
A Structural In Silico Analysis of Novel Epitopes from Toxoplasma gondii Proteins for the Serodiagnosis of Toxoplasmosis

 , , and
, , and
Abstract
1. Introduction
2. Results
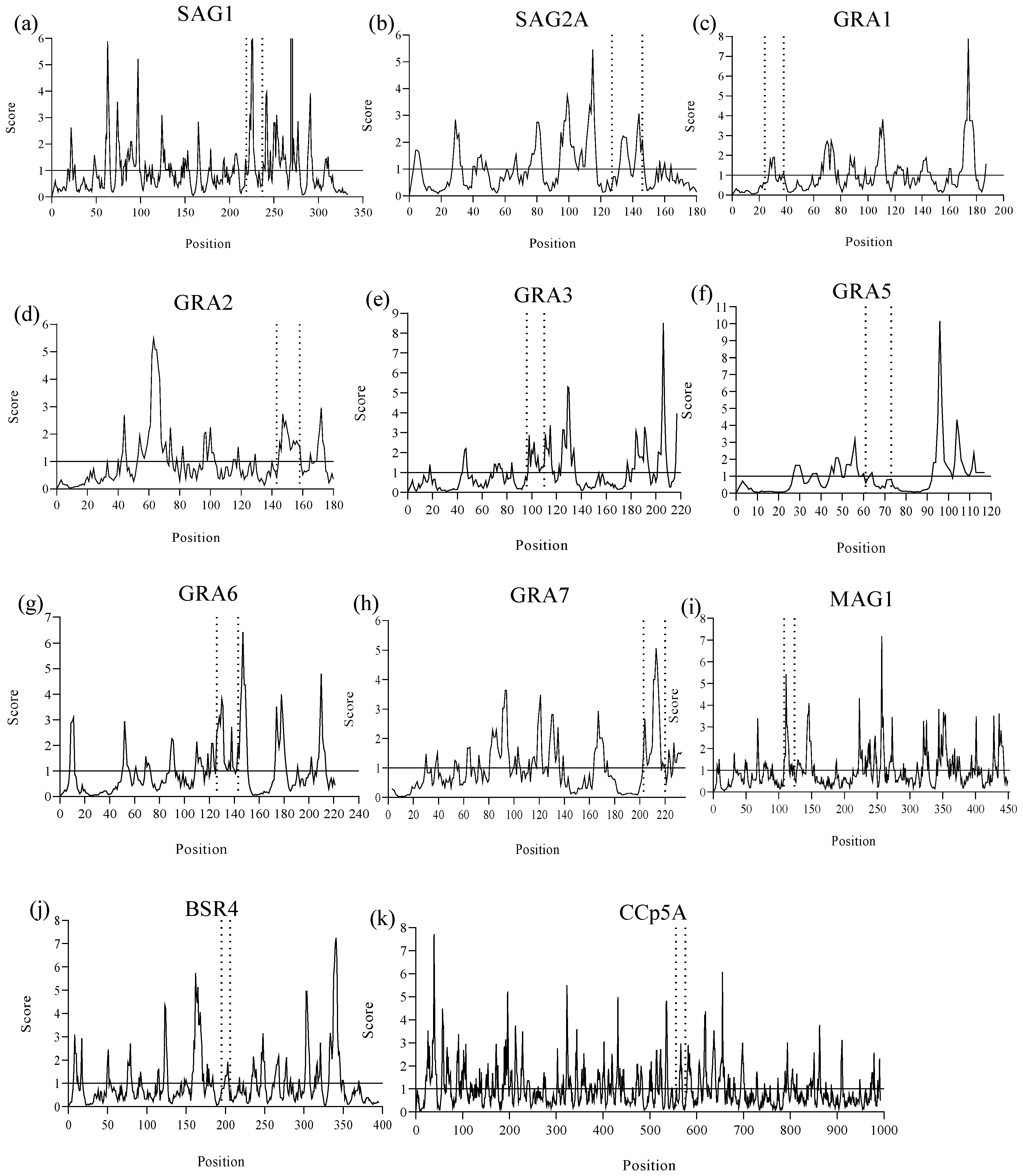
2.1. B-Cell Epitope Prediction Design
2.2. Homology and Phylogenetic Relationship
2.3. General Characteristics of Peptides: Physicochemical Properties, Accessibility, and Antigenicity
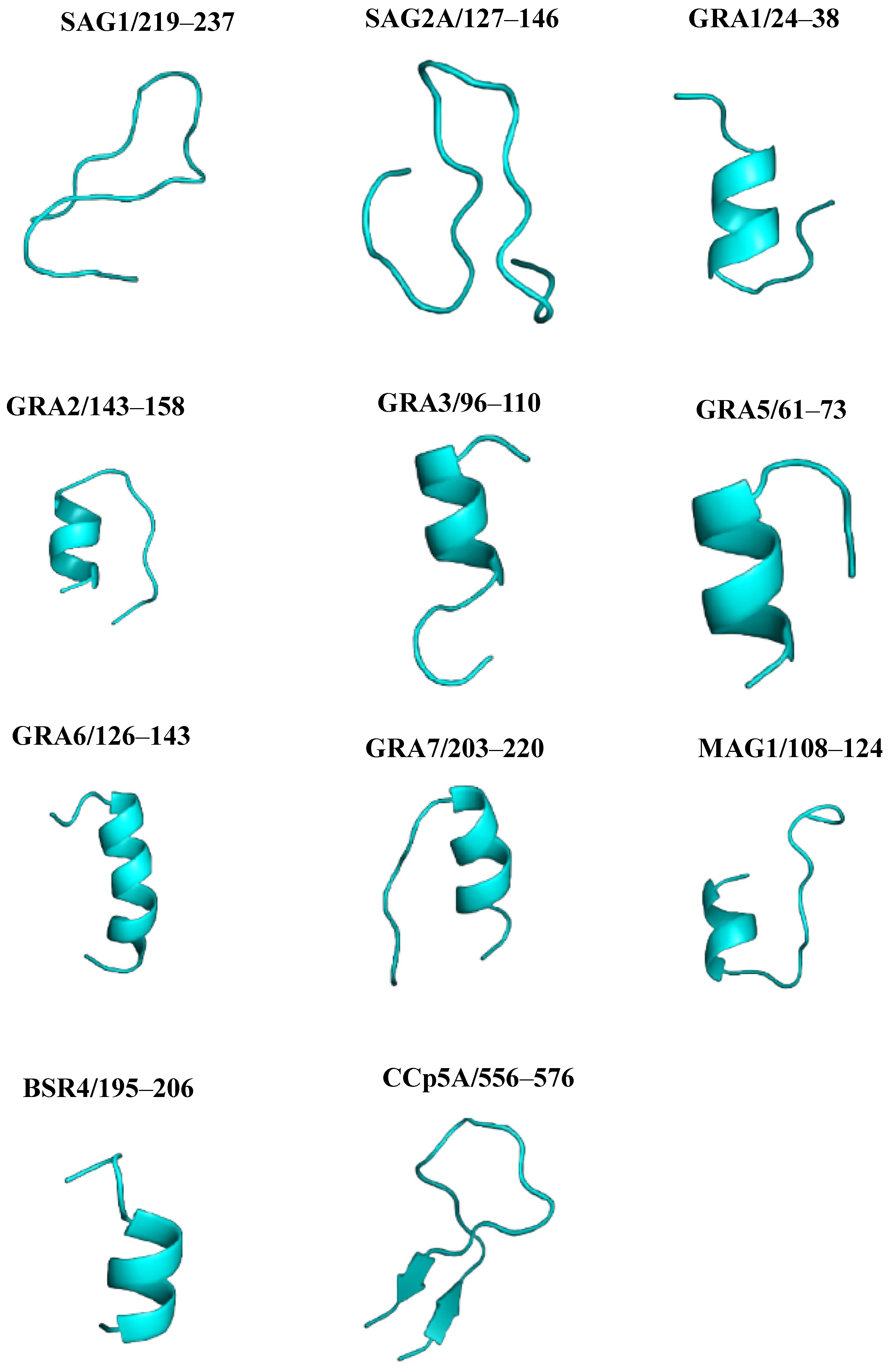
2.4. Three-Dimensional Modeling of Proteins and Validation
2.5. Three-Dimensional Modeling of Peptides and Validation
2.6. PYMOL Alignment
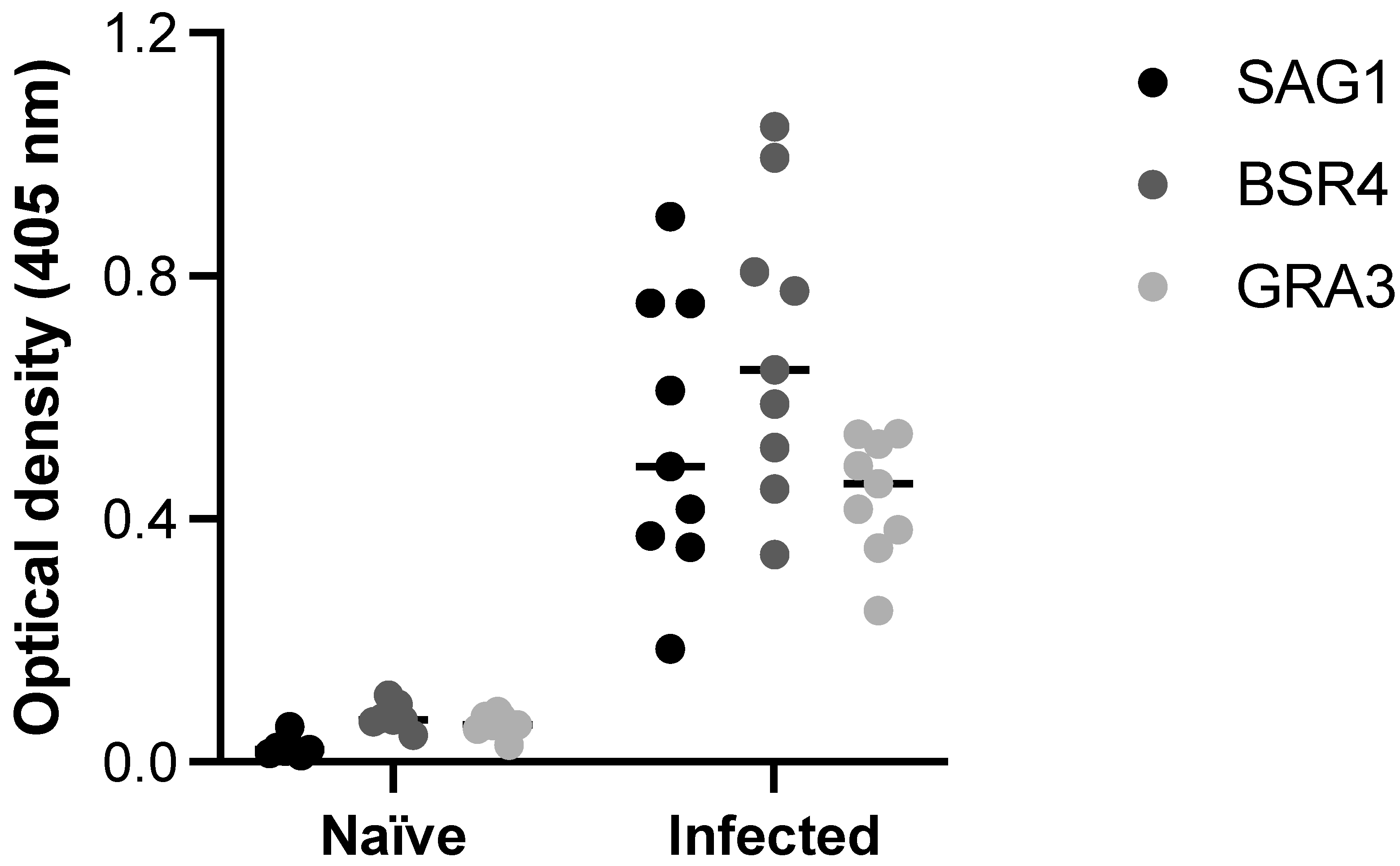
2.7. Biological Activity of the Selected Peptides
3. Discussion
4. Materials and Methods
4.1. Prediction and Analysis of Linear B-Cell Epitopes
4.2. Sequence Alignment and Estimation of Phylogenetic Trees
4.3. Prediction of Physicochemical Properties, Accessibility, and Antigenicity
4.4. Prediction of the Three-Dimensional (3D) Model of the T. gondii Proteins
4.5. Prediction of the Three-Dimensional (3D) Model of the T. gondii Peptides
4.6. Validation of the 3D Modeling of Proteins and Peptides
4.7. Three-Dimensional Structure Alignment of Proteins and Peptides
4.8. Serum Samples from Uninfected and Experimentally Infected Pigs
4.9. ELISA to Evaluate the Biological Activity of the Selected Peptides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Macêdo, A.G., Jr.; Cunha, J.P., Jr.; Cardoso., T.H.S.; Silva, M.V.; Santiago, F.M.; Silva, J.S.; Pirovani, C.P.; Silva, D.A.O.; Mineo, J.R.; Mineo, T.W.P. SAG2A protein from Toxoplasma gondii interacts with both innate and adaptive immune compartments of infected hosts. Parasit. Vectors 2013, 6, 163. [Google Scholar] [CrossRef]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [PubMed]
- Ben-Harari, R.R.; Connolly, M.P. High burden and low awareness of toxoplasmosis in the United States. Postgrad. Med. 2019, 131, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Robert-Gangneux, F.; Dardé, M.L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Teixeira, C.; Gomes, P.; Borges, M. Promising drug targets and compounds with anti-Toxoplasma gondii activity. Microorganisms 2021, 9, 1960. [Google Scholar] [CrossRef]
- Dubey, J.P. Outbreaks of clinical toxoplasmosis in humans: Five decades of personal experience, perspectives and lessons learned. Parasit. Vectors 2021, 14, 263. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, G.; Cai, J.P. Identifying novel B cell epitopes within Toxoplasma gondii GRA6. Korean J. Parasitol. 2016, 54, 431–437. [Google Scholar] [CrossRef]
- Dubey, J.P. Toxoplasmosis—An overview. Southeast Asian J. Trop. Med. Public Health 1991, 22, 88–92. [Google Scholar]
- Luft, B.J.; Remington, J.S. Toxoplasmic encephalitis in AIDS. Clin. Infect. Dis. 1992, 15, 211–222. [Google Scholar] [CrossRef]
- Ambroise-Thomas, P. Parasitic diseases and immunodeficiencies. Parasitology 2001, 122, S65–S71. [Google Scholar] [CrossRef]
- Tomasz Ferra, B.; Holec-Gąsior, L.; Gatkowska, J.; Dziadek, B.; Dzitko, K.; Grąźlewska, W.; Lautenbach, D. The first study on the usefulness of recombinant tetravalent chimeric proteins containing fragments of SAG2, GRA1, ROP1 and AMA1 antigens in the detection of specific anti-Toxoplasma gondii antibodies in mouse and human sera. PLoS ONE 2019, 14, e0217866. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0217866 (accessed on 23 April 2025).
- Ybañez, R.H.D.; Ybañez, A.P.; Nishikawa, Y. Review on the current trends of toxoplasmosis serodiagnosis in humans. Front. Cell. Infect. Microbiol. 2020, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Sudan, V.; Jaiswal, A.K.; Shanker, D. Recent trends in the diagnosis of toxoplasmosis. Clin. Rev. Opin. 2013, 5, 11–17. [Google Scholar] [CrossRef]
- Murat, J.B.; Hidalgo, H.F.; Brenier-Pinchart, M.P.; Pelloux, H. Human toxoplasmosis: Which biological diagnostic tests are best suited to which clinical situations? Expert Rev. Anti Infect. Ther. 2013, 11, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Roiko, M.S.; LaFavers, K.; Leland, D.; Arrizabalaga, G. Toxoplasma gondii-positive human sera recognize intracellular tachyzoites and bradyzoites with diverse patterns of immunoreactivity. Int. J. Parasitol. 2018, 48, 225–232. [Google Scholar] [CrossRef]
- Santana, S.S.; Paiva, C.F.; Carvalho, F.R.; Barros, H.L.S.; Silva, T.L.; Barros, P.S.C.; Pajuaba, A.C.A.M.; Barros, G.B.; Dietze, R.; Mineo, T.W.P.; et al. A peptide originated from Toxoplasma gondii microneme 8 displaying serological evidence to differentiate recent from chronic human infection. Parasitol. Int. 2021, 84, 102394. [Google Scholar] [CrossRef]
- Sun, X.; Lu, H.; Jia, B.; Chang, Z.; Peng, S.; Yin, J.; Chen, Q.; Jiang, N. A comparative study of Toxoplasma gondii seroprevalence in three healthy Chinese populations detected using native and recombinant antigens. Parasit. Vectors 2013, 6, 241. [Google Scholar] [CrossRef]
- Santana, S.S.; Gebrim, L.C.; Carvalho, F.R.; Barros, H.S.; Barros, P.C.; Pajuaba, A.C.A.M.; Messina, V.; Possenti, A.; Cherchi, S.; Reiche, E.M.V.; et al. CCp5A protein from Toxoplasma gondii as a serological marker of oocyst-driven infections in humans and domestic animals. Front. Microbiol. 2015, 6, 1305. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Bhattacharya, M.; Sharma, G.; Lee, S.S. Immunoinformatics approach for the identification and characterization of T cell and B cell epitopes towards the peptide-based vaccine against SARS-CoV-2. Arch. Med. Res. 2021, 52, 362–370. [Google Scholar] [CrossRef]
- Arranz-Solís, D.; Cordeiro, C.; Young, L.H.; Dardé, M.L.; Commodaro, A.G.; Grigg, M.E.; Saeij, J.P.J. Serotyping of Toxoplasma gondii infection using peptide membrane arrays. Front. Cell. Infect. Microbiol. 2019, 9, 408. [Google Scholar] [CrossRef]
- Kong, J.T.; Grigg, M.E.; Uyetake, L.; Parmley, S.; Boothroyd, J.C. Serotyping of Toxoplasma gondii infections in humans using synthetic peptides. J. Infect. Dis. 2003, 187, 1484–1495. [Google Scholar] [CrossRef] [PubMed]
- Lekutis, C.; Ferguson, D.J.P.; Grigg, M.E.; Camps, M.; Boothroyd, J.C. Surface antigens of Toxoplasma gondii: Variations on a theme. Inter. J. Parasitol. 2001, 31, 1285–1292. [Google Scholar] [CrossRef]
- Tomavo, S.; Schwarz, R.T.; Dubremetz, J.F. Evidence for glycosyl-phosphatidylinositol anchoring of Toxoplasma gondii major surface antigens. Mol. Cell. Biol. 1989, 9, 4576. [Google Scholar] [CrossRef]
- Jalallou, N.; Bandepour, M.; Khazan, H.; Haghighi, A.; Abdollahi, S.; Kazemi, B. Recombinant SAG1 antigen to detect Toxoplasma gondii specific Immunoglobulin G in human sera by ELISA test. Iran. J. Parasitol. 2010, 5, 1–9. [Google Scholar] [PubMed]
- Bel-Ochi, N.C.; Bouratbine, A.; Mousli, M. Enzyme-Linked Immunosorbent Assay using recombinant SAG1 antigen to detect Toxoplasma gondii-specific Immunoglobulin G antibodies in human sera and saliva. Clin. Vaccine Immunol. 2013, 20, 468–473. [Google Scholar] [CrossRef]
- Béla, S.R.; Silva, D.A.O.; Cunha-Júnior, J.P.; Pirovani, C.P.; Chaves-Borges, F.A.; Carvalho, F.R.; Oliveira, T.C.; Mineo, J.R. Use of SAG2A recombinant Toxoplasma gondii surface antigen as a diagnostic marker for human acute toxoplasmosis: Analysis of titers and avidity of IgG and IgG1 antibodies. Diagn. Microbiol. Infect. Dis. 2008, 62, 245–254. [Google Scholar] [CrossRef]
- Beghetto, E.; Spadoni, A.; Bruno, L.; Buffolano, W.; Gargano, N. Chimeric antigens of Toxoplasma gondii: Toward standardization of toxoplasmosis serodiagnosis using recombinant products. J. Clin. Microbiol. 2006, 44, 2133–2140. [Google Scholar] [CrossRef]
- Costa, J.G.; Vilariño, M.J. Semiquantitative dot blot with the GRA8 antigen to differentiate the stages of toxoplasmosis infection. J. Microbiol. Methods 2018, 149, 9–13. [Google Scholar] [CrossRef]
- Kotresha, D.; Poonam, D.; Hafiznur, Y.M.; Saadatnia, G.; Nurulhasanah, O.; Sabariah, O.; Tan, S.Y.; Zahidah, A.K.I.; Rahmah, N. Recombinant proteins from new constructs of SAG1 and GRA7 sequences and their usefulness to detect acute toxoplasmosis. Trop. Biomed. 2012, 29, 129–137. [Google Scholar]
- Pietkiewicz, H.; Hiszczyńska-Sawicka, E.; Kur, J.; Petersen, E.; Nielsen, H.V.; Stankiewicz, M.; Andrzejewska, I.; Myjak, P. Usefulness of Toxoplasma gondii-specific recombinant antigens in serodiagnosis of human toxoplasmosis. J. Clin. Microbiol. 2004, 42, 1779–1781. [Google Scholar] [CrossRef]
- Holec-Ga, L.; Kur, J.; Hiszczyńska-Sawicka, E. GRA2 and ROP1 recombinant antigens as potential markers for detection of Toxoplasma gondii-specific Immunoglobulin G in humans with acute toxoplasmosis. Clin. Vaccine Immunol. 2009, 16, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Holec-Ga, L.; Kur, J. Toxoplasma gondii: Recombinant GRA5 antigen for detection of immunoglobulin G antibodies using enzyme-linked immunosorbent assay. Exp. Parasitol. 2010, 124, 272–278. [Google Scholar] [CrossRef]
- Elyasi, H.; Babaie, J.; Fricker-Hidalgo, H.; Brenier-Pinchart, M.P.; Zare, M.; Sadeghiani, G.; Assmar, M.; Pelloux, H.; Golkar, M. Use of Dense Granule Antigen GRA6 in an Immunoglobulin G avidity test to exclude acute Toxoplasma gondii infection during pregnancy. Clin. Vaccine Immunol. 2010, 17, 1349–1355. [Google Scholar] [CrossRef]
- Xing-Zhi, C.; Guo, K.; Chen, Y.; Liu, L.-L.; Shen, J.-L.; Jiao, Y.-M.; Fang, Q.; Sun, X. Recombinant expression and immunologic identification of bradyzoite-specific antigen BSR4 of Toxoplasma gondii. Chin. J. Parasitol. Parasit. Dis. 2012, 30, 170–173. [Google Scholar]
- Di Cristina, M.; Del Porto, P.; Buffolano, W.; Beghetto, E.; Spadoni, A.; Guglietta, S.; Piccolella, E.; Felici, F.; Gargano, N. The Toxoplasma gondii bradyzoite antigens BAG1 and MAG1 induce early humoral and cell-mediated immune responses upon human infection. Microbes Infect. 2004, 6, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Holec, L.; Hiszczyńska-Sawicka, E.; Ga, A.; Brillowska-Da, A.; Kur, J. Use of MAG1 recombinant dntigen for Diagnosis of Toxoplasma gondii infection in humans. Clin. Vaccine Immunol. 2007, 14, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Ybañez, R.H.; Nishikawa, Y. Comparative performance of recombinant GRA6, GRA7, and GRA14 for the serodetection of T. gondii infection and analysis of IgG subclasses in human sera from the Philippines. Pathogens 2022, 11, 277. [Google Scholar] [CrossRef]
- Potocnakova, L.; Bhide, M.; Pulzova, L.B. An introduction to B-cell epitope mapping and in silico epitope prediction. J. Immunol. Res. 2016, 1, 6760830. [Google Scholar] [CrossRef]
- Sanchez-Trincado, J.L.; Gomez-Perosanz, M.; Reche, P.A. Fundamentals and methods for T- and B-cell epitope prediction. J. Immunol. Res. 2017, 1, 2680160. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools on the ExPASy Server, In 2-D Proteome Analysis Protocols; Link, A.J., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 1999. [Google Scholar] [CrossRef]
- Bai, Y.; He, S.; Zhao, G.; Chen, L.; Shi, N.; Zhou, H.; Cong, H.; Zhao, Q.; Zhu, X.-Q. Toxoplasma gondii: Bioinformatics analysis, cloning and expression of a novel protein TgIMP1. Exp. Parasitol. 2012, 132, 458–464. [Google Scholar] [CrossRef]
- Dardé, M.L. Toxoplasma gondii, “new” genotypes and virulence. Parasite 2008, 15, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Shaddel, M.; Ebrahimi, M.; Tabandeh, M.R. Bioinformatics analysis of single and multi-hybrid epitopes of GRA-1, GRA-4, GRA-6 and GRA-7 proteins to improve DNA vaccine design against Toxoplasma gondii. J. Parasit. Dis. 2018, 42, 269–276. [Google Scholar] [CrossRef]
- Javadi-Mamaghani, A.; Seyyed-Tabaei, S.J.; Ranjbar, M.M.; Haghighi, A.; Spotin, A.; Rezaee, H. Designing diagnostic kit for Toxoplasma gondii based on GRA7, SAG1, and ROP1 antigens: An in silico strategy. Int. J. Pept. Res. Ther. 2020, 26, 2269–2283. [Google Scholar] [CrossRef]
- Burg, J.L.; Perelman, D.; Kasper, L.H.; Ware, P.L.; Boothroyd, J.C. Molecular analysis of the gene encoding the major surface antigen of Toxoplasma gondii. J. Immunol. 1988, 141, 3584–3591. [Google Scholar] [CrossRef] [PubMed]
- Cardona, N.; de-la-Torre, A.; Siachoque, H.; Patarroyo, M.A.; Gomez-Marin, J.E. Toxoplasma gondii: P30 peptides recognition pattern in human toxoplasmosis. Exp. Parasitol. 2009, 123, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gąsior, L.; Ferra, B.; Grąźlewska, W. Toxoplasma gondii tetravalent chimeric proteins as novel antigens for detection of specific Immunoglobulin G in sera of small ruminants. Animals 2019, 9, 1146. [Google Scholar] [CrossRef]
- Holec-Ga, L.; Ferra, B.; Drapała, D. MIC1-MAG1-SAG1 chimeric protein, a most effective antigen for detection of human toxoplasmosis. Clin. Vaccine Immunol. 2012, 19, 1977–1979. [Google Scholar] [CrossRef]
- Dai, J.-F.; Jiang, M.; Qu, L.-L.; Sun, L.; Wang, Y.-Y.; Gong, L.-L.; Gong, R.-J.; Si, J. Toxoplasma gondii: Enzyme-linked immunosorbent assay based on a recombinant multi-epitope peptide for distinguishing recent from past infection in human sera. Exp. Parasitol. 2013, 133, 95–100. [Google Scholar] [CrossRef]
- Holec-Ga, L. Toxoplasma gondii recombinant antigens as tools for serodiagnosis of human toxoplasmosis: Current status of studies. Clin. Vaccine Immunol. 2013, 20, 1343–1351. [Google Scholar] [CrossRef]
- de Souza, M.Q.; Galdino, A.S.; Santos, J.C.D.; Soares, M.V.; de Nóbrega, Y.C.; Alvares, A.C.M.; de Freitas, S.M.; Torres, F.A.G.; Felipe, M.S.S. A recombinant multiepitope protein for hepatitis B diagnosis. BioMed Res. Inter. 2013, 1, 148317. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Emini, E.A.; Hughes, J.V.; Perlow, D.S.; Boger, J. Induction of hepatitis A virus-neutralizing antibody by a virus-specific synthetic peptide. J. Virol. 1985, 55, 836–839. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumor antigens and subunit vaccines. BMC Bioinform. 2007, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Chivian, D.; Baker, D. Protein structure prediction and analysis using the Robetta server. Nucleic Acids Res. 2004, 32, W526–W531. [Google Scholar] [CrossRef]
- Shen, Y.; Maupetit, J.; Derreumaux, P.; Tufféry, P. Improved PEP-FOLD approach for peptide and miniprotein structure prediction. J. Chem. Theory Comput. 2014, 10, 4745–4758. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.L.; MacArthur, M.W.; Hutchinson, E.G.; Thornton, J.M. Stereochemical quality of protein structure coordinates. Proteins 1992, 12, 345–364. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef]
- Pontius, J.; Richelle, J.; Wodak, S.J. Deviations from standard atomic volumes as a quality measure for protein crystal structures. J. Mol. Biol. 1996, 264, 121–136. [Google Scholar] [CrossRef]
- Garcia, J.L.; Gennari, S.M.; Navarro, I.T.; Machado, R.Z.; Sinhorini, I.L.; Freire, R.L.; Marana, E.R.M.; Tsutsui, V.; Contente, A.P.A.; Begale, L.P. Partial protection against tissue cysts formation in pigs vaccinated with crude rhoptry proteins of Toxoplasma gondii. Vet. Parasitol. 2005, 129, 209–217. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Peptide Identification | Peptide Sequences | Length | Position |
|---|---|---|---|---|
| SAG1 | Pep1 | GVKVPQDNNQYCSGTTLTG | 19 | 219–237 |
| SAG2A | Pep2 | EAGAPAGRNNDGGSSAPTPK | 20 | 127–146 |
| GRA1 | Pep3 | AAEGGDNQSSAVSDR | 15 | 24–38 |
| GRA2 | Pep4 | NVESDRSTTTTQAPDS | 16 | 143–158 |
| GRA3 | Pep5 | DAESDKVDNQAEGGE | 15 | 96–110 |
| GRA5 | Pep6 | FERGRAAVTGHPV | 13 | 61–73 |
| GRA6 | Pep7 | QGTRRRYSSVQEPQAKVP | 18 | 126–143 |
| GRA7 | Pep8 | RTYRHFSPRKNRSRQPAL | 18 | 203–220 |
| MAG1 | Pep9 | CEEQQEQGDTTLSDHDF | 17 | 108–124 |
| BSR4 | Pep10 | TSAARGTESGKT | 12 | 195–206 |
| CCp5A | Pep11 | NAYADCSASSEESDVYGCAAG | 21 | 556–576 |
| Peptide | Molecular Weight (Da) | Theoretic pI | Aliphatic Index | Instability Index | GRAVY |
|---|---|---|---|---|---|
| SAG1 | 1982.15 | 5.83 | 51.05 | 17.03 | −0.721 |
| SAG2A | 1853.92 | 6.17 | 20.00 | 56.12 | −1.195 |
| GRA1 | 1463.44 | 4.03 | 39.33 | 49.64 | −1.040 |
| GRA2 | 1708.71 | 4.03 | 24.38 | 58.81 | −1.425 |
| GRA3 | 1563.51 | 3.71 | 32.67 | −0.99 | −1.713 |
| GRA5 | 1396.57 | 9.61 | 60.00 | 2.24 | −0.308 |
| GRA6 | 2087.33 | 10.90 | 37.78 | 112.97 | −1.578 |
| GRA7 | 2270.59 | 12.18 | 27.22 | 92.42 | −1.944 |
| MAG1 | 1981.98 | 3.71 | 22.94 | 60.63 | −1.659 |
| BSR4 | 1165.23 | 8.41 | 16.67 | 20.04 | −1.067 |
| CCp5A | 2070.10 | 3.43 | 37.62 | 52.16 | −0.281 |
| Peptide | VaxiJen 2.0 Antigenicity Value |
|---|---|
| SAG1 | 1.1163 |
| SAG2A | 1.2736 |
| GRA1 | 1.2842 |
| GRA2 | 1.0249 |
| GRA3 | 1.2841 |
| GRA5 | 0.7496 |
| GRA6 | 0.3953 |
| GRA7 | 0.6110 |
| MAG1 | 1.2375 |
| BSR4 | 1.4757 |
| CCp5A | 1.3231 |
| PROTEINS | Whole Molecule | Peptide | ||
|---|---|---|---|---|
| ERRAT | PROVE | ERRAT | PROVE | |
| SAG1 | 90.7975 | 1.2% | 100 | 5.0% |
| SAG2A | 91.4286 | 4.5% | 100 | 4.8% |
| GRA1 | 99.4413 | 3.3% | 100 | 0% |
| GRA2 | 91.3295 | 3.2% | 100 | 0% |
| GRA3 | 99.0566 | 1.9% | 100 | 0% |
| GRA5 | 100 | 0.8% | 100 | 0% |
| GRA6 | 92.2330 | 3.1% | 100 | 0% |
| GRA7 | 97.7974 | 2.7% | 100 | 4.0% |
| MAG1 | 92.0635 | 3.0% | 100 | 5.0% |
| BSR4 | 97.1204 | 3.7% | 100 | 0% |
| CCp5A | 89.4467 | 4.2% | 100 | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Betancourt, A.d.V.B.; Silva, T.L.; de Freitas Oliveira, D.K.; Nicolau-Junior, N.; Garcia, J.L.; Fujiwara, R.T.; Mineo, T.W.P.; Mineo, J.R. A Structural In Silico Analysis of Novel Epitopes from Toxoplasma gondii Proteins for the Serodiagnosis of Toxoplasmosis. Int. J. Mol. Sci. 2025, 26, 4689. https://doi.org/10.3390/ijms26104689
Betancourt AdVB, Silva TL, de Freitas Oliveira DK, Nicolau-Junior N, Garcia JL, Fujiwara RT, Mineo TWP, Mineo JR. A Structural In Silico Analysis of Novel Epitopes from Toxoplasma gondii Proteins for the Serodiagnosis of Toxoplasmosis. International Journal of Molecular Sciences. 2025; 26(10):4689. https://doi.org/10.3390/ijms26104689
Chicago/Turabian StyleBetancourt, Angelis del Valle Benitez, Tamires Lopes Silva, Débora Karolla de Freitas Oliveira, Nilson Nicolau-Junior, João Luis Garcia, Ricardo Toshio Fujiwara, Tiago Wilson Patriarca Mineo, and José Roberto Mineo. 2025. "A Structural In Silico Analysis of Novel Epitopes from Toxoplasma gondii Proteins for the Serodiagnosis of Toxoplasmosis" International Journal of Molecular Sciences 26, no. 10: 4689. https://doi.org/10.3390/ijms26104689
APA StyleBetancourt, A. d. V. B., Silva, T. L., de Freitas Oliveira, D. K., Nicolau-Junior, N., Garcia, J. L., Fujiwara, R. T., Mineo, T. W. P., & Mineo, J. R. (2025). A Structural In Silico Analysis of Novel Epitopes from Toxoplasma gondii Proteins for the Serodiagnosis of Toxoplasmosis. International Journal of Molecular Sciences, 26(10), 4689. https://doi.org/10.3390/ijms26104689