
Protein-Based Mechanism of Wheat Growth Under Salt Stress in Seeds Irradiated with Millimeter Waves

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Root Growth of Wheat Under Salt Stress Was Improved by MMW Irradiation
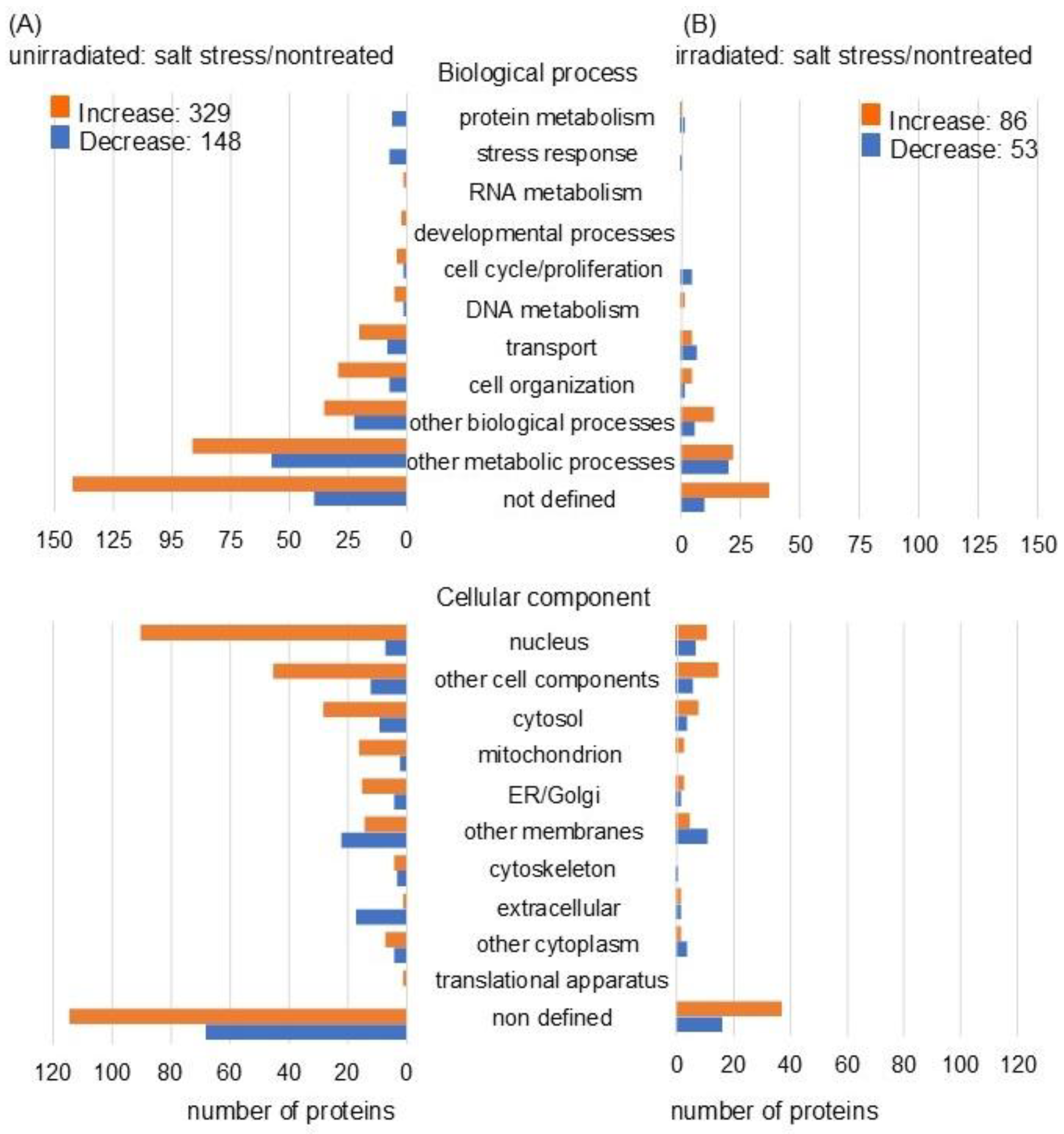
2.2. Salt Stress-Affected Wheat Proteins, Which Were Restored at the Protein Level by MMW Irradiation
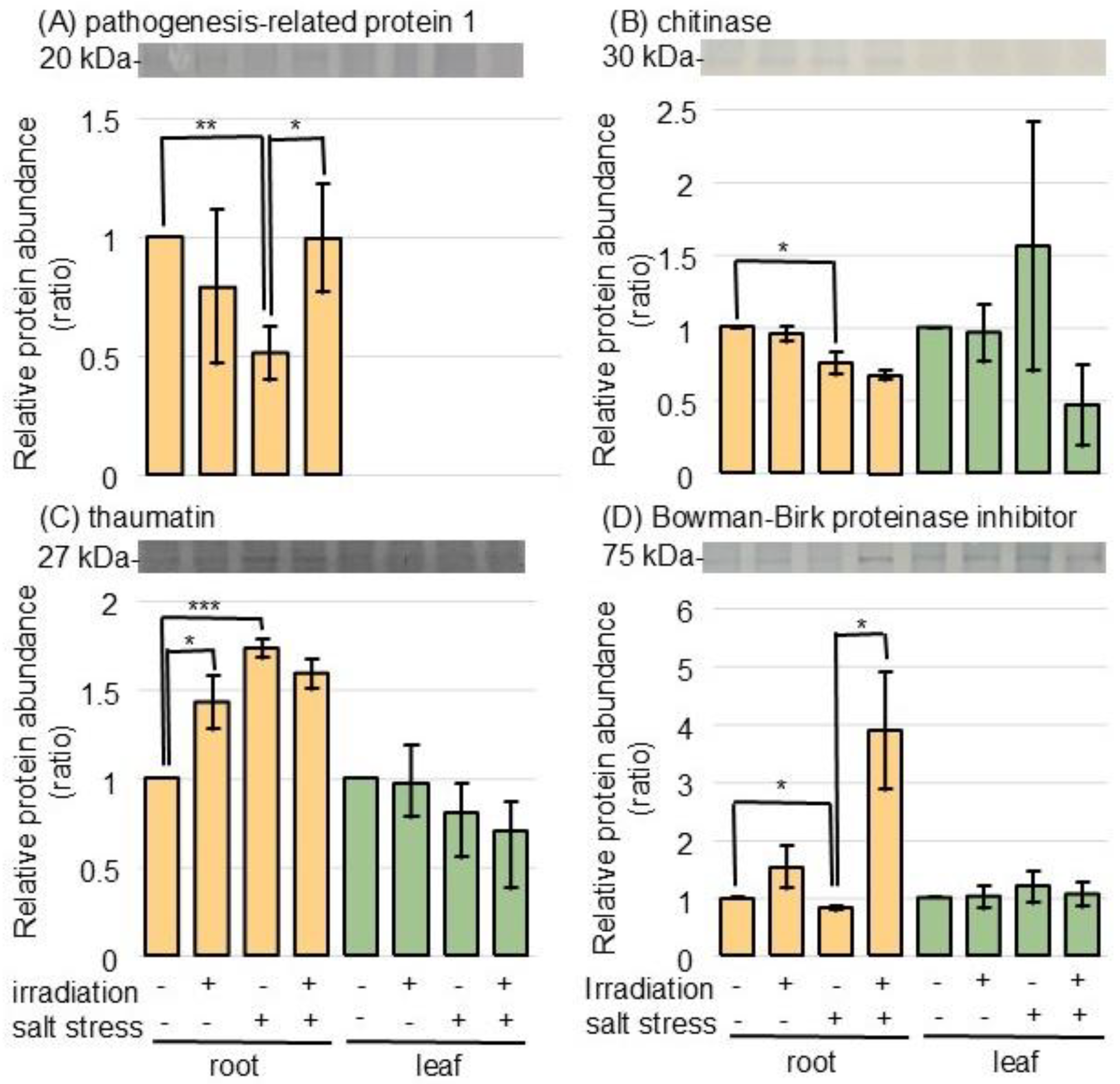
2.3. Protein Accumulation Was Altered Under Salt Stress in Wheat Irradiated with MMW, as Determined Using Immunoblot Analysis
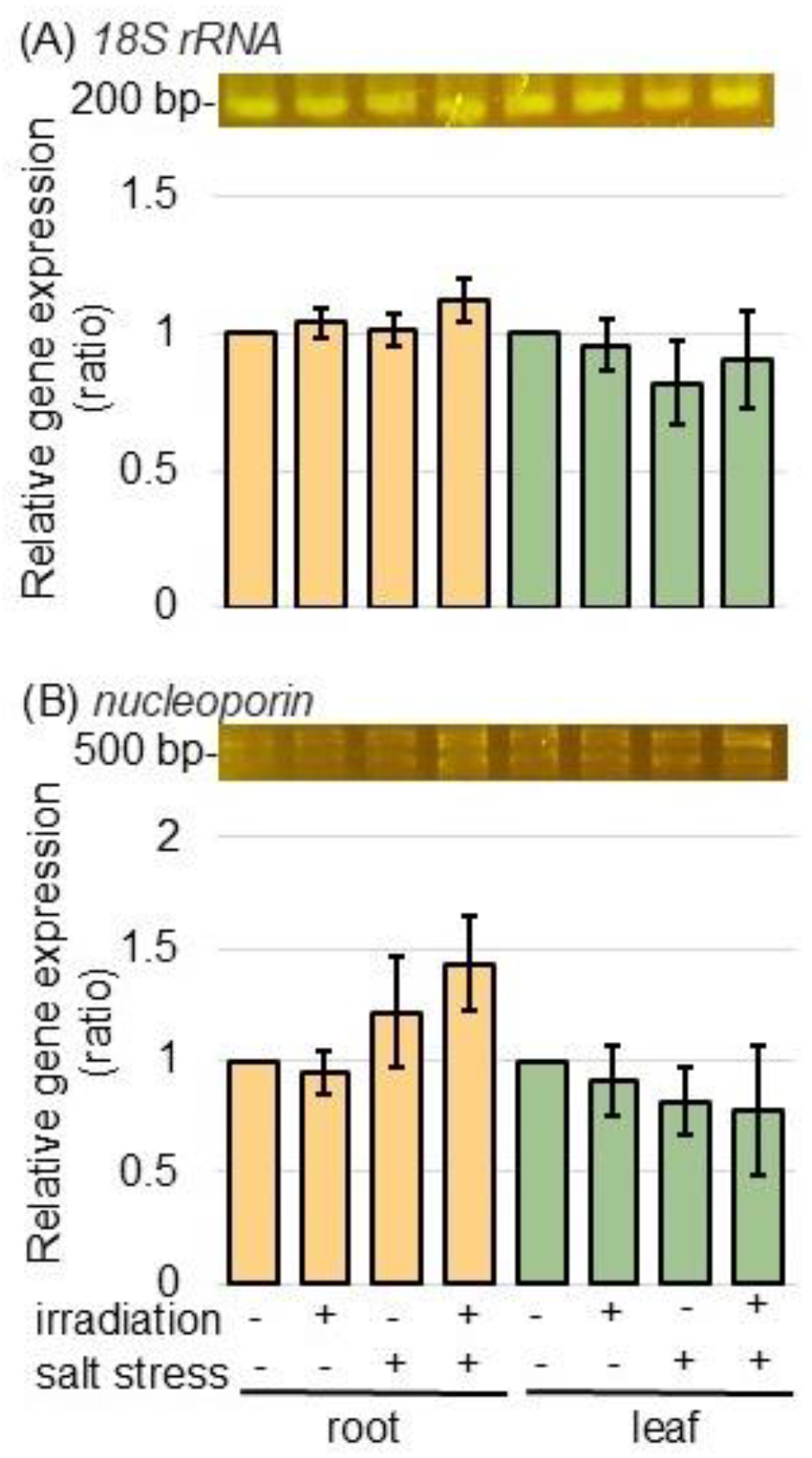
2.4. Gene Expression Was Analyzed Under Salt Stress in Wheat Irradiated with MMW Using PCR Technique
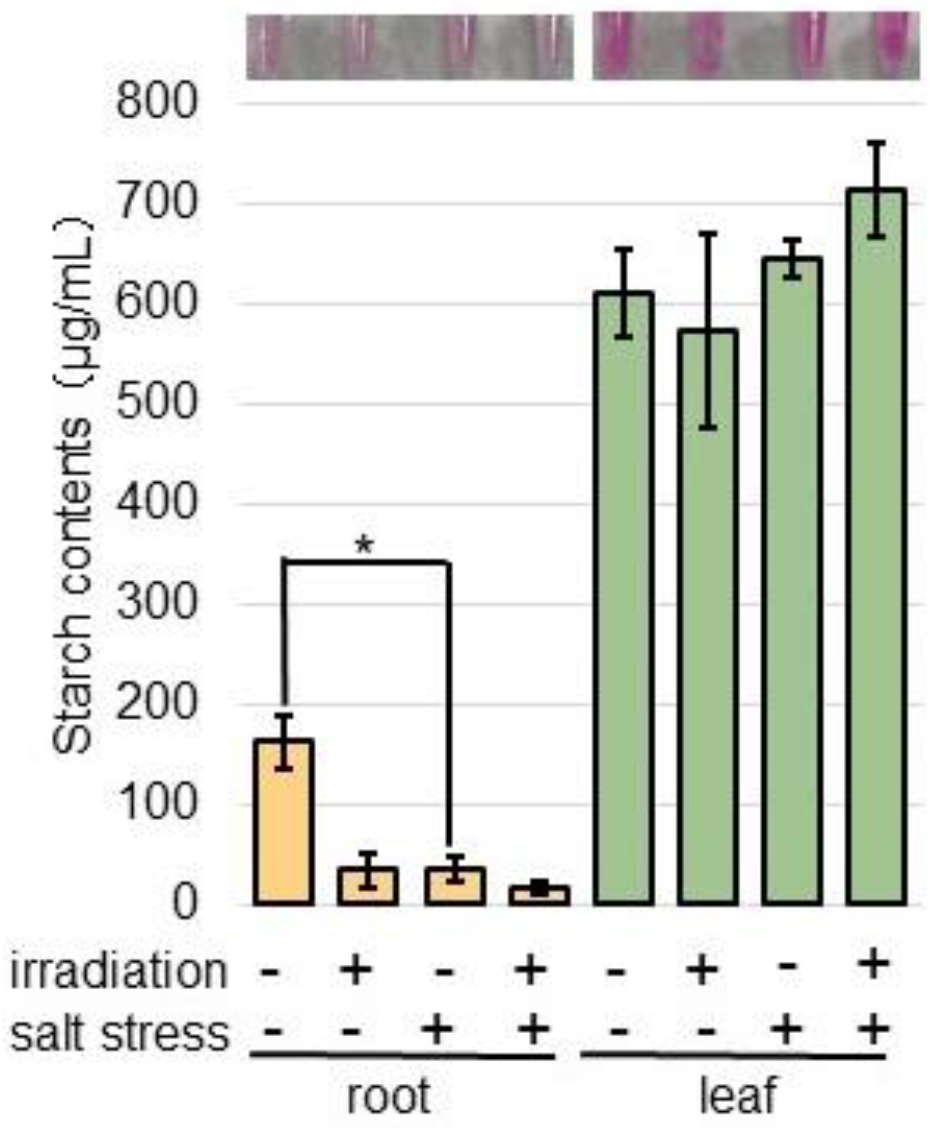
2.5. Starch Content Decreased Under Salt Stress in Wheat Irradiated with MMW
3. Discussion
3.1. MMW Irradiation Positively Affects Wheat Growth Under Salt Stress
3.2. MMW Irradiation Regulates the ROS Scavenging Process in Wheat Under Salt Stress
3.3. MMW Irradiation Regulates Pathogen-Related Proteins in Wheat Under Salt Stress
4. Materials and Methods
4.1. Plant Materials and Salt Treatment
4.2. Protein Extraction and Concentration Measurement
4.3. Protein Enrichment, Reduction, Alkylation, and Digestion
4.4. Protein Identification Using LC-MS/MS
4.5. Analysis of MS Data
4.6. Differential Analysis of Proteins Using MS Data
4.7. Immunoblot Analysis
4.8. RNA Extraction, cDNA Synthesis, and PCR Analysis
4.9. Starch-Content Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Crops and Livestock Products. Online Database on Crop Production, Yield and Harvested Area. 2022, pp. 1–5. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 1 October 2024).
- Ortiz, R.; Sayre, K.D.; Govaerts, B.; Gupta, R.; Subbarao, G.V.; Ban, T.; Hodson, D.; Dixon, J.M.; Iván Ortiz-Monasterio, J.; Reynolds, M. Climate change: Can wheat beat the heat? Agric. Ecosys. Environ. 2008, 126, 46–58. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.H.; Foster, K.J.; et al. Energy costs of salt tolerance in crop plants. New Phytol. 2020, 225, 1072–1090. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Bohra, A.; Jha, R.; Parida, S.K. Salinity stress response and ‘omics’ approaches for improving salinity stress tolerance in major grain legumes. Plant Cell Rep. 2019, 38, 255–277. [Google Scholar] [CrossRef]
- Kumar, P.; Choudhary, M.; Halder, T.; Prakash, N.R.; Singh, V.T.; Sheoran, S.; Longmei, N.; Rakshit, S.; Siddique, K.H.M. Salinity stress tolerance and omics approaches: Revisiting the progress and achievements in major cereal crops. Heredity 2022, 128, 497–518. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 2020, 10, 928. [Google Scholar] [CrossRef]
- Feki, K.; Quintero, F.J.; Pardo, J.M.; Masmoudi, K. Regulation of durum wheat Na+/H+ exchanger TdSOS1 by phosphorylation. Plant Mol. Biol. 2011, 76, 545–556. [Google Scholar] [CrossRef]
- Sharma, H.; Taneja, M.; Upadhyay, S.K. Identification, characterization and expression profiling of cation-proton antiporter superfamily in Triticum aestivum L. and functional analysis of TaNHX4-B. Genomics 2020, 112, 356–370. [Google Scholar] [CrossRef]
- Amirbakhtiar, N.; Ismaili, A.; Ghaffari, M.R.; Nazarian Firouzabadi, F.; Shobbar, Z.S. Transcriptome response of roots to salt stress in a salinity-tolerant bread wheat cultivar. PLoS ONE 2019, 14, e0213305. [Google Scholar] [CrossRef]
- Shakirova, F.M.; Sakhabutdinova, A.R.; Bezrukova, M.V.; Atkhutdinova, R.A.; Fatkhutdinova, D.R. Changes in the hormonal status of wheat seedlings induced by salicylic acid and salinity. Plant Sci. 2003, 164, 317–322. [Google Scholar] [CrossRef]
- Sheen, D.M.; McMakin, D.L.; Hall, T.E. Detection of explosives by millimeter-wave imaging. In Counterterrorist Detection Techniques Explosives; Elsevier Science BV: Amsterdam, The Netherlands, 2007; pp. 237–277. [Google Scholar]
- Mignone, C.; Barnes, R. More than meets the eye: The electromagnetic spectrum. Sci. School 2011, 20, 51–59. [Google Scholar]
- Abbey, L.; Udenigwe, C.; Mohan, A.; Anom, E. Microwave irradiation effects on vermicasts potency, and plant growth and antioxidant activity in seedlings of Chinese cabbage (Brassica rapa subsp. pekinensis). J. Radiat. Res. Appl. Sci. 2017, 10, 110–116. [Google Scholar] [CrossRef]
- Seo, D.H.; Kim, M.S.; Choi, H.W.; Sung, J.M.; Park, J.D.; Kum, J.S. Effects of millimeter wave treatment on the germination rate and antioxidant potentials and gamma-aminobutyric acid of the germinated brown rice. Food Sci. Biotechnol. 2016, 25, 111–114. [Google Scholar] [CrossRef]
- Betskii, O.V.; Lebedeva, N.N.; Tambiev, A.H.; Kirikova, N.N.; Slavin, V.E. Millimeter waves in the newest agricultural biotechnologies. J. Sci. Eng. 2007, 23, 236–252. [Google Scholar]
- Poghosyan, G.H.; Mukhaelyan, Z.H. The influence of low-intensity EMI treatment on seed germination and early growth of theat. Proc. Yerevan State Univ. Chem. Biol. 2018, 52, 110–115. [Google Scholar]
- Mukhaelyan, Z.H.; Shahinyan, M.A.; Poghosyan, G.H.; Vardevanyan, P.O. Wheat seedlings growth and antioxidant system activity changes as responses to extremely high frequency electromagnetic irradiation. Am. J. Plant Bio. 2016, 2, 1–10. [Google Scholar]
- Zhong, Z.; Furuya, T.; Ueno, K.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Tani, M.; Tian, J.; Komatsu, S. Proteomic analysis of irradiation with millimeter waves on soybean growth under flooding conditions. Int. J. Mol. Sci. 2020, 21, 486. [Google Scholar] [CrossRef]
- Komatsu, S.; Maruyama, J.; Furuya, T.; Yin, X.; Yamaguchi, H.; Hitachi, K.; Miyashita, N.; Tsuchida, K.; Tani, M. Proteomic and biological analyses reveal the effect on growth under flooding stress of chickpea irradiated with millimeter waves. J. Proteome Res. 2021, 20, 4718–4727. [Google Scholar] [CrossRef]
- Komatsu, S.; Tsutsui, Y.; Furuya, T.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Tani, M. Proteomic and biochemical approaches elucidate the role of millimeter-wave irradiation in wheat growth under flooding stress. Int. J. Mol. Sci. 2022, 23, 10360. [Google Scholar] [CrossRef]
- Jilani, A.; Ansari, M.O.; Rehman, G.U.; Shakoor, M.B.; Hussain, S.Z.; Othman, M.H.D.; Ahmad, S.R.; Dustgeer, M.R.; Alshahrie, A. Phenol removal and hydrogen production from water: Silver nanoparticles decorated on polyaniline wrapped zinc oxide nanorods. J. Ind. Eng. Chem. 2022, 109, 347–358. [Google Scholar] [CrossRef]
- Komatsu, S.; Hamada, K.; Furuya, T.; Nishiuchi, T.; Tani, M. Membrane proteomics to understand enhancement effects of millimeter-wave irradiation on wheat root under flooding stress. Int. J. Mol. Sci. 2023, 24, 9014. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Nishiuchi, T.; Furuya, T.; Tani, M. Millimeter-wave irradiation regulates mRNA-expression and the ubiquitin-proteasome system in wheat exposed to flooding stress. J. Proteomics 2024, 294, 105073. [Google Scholar] [CrossRef] [PubMed]
- Talaat, N.B.; Todorova, D. Antioxidant machinery and glyoxalase system regulation confers salt stress tolerance to wheat (Triticum aestivum L.) plants treated with melatonin and salicylic Acid. J. Soil Sci. Plant Nutr. 2022, 22, 3527–3540. [Google Scholar] [CrossRef]
- Ben Saad, R.; Ben Romdhane, W.; Wiszniewska, A.; Baazaoui, N.; Taieb Bouteraa, M.; Chouaibi, Y.; Alfaifi, M.Y.; Kačániová, M.; Čmiková, N.; Ben Hsouna, A.; et al. Rosmarinus officinalis L. essential oil enhances salt stress tolerance of durum wheat seedlings through ROS detoxification and stimulation of antioxidant defense. Protoplasma, 2024; ahead of print. [Google Scholar] [CrossRef]
- Hameed, A.; Maqsood, W.; Hameed, A.; Qayyum, M.A.; Ahmed, T.; Farooq, T. Chitosan nanoparticles encapsulating curcumin counteract salt-mediated ionic toxicity in wheat seedlings: An ecofriendly and sustainable approach. Environ. Sci. Pollut. Res. Int. 2024, 31, 8917–8929. [Google Scholar] [CrossRef]
- Wang, M.; Xia, G.M. The landscape of molecular mechanisms for salt tolerance in wheat. Crop J. 2018, 6, 42–47. [Google Scholar] [CrossRef]
- Liu, S.; Liu, S.; Wang, M.; Wei, T.; Meng, C.; Wang, M.; Xia, G. A wheat SIMILAR TO RCD-ONE gene enhances seedling growth and abiotic stress resistance by modulating redox homeostasis and maintaining genomic integrity. Plant Cell 2014, 26, 164–180. [Google Scholar] [CrossRef]
- Hou, L.; Liu, Z.; Zhang, D.; Liu, S.; Chen, Z.; Wu, Q.; Shang, Z.; Wang, J.; Wang, J. BR regulates wheat root salt tolerance by maintaining ROS homeostasis. Planta 2024, 260, 5. [Google Scholar] [CrossRef]
- Yang, R.; Yang, Z.; Xing, M.; Jing, Y.; Zhang, Y.; Zhang, K.; Zhou, Y.; Zhao, H.; Qiao, W.; Sun, J. TaBZR1 enhances wheat salt tolerance via promoting ABA biosynthesis and ROS scavenging. J. Genet. Genom. 2023, 50, 861–871. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, X.; Xiao, Z.; Yin, X.; Xing, T.; Xia, G. A wheat superoxide dismutase gene TaSOD2 enhances salt resistance through modulating redox homeostasis by promoting NADPH oxidase activity. Plant Mol. Biol. 2016, 91, 115–130. [Google Scholar] [CrossRef]
- Wang, J.Y.; Li, Q.; Mao, X.G.; Li, A.; Jing, R.L. Wheat transcription factor TaAREB3 participates in drought and freezing tolerances in Arabidopsis. Int. J. Biol. Sci. 2016, 12, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.J.; Ding, L.; Zhu, J.K. SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell 1996, 8, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.Y.; Du, Y.T.; Fu, J.D.; Yu, T.F.; Wang, C.T.; Chen, M.; Chen, J.; Ma, Y.Z.; Xu, Z.S. Wheat CBL-interacting protein kinase 23 positively regulates drought stress and ABA responses. BMC Plant Biol. 2018, 18, 93. [Google Scholar] [CrossRef] [PubMed]
- Delaney, T.P.; Uknes, S.; Vernooij, B.; Friedrich, L.; Weymann, K.; Negrotto, D.; Gaffney, T.; Gut-Rella, M.; Kessmann, H.; Ward, E.; et al. A central role of salicylic acid in plant disease resistance. Science 1994, 266, 1247–1250. [Google Scholar] [CrossRef]
- Gruner, R.; Strompen, G.; Pfitzner, A.J.P.; Pfitzner, U.M. Salicylic acid and the hypersensitive response initiate distinct signal transduction pathways in tobacco that converge on the as-1-like element of the PR-1a promoter. Eur. J. Biochem. 2003, 270, 4876–4886. [Google Scholar] [CrossRef]
- Kiba, A.; Nishihara, M.; Nakatsuka, T.; Yamamura, S. Pathogenesis-related protein 1 homologue is an antifungal protein in Wasabia japonica leaves and confers resistance to Botrytis cinerea in transgenic tobacco. Plant Biotechnol.-Nar. 2007, 24, 247–253. [Google Scholar] [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef]
- Seo, P.J.; Lee, A.K.; Xiang, F.; Park, C.M. Molecular and functional profiling of Arabidopsis pathogenesis-related genes: Insights into their roles in salt response of seed germination. Plant Cell Physiol. 2008, 49, 334–344. [Google Scholar] [CrossRef]
- Liu, W.X.; Zhang, F.C.; Zhang, W.Z.; Song, L.F.; Wu, W.H.; Chen, Y.F. Arabidopsis Di19 functions as a transcription factor and modulates PR1, PR2, and PR5 expression in response to drought stress. Mol. Plant 2013, 6, 1487–1502. [Google Scholar] [CrossRef]
- Vaghela, B.; Vashi, R.; Rajput, K.; Joshi, R. Plant chitinases and their role in plant defense: A comprehensive review. Enzyme Microb. Technol. 2022, 159, 110055. [Google Scholar] [CrossRef]
- Agarwal, P.; Agarwal, P.K. Pathogenesis related-10 proteins are small, structurally similar but with diverse role in stress signaling. Mol. Biol. Rep. 2014, 41, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Long, J.; He, X.; Li, S.; Xu, H. Molecular cloning and characterization of pathogenesis-related protein family 10 gene from spinach (SoPR10). Biosci. Biotechnol. Biochem. 2014, 78, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, H.; Rajput, R.; Pandey, A.; Upadhyay, S.K. Molecular characterization revealed the role of thaumatin-like proteins of bread wheat in stress response. Front. Plant Sci. 2022, 12, 807448. [Google Scholar] [CrossRef] [PubMed]
- Kothari, K.S.; Dansana, P.K.; Giri, J.; Tyagi, A.K. Rice stress associated protein 1 (OsSAP1) interacts with aminotransferase (OsAMTR1) and pathogenesis-related 1a protein (OsSCP) and regulates abiotic stress responses. Front. Plant Sci. 2016, 7, 1057–1072. [Google Scholar] [CrossRef]
- Garcia-Cristobal, J.; Garcia-Villaraco, A.; Ramos, B.; Gutierrez-Manero, J.; Lucas, J.A. Priming of pathogenesis related-proteins and enzymes related to oxidative stress by plant growth promoting rhizobacteria on rice plants upon abiotic and biotic stress challenge. J. Plant Physiol. 2015, 188, 72–79. [Google Scholar] [CrossRef]
- Wang, J.; Mao, X.; Wang, R.; Li, A.; Zhao, G.; Zhao, J.; Jing, R. Identification of wheat stress-responding genes and TaPR-1-1 function by screening a cDNA yeast library prepared following abiotic stress. Sci. Rep. 2019, 9, 141. [Google Scholar] [CrossRef]
- Richards, K.D.; Snowden, K.C.; Cardner, R.C. Wali6 and wali7, genes induced by aluminum in wheat (Triticum aestivum) roots. Plant Physiol. 1994, 105, 1455–1456. [Google Scholar] [CrossRef]
- Shan, L.; Li, C.; Chen, F.; Zhao, S.; Xia, G. A Bowman-Birk type protease inhibitor is involved in the tolerance to salt stress in wheat. Plant Cell Environ. 2008, 31, 1128–1137. [Google Scholar] [CrossRef]
- Komatsu, S.; Diniyah, A.; Zhu, W.; Nakano, M.; Rehman, S.U.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K. Metabolomic and proteomic analyses to reveal the role of plant-derived smoke solution on wheat under salt stress. Int. J. Mol. Sci. 2024, 25, 8216. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Komatsu, S.; Han, C.; Nanjo, Y.; Altaf-Un-Nahar, M.; Wang, K.; He, D.; Yang, P. Label-free quantitative proteomic analysis of abscisic acid effect in early-stage soybean under flooding. J. Proteome Res. 2013, 12, 4769–4784. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Rehman, S.U.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Yamaguchi, T.; Sunohara, Y.; Matsumoto, H.; Komatsu, S. Proteomic analysis of the effect of plant-derived smoke on soybean during recovery from flooding stress. J. Proteom. 2018, 181, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote) omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Yamamoto, A.; Nakamura, T.; Nouri, M.Z.; Nanjo, Y.; Nishizawa, K.; Furukawa, K. Comprehensive analysis of mitochondria in roots and hypocotyls of soybean under flooding stress using proteomics and metabolomics techniques. J. Proteome Res. 2011, 10, 3993–4004. [Google Scholar] [CrossRef]
- Haque, M.E.; Abe, F.; Mori, M.; Oyanagi, A.; Komatsu, S.; Kawaguchi, K. Characterization of a wheat pathogenesis-related protein, TaBWPR-1.2, in seminal roots in response to waterlogging stress. J. Plant Physiol. 2014, 171, 602–609. [Google Scholar] [CrossRef]
- Masuda, T.; Yonayama, T.; Kataoka, T.; Komatsu, S. Analysis of protein in low-phosphorus-adapted ruzigrass (Brachiaria ruziziensis). In Plant Nutrition for Sustainable Food Production and Environment; Ando, T., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 331–332. [Google Scholar]
- Konishi, H.; Ishiguro, K.; Komatsu, S. A proteomics approach towards understanding blast fungus infection of rice grown under different levels of nitrogen fertilization. Proteomics 2001, 1, 1162–1171. [Google Scholar] [CrossRef]
- Rakwal, R.; Komatsu, S. Role of jasmonate in the rice (Oryza sativa L.) self-defense mechanism using proteome analysis. Electrophoresis 2000, 21, 2492–2500. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef]
- Vizcaíno, J.A.; Côté, R.G.; Csordas, A.; Dianes, J.A.; Fabregat, A.; Foster, J.M.; Griss, J.; Alpi, E.; Birim, M.; Contell, J.; et al. The PRoteomics IDEntifications (PRIDE) database and associated tools: Status in 2013. Nucleic Acids Res. 2013, 41, D1063–D1069. [Google Scholar] [CrossRef]
- Okuda, S.; Watanabe, Y.; Moriya, Y.; Kawano, S.; Yamamoto, T.; Matsumoto, M.; Takami, T.; Kobayashi, D.; Araki, N.; Yoshizawa, A.C.; et al. jPOSTTrepo: An international standard data repository for proteomes. Nucleic Acids Res. 2017, 45, D1107–D1111. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komatsu, S.; Koh, R.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K. Protein-Based Mechanism of Wheat Growth Under Salt Stress in Seeds Irradiated with Millimeter Waves. Int. J. Mol. Sci. 2025, 26, 253. https://doi.org/10.3390/ijms26010253
Komatsu S, Koh R, Yamaguchi H, Hitachi K, Tsuchida K. Protein-Based Mechanism of Wheat Growth Under Salt Stress in Seeds Irradiated with Millimeter Waves. International Journal of Molecular Sciences. 2025; 26(1):253. https://doi.org/10.3390/ijms26010253
Chicago/Turabian StyleKomatsu, Setsuko, Rachel Koh, Hisateru Yamaguchi, Keisuke Hitachi, and Kunihiro Tsuchida. 2025. "Protein-Based Mechanism of Wheat Growth Under Salt Stress in Seeds Irradiated with Millimeter Waves" International Journal of Molecular Sciences 26, no. 1: 253. https://doi.org/10.3390/ijms26010253
APA StyleKomatsu, S., Koh, R., Yamaguchi, H., Hitachi, K., & Tsuchida, K. (2025). Protein-Based Mechanism of Wheat Growth Under Salt Stress in Seeds Irradiated with Millimeter Waves. International Journal of Molecular Sciences, 26(1), 253. https://doi.org/10.3390/ijms26010253