115100
APA StyleLin, T., Liang, T., Shen, Y., & Gao, F. (2025). BACE1 Inhibition Protects Against Type 2 Diabetes Mellitus by Restoring Insulin Receptor in Mice. International Journal of Molecular Sciences, 26(11), 5100. https://doi.org/10.3390/ijms26115100