
Angolensin Isolated from Pterocarpus indicus Willd. Attenuates LPS-Induced Sickness Behaviors in Mice and Exhibits CNS Safety

 , ,
, ,  , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Cytotoxicity Profiles of Angolensin in BV-2 Microglial Cells
2.2. In Vitro Anti-Inflammatory Effects of Angolensin in LPS-Stimulated BV-2 Cells
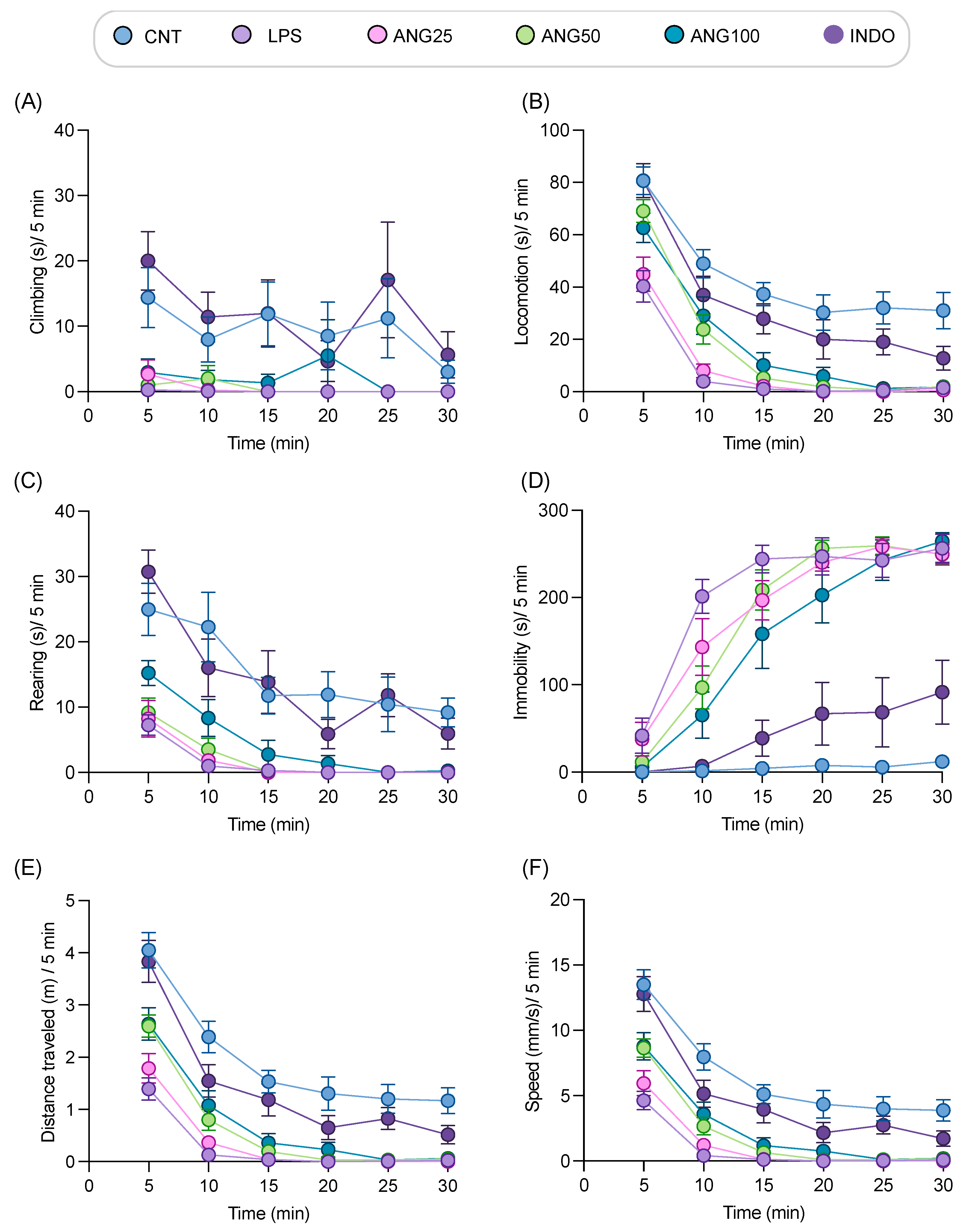
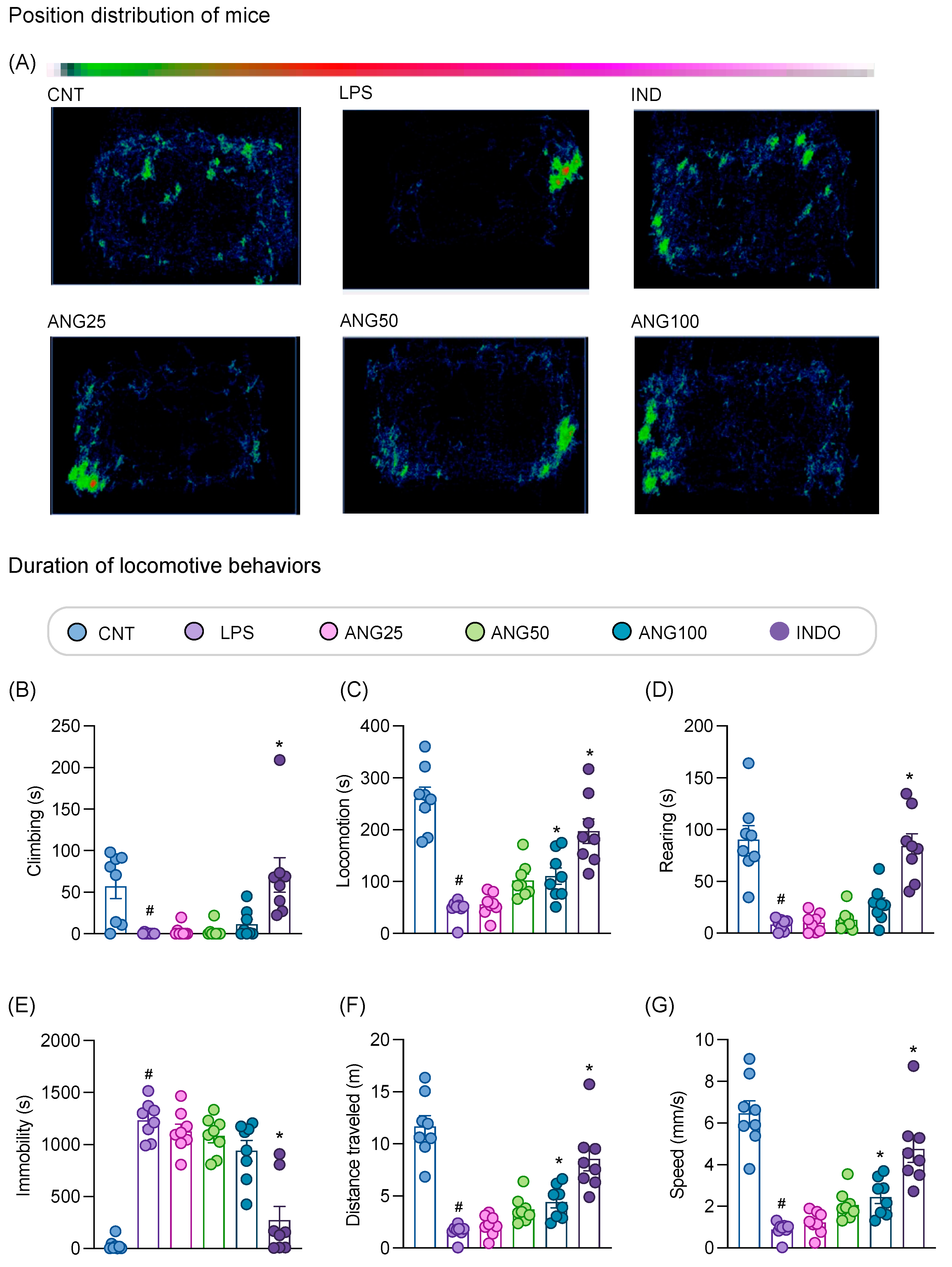
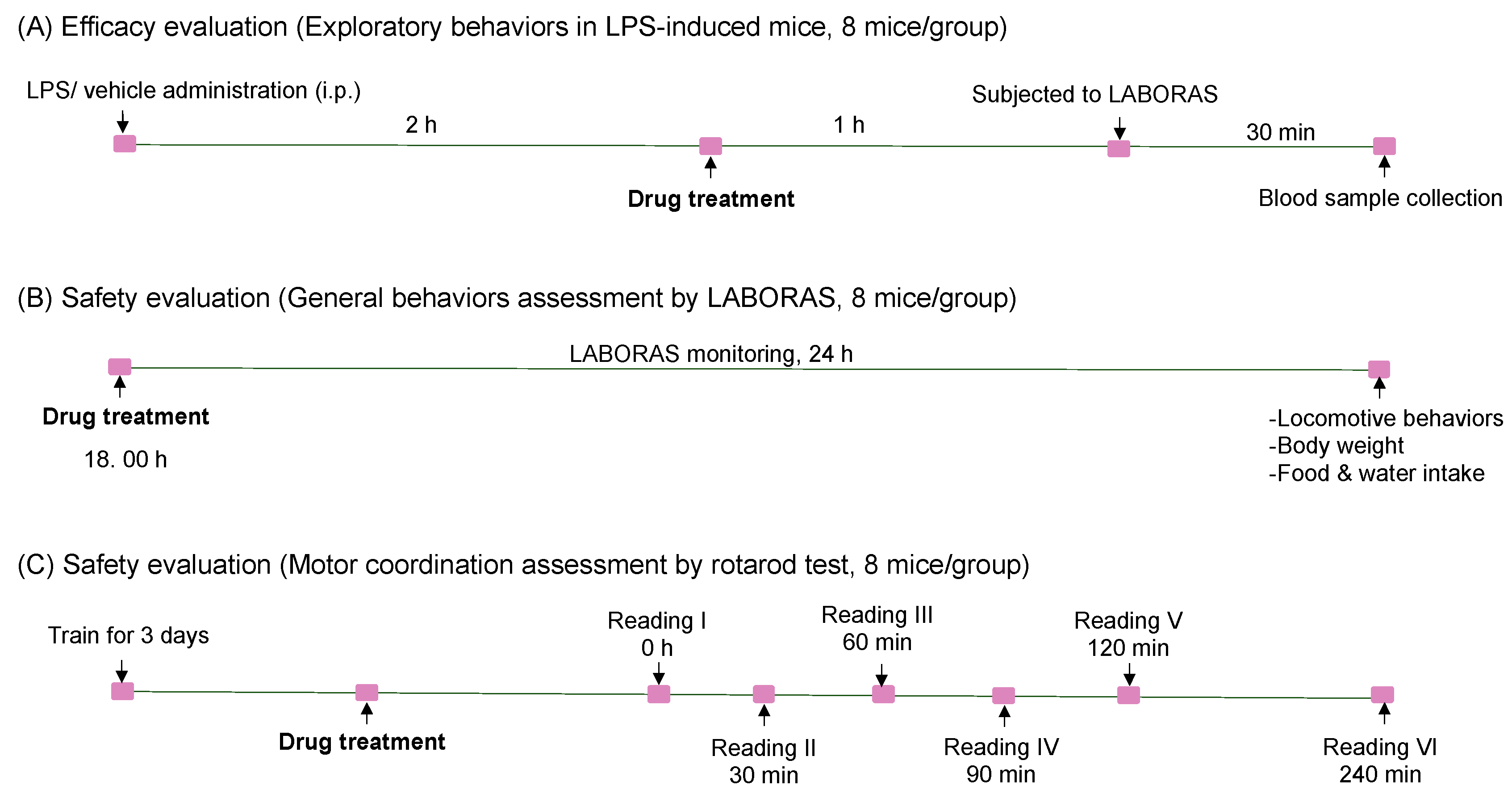
2.3. The Effect of Angolensin on LPS-Induced Sickness Behaviors
2.4. The Effects of Angolensin on the Expression of Proinflammatory Cytokines in LPS-Induced Mice
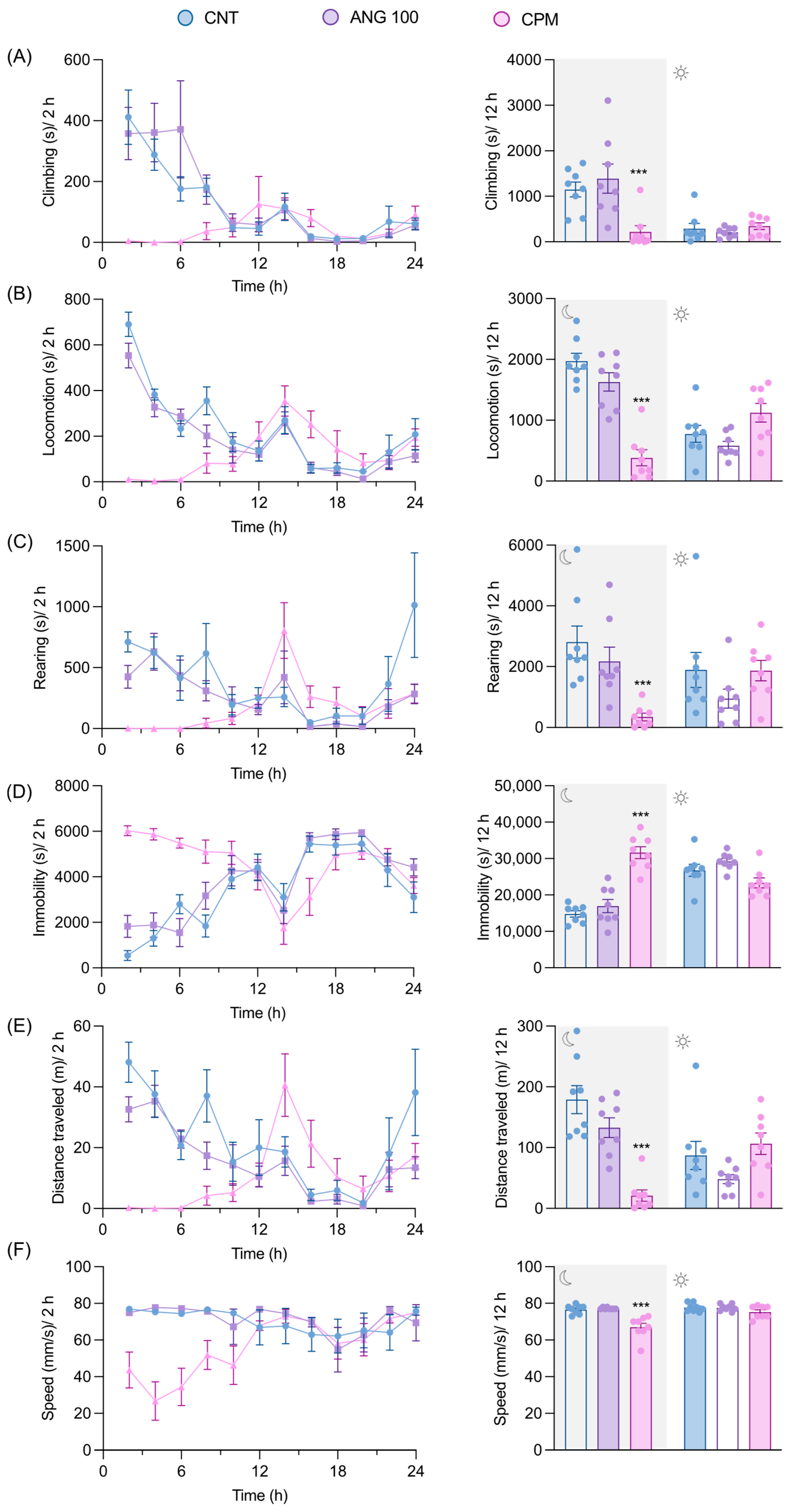
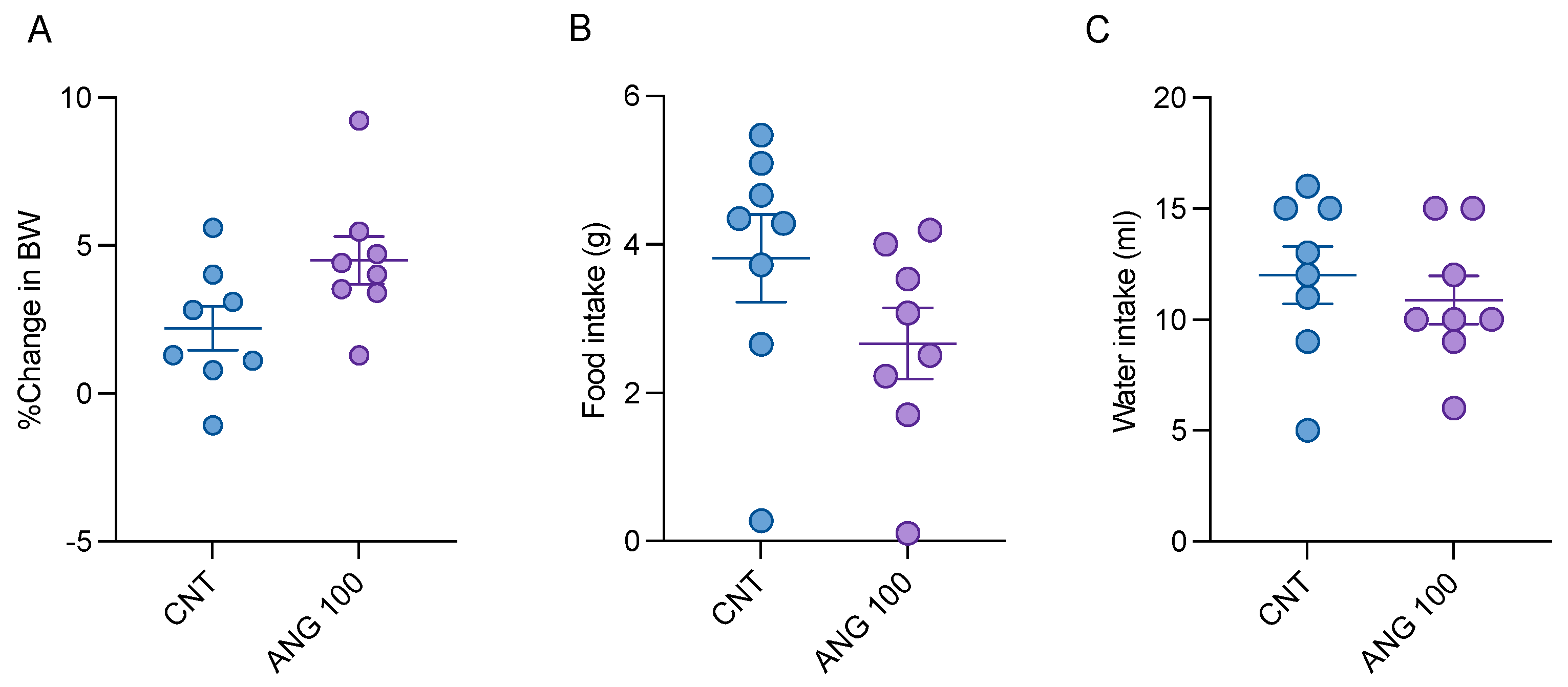
2.5. The Effects of Angolensin on General Behaviors and Well-Being of Mice
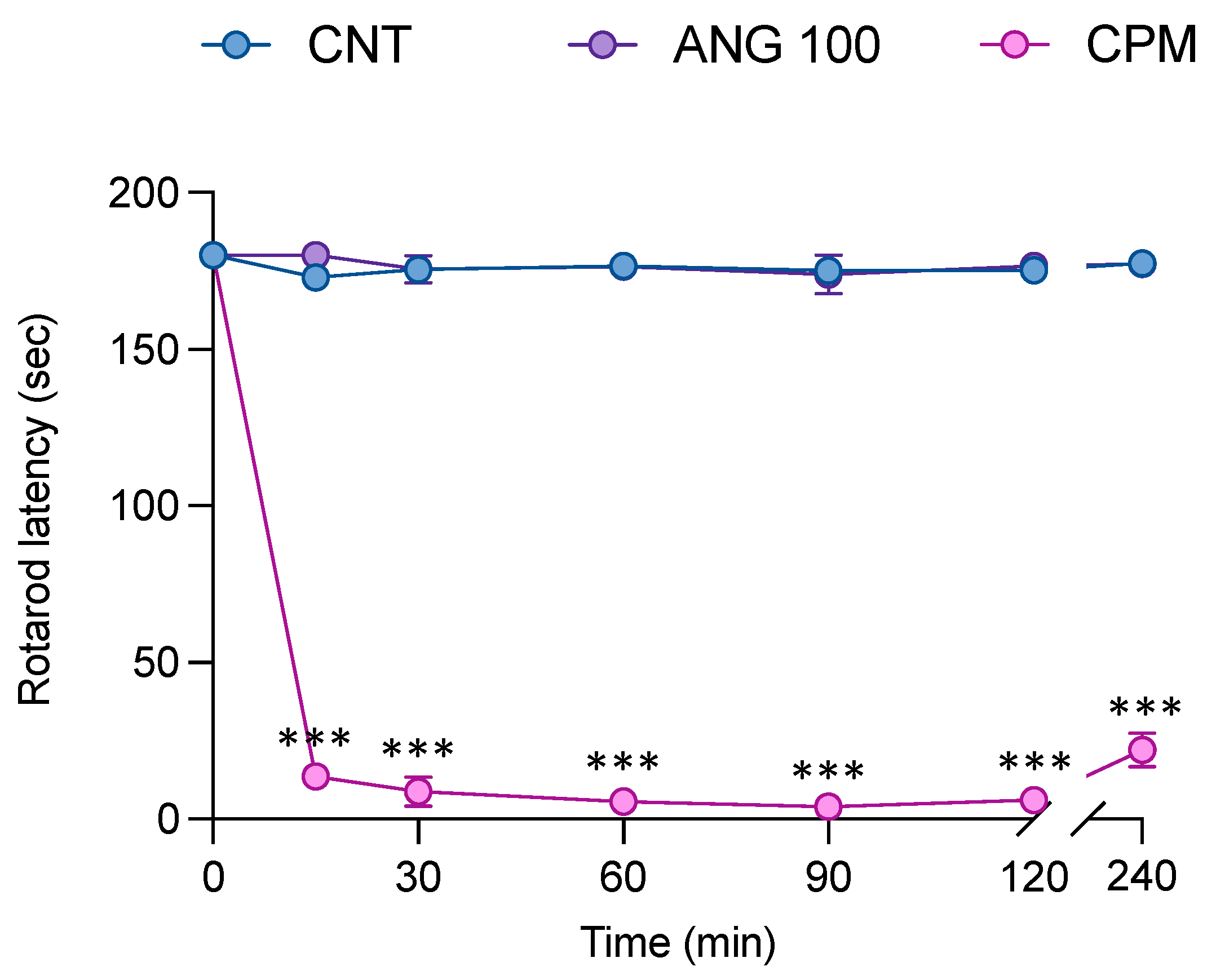
2.6. The Effect of Angolensin on Motor Coordination
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Authentication
4.2. Isolation of Angolensin
4.3. Cell Culture
4.4. MTT Assay
4.5. Double Staining Assay
4.6. Cell Treatment and LPS Stimulation
4.7. Nitrite Assay
4.8. Cytokine Assay
4.9. Animals
4.10. Drugs and Treatment Administration
4.11. Assessment of Exploratory Behavior by LABORAS Automated Home-Cage Behavioral Analysis
4.12. Blood Sample Collection
4.13. Assessment of General Well-Being of Mice by LABORAS Automated Home-Cage Monitoring
4.14. Rotarod Test
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LPS | Lipopolysaccharide |
| LABORAS | Laboratory Animal Behavior Observation, Registration and Analysis System |
| CNS | Central nervous system |
| TNF-α | Tumor necrosis factor-alpha |
| IL-6 | Interleukin-6 |
| NO | Nitric oxide |
References
- Harden, L.M.; Kent, S.; Pittman, Q.J.; Roth, J. Fever and sickness behavior: Friend or foe? Brain Behav. Immun. 2015, 50, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Konsman, J.P.; Parnet, P.; Dantzer, R. Cytokine-induced sickness behaviour: Mechanisms and implications. Trends Neurosci. 2002, 25, 154–159. [Google Scholar] [CrossRef]
- Nandeesh, R.; Vijayakumar, S.; Munnolli, A.; Alreddy, A.; Veerapur, V.P.; Chandramohan, V.; Manjunatha, E. Bioactive phenolic fraction of Citrus maxima abate lipopolysaccharide-induced sickness behaviour and anorexia in mice: In-silico molecular docking and dynamic studies of biomarkers against NF-κB. Biomed. Pharmacother. 2018, 108, 1535–1545. [Google Scholar] [CrossRef]
- Khodr, B.; Khalil, Z. Modulation of inflammation by reactive oxygen species: Implications for aging and tissue repair. Free Radic. Biol. Med. 2001, 30, 1–8. [Google Scholar] [CrossRef]
- Lasselin, J.; Schedlowski, M.; Karshikoff, B.; Engler, H.; Lekander, M.; Konsman, J.P. Comparison of bacterial lipopolysaccharide-induced sickness behavior in rodents and humans: Relevance for symptoms of anxiety and depression. Neurosci. Biobehav. Rev. 2020, 115, 15–24. [Google Scholar] [CrossRef]
- Anderson, S.T.; Commins, S.; Moynagh, P.N.; Coogan, A.N. Lipopolysaccharide-induced sepsis induces long-lasting affective changes in the mouse. Brain Behav. Immun. 2015, 43, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Hijma, H.J.; Moss, L.M.; Gal, P.; Ziagkos, D.; de Kam, M.L.; Moerland, M.; Groeneveld, G.J. Challenging the challenge: A randomized controlled trial evaluating the inflammatory response and pain perception of healthy volunteers after single-dose LPS administration, as a potential model for inflammatory pain in early-phase drug development. Brain Behav. Immun. 2020, 88, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Tarselli, M.A.; Raehal, K.M.; Brasher, A.K.; Streicher, J.M.; Groer, C.E.; Cameron, M.D.; Bohn, L.M.; Micalizio, G.C. Synthesis of conolidine, a potent non-opioid analgesic for tonic and persistent pain. Nat. Chem. 2011, 3, 449–453. [Google Scholar] [CrossRef]
- Tapsoba, H.; Deschamps, J.-P. Use of medicinal plants for the treatment of oral diseases in Burkina Faso. J. Ethnopharmacol. 2006, 104, 68–78. [Google Scholar] [CrossRef]
- Nadembega, P.; Boussim, J.I.; Nikiema, J.B.; Poli, F.; Antognoni, F. Medicinal plants in Baskoure, Kourittenga Province, Burkina Faso: An ethnobotanical study. J. Ethnopharmacol. 2011, 133, 378–395. [Google Scholar] [CrossRef]
- Cha, H.-S.; Kim, W.-J.; Lee, M.-H.; Kim, S.-Y.; Kim, S.H.; Lee, K.-H.; Kim, T.-J. Inhibitory effect of Pterocarpus indicus Willd water extract on IgE/Ag-induced mast cell and atopic dermatitis-like mouse models. Biosci. Biotechnol. Biochem. 2016, 80, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Majumder, S.; Akter, F.; Islam, F.; Shahriar, M.; Alam, J. Pre-clinical investigation of analgesic, anti-diarrheal and CNS depressant effect of Pterocarpus indicus in Swiss albino mice. Jordan J. Pharm. Sci. 2021, 14, 85–94. [Google Scholar]
- Jagtap, A.G.; Fernandes, B.B. Analgesic potential of Pterocarpus indicus. Indian Drugs 2008, 45, 649–654. [Google Scholar]
- King, F.E.; King, T.J.; Warwick, A.J. 20. The chemistry of extractives from hardwoods. Part VI. Constituents of muninga, the heartwood of Pterocarpus Angolensis, A.: 6:4′-dihydroxy-5:7-dimethoxyisoflavone (muningin). J. Chem. Soc. (Resumed) 1952, 96–100. [Google Scholar] [CrossRef]
- Gupta, V.N.; Seshadri, T.R. Synthesis of angolensin. Proc. Indian Acad. Sci.-Sect. A 1956, 44, 223–227. [Google Scholar] [CrossRef]
- Seshadri, T.R. Polyphenols of Pterocarpus and Dalbergia woods. Phytochemistry 1972, 11, 881–898. [Google Scholar] [CrossRef]
- Pfitscher, A.; Reiter, E.; Jungbauer, A. Receptor binding and transactivation activities of red clover isoflavones and their metabolites. J. Steroid Biochem. Mol. Biol. 2008, 112, 87–94. [Google Scholar] [CrossRef]
- Pilotti, C.A.; Kondo, R.; Shimizu, K.; Sakai, K. An examination of the anti-fungal components in the heartwood extracts of Pterocarpus indicus. Mokuzai Gakkaishi = J. Jpn. Wood Res. Soc. 1995, 41, 593–597. [Google Scholar]
- Sato, M.; Tanaka, H.; Yamaguchi, R.; Oh-Uchi, T.; Etoh, H. Erythrina poeppigiana-derived phytochemical exhibiting antimicrobial activity against Candida albicans and methicillin-resistant Staphylococcus aureus. Lett. Appl. Microbiol. 2003, 37, 81–85. [Google Scholar] [CrossRef]
- Feunaing, R.T.; Tamfu, A.N.; Gbaweng, A.J.Y.; Mekontso Magnibou, L.; Ntchapda, F.; Henoumont, C.; Laurent, S.; Talla, E.; Dinica, R.M. In Vitro Evaluation of α-amylase and α-glucosidase Inhibition of 2,3-Epoxyprocyanidin C1 and Other Constituents from Pterocarpus erinaceus Poir. Molecules 2023, 28, 126. [Google Scholar] [CrossRef]
- Pereira de Souza Goldim, M.; Della Giustina, A.; Mathias, K.; de Oliveira Junior, A.; Fileti, M.E.; De Carli, R.; Zarbato, G.; Garbossa, L.; da Rosa, N.; Oliveira, J.; et al. Sickness Behavior Score Is Associated with Neuroinflammation and Late Behavioral Changes in Polymicrobial Sepsis Animal Model. Inflammation 2020, 43, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Biesmans, S.; Meert, T.F.; Bouwknecht, J.A.; Acton, P.D.; Davoodi, N.; De Haes, P.; Kuijlaars, J.; Langlois, X.; Matthews, L.J.R.; Ver Donck, L.; et al. Systemic Immune Activation Leads to Neuroinflammation and Sickness Behavior in Mice. Mediat. Inflamm. 2013, 2013, 271359. [Google Scholar] [CrossRef] [PubMed]
- Ragasa, C.Y.; De Luna, R.D.; Hofilena, J.G. Antimicrobial terpenoids from Pterocarpus indicus. Nat. Prod. Res. 2005, 19, 305–309. [Google Scholar] [CrossRef]
- Powell, E.; Shanle, E.; Brinkman, A.; Li, J.; Keles, S.; Wisinski, K.B.; Huang, W.; Xu, W. Identification of estrogen receptor dimer selective ligands reveals growth-inhibitory effects on cells that co-express ERα and ERβ. PLoS ONE 2012, 7, e30993. [Google Scholar] [CrossRef]
- Morgan, S.C.; Taylor, D.L.; Pocock, J.M. Microglia release activators of neuronal proliferation mediated by activation of mitogen-activated protein kinase, phosphatidylinositol-3-kinase/Akt and delta-Notch signalling cascades. J. Neurochem. 2004, 90, 89–101. [Google Scholar] [CrossRef]
- Ji, R.R.; Chamessian, A.; Zhang, Y.Q. Pain regulation by non-neuronal cells and inflammation. Science 2016, 354, 572–577. [Google Scholar] [CrossRef]
- Hart, B.L. Beyond Fever: Comparative Perspectives on Sickness Behavior. In Encyclopedia of Animal Behavior; Breed, M.D., Moore, J., Eds.; Academic Press: Oxford, UK, 2010; pp. 205–210. [Google Scholar] [CrossRef]
- Chang, Y.C.; Tsai, M.H.; Sheu, W.H.H.; Hsieh, S.C.; Chiang, A.N. The therapeutic potential and mechanisms of action of quercetin in relation to lipopolysaccharide-induced sepsis in vitro and in vivo. PLoS ONE 2013, 8, e80744. [Google Scholar] [CrossRef]
- Klein, S.L.; Nelson, R.J. Social Behavior and Parasites☆. In Encyclopedia of Animal Behavior, 2nd ed.; Choe, J.C., Ed.; Academic Press: Oxford, UK, 2019; pp. 739–746. [Google Scholar] [CrossRef]
- Hasriadi; Dasuni Wasana, P.W.; Vajragupta, O.; Rojsitthisak, P.; Towiwat, P. Automated home-cage monitoring as a potential measure of sickness behaviors and pain-like behaviors in LPS-treated mice. PLoS ONE 2021, 16, e0256706. [Google Scholar] [CrossRef] [PubMed]
- Teeling, J.L.; Cunningham, C.; Newman, T.A.; Perry, V.H. The effect of non-steroidal anti-inflammatory agents on behavioural changes and cytokine production following systemic inflammation: Implications for a role of COX-1. Brain Behav. Immun. 2010, 24, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Bangasser, D.A.; Valentino, R.J. Sex differences in molecular and cellular substrates of stress. Cell Mol. Neurobiol. 2012, 32, 709–723. [Google Scholar] [CrossRef]
- Senst, L.; Baimoukhametova, D.; Sterley, T.-L.; Bains, J.S. Sexually dimorphic neuronal responses to social isolation. eLife 2016, 5, e18726. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Akad, A.; Aderogba, R.; Chowdhury, T.G.; Aoki, C. Dendrites of the dorsal and ventral hippocampal CA1 pyramidal neurons of singly housed female rats exhibit lamina-specific growths and retractions during adolescence that are responsive to pair housing. Synapse 2018, 72, e22034. [Google Scholar] [CrossRef] [PubMed]
- Kercmar, J.; Tobet, S.A.; Majdic, G. Social isolation during puberty affects female sexual behavior in mice. Front. Behav. Neurosci. 2014, 8, 337. [Google Scholar] [CrossRef] [PubMed]
- Palanza, P.; Gioiosa, L.; Parmigiani, S. Social stress in mice: Gender differences and effects of estrous cycle and social dominance. Physiol. Behav. 2001, 73, 411–420. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef]
- Archie, S.R.; Al Shoyaib, A.; Cucullo, L. Blood-Brain Barrier Dysfunction in CNS Disorders and Putative Therapeutic Targets: An Overview. Pharmaceutics 2021, 13, 1779. [Google Scholar] [CrossRef]
- Banks, W.A.; Gray, A.M.; Erickson, M.A.; Salameh, T.S.; Damodarasamy, M.; Sheibani, N.; Meabon, J.S.; Wing, E.E.; Morofuji, Y.; Cook, D.G.; et al. Lipopolysaccharide-induced blood-brain barrier disruption: Roles of cyclooxygenase, oxidative stress, neuroinflammation, and elements of the neurovascular unit. J. Neuroinflammation 2015, 12, 223. [Google Scholar] [CrossRef]
- Hasriadi; Wasana, P.W.D.; Sritularak, B.; Vajragupta, O.; Rojsitthisak, P.; Towiwat, P. Batatasin III, a Constituent of Dendrobium scabrilingue, Improves Murine Pain-like Behaviors with a Favorable CNS Safety Profile. J. Nat. Prod. 2022, 85, 1816–1825. [Google Scholar] [CrossRef]
- Poon, D.C.-H.; Ho, Y.-S.; Chiu, K.; Chang, R.C.-C. Cytokines: How important are they in mediating sickness? Neurosci. Biobehav. Rev. 2013, 37, 1–10. [Google Scholar] [CrossRef]
- Castagne, V.; Froger-Colleaux, C.; Esneault, E.; Anne Marie, H.; Lemaire, M.; Porsolt, R.D. Central nervous system (CNS) safety pharmacology studies. In Drug Discovery and Evaluation: Safety and Pharmacokinetic Assays; Springer: Berlin/Heidelberg, Germany, 2013; pp. 17–72. [Google Scholar]
- Auriel, E.; Regev, K.; Korczyn, A.D. Chapter 38—Nonsteroidal anti-inflammatory drugs exposure and the central nervous system. In Handbook of Clinical Neurology; Biller, J., Ferro, J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 119, pp. 577–584. [Google Scholar]
- Ripamonti, C.; Bruera, E. CNS Adverse Effects of Opioids in Cancer Patients. CNS Drugs 1997, 8, 21–37. [Google Scholar] [CrossRef]
- Mead, A.N.; Amouzadeh, H.R.; Chapman, K.; Ewart, L.; Giarola, A.; Jackson, S.J.; Jarvis, P.; Jordaan, P.; Redfern, W.; Traebert, M. Assessing the predictive value of the rodent neurofunctional assessment for commonly reported adverse events in phase I clinical trials. Regul. Toxicol. Pharmacol. 2016, 80, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.J.; Castagné, V.; Moser, P.C.; Mittelstadt, S.W. Comparison of methods for the assessment of locomotor activity in rodent safety pharmacology studies. J. Pharmacol. Toxicol. Methods 2011, 64, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Lajoie, Y.; Gallagher, S. Predicting falls within the elderly community: Comparison of postural sway, reaction time, the Berg balance scale and the Activities-specific Balance Confidence (ABC) scale for comparing fallers and non-fallers. Arch. Gerontol. Geriatr. 2004, 38, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Hegeman, J.; van den Bemt, B.J.; Duysens, J.; van Limbeek, J. NSAIDs and the risk of accidental falls in the elderly: A systematic review. Drug Saf. 2009, 32, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.G. Drug Discovery and Evaluation: Safety and Pharmacokinetic Assays; with 125 Tables; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Gauvin, D.V.; Zimmermann, Z.J.; Dalton, J.A.; Baird, T.J.; Kallman, M.J. CNS Safety Screening Under ICH S7A Guidelines Requires Observations of Multiple Behavioral Units to Assess Motor Function. Int. J. Toxicol. 2019, 38, 339–356. [Google Scholar] [CrossRef]
- Salakka, A.; Wähälä, K. Synthesis of α-methyldeoxybenzoins. J. Chem. Soc. Perkin Trans. 1999, 1, 2601–2604. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinalchemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nwe, S.Y.; Dasuni Wasana, P.W.; Hasriadi; Towiwat, P.; Thongphichai, W.; Sritularak, B.; Sukrong, S. Angolensin Isolated from Pterocarpus indicus Willd. Attenuates LPS-Induced Sickness Behaviors in Mice and Exhibits CNS Safety. Int. J. Mol. Sci. 2025, 26, 4887. https://doi.org/10.3390/ijms26104887
Nwe SY, Dasuni Wasana PW, Hasriadi, Towiwat P, Thongphichai W, Sritularak B, Sukrong S. Angolensin Isolated from Pterocarpus indicus Willd. Attenuates LPS-Induced Sickness Behaviors in Mice and Exhibits CNS Safety. International Journal of Molecular Sciences. 2025; 26(10):4887. https://doi.org/10.3390/ijms26104887
Chicago/Turabian StyleNwe, San Yoon, Peththa Wadu Dasuni Wasana, Hasriadi, Pasarapa Towiwat, Wisuwat Thongphichai, Boonchoo Sritularak, and Suchada Sukrong. 2025. "Angolensin Isolated from Pterocarpus indicus Willd. Attenuates LPS-Induced Sickness Behaviors in Mice and Exhibits CNS Safety" International Journal of Molecular Sciences 26, no. 10: 4887. https://doi.org/10.3390/ijms26104887
APA StyleNwe, S. Y., Dasuni Wasana, P. W., Hasriadi, Towiwat, P., Thongphichai, W., Sritularak, B., & Sukrong, S. (2025). Angolensin Isolated from Pterocarpus indicus Willd. Attenuates LPS-Induced Sickness Behaviors in Mice and Exhibits CNS Safety. International Journal of Molecular Sciences, 26(10), 4887. https://doi.org/10.3390/ijms26104887