
Effects of Exercise Training on Cardiac Mitochondrial Functions in Diabetic Heart: A Systematic Review



Abstract

1. Introduction
2. Materials and Methods
2.1. Protocol and Registration
2.2. Information Sources
2.3. Study Design
2.4. Animal Model Type
2.5. Intervention Focus
2.6. Comparators
2.7. Outcomes
2.8. Search Strategy
2.9. Selection Process
2.10. Data Management
2.11. Data Collection Process
2.12. Data Items
2.13. Outcomes and Prioritization
2.14. Risk of Bias Assessment
2.15. Data Synthesis
3. Results
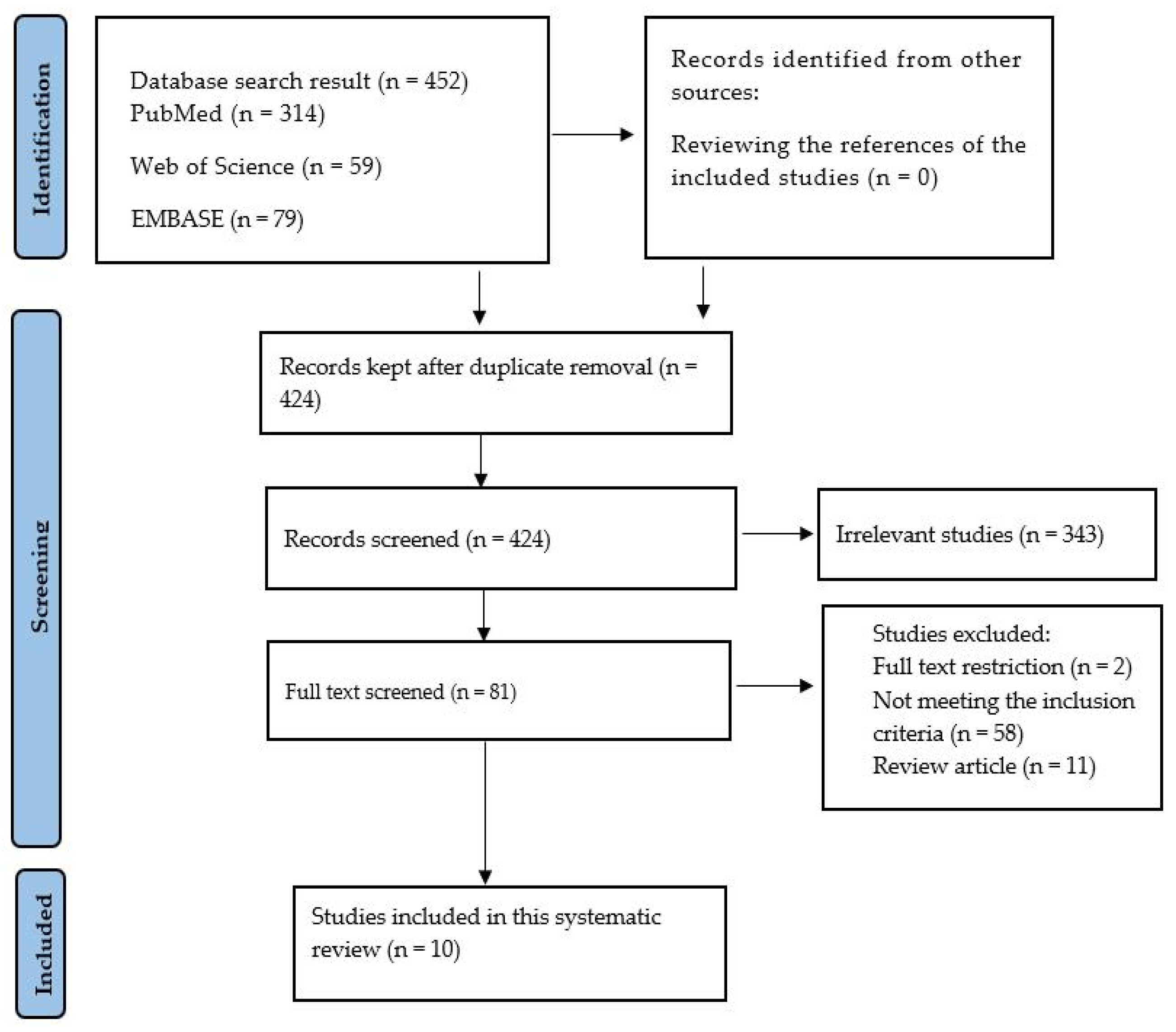
3.1. Search Results
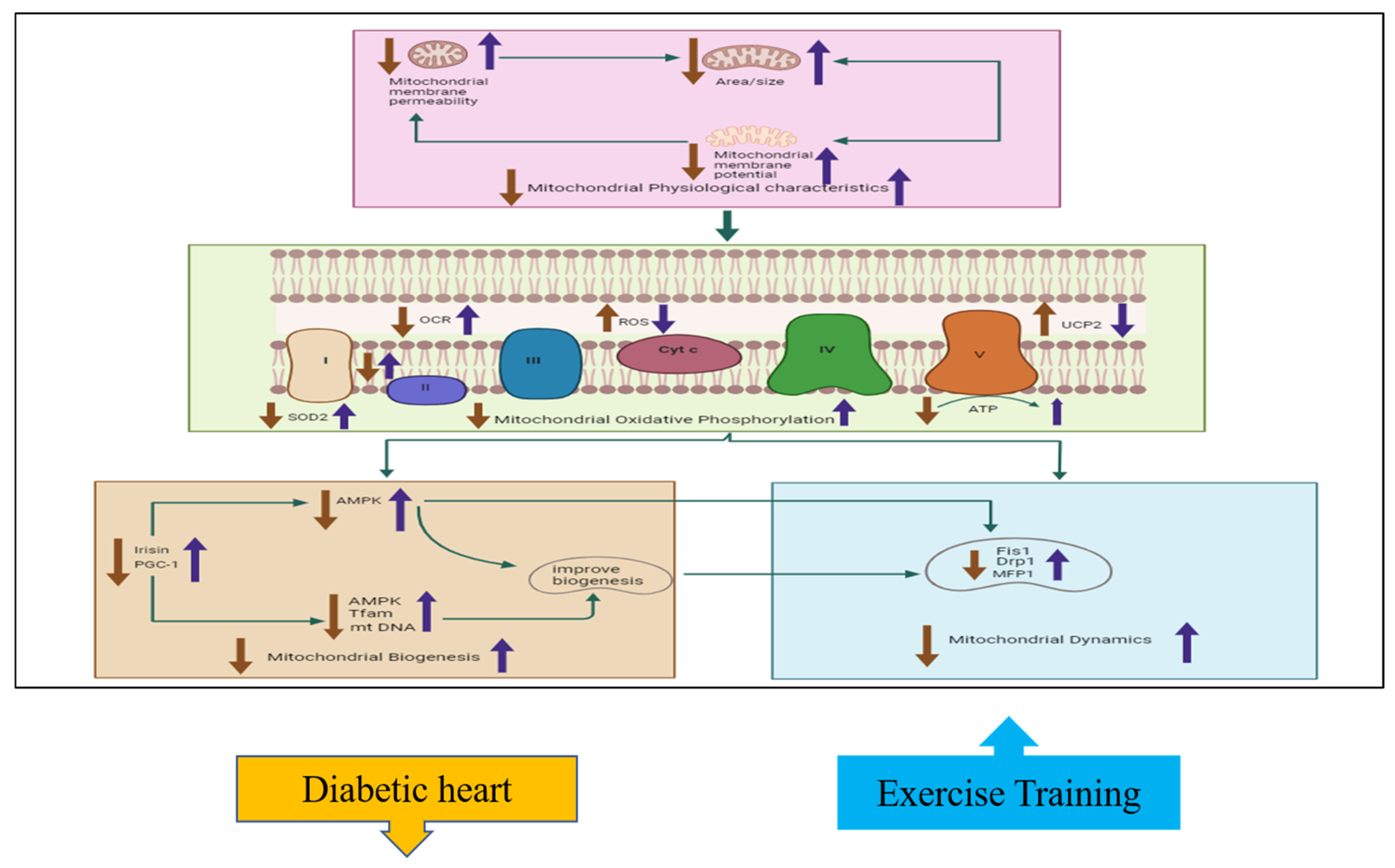
3.2. Effects of Exercise Training on Cardiac Physiological Properties in Diabetic Hearts
3.3. Effects of Exercise Training on Cardiac Oxidative Phosphorylation in Diabetic Hearts
3.4. Effects of Exercise Training on Cardiac Mitochondrial Biogenesis
3.5. Effects of Exercise Training on Cardiac Mitochondrial Dynamics in Diabetic Hearts
3.6. Effects of Exercise Training on Glycemic Parameters and Body Weight
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ritchie, R.H.; Abel, E.D. Basic mechanisms of diabetic heart disease. Circ. Res. 2020, 126, 1501–1525. [Google Scholar] [CrossRef] [PubMed]
- Hafstad, A.D.; Boardman, N.; Aasum, E. How exercise may amend metabolic disturbances in diabetic cardiomyopathy. Antioxid. Redox Signal. 2015, 22, 1587–1605. [Google Scholar] [CrossRef] [PubMed]
- Huynh, K.; Bernardo, B.C.; McMullen, J.R.; Ritchie, R.H. Diabetic cardiomyopathy: Mechanisms and new treatment strategies targeting antioxidant signaling pathways. Pharmacol. Ther. 2014, 142, 375–415. [Google Scholar] [CrossRef] [PubMed]
- Sivitz, W.I.; Yorek, M.A. Mitochondrial dysfunction in diabetes: From molecular mechanisms to functional significance and therapeutic opportunities. Antioxid. Redox Signal. 2010, 12, 537–577. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Geng, L.; Ying, L.; Shu, L.; Ye, K.; Yang, R.; Liu, Y.; Wang, Y.; Cai, Y.; Jiang, X.; et al. FGF21-Sirtuin 3 Axis Confers the Protective Effects of Exercise Against Diabetic Cardiomyopathy by Governing Mitochondrial Integrity. Circulation 2022, 146, 1537–1557. [Google Scholar] [CrossRef] [PubMed]
- Fontana, F.; Limonta, P. The multifaceted roles of mitochondria at the crossroads of cell life and death in cancer. Free Radic. Biol. Med. 2021, 176, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Dabkowski, E.R.; Baseler, W.A.; Williamson, C.L.; Razunguzwa, T.T.; Frisbee, J.C.; Hollander, J.M.; Pinti, M.V.; Fink, G.K.; Hathaway, Q.A.; Durr, A.J.; et al. Mitochondrial dysfunction in the type 2 diabetic heart is associated with alterations in spatially distinct mitochondrial proteomes. Am. J. Physio. Heart Circul. Physiol. 2010, 299, H529–H540. [Google Scholar] [CrossRef]
- Kuznetsov, A.V.; Javadov, S.; Margreiter, R.; Grimm, M.; Hagenbuchner, J.; Ausserlechner, M.J. The role of mitochondria in the mechanisms of cardiac ischemia-reperfusion injury. Antioxidants 2019, 8, 454. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Paulson, D.J.; Mathews, R.; Bowman, J.; Zhao, J.; Loganathan, R.; Bilgen, M.; Al-Hafez, B.; Zhero, S.V.; Alenezy, M.D.; Smirnova, I.V.; et al. Metabolic effects of treadmill exercise training on the diabetic heart. J. Appl. Physiol. 1992, 73, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Marcos, P.J.; Auwerx, J. Regulation of PGC-1α, a nodal regulator of mitochondrial biogenesis. Am. J. Clin. Nutr. 2011, 93, 884S–890S. [Google Scholar] [CrossRef] [PubMed]
- Marin, T.L.; Gongol, B.; Zhang, F.; Martin, M.; Johnson, D.A.; Xiao, H.; Wang, Y.; Subramaniam, S.; Chien, S.; Shyy, J.Y.-J. AMPK promotes mitochondrial biogenesis and function by phosphorylating the epigenetic factors DNMT1, RBBP7, and HAT1. Sci. Signal. 2017, 10, eaaf7478. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.C.; Geiger, P.C.; Han, D.-H.; Jones, T.E.; Holloszy, J.O. Calcium induces increases in peroxisome proliferator-activated receptor γ coactivator-1α and mitochondrial biogenesis by a pathway leading to p38 mitogen-activated protein kinase activation. J. Biol. Chem. 2007, 282, 18793–18799. [Google Scholar] [CrossRef]
- Din, S.; Konstandin, M.H.; Johnson, B.; Emathinger, J.; Völkers, M.; Toko, H.; Collins, B.; Ormachea, L.; Samse, K.; Kubli, D.A.; et al. Metabolic dysfunction consistent with premature aging results from deletion of Pim kinases. Circ. Res. 2014, 115, 376–387. [Google Scholar] [CrossRef]
- Zhang, F.; Lin, J.J.; Tian, H.N.; Wang, J. Effect of exercise on improving myocardial mitochondrial function in decreasing diabetic cardiomyopathy. Exp. Physiol. 2024, 109, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Losón, O.C.; Song, Z.; Chen, H.; Chan, D.C. Fis1, Mff, MiD49, and MiD51 mediate Drp1 recruitment in mitochondrial fission. Mol. Biol. Cell. 2013, 24, 659–667. [Google Scholar] [CrossRef]
- Yu, T.; Robotham, J.L.; Yoon, Y. Increased production of reactive oxygen species in hyperglycemic conditions requires dynamic change of mitochondrial morphology. Proc. Natl. Acad. Sci. USA 2006, 103, 2653–2658. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Robotham, J.L.; Yoon, Y. Mitochondrial fission mediates high glucose-induced cell death through elevated production of reactive oxygen species. Cardiovasc. Res. 2008, 79, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Makino, A.; Scott, B.; Dillmann, W. Mitochondrial fragmentation and superoxide anion production in coronary endothelial cells from a mouse model of type 1 diabetes. Diabetologia 2010, 53, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Trevellin, E.; Scorzeto, M.; Olivieri, M.; Granzotto, M.; Valerio, A.; Tedesco, L.; Fabris, R.; Serra, R.; Quarta, M.; Reggiani, C.; et al. Exercise training induces mitochondrial biogenesis and glucose uptake in subcutaneous adipose tissue through eNOS-dependent mechanisms. Diabetes 2014, 63, 2800–2811. [Google Scholar] [CrossRef] [PubMed]
- Ko, T.H.; Marquez, J.C.; Kim, H.K.; Jeong, S.H.; Lee, S.; Youm, J.B.; Song, I.S.; Seo, D.Y.; Kim, H.J.; Won, D.N.; et al. Resistance exercise improves cardiac function and mitochondrial efficiency in diabetic rat hearts. Pflügers Arch.-Eur. J. Physiol. 2018, 470, 263–275. [Google Scholar] [CrossRef]
- Botta, A.; Laher, I.; Beam, J.; DeCoffe, D.; Brown, K.; Halder, S.; Devlin, A.; Gibson, D.L.; Ghosh, S. Short term exercise induces PGC-1α, ameliorates inflammation and increases mitochondrial membrane proteins but fails to increase respiratory enzymes in aging diabetic hearts. PLoS ONE 2013, 8, e70248. [Google Scholar] [CrossRef]
- Wang, B.; Zhao, C.; Wang, Y.; Tian, X.; Lin, J.; Zhu, B.; Zhou, Y.; Zhang, X.; Li, N.; Sun, Y.; et al. Exercise ameliorating myocardial injury in type 2 diabetic rats by inhibiting excessive mitochondrial fission involving increased irisin expression and AMP-activated protein kinase phosphorylation. J. Diabetes 2023, 16, e13475. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Zhu, S.; Wu, J.; Zhang, M.; Xu, Y.; Xu, W.; Cui, J.; Yu, B.; Cao, W.; Liu, J. Exercise enhances cardiac function by improving mitochondrial dysfunction and maintaining energy homoeostasis in the development of diabetic cardiomyopathy. J. Mol. Med. 2020, 98, 245–261. [Google Scholar] [CrossRef]
- Wang, H.; Bei, Y.; Lu, Y.; Sun, W.; Liu, Q.; Wang, Y.; Cao, Y.; Chen, P.; Xiao, J.; Kong, X. Exercise Prevents Cardiac Injury and Improves Mitochondrial Biogenesis in Advanced Diabetic Cardiomyopathy with PGC-1α and Akt Activation. Cell. Physiol. Biochem. 2015, 35, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- Bækkerud, F.H.; Salerno, S.; Ceriotti, P.; Morland, C.; Storm-Mathisen, J.; Bergersen, L.H.; Høydal, M.A.; Catalucci, D.; Stølen, T.O. High Intensity Interval Training Ameliorates Mitochondrial Dysfunction in the Left Ventricle of Mice with Type 2 Diabetes. Cardiovasc. Toxicol. 2019, 19, 422–431. [Google Scholar] [CrossRef]
- Veeranki, S.; Givvimani, S.; Kundu, S.; Metreveli, N.; Pushpakumar, S.; Tyagi, S.C. Moderate intensity exercise prevents diabetic cardiomyopathy associated contractile dysfunction through restoration of mitochondrial function and connexin 43 levels in db/db mice. J. Mol. Cell. Cardiol. 2016, 92, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, N.; Lavoie, J.P.; Rousseau-Migneron, S.; Nadeau, A. Physical training reverses defect in mitochondrial energy production in heart of chronically diabetic rats. Diabetes 1993, 42, 682–687. [Google Scholar] [CrossRef]
- Lumini-Oliveira, J.; Magalhães, J.; Pereira, C.V.; Moreira, A.C.; Oliveira, P.J.; Ascensão, A. Endurance training reverts heart mitochondrial dysfunction, permeability transition and apoptotic signaling in long-term severe hyperglycemia. Mitochondrion 2011, 11, 54–63. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Shkurat, T.P.; Melnichenko, A.A.; Grechko, A.V.; Orekhov, A.N. The role of mitochondrial dysfunction in cardiovascular disease: A brief review. Ann. Med. 2018, 50, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.M.; Tate, M.; Prakoso, D.; Deo, M.; Willis, A.M.; Nash, D.M.; Donner, D.G.; Crawford, S.; Kiriazis, H.; Granata, C.; et al. Characterisation of the Myocardial Mitochondria Structural and Functional Phenotype in a Murine Model of Diabetic Cardiomyopathy. Front. Physiol. 2021, 12, 672252. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.L.; Dabkowski, E.R.; Baseler, W.A.; Croston, T.L.; Alway, S.E.; Hollander, J.M. Enhanced apoptotic propensity in diabetic cardiac mitochondria: Influence of subcellular spatial location. Am. J. Physiol. Heart C 2010, 298, H633–H642. [Google Scholar] [CrossRef]
- Sloan, R.C.; Moukdar, F.; Frasier, C.R.; Patel, H.D.; Bostian, P.A.; Lust, R.M.; Brown, D.A. Mitochondrial permeability transition in the diabetic heart: Contributions of thiol redox state and mitochondrial calcium to augmented reperfusion injury. J. Mol. Cell. Cardiol. 2012, 52, 1009–1018. [Google Scholar] [CrossRef]
- Jubaidi, F.F.; Zainalabidin, S.; Mariappan, V.; Budin, S.B. Mitochondrial Dysfunction in Diabetic Cardiomyopathy: The Possible Therapeutic Roles of Phenolic Acids. Int. J. Mol. Sci. 2020, 21, 6043. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.X. Mitochondrial dysfunction, oxidative stress and diabetic cardiovascular disorders. Cardiovasc. Hematol. Disord. Drug Targets 2012, 12, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Gollmer, J.; Zirlik, A.; Bugger, H. Mitochondrial mechanisms in diabetic cardiomyopathy. Diabetes Metabol. J. 2020, 44, 33–53. [Google Scholar] [CrossRef] [PubMed]
- Bugger, H.; Abel, E.D. Mitochondria in the diabetic heart. Cardiovasc. Res. 2010, 88, 229–240. [Google Scholar] [CrossRef]
- Dludla, P.V.; Nkambule, B.B.; Tiano, L.; Louw, J.; Jastroch, M.; Mazibuko-Mbeje, S.E. Uncoupling proteins as a therapeutic target to protect the diabetic heart. Pharmacol. Res. 2018, 137, 11–24. [Google Scholar] [CrossRef] [PubMed]
- How, O.-J.; Aasum, E.; Severson, D.L.; Chan, W.A.; Essop, M.F.; Larsen, T.S. Increased myocardial oxygen consumption reduces cardiac efficiency in diabetic mice. Diabetes 2006, 55, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Viloria, M.A.D.; Li, Q.; Lu, W.; Nhu, N.T.; Liu, Y.; Cui, Z.-Y.; Cheng, Y.-J.; Lee, S.-D. Effect of exercise training on cardiac mitochondrial respiration, biogenesis, dynamics, and mitophagy in ischemic heart disease. Front. Cardiovasc. Med. 2022, 9, 949744. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Guo, Y.; Xia, Y.; Li, C.; Xu, X.; Qi, T.; Zhang, F.; Fan, M.; Hu, G.; Zhao, H.; et al. FNDC5/Irisin attenuates diabetic cardiomyopathy in a type 2 diabetes mouse model by activation of integrin αV/β5-AKT signaling and reduction of oxidative/nitrosative stress. J. Mol. Cell Cardiol. 2021, 160, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Huang, X.; Xu, M.; Yang, L.; Hua, F. MiR-144 protects the heart from hyperglycemia-induced injury by regulating mitochondrial biogenesis and cardiomyocyte apoptosis. FASEB J. 2020, 34, 2173–2197. [Google Scholar] [CrossRef]
- Tao, L.-C.; Wang, T.-T.; Zheng, L.; Hua, F.; Li, J.-J. The role of mitochondrial biogenesis dysfunction in diabetic cardiomyopathy. Biomol. Ther. 2022, 30, 399. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; He, C.; Zou, M.-H. AMP-activated protein kinase modulates cardiac autophagy in diabetic cardiomyopathy. Autophagy 2011, 7, 1254–1255. [Google Scholar] [CrossRef]
- Paramesha, B.; Anwar, M.S.; Meghwani, H.; Maulik, S.K.; Arava, S.K.; Banerjee, S.K. Sirt1 and Sirt3 Activation Improved Cardiac Function of Diabetic Rats via Modulation of Mitochondrial Function. Antioxidants 2021, 10, 338. [Google Scholar] [CrossRef]
- Chen, Y.H.; Ta, A.P.; Chen, Y.; Lee, H.C.; Fan, W.; Chen, P.L.; Jordan, M.C.; Roos, K.P.; MacGregor, G.R.; Yang, Q.; et al. Dual roles of myocardial mitochondrial AKT on diabetic cardiomyopathy and whole body metabolism. Cardiovasc. Diabetol. 2023, 22, 294. [Google Scholar] [CrossRef]
- Karamanlidis, G.; Bautista-Hernandez, V.; Fynn-Thompson, F.; del Nido, P.; Tian, R. Impaired mitochondrial biogenesis precedes heart failure in right ventricular hypertrophy in congenital heart disease. Circ. Heart Fail. 2011, 4, 707–713. [Google Scholar] [CrossRef]
- García-Peña, L.M.; Abel, E.D.; Pereira, R.O. Mitochondrial Dynamics, Diabetes, and Cardiovascular Disease. Diabetes 2024, 73, 151–161. [Google Scholar] [CrossRef]
- Hu, L.; Ding, M.; Tang, D.; Gao, E.; Li, C.; Wang, K.; Qi, B.; Qiu, J.; Zhao, H.; Chang, P.; et al. Targeting mitochondrial dynamics by regulating Mfn2 for therapeutic intervention in diabetic cardiomyopathy. Theranostics 2019, 9, 3687. [Google Scholar] [CrossRef]
- Zhou, Y.; Suo, W.; Zhang, X.; Liang, J.; Zhao, W.; Wang, Y.; Li, H.; Ni, Q. Targeting mitochondrial quality control for diabetic cardiomyopathy: Therapeutic potential of hypoglycemic drugs. Biomed. Pharmacother. 2023, 168, 115669. [Google Scholar] [CrossRef]
- Trimarchi, G.; Pizzino, F.; Paradossi, U.; Gueli, I.A.; Palazzini, M.; Gentile, P.; Di Spigno, F.; Ammirati, E.; Garascia, A.; Tedeschi, A.; et al. Charting the Unseen: How Non-Invasive Imaging Could Redefine Cardiovascular Prevention. J. Cardiovasc. Dev. Dis. 2024, 11, 245. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| References | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Wang et al. [23] | √ | √ | √ | √ | √ | √ | √ | 7 | |||
| 2. Wang et al. [24] | √ | √ | √ | √ | √ | 5 | |||||
| 3. Wang et al. [25] | √ | √ | √ | √ | √ | √ | 6 | ||||
| 4. Bækkerud et al. [26] | √ | √ | √ | √ | 4 | ||||||
| 5. Veeranki et al. [27] | √ | √ | √ | √ | 4 | ||||||
| 6. Mokhtar et al. [28] | √ | √ | √ | √ | √ | 5 | |||||
| 7. Jin et al. [5] | √ | √ | √ | √ | √ | √ | √ | 7 | |||
| 8. Ko et al. [21] | √ | √ | √ | √ | √ | 5 | |||||
| 9. Botta et al. [22] | √ | √ | √ | √ | √ | √ | 6 | ||||
| 10. Oliveira et al. [29] | √ | √ | √ | √ | √ | √ | 6 |
| References | Sample and Diabetes Type | Exercise Type | Exercise Parameters | Glycemic Parameters and Body Weight (BW) | Mitochondrial Physiological Characteristics | Mitochondrial Oxidative Phosphorylation | Mitochondrial Biogenesis | Mitochondrial Dynamics |
|---|---|---|---|---|---|---|---|---|
| 1. Wang et al. [23] | S: Male Wistar rats A: 48 weeks DT: Type-II | Treadmill training | Sp: 10–18 m/min D: 10–45 min/day F: 5 days/week P: 8 weeks | DH: BW↑ ↑,hyperglycemia↑ ET: BW↓, hyperglycemia↓ | DH: NADH↓, SDH↓, CCR↓, CCO↓ ET: NADH↑, SDH↑, CCR↑,CCO↑ | DH: AMPK↓ Irisin↓ ET: AMPK↑, Irisin↑ | DH: Drp1↑, Fis1↑,MFF↑ ET: Drp1↓, Fis1↓,MFF↓ | |
| 2. Wang et al. [24] | S: Male C57BL/6 mice A: 6 weeks DT: Type-II | Treadmill training | Sp: 10 m/min D: 1 hr/day F: 4 days/week P: 16 weeks | DH: BW↑, plasma glucose↑ ET: BW↓, ↓plasma glucose | DH: Membrane potential↓ ET: Membrane potential↑ | DH: 4HNE↑,Nox4↑,p47phox↑, p67phox↑,ROS↑ ET: 4HNE↓,Nox4↓, p47phox↓, p67phox↓,ROS↓ DH: UCP2↑, ATP↓, RCR↓, OCR↑ ET: UCP2↓, ATP↑, RCR↑, OCR↓ DH: ETC enzymes like NUDE↓, CCR↓, COX↓, SDH↓, SOD2↓ ET:ETC enzymes like NUDE↑, CCR↑, COX↑,SDH↑,SOD2↑ | DH: PGC-1α↓,AMPK↓ ET:PGC-1α↑,AMPK↑ | |
| 3. Wang et al. [25] | S: Male C57BL/6 mice A: 6 weeks DT: Type-II | Treadmill training | Sp: 10 m/min D: 1 hr/day F: days/week P: 15 weeks | DH: BW↑ ET: BW↓ | DH: 16sRNA↓,ND1↓,ND6↓, CYBT↓,mtDNA↓,SSDPB1↓,Top1mt↓ Twinkle↓ ET: 16sRNA↑,ND1↑,ND6↑, CYBT↑,mtDNA↑,SSDPB1↑, Top1mt ↑, Twinkle↑ DH: PGC1α↓,AKT↓,TFAM↓ ET: PGC1α ↑,AKT↑,TFAM↑ DH: mRNA of NRF1↓ ET: mRNA of NRF1 ↑ | |||
| 4. Bækkerud et al. [26] | S: Male (BKS.cgm+/Lepdb/Bom Tac) db/db mice A: 8 weeks DT: Type-II | Treadmill training | Sp: progressive 0.03 m/s D: 40 min/day F: 5 days/week P: 8 weeks | DH:BW↑,serum glucose↑ ET: BW↓,No significant change In glucose level | DH: Size↓,quantity↑ and density (no change) ET: Slight ↑ in size and quantity and NC in density | DH: ETC complexes (CI, I+II, III, IV) activity↓ ET: ETC complexes (CI+II,CII and CIV) Activity↑ DH: IDH↑,OGDH↑,SDH↑, ET: IDH↓,OGDH↓,SDH↓ DH: MCU↓ ET: MCU ↑ | DH: MFP1↓ ET: MFP1↑ | |
| 5. Veeranki et al. [27] | S: Male C57BL/6 mice A: 2 months DT: Type-II | Treadmill training | Sp: 10–11 m/min F: 5 day/week P: 5 weeks | DH: BW↑, glucose↑ ET: BW↓, glucose↓ | DH: Abnormal membrane permeability ET: Restores membrane permeability | DH: OCR↓, ATP↓ ET: OCR↑, ATP↓ DH: Cytochrome content↓, leakage↑ ET: Cytochrome content↑, leakage↓ | DH: Mitochondrial biogenesis imbalance ET: Mitochondrial biogenesis balanced | DH: Drp1↑,Mfn2(NC) ET: Drp1,Mfn2 (NC) |
| 6. Mokhtar et al. [28] | S: Male Wistar rats A: N/A DT: Type-II | Rodent treadmill training | Sp: Progressive exercise D: 10–60 min/day F: 5 days/week P: 10 week | DH: Same BW, glucose↑ ET: No changes in BW and glucose levels | DH: State-3 respiration↓, ATP ↓ ET: State-3 respiration↑, ATP↑ | |||
| 7. Jin et al. [5] | S: Male C57BL/6J mice A: 6 weeks DT: Type-II | Treadmill training | P: 6 weeks | DH: BW↑, fasting blood glucose↑ ET: BW↓, fasting blood glucose↓ | DH: Area↓, diameter↓, perimeter↓, cristae area↓, mitochondrial area↓ ET: Area↑, diameter↑, perimeter↑, cristae area↑, mitochondrial area↑ | DH: oxidative stress↑, ATP↓, FAO↓, ROS↑ ET: oxidative stress↓, ATP↑, FAO↑, ROS↓ DH: MRC complex (I -V) activity↓ ET: MRC complex(I-V) activity ↑ DH: Mitochondrial acetylation of SOD2↑, LCAD ↑ ET: Mitochondrial acetylation of SOD2↓, LCAD↓ | DH: Sirtuin-3↓,AMPK ↓ ET:Sirtuin-3↑,AMPK↑ | |
| 8. Ko et al. [21] | S: Male Otuska Long– Evans Tokushima Fatty rats A: 28 weeks DT: Type-II | Climbing ladder | Sp: 20 Reps F: 5 days/week P: 12 weeks | DH: BW↑, glucose↑ ET: BW↓, glucose↓ | DH: Membrane potential↓ ET: Membrane potential↑ | DH: GLUT-4↓, PDHE1α↓ ET: GLUT-4↑, PDHE1α↑ DH: State-4 respiration↑, ROS↑ ET: State-4 respiration↓, ROS↓ DH: RCR ↓, ATP↓,SOD2↓ ET: RCR↑, ATP↑, SOD2↑ DH:UCP2↑ UCP3↑, ET: UCP2↓, UCP3↓ | DH: PGC1α↓,TFAM↓ ET: PGC1α↑,TFAM↑ | |
| 9. Botta et al. [22] | S: Male C57BLKS/J mice with A: 6 weeks DT: Type-II | Motorized exercise wheel system | Sp: 5.2 m/min F: 5 days/week P: 3 weeks | DH: Blood glucose and BW changes ET: No changes observed after exercise | DH: TOM-70↓, VDAC-1 ↓ ET:TOM-70↑, VDAC-1↑ DH: ETC proteins lost complexes(I-V) ET: Complex I and activity II↓, IV ↑but III and V remain unchanged DH: SOD2↑ ET:SOD2↓ | DH: mt DNA↓, PGC-1α↓ ET: mt DNA↑, PGC-1α↑ DH: TFAM↓, NRF 1 and 2↓ ET: No changes seen in the above transcription factors | DH: Signs of fission and fusion noted ET: No effect of exercise on fission and fusion | |
| 10. Oliveira et al. [29] | S: Male Wistar rats A: 6–8 weeks DT: Type-II | Motor-driven treadmill | Sp: 25 m/min F: 5 days/week P: 14 weeks | DH: Body weight↓, blood glucose↓ ET: Body weight↑, blood glucose↑ | DH: Mitochondrial permeability ↑ ET: Mitochondrial permeability ↓ DH: Swelling↑ ET: Swelling↓ | DH: State 3 and state 4 respiration↓ ET: State 3 and state 4 respiration↑ DH: Complex I and II activity ↓ ET: Complex I and II activity ↑ DH: RCR↓ ET: RCR↑ | DH: TFAM↑ ET: TFAM↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, I.A.; Ishaq, S.; Lee, S.-D.; Wu, B.-T. Effects of Exercise Training on Cardiac Mitochondrial Functions in Diabetic Heart: A Systematic Review. Int. J. Mol. Sci. 2025, 26, 8. https://doi.org/10.3390/ijms26010008
Shah IA, Ishaq S, Lee S-D, Wu B-T. Effects of Exercise Training on Cardiac Mitochondrial Functions in Diabetic Heart: A Systematic Review. International Journal of Molecular Sciences. 2025; 26(1):8. https://doi.org/10.3390/ijms26010008
Chicago/Turabian StyleShah, Iqbal Ali, Shahid Ishaq, Shin-Da Lee, and Bor-Tsang Wu. 2025. "Effects of Exercise Training on Cardiac Mitochondrial Functions in Diabetic Heart: A Systematic Review" International Journal of Molecular Sciences 26, no. 1: 8. https://doi.org/10.3390/ijms26010008
APA StyleShah, I. A., Ishaq, S., Lee, S.-D., & Wu, B.-T. (2025). Effects of Exercise Training on Cardiac Mitochondrial Functions in Diabetic Heart: A Systematic Review. International Journal of Molecular Sciences, 26(1), 8. https://doi.org/10.3390/ijms26010008