
From Childhood Woes to Adult Blues: Unmasking the Role of Early Traumas, P2X7 Receptor, and Neuroinflammation in Anxiety and Depression

 ,
,
Abstract
1. Introduction
2. Early-Life Trauma and Neuroinflammation: Setting the Stage
2.1. The Potential Role of Neuroinflammation in the Lasting Effects of Early-Life Traumas
2.2. The Role of Early-Life Stress in Shaping Neuroimmune Responses
3. P2X7 Receptor: A Central Hub in Neuroinflammation
3.1. Role of P2X7R in ATP Signalling
3.2. The Role of P2X7R in Neuroinflammatory Processes
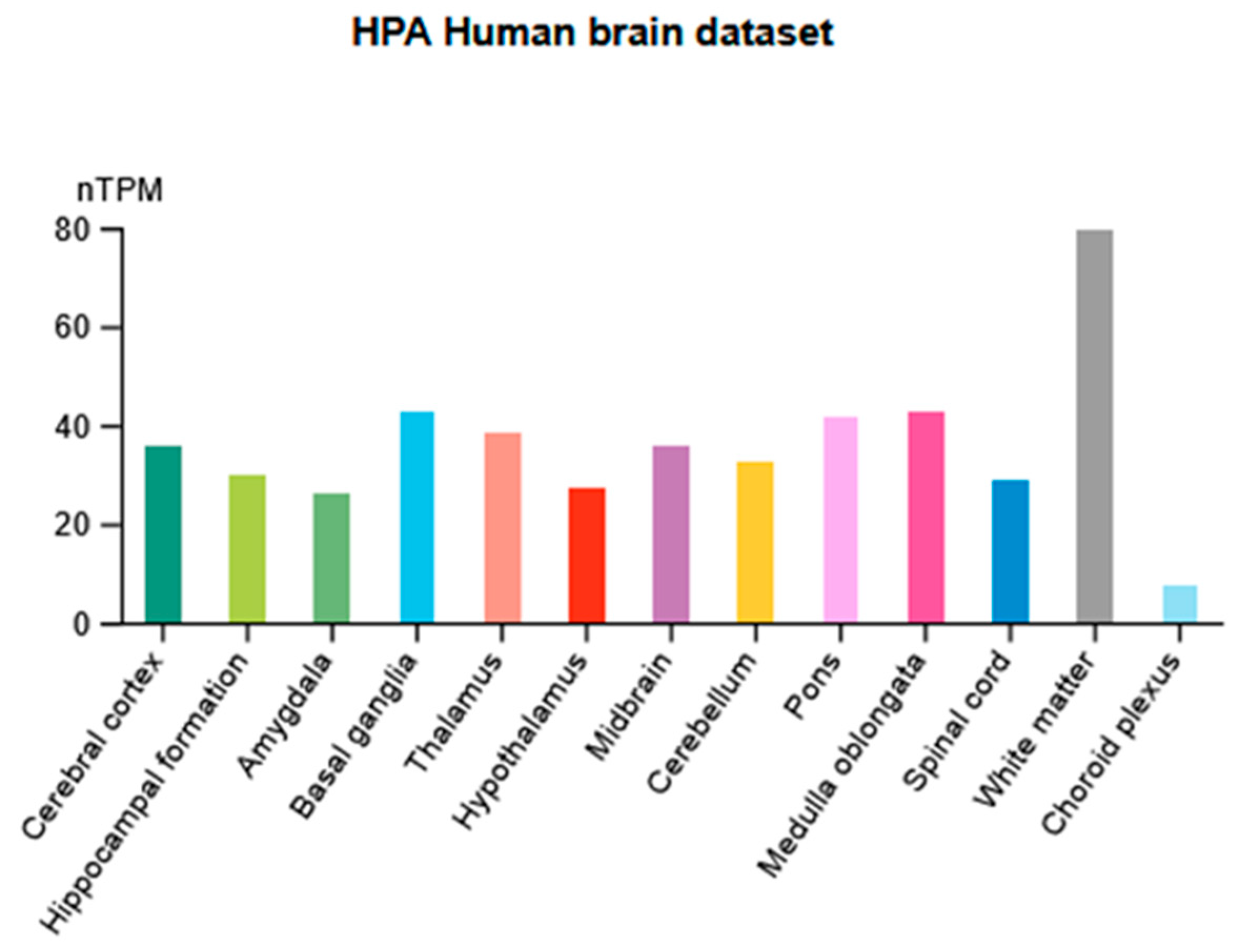
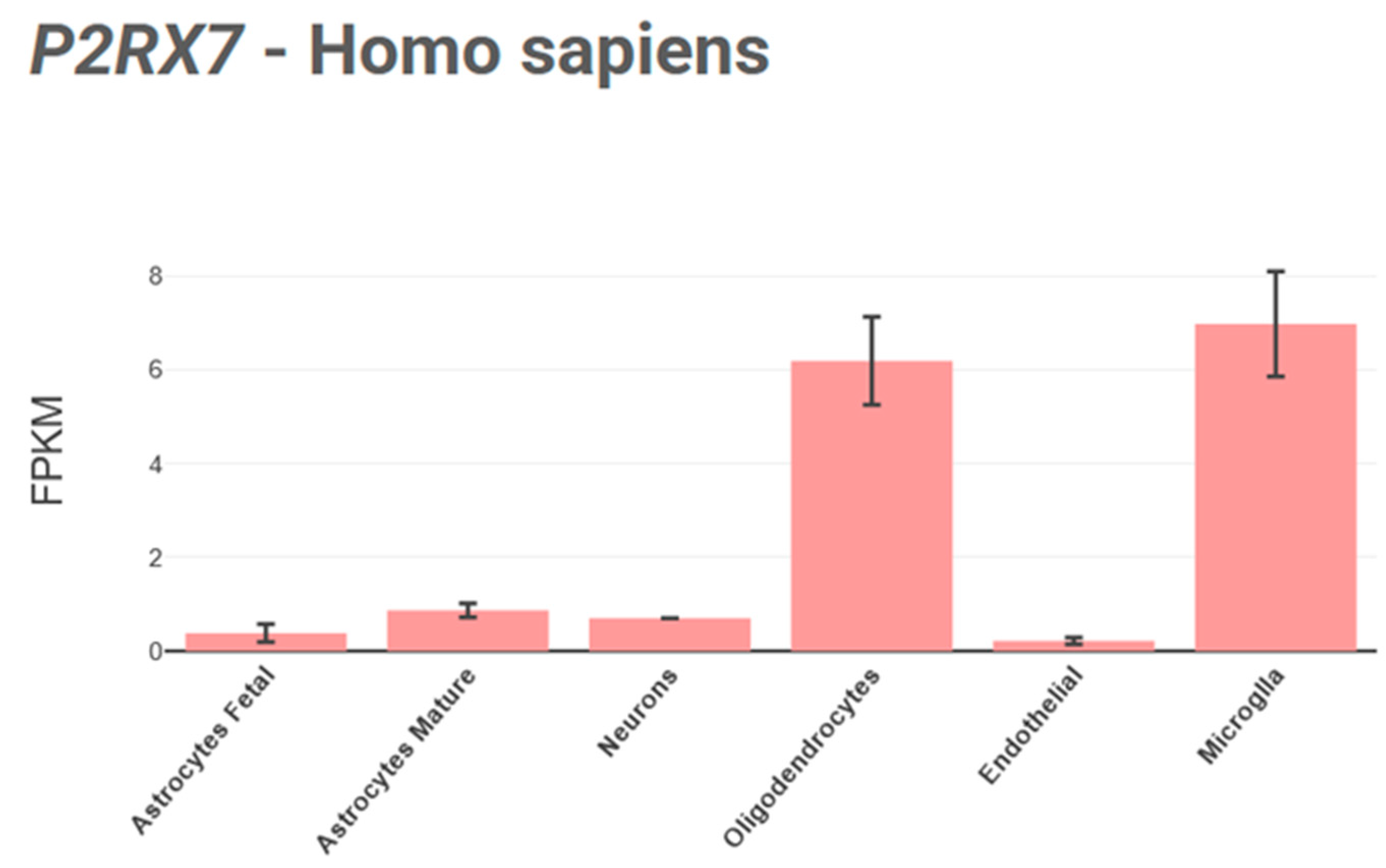
3.3. P2X7 Receptor Distribution and Function in the CNS
3.4. P2X7R’s Involvement in Other Neurological and Psychiatric Conditions
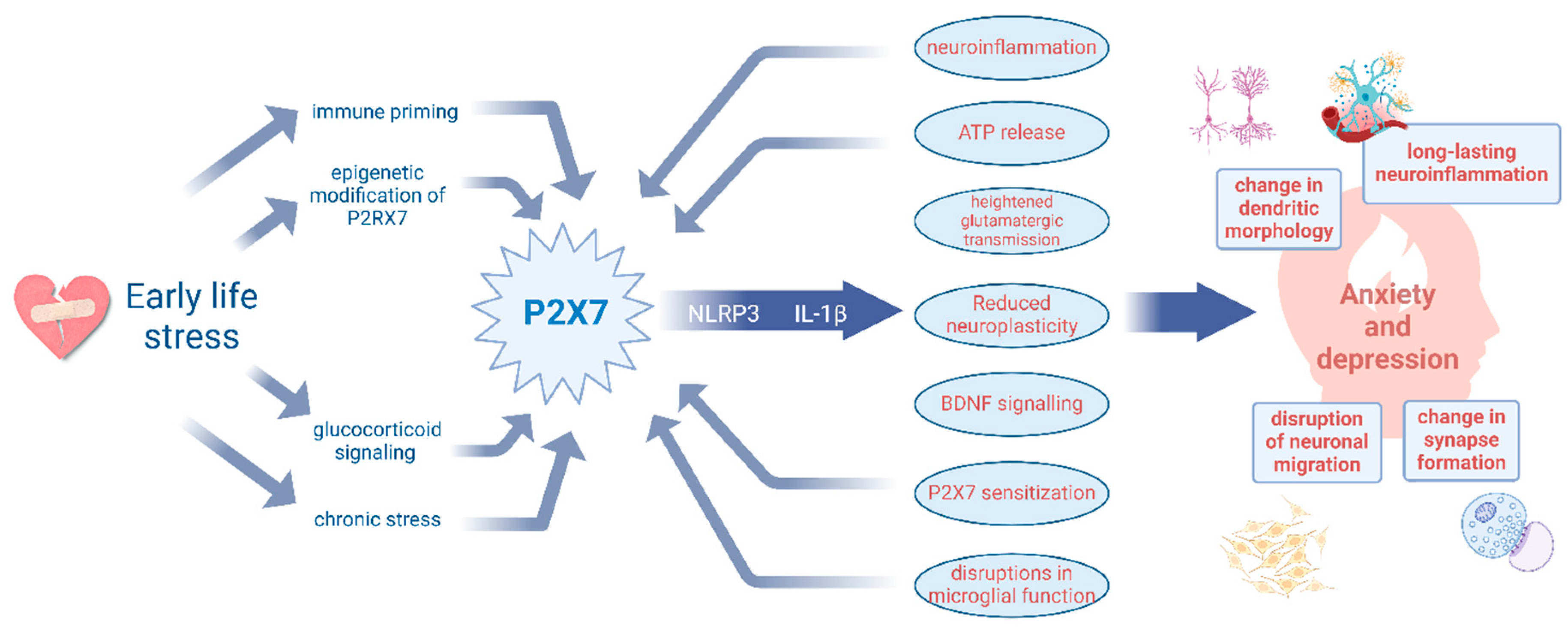
4. P2X7R Activation: A Bridge Between Early Trauma and Anxio-Depressive Behaviours
4.1. Early-Life Stress Leads to Prolonged Enhanced Immune Activation
4.2. Neuroinflammation and Disruption of Brain Development
4.3. Early-Life Immune Activation Leads to Long-Term Neuropsychiatric Disorders
4.4. The Potential Role of the P2RX7 Gene in the Pathogenesis of Depression and Anxiety

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant ID (SNP) | Associated Condition | Effect of Function | Notes | References |
|---|---|---|---|---|
| rs2230912 | MDD BD Anxiety | Gain-of-function | One of the most studied SNPs; associated with increased inflammatory response, increased pore activity of P2X7, increased receptor function | [113,124,130,131,132] |
| rs1718119 | MDD BD (rapid cycling) | Gain-of-function | Associated with increased cytokine release, increased pore activity of P2X7, increased receptor function | [116,131] |
| rs1653625 | MDD BD | Unknown | Possible influence on gene expression through miRNA-mediated regulation | [116,132] |
| rs3751143 | MDD BD | Loss-of-function | Diminished receptor activity and associated with altered immune function | [116,133] |
| rs7958311 | MDD | Gain-of-function and Loss-of-function | Enhances the P2X7 receptor’s channel activity, impairs the receptor’s ability to form large pores (a critical function for processes such as cytokine release and cell death) | [134] |
| rs208294 | MD BD | Gain-of- function | Increases both channel and pore functions of the P2X7 receptor | [125,126] |
| rs67881993 | Anxiety | Unknown | Modulating anxiety symptoms through gene–environment interactions conveying a protective effect against increased anxiety in individuals exposed to ELA | [122] |
| rs74892325 | MDD | Unknown | Modulating the impact of environmental stressors on mood regulation; the presence of the minor allele was associated with a protective effect against increased depression severity in individuals exposed to ELA | [121] |
| rs11615992 | Suicide risk | Regulatory role variant | Potential role in modulating the effects of early-childhood maltreatment on suicidal ideation (protective effect) | [123] |
| rs78473339 | Suicide risk | Unknown | Potential role in modulating the effects of early-childhood maltreatment on hopelessness (protective effect) | [123] |
5. P2X7R Modulation and Its Therapeutic Potential in Anxiety and Depression
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Nettis, M.A.; Pariante, C.M.; Mondelli, V. Early-Life Adversity, Systemic Inflammation and Comorbid Physical and Psychiatric Illnesses of Adult Life. Curr. Top. Behav. Neurosci. 2020, 44, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.E.; White, S.F.; Chen, E.; Nusslock, R. Association of Inflammatory Activity With Larger Neural Responses to Threat and Reward Among Children Living in Poverty. Am. J. Psychiatry 2021, 178, 313–320. [Google Scholar] [CrossRef]
- Mattei, D.; Notter, T. Basic Concept of Microglia Biology and Neuroinflammation in Relation to Psychiatry. Curr. Top. Behav. Neurosci. 2020, 44, 9–34. [Google Scholar] [CrossRef]
- Liu, J.; Liu, T.T.; Mou, L.; Zhang, Y.; Chen, X.; Wang, Q.; Deng, B.L.; Liu, J. P2X7 receptor: A potential target for treating comorbid anxiety and depression. Purinergic Signal. 2024, 1–11. [Google Scholar] [CrossRef]
- Wohleb, E.S. Neuron-Microglia Interactions in Mental Health Disorders: “For Better, and For Worse”. Front. Immunol. 2016, 7, 544. [Google Scholar] [CrossRef]
- Dantzer, R.; Cohen, S.; Russo, S.J.; Dinan, T.G. Resilience and immunity. Brain Behav. Immun. 2018, 74, 28–42. [Google Scholar] [CrossRef]
- Johnson, J.D.; Barnard, D.F.; Kulp, A.C.; Mehta, D.M. Neuroendocrine Regulation of Brain Cytokines After Psychological Stress. J. Endocr. Soc. 2019, 3, 1302–1320. [Google Scholar] [CrossRef]
- Kokkosis, A.G.; Madeira, M.M.; Hage, Z.; Valais, K.; Koliatsis, D.; Resutov, E.; Tsirka, S.E. Chronic psychosocial stress triggers microglial-/macrophage-induced inflammatory responses leading to neuronal dysfunction and depressive-related behavior. Glia 2024, 72, 111–132. [Google Scholar] [CrossRef]
- Sequeira, M.K.; Bolton, J.L. Stressed Microglia: Neuroendocrine-Neuroimmune Interactions in the Stress Response. Endocrinology 2023, 164, bqad088. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Biber, K. The microglial ATP-gated ion channel P2X7 as a CNS drug target. Glia 2016, 64, 1772–1787. [Google Scholar] [CrossRef]
- Jiang, L.H.; Caseley, E.A.; Muench, S.P.; Roger, S. Structural basis for the functional properties of the P2X7 receptor for extracellular ATP. Purinergic Signal. 2021, 17, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Caseley, E.A.; Muench, S.P.; Roger, S.; Mao, H.J.; Baldwin, S.A.; Jiang, L.H. Non-synonymous single nucleotide polymorphisms in the P2X receptor genes: Association with diseases, impact on receptor functions and potential use as diagnosis biomarkers. Int. J. Mol. Sci. 2014, 15, 13344–13371. [Google Scholar] [CrossRef] [PubMed]
- Vinci, M.; Vitello, G.A.; Greco, D.; Treccarichi, S.; Ragalmuto, A.; Musumeci, A.; Fallea, A.; Federico, C.; Calì, F.; Saccone, S.; et al. Next Generation Sequencing and Electromyography Reveal the Involvement of the. Curr. Issues Mol. Biol. 2024, 46, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Lu, Y.; Zhang, Q.; Liu, W.; Yang, R.; Jiao, J.; Liu, J.; Gao, G.; Yang, H. Piperine promotes autophagy flux by P2RX4 activation in SNCA/α-synuclein-induced Parkinson disease model. Autophagy 2022, 18, 559–575. [Google Scholar] [CrossRef]
- Li, Z.; Huang, Z.; Zhang, H.; Lu, J.; Tian, Y.; Wei, Y.; Yang, Y.; Bai, L. P2X7 Receptor Induces Pyroptotic Inflammation and Cartilage Degradation in Osteoarthritis via NF-κB/NLRP3 Crosstalk. Oxid. Med. Cell Longev. 2021, 2021, 8868361. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, L.; Zhang, Y.; Wang, H.; Sun, S.; Wu, L.A. Involvement of microglial P2X7 receptor in pain modulation. CNS Neurosci. Ther. 2024, 30, e14496. [Google Scholar] [CrossRef]
- Wang, M.; Pan, W.; Xu, Y.; Zhang, J.; Wan, J.; Jiang, H. Microglia-Mediated Neuroinflammation: A Potential Target for the Treatment of Cardiovascular Diseases. J. Inflamm. Res. 2022, 15, 3083–3094. [Google Scholar] [CrossRef]
- Jiang, L.H.; Baldwin, J.M.; Roger, S.; Baldwin, S.A. Insights into the Molecular Mechanisms Underlying Mammalian P2X7 Receptor Functions and Contributions in Diseases, Revealed by Structural Modeling and Single Nucleotide Polymorphisms. Front. Pharmacol. 2013, 4, 55. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Stefani, C.; Badiale, G.; Campione, G.; Martini, F.; Tognon, M. The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications. Cancers 2022, 14, 1116. [Google Scholar] [CrossRef] [PubMed]
- Sluyter, R.; Adriouch, S.; Fuller, S.J.; Nicke, A.; Sophocleous, R.A.; Watson, D. Animal Models for the Investigation of P2X7 Receptors. Int. J. Mol. Sci. 2023, 24, 8225. [Google Scholar] [CrossRef]
- Csölle, C.; Andó, R.D.; Kittel, Á.; Gölöncsér, F.; Baranyi, M.; Soproni, K.; Zelena, D.; Haller, J.; Németh, T.; Mócsai, A.; et al. The absence of P2X7 receptors (P2rx7) on non-haematopoietic cells leads to selective alteration in mood-related behaviour with dysregulated gene expression and stress reactivity in mice. Int. J. Neuropsychopharmacol. 2013, 16, 213–233. [Google Scholar] [CrossRef] [PubMed]
- Sidoryk-Węgrzynowicz, M.; Strużyńska, L. Astroglial and Microglial Purinergic P2X7 Receptor as a Major Contributor to Neuroinflammation during the Course of Multiple Sclerosis. Int. J. Mol. Sci. 2021, 22, 8404. [Google Scholar] [CrossRef] [PubMed]
- Thawkar, B.S.; Kaur, G. Inhibitors of NF-κB and P2X7/NLRP3/Caspase 1 pathway in microglia: Novel therapeutic opportunities in neuroinflammation induced early-stage Alzheimer’s disease. J. Neuroimmunol. 2019, 326, 62–74. [Google Scholar] [CrossRef]
- Engel, T. The P2X7 Receptor as a Mechanistic Biomarker for Epilepsy. Int. J. Mol. Sci. 2023, 24, 5410. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, H.Y.; Rubini, P.; Tang, Y.; Illes, P. A Possible Causal Involvement of Neuroinflammatory, Purinergic P2X7 Receptors in Psychiatric Disorders. Curr. Neuropharmacol. 2022, 20, 2142–2155. [Google Scholar] [CrossRef]
- Szabó, D.; Tod, P.; Gölöncsér, F.; Román, V.; Lendvai, B.; Otrokocsi, L.; Sperlágh, B. Maternal P2X7 receptor inhibition prevents autism-like phenotype in male mouse offspring through the NLRP3-IL-1β pathway. Brain Behav. Immun. 2022, 101, 318–332. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Jones, D.N.C. Emerging role of the P2X7-NLRP3-IL1beta pathway in mood disorders. Psychoneuroendocrinology 2018, 98, 95–100. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Wang, Q.; Ao, H.; Shoblock, J.R.; Lord, B.; Aluisio, L.; Fraser, I.; Nepomuceno, D.; Neff, R.A.; Welty, N.; et al. Pharmacological characterization of a novel centrally permeable P2X7 receptor antagonist: JNJ-47965567. Br. J. Pharmacol. 2013, 170, 624–640. [Google Scholar] [CrossRef]
- Monif, M.; Burnstock, G.; Williams, D.A. Microglia: Proliferation and activation driven by the P2X7 receptor. Int. J. Biochem. Cell Biol. 2010, 42, 1753–1756. [Google Scholar] [CrossRef]
- Ribeiro, D.E.; Roncalho, A.L.; Glaser, T.; Ulrich, H.; Wegener, G.; Joca, S. P2X7 Receptor Signaling in Stress and Depression. Int. J. Mol. Sci. 2019, 20, 2778. [Google Scholar] [CrossRef]
- Janke, K.L.; Cominski, T.P.; Kuzhikandathil, E.V.; Servatius, R.J.; Pang, K.C. Investigating the Role of Hippocampal BDNF in Anxiety Vulnerability Using Classical Eyeblink Conditioning. Front. Psychiatry 2015, 6, 106. [Google Scholar] [CrossRef] [PubMed]
- Rana, T.; Behl, T.; Sehgal, A.; Srivastava, P.; Bungau, S. Unfolding the Role of BDNF as a Biomarker for Treatment of Depression. J. Mol. Neurosci. 2021, 71, 2008–2021. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.W.; Walser, S.M.; Dedic, N.; Aprile-Garcia, F.; Jakubcakova, V.; Adamczyk, M.; Webb, K.J.; Uhr, M.; Refojo, D.; Schmidt, M.V. Heterozygosity for the mood disorder-associated variant Gln460Arg alters P2X7 receptor function and sleep quality. J. Neurosci. 2017, 37, 11688–11700. [Google Scholar] [CrossRef] [PubMed]
- von Muecke-Heim, I.A.; Ries, C.; Urbina, L.; Deussing, J.M. P2X7R antagonists in chronic stress-based depression models: A review. Eur. Arch. Psychiatry Clin. Neurosci. 2021, 271, 1343–1358. [Google Scholar] [CrossRef]
- Urbina-Treviño, L.; von Mücke-Heim, I.A.; Deussing, J.M. P2X7 Receptor-Related Genetic Mouse Models—Tools for Translational Research in Psychiatry. Front. Neural Circuits 2022, 16, 876304. [Google Scholar] [CrossRef]
- von Mücke-Heim, I.A.; Deussing, J.M. The P2X7 receptor in mood disorders: Emerging target in immunopsychiatry, from bench to bedside. Neuropharmacology 2022, 224, 109366. [Google Scholar] [CrossRef]
- Bauer, M.E.; Teixeira, A.L. Neuroinflammation in Mood Disorders: Role of Regulatory Immune Cells. Neuroimmunomodulation 2021, 28, 99–107. [Google Scholar] [CrossRef]
- Chang, J.; Jiang, T.; Shan, X.; Zhang, M.; Li, Y.; Qi, X.; Bian, Y.; Zhao, L. Pro-inflammatory cytokines in stress-induced depression: Novel insights into mechanisms and promising therapeutic strategies. Prog. Neuropsychopharmacol. Biol. Psychiatry 2024, 131, 110931. [Google Scholar] [CrossRef]
- Hostinar, C.E.; Nusslock, R.; Miller, G.E. Future Directions in the Study of Early-Life Stress and Physical and Emotional Health: Implications of the Neuroimmune Network Hypothesis. J. Clin. Child. Adolesc. Psychol. 2018, 47, 142–156. [Google Scholar] [CrossRef]
- Baumeister, D.; Akhtar, R.; Ciufolini, S.; Pariante, C.M.; Mondelli, V. Childhood trauma and adulthood inflammation: A meta-analysis of peripheral C-reactive protein, interleukin-6 and tumour necrosis factor-α. Mol. Psychiatry 2016, 21, 642–649. [Google Scholar] [CrossRef]
- Miller, G.E.; Cole, S.W. Clustering of depression and inflammation in adolescents previously exposed to childhood adversity. Biol. Psychiatry 2012, 72, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Nusslock, R.; Miller, G.E. Early-Life Adversity and Physical and Emotional Health Across the Lifespan: A Neuroimmune Network Hypothesis. Biol. Psychiatry 2016, 80, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.V.; Wang, X.D.; Meijer, O.C. Early life stress paradigms in rodents: Potential animal models of depression? Psychopharmacology 2011, 214, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Zanta, N.C.; Assad, N.; Suchecki, D. Neurobiological mechanisms involved in maternal deprivation-induced behaviours relevant to psychiatric disorders. Front. Mol. Neurosci. 2023, 16, 1099284. [Google Scholar] [CrossRef]
- Orso, R.; Creutzberg, K.C.; Wearick-Silva, L.E.; Wendt Viola, T.; Tractenberg, S.G.; Benetti, F.; Grassi-Oliveira, R. How Early Life Stress Impact Maternal Care: A Systematic Review of Rodent Studies. Front. Behav. Neurosci. 2019, 13, 197. [Google Scholar] [CrossRef]
- Birnie, M.T.; Kooiker, C.L.; Short, A.K.; Bolton, J.L.; Chen, Y.; Baram, T.Z. Plasticity of the Reward Circuitry After Early-Life Adversity: Mechanisms and Significance. Biol. Psychiatry 2020, 87, 875–884. [Google Scholar] [CrossRef]
- Tzanoulinou, S.; Sandi, C. The Programming of the Social Brain by Stress During Childhood and Adolescence: From Rodents to Humans. Curr. Top. Behav. Neurosci. 2017, 30, 411–429. [Google Scholar] [CrossRef]
- Slopen, N.; Kubzansky, L.D.; McLaughlin, K.A.; Koenen, K.C. Childhood adversity and inflammatory processes in youth: A prospective study. Psychoneuroendocrinology 2013, 38, 188–200. [Google Scholar] [CrossRef]
- Danese, A.; Caspi, A.; Williams, B.; Ambler, A.; Sugden, K.; Mika, J.; Werts, H.; Freeman, J.; Pariante, C.M.; Moffitt, T.E.; et al. Biological embedding of stress through inflammation processes in childhood. Mol. Psychiatry 2011, 16, 244–246. [Google Scholar] [CrossRef]
- Slopen, N.; Loucks, E.B.; Appleton, A.A.; Kawachi, I.; Kubzansky, L.D.; Non, A.L.; Buka, S.; Gilman, S.E. Early origins of inflammation: An examination of prenatal and childhood social adversity in a prospective cohort study. Psychoneuroendocrinology 2015, 51, 403–413. [Google Scholar] [CrossRef]
- Delpech, J.C.; Wei, L.; Hao, J.; Yu, X.; Madore, C.; Butovsky, O.; Kaffman, A. Early life stress perturbs the maturation of microglia in the developing hippocampus. Brain Behav. Immun. 2016, 57, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Simen, A.; Mane, S.; Kaffman, A. Early life stress inhibits expression of a novel innate immune pathway in the developing hippocampus. Neuropsychopharmacology 2012, 37, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; David, A.; Duman, R.S.; Anisman, H.; Kaffman, A. Early life stress increases anxiety-like behavior in Balb c mice despite a compensatory increase in levels of postnatal maternal care. Horm. Behav. 2010, 57, 396–404. [Google Scholar] [CrossRef]
- Coelho, R.; Viola, T.W.; Walss-Bass, C.; Brietzke, E.; Grassi-Oliveira, R. Childhood maltreatment and inflammatory markers: A systematic review. Acta Psychiatr. Scand. 2014, 129, 180–192. [Google Scholar] [CrossRef]
- Zhou, L.; Wu, Z.; Wang, G.; Xiao, L.; Wang, H.; Sun, L.; Xie, Y. Long-term maternal separation potentiates depressive-like behaviours and neuroinflammation in adult male C57/BL6J mice. Pharmacol. Biochem. Behav. 2020, 196, 172953. [Google Scholar] [CrossRef]
- Roque, A.; Ochoa-Zarzosa, A.; Torner, L. Maternal separation activates microglial cells and induces an inflammatory response in the hippocampus of male rat pups, independently of hypothalamic and peripheral cytokine levels. Brain Behav. Immun. 2016, 55, 39–48. [Google Scholar] [CrossRef]
- Harry, G.J.; Kraft, A.D. Microglia in the developing brain: A potential target with lifetime effects. Neurotoxicology 2012, 33, 191–206. [Google Scholar] [CrossRef]
- Schafer, D.P.; Stevens, B. Microglia Function in Central Nervous System Development and Plasticity. Cold Spring Harb. Perspect. Biol. 2015, 7, a020545. [Google Scholar] [CrossRef]
- Johnson, F.K.; Kaffman, A. Early life stress perturbs the function of microglia in the developing rodent brain: New insights and future challenges. Brain Behav. Immun. 2018, 69, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Paolicelli, R.C.; Sforazzini, F.; Weinhard, L.; Bolasco, G.; Pagani, F.; Vyssotski, A.L.; Bifone, A.; Gozzi, A.; Ragozzino, D.; et al. Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat. Neurosci. 2014, 17, 400–406. [Google Scholar] [CrossRef]
- Dawidowski, B.; Górniak, A.; Podwalski, P.; Lebiecka, Z.; Misiak, B.; Samochowiec, J. The Role of Cytokines in the Pathogenesis of Schizophrenia. J. Clin. Med. 2021, 10, 3849. [Google Scholar] [CrossRef] [PubMed]
- Andrejew, R.; Oliveira-Giacomelli, Á.; Ribeiro, D.E.; Glaser, T.; Arnaud-Sampaio, V.F.; Lameu, C.; Ulrich, H. The P2X7 Receptor: Central Hub of Brain Diseases. Front. Mol. Neurosci. 2020, 13, 124. [Google Scholar] [CrossRef] [PubMed]
- Fields, R.D. Nonsynaptic and nonvesicular ATP release from neurons and relevance to neuron-glia signaling. Semin. Cell Dev. Biol. 2011, 22, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, S. Synchronization of Ca2+ oscillations: Involvement of ATP release in astrocytes. FEBS J. 2010, 277, 286–292. [Google Scholar] [CrossRef]
- Chen, Y.H.; Lin, S.; Jin, S.Y.; Gao, T.M. Extracellular ATP Is a Homeostatic Messenger That Mediates Cell-Cell Communication in Physiological Processes and Psychiatric Diseases. Biol. Psychiatry 2025, 97, 41–53. [Google Scholar] [CrossRef]
- Giuliani, A.L.; Sarti, A.C.; Di Virgilio, F. Ectonucleotidases in Acute and Chronic Inflammation. Front. Pharmacol. 2020, 11, 619458. [Google Scholar] [CrossRef]
- Rodrigues, R.J.; Tomé, A.R.; Cunha, R.A. ATP as a multi-target danger signal in the brain. Front. Neurosci. 2015, 9, 148. [Google Scholar] [CrossRef]
- Sperlágh, B.; Illes, P. P2X7 receptor: An emerging target in central nervous system diseases. Trends Pharmacol. Sci. 2014, 35, 537–547. [Google Scholar] [CrossRef]
- Bidula, S.; Dhuna, K.; Helliwell, R.; Stokes, L. Positive allosteric modulation of P2X7 promotes apoptotic cell death over lytic cell death responses in macrophages. Cell Death Dis. 2019, 10, 882. [Google Scholar] [CrossRef]
- Savio, L.E.B.; de Andrade Mello, P.; da Silva, C.G.; Coutinho-Silva, R. The P2X7 Receptor in Inflammatory Diseases: Angel or Demon? Front. Pharmacol. 2018, 9, 52. [Google Scholar] [CrossRef]
- Grassi, F.; Salina, G. The P2X7 Receptor in Autoimmunity. Int. J. Mol. Sci. 2023, 24, 14116. [Google Scholar] [CrossRef] [PubMed]
- Grassi, F.; Marino, R. The P2X7 receptor in mucosal adaptive immunity. Purinergic Signal. 2024, 20, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Wiley, J.S.; Sluyter, R.; Gu, B.J.; Stokes, L.; Fuller, S.J. The human P2X7 receptor and its role in innate immunity. Tissue Antigens 2011, 78, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Acioglu, C.; Heary, R.F.; Elkabes, S. Roles of neuronal toll-like receptors in neuropathic pain and central nervous system injuries and diseases. Brain Behav. Immun. 2022, 102, 163–178. [Google Scholar] [CrossRef]
- Ospelt, C.; Gay, S. TLRs and chronic inflammation. Int. J. Biochem. Cell Biol. 2010, 42, 495–505. [Google Scholar] [CrossRef]
- Mouihate, A.; Galic, M.A.; Ellis, S.L.; Spencer, S.J.; Tsutsui, S.; Pittman, Q.J. Early life activation of toll-like receptor 4 reprograms neural anti-inflammatory pathways. J. Neurosci. 2010, 30, 7975–7983. [Google Scholar] [CrossRef]
- Viola, T.W.; Creutzberg, K.C.; Zaparte, A.; Kestering-Ferreira, É.; Tractenberg, S.G.; Centeno-Silva, A.; Orso, R.; Lumertz, F.S.; Brietzke, E.; Wearick-Silva, L.E.; et al. Acute neuroinflammation elicited by TLR-3 systemic activation combined with early life stress induces working memory impairments in male adolescent mice. Behav. Brain Res. 2019, 376, 112221. [Google Scholar] [CrossRef]
- Wang, M.; Krauss, J.L.; Domon, H.; Hosur, K.B.; Liang, S.; Magotti, P.; Triantafilou, M.; Triantafilou, K.; Lambris, J.D.; Hajishengallis, G. Microbial hijacking of complement-toll-like receptor crosstalk. Sci. Signal. 2010, 3, ra11. [Google Scholar] [CrossRef]
- Raby, A.C.; Holst, B.; Davies, J.; Colmont, C.; Laumonnier, Y.; Coles, B.; Shah, S.; Hall, J.; Topley, N.; Köhl, J.; et al. TLR activation enhances C5a-induced pro-inflammatory responses by negatively modulating the second C5a receptor, C5L2. Eur. J. Immunol. 2011, 41, 2741–2752. [Google Scholar] [CrossRef]
- Karmakar, M.; Katsnelson, M.A.; Dubyak, G.R.; Pearlman, E. Neutrophil P2X7 receptors mediate NLRP3 inflammasome-dependent IL-1β secretion in response to ATP. Nat. Commun. 2016, 7, 10555. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Li, K.-Y.; Lan, Y.; Zeng, H.-Y.; Chen, S.-Q.; Wang, H. NLRP3 Inflammasome: A key contributor to the inflammation formation. Food Chem. Toxicol. 2023, 174, 113683. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The role of interleukin-1 in general pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef] [PubMed]
- Yue, N.; Huang, H.; Zhu, X.; Han, Q.; Wang, Y.; Li, B.; Liu, Q.; Wu, G.; Zhang, Y.; Yu, J. Activation of P2X7 receptor and NLRP3 inflammasome assembly in hippocampal glial cells mediates chronic stress-induced depressive-like behaviors. J. Neuroinflammation 2017, 14, 102. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Tang, Y.; Sperlagh, B. Glial Purinergic Signaling-Mediated Oxidative Stress (GPOS) in Neuropsychiatric Disorders. Oxid. Med. Cell Longev. 2022, 2022, 1075440. [Google Scholar] [CrossRef]
- Zhao, Y.F.; Tang, Y.; Illes, P. Astrocytic and Oligodendrocytic P2X7 Receptors Determine Neuronal Functions in the CNS. Front. Mol. Neurosci. 2021, 14, 641570. [Google Scholar] [CrossRef]
- Campagno, K.E.; Mitchell, C.H. The P2X(7) Receptor in Microglial Cells Modulates the Endolysosomal Axis, Autophagy, and Phagocytosis. Front. Cell Neurosci. 2021, 15, 645244. [Google Scholar] [CrossRef]
- Uhlén, M.; Karlsson, M.J.; Hober, A.; Svensson, A.S.; Scheffel, J.; Kotol, D.; Zhong, W.; Tebani, A.; Strandberg, L.; Edfors, F.; et al. The human secretome. Sci. Signal. 2019, 12, eaaz0274. [Google Scholar] [CrossRef]
- Karlsson, M.; Zhang, C.; Méar, L.; Zhong, W.; Digre, A.; Katona, B.; Sjöstedt, E.; Butler, L.; Odeberg, J.; Dusart, P.; et al. A single-cell type transcriptomics map of human tissues. Sci. Adv. 2021, 7, eabh2169. [Google Scholar] [CrossRef]
- He, Y.; Taylor, N.; Fourgeaud, L.; Bhattacharya, A. The role of microglial P2X7: Modulation of cell death and cytokine release. J. Neuroinflammation 2017, 14, 135. [Google Scholar] [CrossRef]
- Kaczmarek-Hajek, K.; Zhang, J.; Kopp, R.; Grosche, A.; Rissiek, B.; Saul, A.; Bruzzone, S.; Engel, T.; Jooss, T.; Krautloher, A. Re-evaluation of neuronal P2X7 expression using novel mouse models and a P2X7-specific nanobody. eLife 2018, 7, e36217. [Google Scholar] [CrossRef]
- Illes, P.; Khan, T.M.; Rubini, P. Neuronal P2X7 receptors revisited: Do they really exist? J. Neurosci. 2017, 37, 7049–7062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sloan, S.A.; Clarke, L.E.; Caneda, C.; Plaza, C.A.; Blumenthal, P.D.; Vogel, H.; Steinberg, G.K.; Edwards, M.S.; Li, G.; et al. Purification and Characterization of Progenitor and Mature Human Astrocytes Reveals Transcriptional and Functional Differences with Mouse. Neuron 2016, 89, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Engel, T.; Nicke, A.; Deussing, J.M.; Sperlagh, B.; Diaz-Hernandez, M. Editorial: P2X7 as Common Therapeutic Target in Brain Diseases. Front. Mol. Neurosci. 2021, 14, 656011. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Ceusters, M. Targeting neuroinflammation with brain penetrant P2X7 antagonists as novel therapeutics for neuropsychiatric disorders. Neuropsychopharmacology 2020, 45, 234–235. [Google Scholar] [CrossRef] [PubMed]
- Territo, P.R.; Zarrinmayeh, H. P2X(7) Receptors in Neurodegeneration: Potential Therapeutic Applications From Basic to Clinical Approaches. Front. Cell Neurosci. 2021, 15, 617036. [Google Scholar] [CrossRef]
- Calzaferri, F.; Ruiz-Ruiz, C.; de Diego, A.M.G.; de Pascual, R.; Méndez-López, I.; Cano-Abad, M.F.; Maneu, V.; de Los Ríos, C.; Gandía, L.; García, A.G. The purinergic P2X7 receptor as a potential drug target to combat neuroinflammation in neurodegenerative diseases. Med. Res. Rev. 2020, 40, 2427–2465. [Google Scholar] [CrossRef]
- Kim, J.; Suh, Y.-H.; Chang, K.-A. Interleukin-17 induced by cumulative mild stress promoted depression-like behaviors in young adult mice. Mol. Brain 2021, 14, 11. [Google Scholar] [CrossRef]
- Sorrells, S.F.; Caso, J.R.; Munhoz, C.D.; Sapolsky, R.M. The stressed CNS: When glucocorticoids aggravate inflammation. Neuron 2009, 64, 33–39. [Google Scholar] [CrossRef]
- Andersen, S.L. Neuroinflammation, Early-Life Adversity, and Brain Development. Harv. Rev. Psychiatry 2022, 30, 24–39. [Google Scholar] [CrossRef]
- Busillo, J.M.; Azzam, K.M.; Cidlowski, J.A. Glucocorticoids sensitize the innate immune system through regulation of the NLRP3 inflammasome. J. Biol. Chem. 2011, 286, 38703–38713. [Google Scholar] [CrossRef]
- Calcia, M.A.; Bonsall, D.R.; Bloomfield, P.S.; Selvaraj, S.; Barichello, T.; Howes, O.D. Stress and neuroinflammation: A systematic review of the effects of stress on microglia and the implications for mental illness. Psychopharmacology 2016, 233, 1637–1650. [Google Scholar] [CrossRef] [PubMed]
- Perlman, W.R.; Webster, M.J.; Herman, M.M.; Kleinman, J.E.; Weickert, C.S. Age-related differences in glucocorticoid receptor mRNA levels in the human brain. Neurobiol. Aging 2007, 28, 447–458. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Magarinos, A.M. Stress effects on morphology and function of the hippocampus. Ann. N. Y. Acad. Sci. 1997, 821, 271–284. [Google Scholar] [CrossRef]
- Gottfried, C.; Bambini-Junior, V. Insights into the Relationship of the Immune System with Neurodevelopmental and Psychiatric Disorders. Neuroimmunomodulation 2018, 25, 243–245. [Google Scholar] [CrossRef]
- Hashimoto, O.; Kuniishi, H.; Nakatake, Y.; Yamada, M.; Wada, K.; Sekiguchi, M. Early life stress from allergic dermatitis causes depressive-like behaviors in adolescent male mice through neuroinflammatory priming. Brain Behav. Immun. 2020, 90, 319–331. [Google Scholar] [CrossRef]
- da Silva, G.L.; Sperotto, N.D.; Borges, T.J.; Bonorino, C.; Takyia, C.M.; Coutinho-Silva, R.; Campos, M.M.; Zanin, R.F.; Morrone, F.B. P2X7 receptor is required for neutrophil accumulation in a mouse model of irritant contact dermatitis. Exp. Dermatol. 2013, 22, 184–188. [Google Scholar] [CrossRef]
- Mizawa, M.; Yamaguchi, M.; Ueda, C.; Makino, T.; Shimizu, T. Stress evaluation in adult patients with atopic dermatitis using salivary cortisol. Biomed. Res. Int. 2013, 2013, 138027. [Google Scholar] [CrossRef]
- Silverberg, J.I.; Gelfand, J.M.; Margolis, D.J.; Boguniewicz, M.; Fonacier, L.; Grayson, M.H.; Ong, P.Y.; Chiesa Fuxench, Z.C.; Simpson, E.L. Symptoms and diagnosis of anxiety and depression in atopic dermatitis in U.S. adults. Br. J. Dermatol. 2019, 181, 554–565. [Google Scholar] [CrossRef]
- North, R.A. Molecular physiology of P2X receptors. Physiol. Rev. 2002, 82, 1013–1067. [Google Scholar] [CrossRef]
- Erhardt, A.; Lucae, S.; Unschuld, P.G.; Ising, M.; Kern, N.; Salyakina, D.; Lieb, R.; Uhr, M.; Binder, E.B.; Keck, M.E.; et al. Association of polymorphisms in P2RX7 and CaMKKb with anxiety disorders. J. Affect. Disord. 2007, 101, 159–168. [Google Scholar] [CrossRef]
- Feng, Y.; Li, X.; Zeng, R.; Gorodeski, G. Endogenously expressed truncated P2X7 receptor lacking the C-terminus is preferentially upregulated in epithelial cancer cells and fails to mediate ligand-induced pore formation and apoptosis. Nucleosides Nucleotides Nucleic Acids 2006, 25, 1271–1276. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, E.; Cirillo, M.; Woltersdorf, R.; Falzoni, S.; Chiozzi, P.; Pellegatti, P.; Callegari, M.G.; Sandona, D.; Markwardt, F.; Schmalzing, G. Trophic activity of a naturally occurring truncated isoform of the P2X7 receptor. FASEB J. 2010, 24, 3393–3404. [Google Scholar] [CrossRef] [PubMed]
- Czamara, D.; Muller-Myhsok, B.; Lucae, S. The P2RX7 polymorphism rs2230912 is associated with depression: A meta-analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 82, 272–277. [Google Scholar] [CrossRef]
- Stokes, L.; Fuller, S.J.; Sluyter, R.; Skarratt, K.K.; Gu, B.J.; Wiley, J.S. Two haplotypes of the P2X(7) receptor containing the Ala-348 to Thr polymorphism exhibit a gain-of-function effect and enhanced interleukin-1beta secretion. FASEB J. 2010, 24, 2916–2927. [Google Scholar] [CrossRef]
- Bennett, M.R. Synaptic P2X7 receptor regenerative-loop hypothesis for depression. Aust. N. Z. J. Psychiatry 2007, 41, 563–571. [Google Scholar] [CrossRef]
- Vereczkei, A.; Abdul-Rahman, O.; Halmai, Z.; Nagy, G.; Szekely, A.; Somogyi, A.; Faludi, G.; Nemoda, Z. Association of purinergic receptor P2RX7 gene polymorphisms with depression symptoms. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 92, 207–216. [Google Scholar] [CrossRef]
- Arabzadeh, S.; Ameli, N.; Zeinoddini, A.; Rezaei, F.; Farokhnia, M.; Mohammadinejad, P.; Ghaleiha, A.; Akhondzadeh, S. Celecoxib adjunctive therapy for acute bipolar mania: A randomized, double-blind, placebo-controlled trial. Bipolar Disord. 2015, 17, 606–614. [Google Scholar] [CrossRef]
- Leboyer, M.; Oliveira, J.; Tamouza, R.; Groc, L. Is it time for immunopsychiatry in psychotic disorders? Psychopharmacology 2016, 233, 1651–1660. [Google Scholar] [CrossRef]
- Ursu, D.; Ebert, P.; Langron, E.; Ruble, C.; Munsie, L.; Zou, W.; Fijal, B.; Qian, Y.W.; McNearney, T.A.; Mogg, A.; et al. Gain and loss of function of P2X7 receptors: Mechanisms, pharmacology and relevance to diabetic neuropathic pain. Mol. Pain. 2014, 10, 37. [Google Scholar] [CrossRef]
- Winham, S.J.; Bobo, W.V.; Liu, J.; Coombes, B.; Backlund, L.; Frye, M.A.; Biernacka, J.M.; Schalling, M.; Lavebratt, C. Sex-specific effects of gain-of-function P2RX7 variation on bipolar disorder. J. Affect. Disord. 2019, 245, 597–601. [Google Scholar] [CrossRef]
- Kristof, Z.; Eszlari, N.; Sutori, S.; Gal, Z.; Torok, D.; Baksa, D.; Petschner, P.; Sperlagh, B.; Anderson, I.M.; Deakin, J.F.W.; et al. P2RX7 gene variation mediates the effect of childhood adversity and recent stress on the severity of depressive symptoms. PLoS ONE 2021, 16, e0252766. [Google Scholar] [CrossRef] [PubMed]
- Kristof, Z.; Gal, Z.; Torok, D.; Eszlari, N.; Sutori, S.; Erdelyi-Hamza, B.; Petschner, P.; Sperlagh, B.; Anderson, I.M.; Deakin, J.F.W.; et al. Variation along P2RX7 interacts with early traumas on severity of anxiety suggesting a role for neuroinflammation. Sci. Rep. 2023, 13, 7757. [Google Scholar] [CrossRef] [PubMed]
- Kristof, Z.; Gal, Z.; Torok, D.; Eszlari, N.; Sutori, S.; Sperlagh, B.; Anderson, I.M.; Deakin, B.; Bagdy, G.; Juhasz, G.; et al. Embers of the Past: Early Childhood Traumas Interact with Variation in P2RX7 Gene Implicated in Neuroinflammation on Markers of Current Suicide Risk. Int. J. Mol. Sci. 2024, 25, 865. [Google Scholar] [CrossRef] [PubMed]
- Hejjas, K.; Szekely, A.; Domotor, E.; Halmai, Z.; Balogh, G.; Schilling, B.; Sarosi, A.; Faludi, G.; Sasvari-Szekely, M.; Nemoda, Z. Association between depression and the Gln460Arg polymorphism of P2RX7 gene: A dimensional approach. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2009, 150B, 295–299. [Google Scholar] [CrossRef]
- Soronen, P.; Mantere, O.; Melartin, T.; Suominen, K.; Vuorilehto, M.; Rytsälä, H.; Arvilommi, P.; Holma, I.; Holma, M.; Jylhä, P.; et al. P2RX7 gene is associated consistently with mood disorders and predicts clinical outcome in three clinical cohorts. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2011, 156B, 435–447. [Google Scholar] [CrossRef]
- Mantere, O.; Soronen, P.; Uher, R.; Ketokivi, M.; Jylhä, P.; Melartin, T.; Paunio, T.; Isometsä, E. Neuroticism mediates the effect of P2RX7 on outcomes of mood disorders. Depress. Anxiety 2012, 29, 816–823. [Google Scholar] [CrossRef]
- Deussing, J.M.; Arzt, E. P2X7 Receptor: A Potential Therapeutic Target for Depression? Trends Mol. Med. 2018, 24, 736–747. [Google Scholar] [CrossRef]
- Su, W.J.; Zhang, T.; Jiang, C.L.; Wang, W. Clemastine Alleviates Depressive-Like Behavior Through Reversing the Imbalance of Microglia-Related Pro-inflammatory State in Mouse Hippocampus. Front. Cell Neurosci. 2018, 12, 412. [Google Scholar] [CrossRef]
- Tan, S.; Wang, Y.; Chen, K.; Long, Z.; Zou, J. Ketamine Alleviates Depressive-Like Behaviors via Down-Regulating Inflammatory Cytokines Induced by Chronic Restraint Stress in Mice. Biol. Pharm. Bull. 2017, 40, 1260–1267. [Google Scholar] [CrossRef]
- Lucae, S.; Salyakina, D.; Barden, N.; Harvey, M.; Gagné, B.; Labbé, M.; Binder, E.B.; Uhr, M.; Paez-Pereda, M.; Sillaber, I.; et al. P2RX7, a gene coding for a purinergic ligand-gated ion channel, is associated with major depressive disorder. Hum. Mol. Genet. 2006, 15, 2438–2445. [Google Scholar] [CrossRef]
- Backlund, L.; Lavebratt, C.; Frisén, L.; Nikamo, P.; Hukic Sudic, D.; Träskman-Bendz, L.; Landén, M.; Edman, G.; Vawter, M.P.; Ösby, U.; et al. P2RX7: Expression responds to sleep deprivation and associates with rapid cycling in bipolar disorder type 1. PLoS ONE 2012, 7, e43057. [Google Scholar] [CrossRef] [PubMed]
- Halmai, Z.; Dome, P.; Vereczkei, A.; Abdul-Rahman, O.; Szekely, A.; Gonda, X.; Faludi, G.; Sasvari-Szekely, M.; Nemoda, Z. Associations between depression severity and purinergic receptor P2RX7 gene polymorphisms. J. Affect. Disord. 2013, 150, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Fuller, S.J.; Stokes, L.; Skarratt, K.K.; Gu, B.J.; Wiley, J.S. Genetics of the P2X7 receptor and human disease. Purinergic Signal. 2009, 5, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Gonda, X.; Hullam, G.; Antal, P.; Eszlari, N.; Petschner, P.; Hökfelt, T.G.; Anderson, I.M.; Deakin, J.F.W.; Juhasz, G.; Bagdy, G. Significance of risk polymorphisms for depression depends on stress exposure. Sci. Rep. 2018, 8, 3946. [Google Scholar] [CrossRef]
- Basso, A.M.; Bratcher, N.A.; Harris, R.R.; Jarvis, M.F.; Decker, M.W.; Rueter, L.E. Behavioral profile of P2X7 receptor knockout mice in animal models of depression and anxiety: Relevance for neuropsychiatric disorders. Behav. Brain Res. 2009, 198, 83–90. [Google Scholar] [CrossRef]
- Boucher, A.A.; Arnold, J.C.; Hunt, G.E.; Spiro, A.; Spencer, J.; Brown, C.; McGregor, I.S.; Bennett, M.R.; Kassiou, M. Resilience and reduced c-Fos expression in P2X7 receptor knockout mice exposed to repeated forced swim test. Neuroscience 2011, 189, 170–177. [Google Scholar] [CrossRef]
- Domingos, L.B.; Hott, S.C.; Terzian, A.L.B.; Resstel, L.B.M. P2X7 purinergic receptors participate in the expression and extinction processes of contextual fear conditioning memory in mice. Neuropharmacology 2018, 128, 474–481. [Google Scholar] [CrossRef]
- Xie, B.; Chen, Y.; Zhang, S.; Wu, X.; Zhang, Z.; Peng, Y.; Huang, X. The expression of P2X7 receptors on peripheral blood mononuclear cells in patients with primary Sjögren’s syndrome and its correlation with anxiety and depression. Clin. Exp. Rheumatol. 2014, 32, 354–360. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kristof, Z.; Szabo, D.; Sperlagh, B.; Torok, D.; Gonda, X. From Childhood Woes to Adult Blues: Unmasking the Role of Early Traumas, P2X7 Receptor, and Neuroinflammation in Anxiety and Depression. Int. J. Mol. Sci. 2025, 26, 4687. https://doi.org/10.3390/ijms26104687
Kristof Z, Szabo D, Sperlagh B, Torok D, Gonda X. From Childhood Woes to Adult Blues: Unmasking the Role of Early Traumas, P2X7 Receptor, and Neuroinflammation in Anxiety and Depression. International Journal of Molecular Sciences. 2025; 26(10):4687. https://doi.org/10.3390/ijms26104687
Chicago/Turabian StyleKristof, Zsuliet, Dorottya Szabo, Beata Sperlagh, Dora Torok, and Xenia Gonda. 2025. "From Childhood Woes to Adult Blues: Unmasking the Role of Early Traumas, P2X7 Receptor, and Neuroinflammation in Anxiety and Depression" International Journal of Molecular Sciences 26, no. 10: 4687. https://doi.org/10.3390/ijms26104687
APA StyleKristof, Z., Szabo, D., Sperlagh, B., Torok, D., & Gonda, X. (2025). From Childhood Woes to Adult Blues: Unmasking the Role of Early Traumas, P2X7 Receptor, and Neuroinflammation in Anxiety and Depression. International Journal of Molecular Sciences, 26(10), 4687. https://doi.org/10.3390/ijms26104687