
A New Method of Canine CD4+ T Lymphocyte Differentiation Towards the Th17 Phenotype with Analysis of Properties and Mitochondrial Activity

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
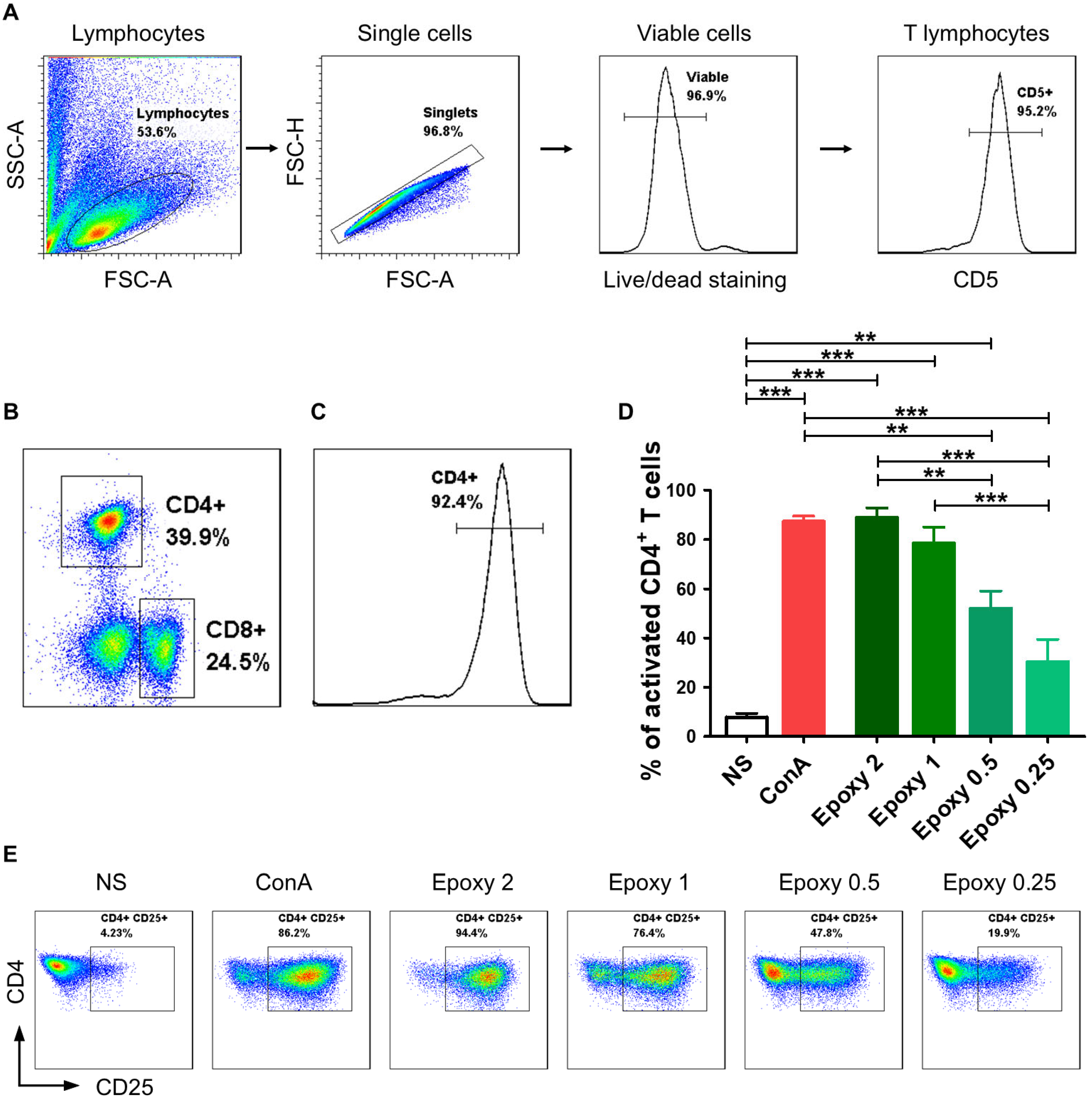
2.1. CD4+ T Cells Isolated from Canine Peripheral Blood Are Effectively Activated by Epoxylated Magnetic Beads
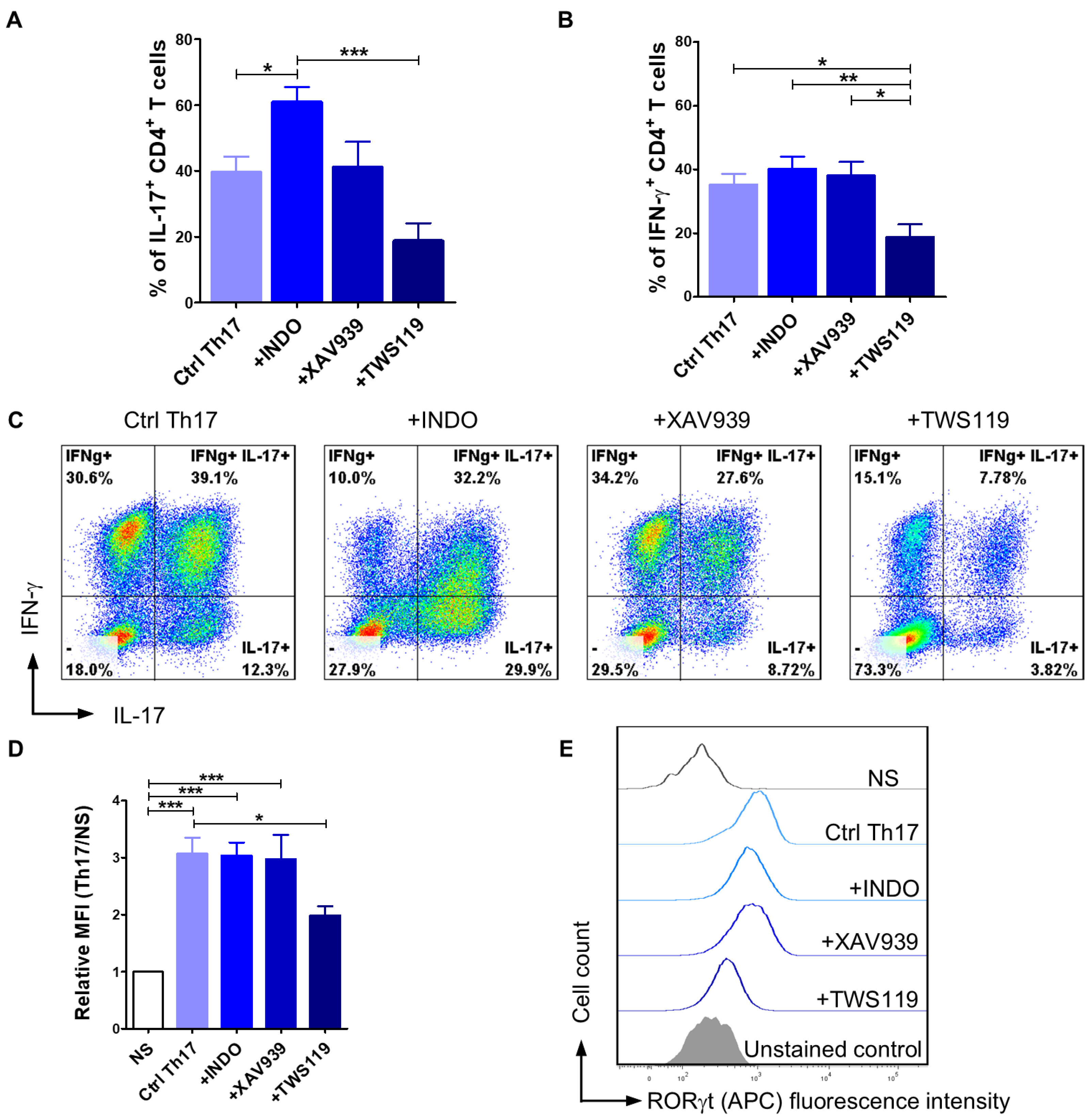
2.2. Differentiation of Activated T Lymphocytes Towards the Th17 Phenotype Can Be Achieved with the Appropriate Programming Medium
2.3. The Application of Wnt/β-Catenin Signaling Pathway Modulators Affects the Efficiency of CD4+ T Lymphocyte Programming Towards the Th17 Phenotype
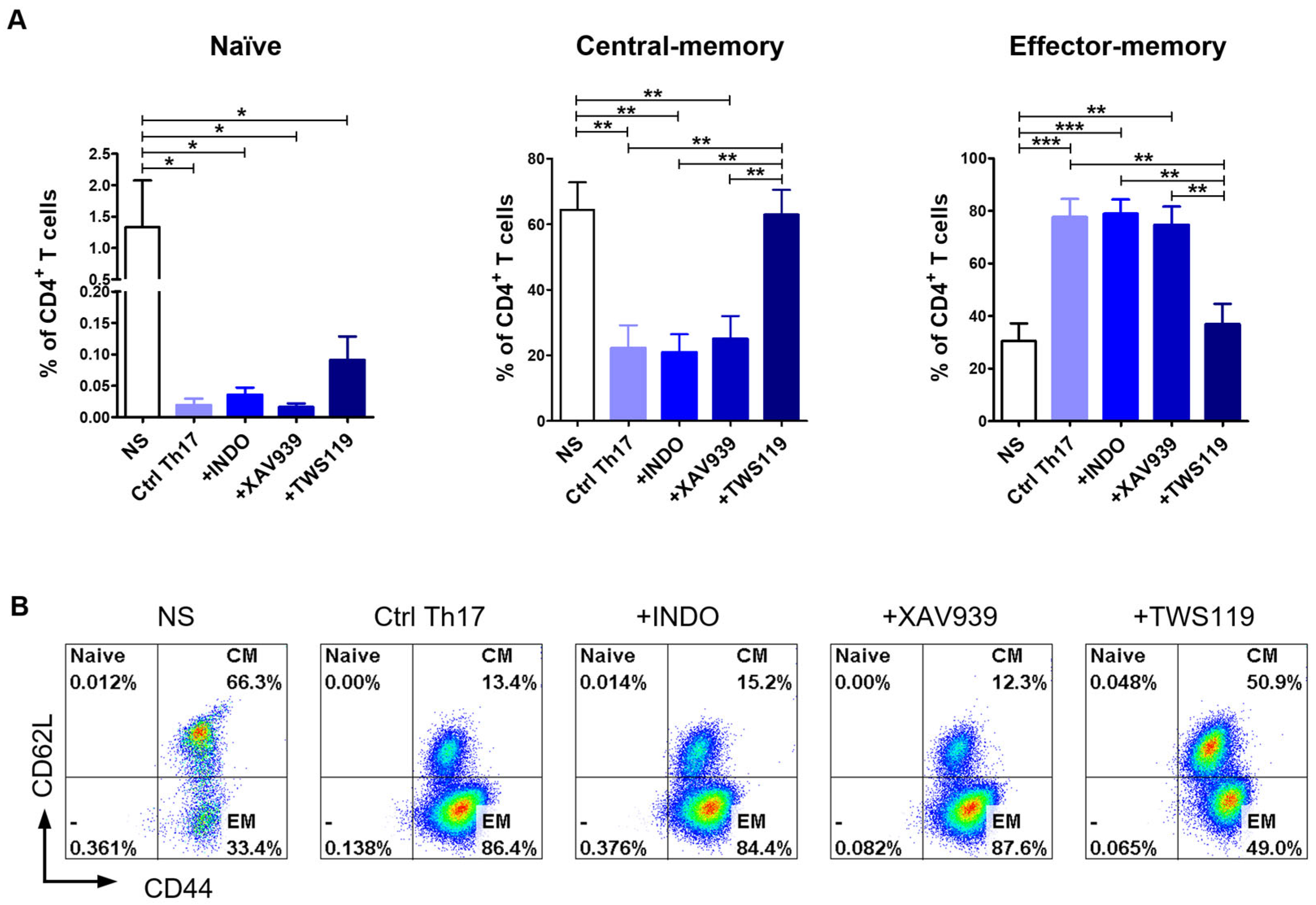
2.4. Th17 Polarization Promotes the Acquiring of an Effector-Memory T Cell Phenotype
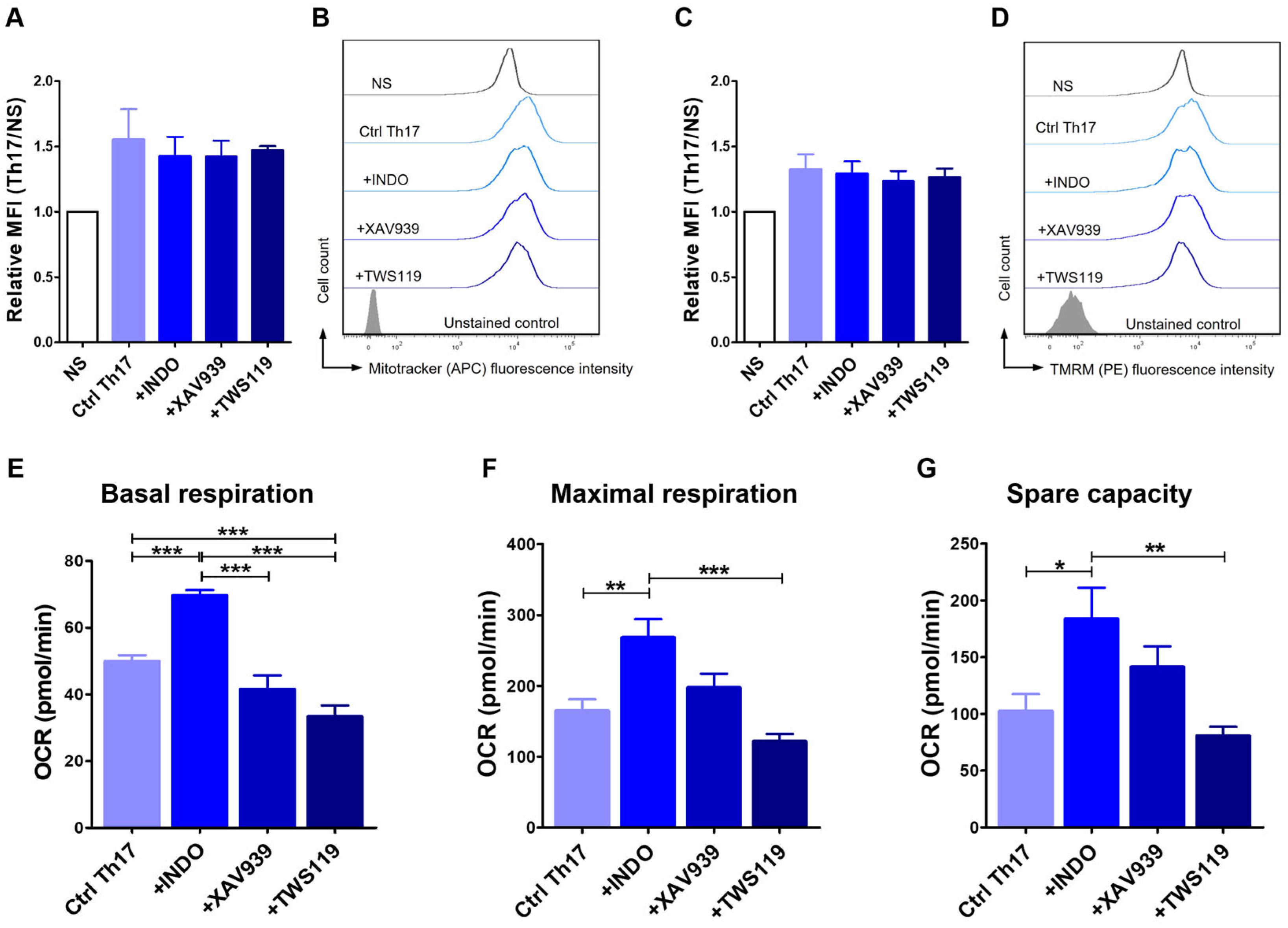
2.5. Differentiation Towards the Th17 Phenotype Involves Changes in Mitochondrial Activity
3. Discussions
4. Materials and Methods
4.1. Isolation of Canine Peripheral Blood CD4+ T Lymphocytes
4.2. Activation and Differentiation of CD4+ T Cells
4.3. Flow Cytometry Analysis
4.4. Flow Cytometric Analysis of Cytokines and Transcription Factors Characteristic of Th17 Cells
4.5. Seahorse Assay
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.-H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A Distinct Lineage of CD4 T Cells Regulates Tissue Inflammation by Producing Interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Noack, M.; Miossec, P. Th17 and Regulatory T Cell Balance in Autoimmune and Inflammatory Diseases. Autoimmun. Rev. 2014, 13, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Maddur, M.S.; Miossec, P.; Kaveri, S.V.; Bayry, J. Th17 Cells. Am. J. Pathol. 2012, 181, 8–18. [Google Scholar] [CrossRef]
- Mensikova, M.; Stepanova, H.; Faldyna, M. Interleukin-17 in Veterinary Animal Species and Its Role in Various Diseases: A Review. Cytokine 2013, 64, 11–17. [Google Scholar] [CrossRef]
- Martinez, N.E.; Sato, F.; Kawai, E.; Omura, S.; Chervenak, R.P.; Tsunoda, I. Regulatory T Cells and Th17 Cells in Viral Infections: Implications for Multiple Sclerosis and Myocarditis. Future Virol. 2012, 7, 593–608. [Google Scholar] [CrossRef]
- Ouyang, W.; Kolls, J.K.; Zheng, Y. The Biological Functions of T Helper 17 Cell Effector Cytokines in Inflammation. Immunity 2008, 28, 454–467. [Google Scholar] [CrossRef]
- Marques, H.S.; de Brito, B.B.; da Silva, F.A.F.; Santos, M.L.C.; de Souza, J.C.B.; Correia, T.M.L.; Lopes, L.W.; Neres, N.S.d.M.; Dórea, R.S.D.M.; Dantas, A.C.S.; et al. Relationship between Th17 Immune Response and Cancer. World J. Clin. Oncol. 2021, 12, 845–867. [Google Scholar] [CrossRef] [PubMed]
- Moser, T.; Akgün, K.; Proschmann, U.; Sellner, J.; Ziemssen, T. The Role of TH17 Cells in Multiple Sclerosis: Therapeutic Implications. Autoimmun. Rev. 2020, 19, 102647. [Google Scholar] [CrossRef]
- Lubberts, E. The IL-23–IL-17 Axis in Inflammatory Arthritis. Nat. Rev. Rheumatol. 2015, 11, 415–429. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Carding, S.R. Inflammatory Bowel Disease: Cause and Immunobiology. Lancet 2007, 369, 1627–1640. [Google Scholar] [CrossRef]
- Martin, J.C.; Baeten, D.L.; Josien, R. Emerging Role of IL-17 and Th17 Cells in Systemic Lupus Erythematosus. Clin. Immunol. 2014, 154, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, X.; Zhu, Q.; Wang, M.; Xie, Z.; Zhao, T. Imbalance of Th17 Cells, Treg Cells and Associated Cytokines in Patients with Systemic Lupus Erythematosus: A Meta-Analysis. Front. Immunol. 2024, 15, 1425847. [Google Scholar] [CrossRef]
- Marinoni, B.; Ceribelli, A.; Massarotti, M.S.; Selmi, C. The Th17 Axis in Psoriatic Disease: Pathogenetic and Therapeutic Implications. Autoimmun. Highlights 2014, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Newcomb, D.C.; Peebles, R.S. Th17-Mediated Inflammation in Asthma. Curr. Opin. Immunol. 2013, 25, 755–760. [Google Scholar] [CrossRef]
- Tamamoto-Mochizuki, C.; Santoro, D.; Saridomikelakis, M.N.; Eisenschenk, M.N.C.; Hensel, P.; Pucheu-Haston, C.; International Committee on Allergic Diseases of Animals (ICADA). Update on the Role of Cytokines and Chemokines in Canine Atopic Dermatitis. Vet. Dermatol. 2024, 35, 25–39. [Google Scholar] [CrossRef]
- Marsella, R. Advances in Our Understanding of Canine Atopic Dermatitis. Vet. Dermatol. 2021, 32, 547. [Google Scholar] [CrossRef] [PubMed]
- Freundt-Revilla, J.; Maiolini, A.; Carlson, R.; Beyerbach, M.; Rentmeister, K.; Flegel, T.; Fischer, A.; Tipold, A. Th17-Skewed Immune Response and Cluster of Differentiation 40 Ligand Expression in Canine Steroid-Responsive Meningitis-Arteritis, a Large Animal Model for Neutrophilic Meningitis. J. Neuroinflamm. 2017, 14, 20. [Google Scholar] [CrossRef]
- Barber, R.; Barber, J. Differential T-Cell Responses in Dogs with Meningoencephalomyelitis of Unknown Origin Compared to Healthy Controls. Front. Vet. Sci. 2022, 9, 925770. [Google Scholar] [CrossRef]
- Muranski, P.; Boni, A.; Antony, P.A.; Cassard, L.; Irvine, K.R.; Kaiser, A.; Paulos, C.M.; Palmer, D.C.; Touloukian, C.E.; Ptak, K.; et al. Tumor-Specific Th17-Polarized Cells Eradicate Large Established Melanoma. Blood 2008, 112, 362–373. [Google Scholar] [CrossRef]
- Muranski, P.; Borman, Z.A.; Kerkar, S.P.; Klebanoff, C.A.; Ji, Y.; Sanchez-Perez, L.; Sukumar, M.; Reger, R.N.; Yu, Z.; Kern, S.J.; et al. Th17 Cells Are Long Lived and Retain a Stem Cell-like Molecular Signature. Immunity 2011, 35, 972–985. [Google Scholar] [CrossRef]
- Martin-Orozco, N.; Muranski, P.; Chung, Y.; Yang, X.O.; Yamazaki, T.; Lu, S.; Hwu, P.; Restifo, N.P.; Overwijk, W.W.; Dong, C. T Helper 17 Cells Promote Cytotoxic T Cell Activation in Tumor Immunity. Immunity 2009, 31, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Kryczek, I.; Zhao, E.; Liu, Y.; Wang, Y.; Vatan, L.; Szeliga, W.; Moyer, J.; Klimczak, A.; Lange, A.; Zou, W. Human TH17 Cells Are Long-Lived Effector Memory Cells. Sci. Transl. Med. 2011, 3, 104ra100. [Google Scholar] [CrossRef] [PubMed]
- Bowers, J.S.; Nelson, M.H.; Majchrzak, K.; Bailey, S.R.; Rohrer, B.; Kaiser, A.D.; Atkinson, C.; Gattinoni, L.; Paulos, C.M. Th17 Cells Are Refractory to Senescence and Retain Robust Antitumor Activity after Long-Term Ex Vivo Expansion. JCI Insight 2017, 2, e90772. [Google Scholar] [CrossRef]
- Majchrzak, K.; Nelson, M.H.; Bailey, S.R.; Bowers, J.S.; Yu, X.-Z.; Rubinstein, M.P.; Himes, R.A.; Paulos, C.M. Exploiting IL-17-Producing CD4+ and CD8+ T Cells to Improve Cancer Immunotherapy in the Clinic. Cancer Immunol. Immunother. 2016, 65, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Zambrano-Zaragoza, J.F.; Romo-Martínez, E.J.; Durán-Avelar, M.D.J.; García-Magallanes, N.; Vibanco-Pérez, N. Th17 Cells in Autoimmune and Infectious Diseases. Int. J. Inflamm. 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFbeta in the Context of an Inflammatory Cytokine Milieu Supports de Novo Differentiation of IL-17-Producing T Cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef]
- Alizadeh, D.; Katsanis, E.; Larmonier, N. The Multifaceted Role of Th17 Lymphocytes and Their Associated Cytokines in Cancer. Clin. Dev. Immunol. 2013, 2013, 957878. [Google Scholar] [CrossRef]
- Yang, L.; Anderson, D.E.; Baecher-Allan, C.; Hastings, W.D.; Bettelli, E.; Oukka, M.; Kuchroo, V.K.; Hafler, D.A. IL-21 and TGF-Beta Are Required for Differentiation of Human T(H)17 Cells. Nature 2008, 454, 350–352. [Google Scholar] [CrossRef]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Stritesky, G.L.; Yeh, N.; Kaplan, M.H. IL-23 Promotes Maintenance but Not Commitment to the Th17 Lineage. J. Immunol. 2008, 181, 5948–5955. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/β-Catenin Signaling in Development and Disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, R.; Fang, X.; Sun, Z. β-Catenin/TCF-1 Pathway in T Cell Development and Differentiation. J. Neuroimmune Pharmacol. 2012, 7, 750–762. [Google Scholar] [CrossRef]
- Muralidharan, S.; Hanley, P.J.; Liu, E.; Chakraborty, R.; Bollard, C.; Shpall, E.; Rooney, C.; Savoldo, B.; Rodgers, J.; Dotti, G. Activation of Wnt Signaling Arrests Effector Differentiation in Human Peripheral and Cord Blood-Derived T Lymphocytes. J. Immunol. 2011, 187, 5221–5232. [Google Scholar] [CrossRef]
- Reya, T.; Duncan, A.W.; Ailles, L.; Domen, J.; Scherer, D.C.; Willert, K.; Hintz, L.; Nusse, R.; Weissman, I.L. A Role for Wnt Signalling in Self-Renewal of Haematopoietic Stem Cells. Nature 2003, 423, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Gattinoni, L.; Ji, Y.; Restifo, N.P. Wnt/β-Catenin Signaling in T Cell Immunity and Cancer Immunotherapy. Clin. Cancer Res. 2010, 16, 4695–4701. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Lee, K.-A.; Yoon, H.-B.; Yoo, S.-A.; Park, Y.W.; Chung, Y.; Kim, W.-U.; Kang, C.-Y. The Wnt Inhibitor Secreted Frizzled-Related Protein 1 (sFRP1) Promotes Human Th17 Differentiation. Eur. J. Immunol. 2012, 42, 2564–2573. [Google Scholar] [CrossRef]
- Majchrzak, K.; Nelson, M.H.; Bowers, J.S.; Bailey, S.R.; Wyatt, M.M.; Wrangle, J.M.; Rubinstein, M.P.; Varela, J.C.; Li, Z.; Himes, R.A.; et al. β-Catenin and PI3Kδ Inhibition Expands Precursor Th17 Cells with Heightened Stemness and Antitumor Activity. JCI Insight 2017, 2, e90547. [Google Scholar] [CrossRef] [PubMed]
- Kol, A.; Walker, N.J.; Nordstrom, M.; Borjesson, D.L. Th17 Pathway As a Target for Multipotent Stromal Cell Therapy in Dogs: Implications for Translational Research. PLoS ONE 2016, 11, e0148568. [Google Scholar] [CrossRef]
- Ritt, M.; Lindborg, B.; O’Brien, T.; Bisignano, J.; Modiano, J. Stimulation with Concanavalin-A Induces IL-17 Production by Canine Peripheral T Cells. Vet. Sci. 2015, 2, 43–51. [Google Scholar] [CrossRef]
- Bujak, J.K.; Pingwara, R.; Nelson, M.H.; Majchrzak, K. Adoptive Cell Transfer: New Perspective Treatment in Veterinary Oncology. Acta Vet. Scand. 2018, 60, 60. [Google Scholar] [CrossRef]
- Park, J.S.; Withers, S.S.; Modiano, J.F.; Kent, M.S.; Chen, M.; Luna, J.I.; Culp, W.T.N.; Sparger, E.E.; Rebhun, R.B.; Monjazeb, A.M.; et al. Canine Cancer Immunotherapy Studies: Linking Mouse and Human. J. Immunother. Cancer 2016, 4, 97. [Google Scholar] [CrossRef] [PubMed]
- Khanna, C.; Lindblad-Toh, K.; Vail, D.; London, C.; Bergman, P.; Barber, L.; Breen, M.; Kitchell, B.; McNeil, E.; Modiano, J.F.; et al. The Dog as a Cancer Model. Nat. Biotechnol. 2006, 24, 1065–1066. [Google Scholar] [CrossRef] [PubMed]
- Paoloni, M.; Khanna, C. Translation of New Cancer Treatments from Pet Dogs to Humans. Nat. Rev. Cancer 2008, 8, 147–156. [Google Scholar] [CrossRef]
- Chow, L.; Wheat, W.; Ramirez, D.; Impastato, R.; Dow, S. Direct Comparison of Canine and Human Immune Responses Using Transcriptomic and Functional Analyses. Sci. Rep. 2024, 14, 2207. [Google Scholar] [CrossRef]
- Kulka, M.; Brennan, K.; Mc Gee, M. Investigation of Canine Extracellular Vesicles in Diffuse Large B-Cell Lymphomas. PLoS ONE 2022, 17, e0274261. [Google Scholar] [CrossRef]
- Rothe, K.; Bismarck, D.; Büttner, M.; Alber, G.; von Buttlar, H. Canine Peripheral Blood CD4+ CD8+ Double-Positive T Cell Subpopulations Exhibit Distinct T Cell Phenotypes and Effector Functions. Vet. Immunol. Immunopathol. 2017, 185, 48–56. [Google Scholar] [CrossRef]
- Szopa, I.M.; Granica, M.; Bujak, J.K.; Łabędź, A.; Błaszczyk, M.; Paulos, C.M.; Majchrzak-Kuligowska, K. Effective Activation and Expansion of Canine Lymphocytes Using a Novel Nano-Sized Magnetic Beads Approach. Front. Immunol. 2021, 12, 604066. [Google Scholar] [CrossRef] [PubMed]
- Panjwani, M.K.; Smith, J.B.; Schutsky, K.; Gnanandarajah, J.; O’Connor, C.M.; Powell, D.J.; Mason, N.J. Feasibility and Safety of RNA-Transfected CD20-Specific Chimeric Antigen Receptor T Cells in Dogs with Spontaneous B Cell Lymphoma. Mol. Ther. 2016, 24, 1602–1614. [Google Scholar] [CrossRef]
- Panjwani, M.K.; Atherton, M.J.; MaloneyHuss, M.A.; Haran, K.P.; Xiong, A.; Gupta, M.; Kulikovsaya, I.; Lacey, S.F.; Mason, N.J. Establishing a Model System for Evaluating CAR T Cell Therapy Using Dogs with Spontaneous Diffuse Large B Cell Lymphoma. Oncoimmunology 2020, 9, 1676615. [Google Scholar] [CrossRef]
- Rotolo, A.; Atherton, M.J.; Kasper, B.T.; Haran, K.P.; Mason, N.J. Genetic Re-Direction of Canine Primary T Cells for Clinical Trial Use in Pet Dogs with Spontaneous Cancer. STAR Protoc. 2021, 2, 100905. [Google Scholar] [CrossRef]
- Muranski, P.; Restifo, N.P. Essentials of Th17 Cell Commitment and Plasticity. Blood 2013, 121, 2402–2414. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.L.; Dorneles, E.M.S.; Soares, R.P.; Magalhães, C.P.; Costa-Pereira, C.; Lage, A.P.; Teixeira-Carvalho, A.; Martins-Filho, O.A.; Araújo, M.S.S. Cross-Reactivity of Commercially Available Anti-Human Monoclonal Antibodies with Canine Cytokines: Establishment of a Reliable Panel to Detect the Functional Profile of Peripheral Blood Lymphocytes by Intracytoplasmic Staining. Acta Vet. Scand. 2015, 57, 51. [Google Scholar] [CrossRef] [PubMed]
- Staal, F.J.T.; Luis, T.C.; Tiemessen, M.M. WNT Signalling in the Immune System: WNT Is Spreading Its Wings. Nat. Rev. Immunol. 2008, 8, 581–593. [Google Scholar] [CrossRef]
- Fleming, H.E.; Janzen, V.; Lo Celso, C.; Guo, J.; Leahy, K.M.; Kronenberg, H.M.; Scadden, D.T. Wnt Signaling in the Niche Enforces Hematopoietic Stem Cell Quiescence and Is Necessary to Preserve Self-Renewal in Vivo. Cell Stem Cell 2008, 2, 274–283. [Google Scholar] [CrossRef]
- Mulroy, T.; McMahon, J.A.; Burakoff, S.J.; McMahon, A.P.; Sen, J. Wnt-1 and Wnt-4 Regulate Thymic Cellularity. Eur. J. Immunol. 2002, 32, 967–971. [Google Scholar] [CrossRef]
- Okamura, R.M.; Sigvardsson, M.; Galceran, J.; Verbeek, S.; Clevers, H.; Grosschedl, R. Redundant Regulation of T Cell Differentiation and TCRalpha Gene Expression by the Transcription Factors LEF-1 and TCF-1. Immunity 1998, 8, 11–20. [Google Scholar] [CrossRef]
- Willinger, T.; Freeman, T.; Herbert, M.; Hasegawa, H.; McMichael, A.J.; Callan, M.F.C. Human Naive CD8 T Cells Down-Regulate Expression of the WNT Pathway Transcription Factors Lymphoid Enhancer Binding Factor 1 and Transcription Factor 7 (T Cell Factor-1) Following Antigen Encounter in Vitro and in Vivo. J. Immunol. 2006, 176, 1439–1446. [Google Scholar] [CrossRef]
- Dihlmann, S.; von Knebel Doeberitz, M. Wnt/Beta-Catenin-Pathway as a Molecular Target for Future Anti-Cancer Therapeutics. Int. J. Cancer 2005, 113, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-M.A.; Mishina, Y.M.; Liu, S.; Cheung, A.; Stegmeier, F.; Michaud, G.A.; Charlat, O.; Wiellette, E.; Zhang, Y.; Wiessner, S.; et al. Tankyrase Inhibition Stabilizes Axin and Antagonizes Wnt Signalling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef]
- Wu, X.; Luo, F.; Li, J.; Zhong, X.; Liu, K. Tankyrase 1 Inhibitior XAV939 Increases Chemosensitivity in Colon Cancer Cell Lines via Inhibition of the Wnt Signaling Pathway. Int. J. Oncol. 2016, 48, 1333–1340. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, L.; Aldarouish, M.; Zhou, Z.; Pan, N.; Liu, J.; Chen, F.; Wang, L. Wnt Pathway Activator TWS119 Enhances the Proliferation and Cytolytic Activity of Human γδT Cells against Colon Cancer. Exp. Cell Res. 2018, 362, 63–71. [Google Scholar] [CrossRef]
- Blobaum, A.L.; Uddin, J.; Felts, A.S.; Crews, B.C.; Rouzer, C.A.; Marnett, L.J. The 2′-Trifluoromethyl Analogue of Indomethacin Is a Potent and Selective COX-2 Inhibitor. ACS Med. Chem. Lett. 2013, 4, 486–490. [Google Scholar] [CrossRef]
- Luz-Crawford, P.; Kurte, M.; Bravo-Alegría, J.; Contreras, R.; Nova-Lamperti, E.; Tejedor, G.; Noël, D.; Jorgensen, C.; Figueroa, F.; Djouad, F.; et al. Mesenchymal Stem Cells Generate a CD4+ CD25+ Foxp3+ Regulatory T Cell Population during the Differentiation Process of Th1 and Th17 Cells. Stem Cell Res. Ther. 2013, 4, 65. [Google Scholar] [CrossRef]
- Shen, Y.; Yang, T.; Wang, J.; Xu, Q.; Li, R.; Pan, W.; Li, G.; Wang, Z.; Tan, J.; Wu, J.; et al. Indomethacin Enhances the Cytotoxicity of Recombinant Human Lymphotoxin α on Tumor Cells by Suppressing NF-κB Signaling. Cancer Biol. Ther. 2007, 6, 1424–1429. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Adhikary, B.; Chand, S.; Maity, B.; Bandyopadhyay, S.K.; Chattopadhyay, S. Molecular Mechanism of Indomethacin-Induced Gastropathy. Free Radic. Biol. Med. 2012, 52, 1175–1187. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, T.; Li, N.; Meng, Z.; Wang, W.; Wang, B.; Song, D. Potential Effects of Indomethacin on Alleviating Osteoarthritis Progression in Vitro. J. Musculoskelet. Neuronal Interact. 2024, 24, 409–419. [Google Scholar]
- Hawcroft, G.; D’Amico, M.; Albanese, C.; Markham, A.F.; Pestell, R.G.; Hull, M.A. Indomethacin Induces Differential Expression of β-Catenin, γ-Catenin and T-Cell Factor Target Genes in Human Colorectal Cancer Cells. Carcinogenesis 2002, 23, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Dihlmann, S.; Siermann, A.; von Knebel Doeberitz, M. The Nonsteroidal Anti-Inflammatory Drugs Aspirin and Indomethacin Attenuate Beta-Catenin/TCF-4 Signaling. Oncogene 2001, 20, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Dihlmann, S.; Klein, S.; Doeberitz, M.V.K. Reduction of Beta-Catenin/T-Cell Transcription Factor Signaling by Aspirin and Indomethacin Is Caused by an Increased Stabilization of Phosphorylated Beta-Catenin. Mol. Cancer Ther. 2003, 2, 509–516. [Google Scholar]
- Gattinoni, L.; Zhong, X.-S.; Palmer, D.C.; Ji, Y.; Hinrichs, C.S.; Yu, Z.; Wrzesinski, C.; Boni, A.; Cassard, L.; Garvin, L.M.; et al. Wnt Signaling Arrests Effector T Cell Differentiation and Generates CD8+ Memory Stem Cells. Nat. Med. 2009, 15, 808–813. [Google Scholar] [CrossRef]
- Yang, S.; Liu, F.; Wang, Q.J.; Rosenberg, S.A.; Morgan, R.A. The Shedding of CD62L (L-Selectin) Regulates the Acquisition of Lytic Activity in Human Tumor Reactive T Lymphocytes. PLoS ONE 2011, 6, e22560. [Google Scholar] [CrossRef] [PubMed]
- Baaten, B.J.; Li, C.-R.; Bradley, L.M. Multifaceted Regulation of T Cells by CD44. Commun. Integr. Biol. 2010, 3, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Gattinoni, L.; Speiser, D.E.; Lichterfeld, M.; Bonini, C. T Memory Stem Cells in Health and Disease. Nat. Med. 2017, 23, 18–27. [Google Scholar] [CrossRef]
- Gattinoni, L.; Klebanoff, C.A.; Palmer, D.C.; Wrzesinski, C.; Kerstann, K.; Yu, Z.; Finkelstein, S.E.; Theoret, M.R.; Rosenberg, S.A.; Restifo, N.P. Acquisition of Full Effector Function in Vitro Paradoxically Impairs the in Vivo Antitumor Efficacy of Adoptively Transferred CD8+ T Cells. J. Clin. Investig. 2005, 115, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Rangel Rivera, G.O.; Knochelmann, H.M.; Dwyer, C.J.; Smith, A.S.; Wyatt, M.M.; Rivera-Reyes, A.M.; Thaxton, J.E.; Paulos, C.M. Fundamentals of T Cell Metabolism and Strategies to Enhance Cancer Immunotherapy. Front. Immunol. 2021, 12, 645242. [Google Scholar] [CrossRef]
- Chapman, N.M.; Boothby, M.R.; Chi, H. Metabolic Coordination of T Cell Quiescence and Activation. Nat. Rev. Immunol. 2020, 20, 55–70. [Google Scholar] [CrossRef]
- Oberholtzer, N.; Quinn, K.M.; Chakraborty, P.; Mehrotra, S. New Developments in T Cell Immunometabolism and Implications for Cancer Immunotherapy. Cells 2022, 11, 708. [Google Scholar] [CrossRef]
- Ma, S.; Ming, Y.; Wu, J.; Cui, G. Cellular Metabolism Regulates the Differentiation and Function of T-Cell Subsets. Cell. Mol. Immunol. 2024, 21, 419–435. [Google Scholar] [CrossRef]
- Shin, B.; Benavides, G.A.; Geng, J.; Koralov, S.B.; Hu, H.; Darley-Usmar, V.M.; Harrington, L.E. Mitochondrial Oxidative Phosphorylation Regulates the Fate Decision between Pathogenic Th17 and Regulatory T Cells. Cell Rep. 2020, 30, 1898–1909.e4. [Google Scholar] [CrossRef]
- Chatterjee, S.; Daenthanasanmak, A.; Chakraborty, P.; Wyatt, M.W.; Dhar, P.; Selvam, S.P.; Fu, J.; Zhang, J.; Nguyen, H.; Kang, I.; et al. CD38-NAD + Axis Regulates Immunotherapeutic Anti-Tumor T Cell Response. Cell Metab. 2018, 27, 85–100.e8. [Google Scholar] [CrossRef]
- Dwyer, C.J.; Arhontoulis, D.C.; Rangel Rivera, G.O.; Knochelmann, H.M.; Smith, A.S.; Wyatt, M.M.; Rubinstein, M.P.; Atkinson, C.; Thaxton, J.E.; Neskey, D.M.; et al. Ex Vivo Blockade of PI3K Gamma or Delta Signaling Enhances the Antitumor Potency of Adoptively Transferred CD8+ T Cells. Eur. J. Immunol. 2020, 50, 1386–1399. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Guillevin, R.; Vallée, J.-N. Vasculogenesis and Angiogenesis Initiation under Normoxic Conditions through Wnt/β-Catenin Pathway in Gliomas. Rev. Neurosci. 2018, 29, 71–91. [Google Scholar] [CrossRef] [PubMed]
- Karmaus, P.W.F.; Chen, X.; Lim, S.A.; Herrada, A.A.; Nguyen, T.-L.M.; Xu, B.; Dhungana, Y.; Rankin, S.; Chen, W.; Rosencrance, C.; et al. Metabolic Heterogeneity Underlies Reciprocal Fates of TH17 Cell Stemness and Plasticity. Nature 2019, 565, 101–105. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szopa, I.M.; Majchrzak-Kuligowska, K.; Pingwara, R.; Kulka, M.; Taşdemir, M.; Gajewska, M. A New Method of Canine CD4+ T Lymphocyte Differentiation Towards the Th17 Phenotype with Analysis of Properties and Mitochondrial Activity. Int. J. Mol. Sci. 2025, 26, 4946. https://doi.org/10.3390/ijms26104946
Szopa IM, Majchrzak-Kuligowska K, Pingwara R, Kulka M, Taşdemir M, Gajewska M. A New Method of Canine CD4+ T Lymphocyte Differentiation Towards the Th17 Phenotype with Analysis of Properties and Mitochondrial Activity. International Journal of Molecular Sciences. 2025; 26(10):4946. https://doi.org/10.3390/ijms26104946
Chicago/Turabian StyleSzopa, Iwona Monika, Kinga Majchrzak-Kuligowska, Rafał Pingwara, Marek Kulka, Monika Taşdemir, and Małgorzata Gajewska. 2025. "A New Method of Canine CD4+ T Lymphocyte Differentiation Towards the Th17 Phenotype with Analysis of Properties and Mitochondrial Activity" International Journal of Molecular Sciences 26, no. 10: 4946. https://doi.org/10.3390/ijms26104946
APA StyleSzopa, I. M., Majchrzak-Kuligowska, K., Pingwara, R., Kulka, M., Taşdemir, M., & Gajewska, M. (2025). A New Method of Canine CD4+ T Lymphocyte Differentiation Towards the Th17 Phenotype with Analysis of Properties and Mitochondrial Activity. International Journal of Molecular Sciences, 26(10), 4946. https://doi.org/10.3390/ijms26104946