
PTHrP Promotes RBP4 Expression Under the Control of PPARγ in the Kidney

 ,
,  and
and
Abstract
1. Introduction
2. Results
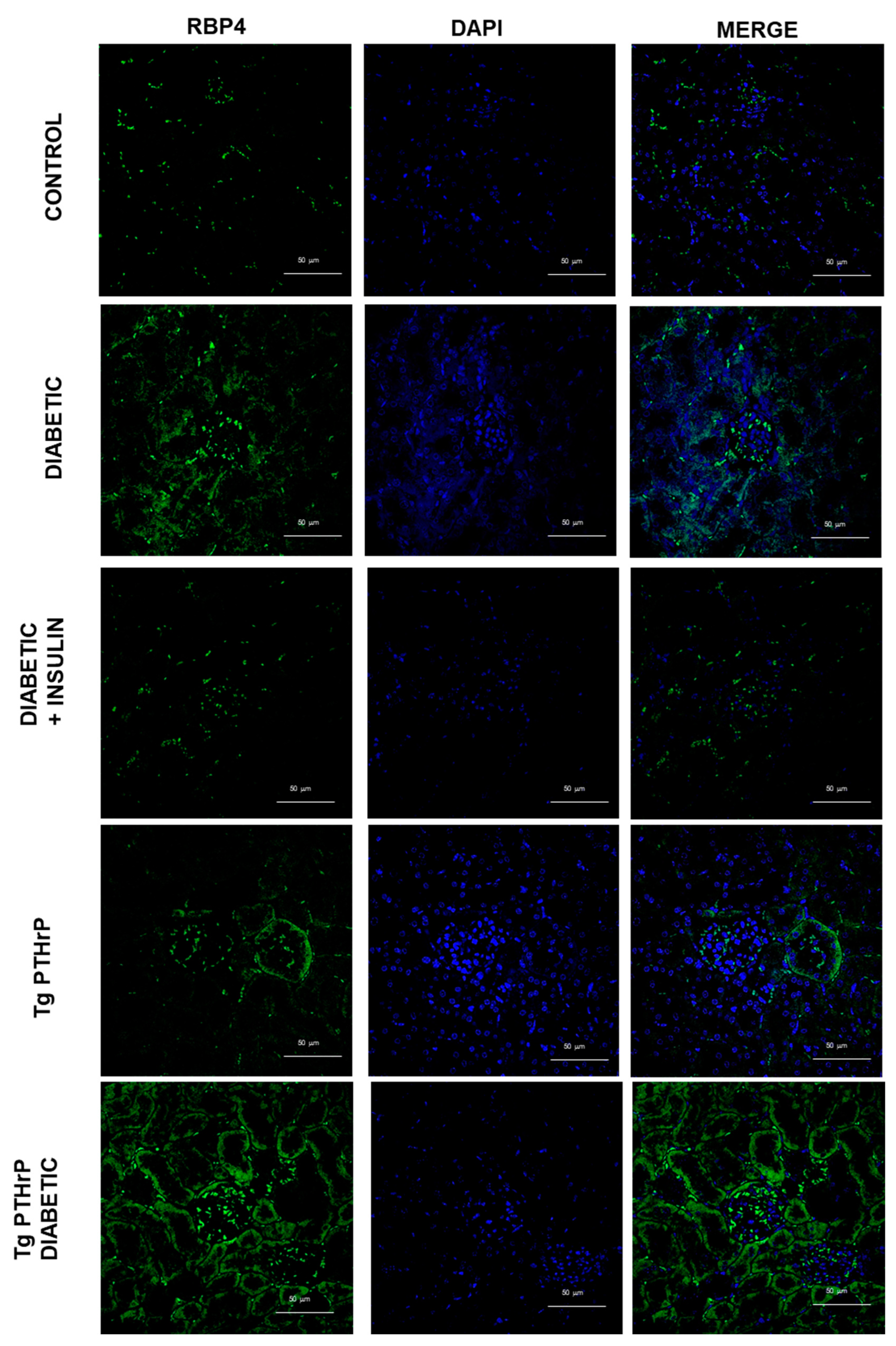
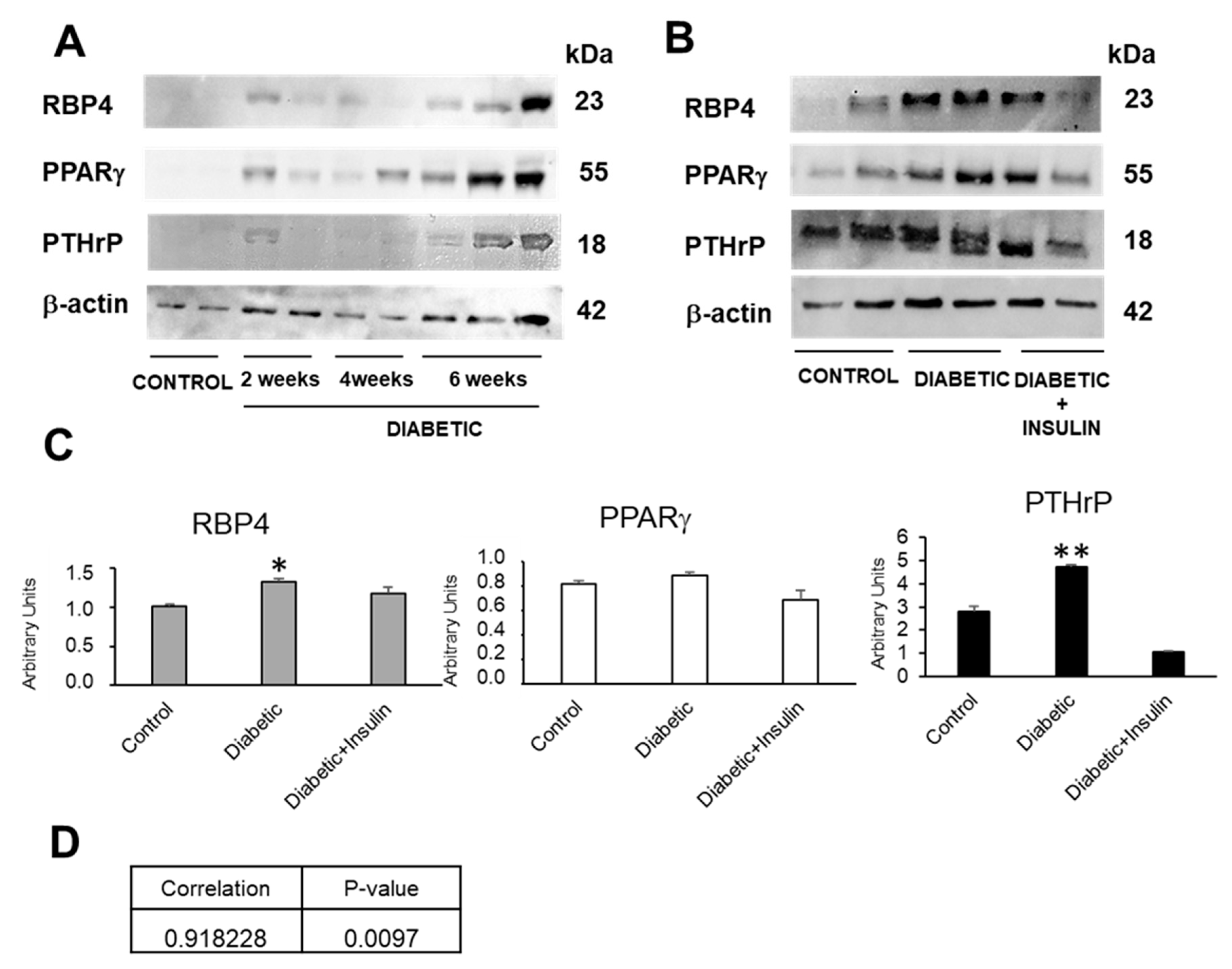
2.1. PTHrP and Diabetes Promote an Increased RBP4 Expression in the Kidney
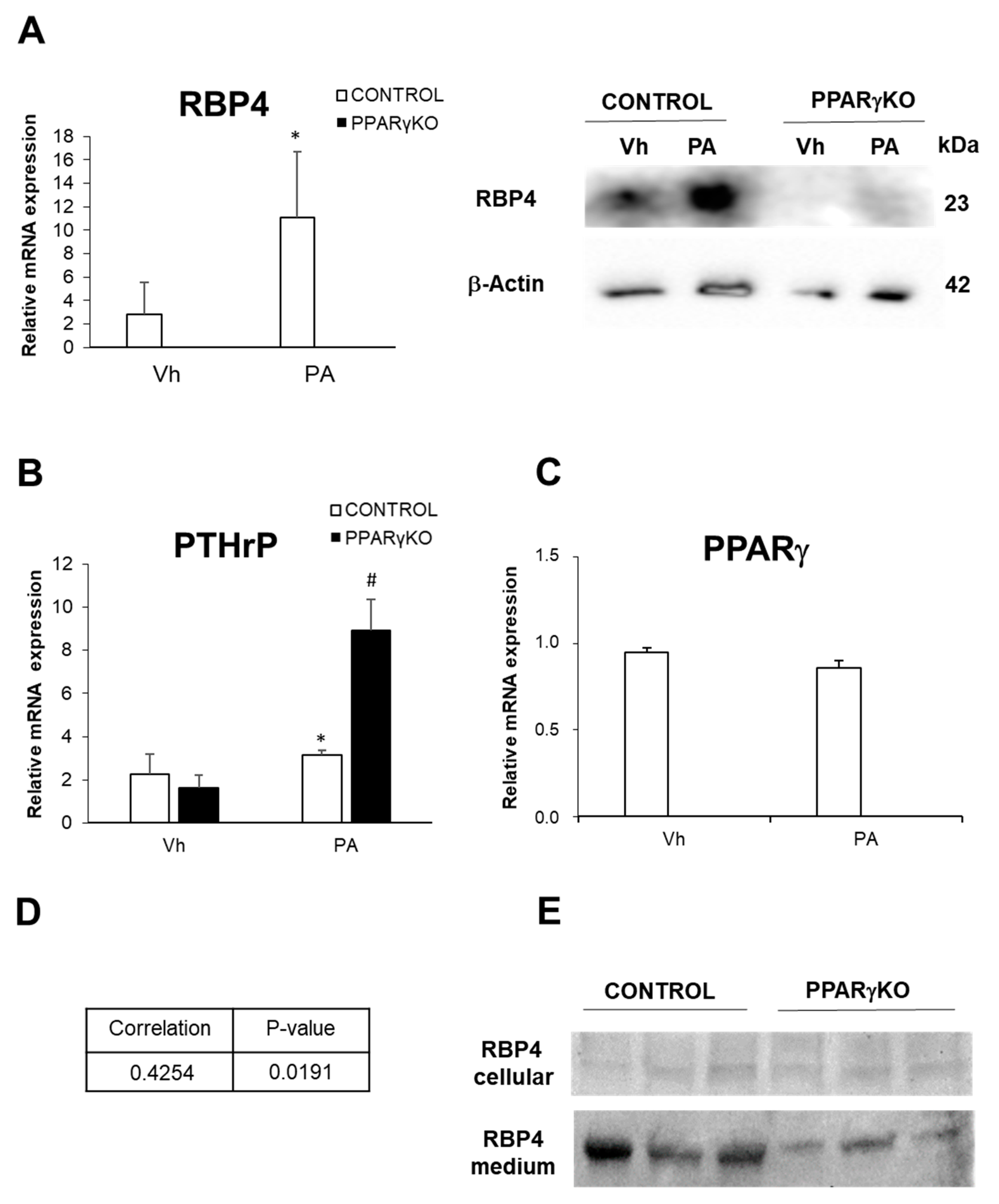
2.2. PA Stimulates RBP4 and PTHrP Expression in Podocytes
2.3. PPARγ Deficiency in Podocytes Promotes a Significant Increase in PTHrP and a Decrease in RBP4 Expression Following Lipotoxic Damage by Palmitic Acid
2.4. Podocytes Excrete RBP4
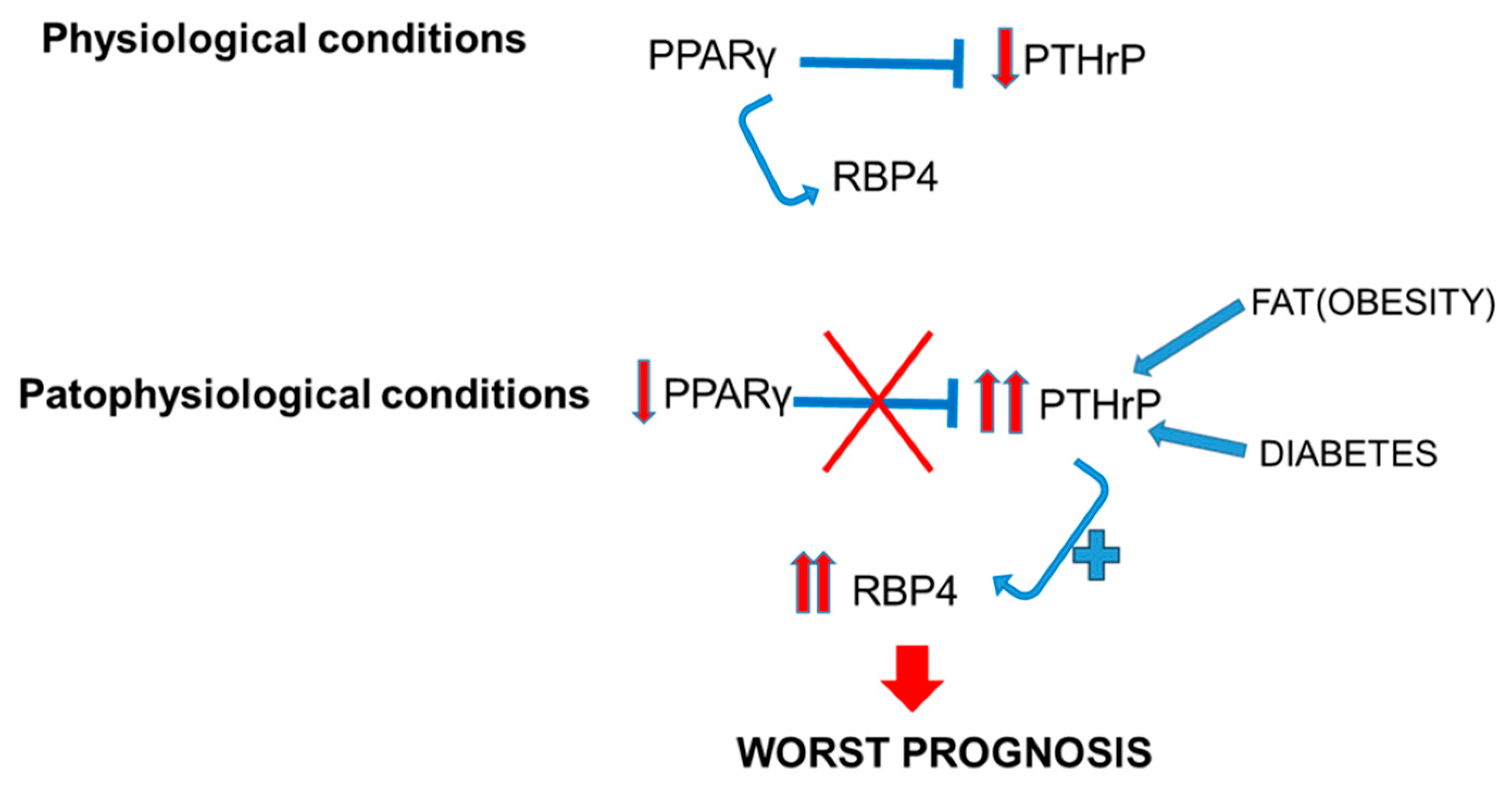
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Lines, Culture, and Treatments
4.3. mRNA Extraction and Quantitative RT-PCR
4.4. Protein Extraction and Western Blotting
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steinhoff, J.S.; Lass, A.; Schupp, M. Biological Functions of RBP4 and Its Relevance for Human Diseases. Front. Physiol. 2021, 12, 659977. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, A.G.; Izquierdo-Lahuerta, A.; Valverde, Á.M.; Ni, L.; Flores-Salguero, E.; Coward, R.J.; Medina-Gómez, G. The Protective Role of Peroxisome Proliferator-Activated Receptor Gamma in Lipotoxic Podocytes. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2023, 1868, 159329. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Zhong, G.; Jin, T.; Hu, W.; Wang, J.; Shi, B.; Wei, R. Diagnostic Value of Retinol-Binding Protein 4 in Diabetic Nephropathy: A Systematic Review and Meta-Analysis. Front. Endocrinol. 2024, 15, 1356131. [Google Scholar] [CrossRef] [PubMed]
- Corrales, P.; Izquierdo-Lahuerta, A.; Medina-Gómez, G. Maintenance of Kidney Metabolic Homeostasis by PPAR Gamma. Int. J. Mol. Sci. 2018, 19, 2063. [Google Scholar] [CrossRef]
- Platt, C.; Coward, R.J. Peroxisome Proliferator Activating Receptor-γ and the Podocyte. Nephrol. Dial. Transplant. 2016, 32, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Ahn, J.; Suh, Y.; Ziouzenkova, O.; Lee, J.W.; Lee, K. Retinol Binding Protein 7 Promotes Adipogenesis in Vitro and Regulates Expression of Genes Involved in Retinol Metabolism. Front. Cell Dev. Biol. 2022, 10, 876031. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Keen, H.L.; Lu, K.T.; Liu, X.; Wu, J.; Davis, D.R.; Ibeawuchi, S.R.C.; Vogel, S.; Quelle, F.W.; Sigmund, C.D. Retinol-Binding Protein 7 Is an Endothelium-Specific PPARγ Cofactor Mediating an Antioxidant Response through Adiponectin. JCI Insight 2017, 2, e91738. [Google Scholar] [CrossRef]
- Woll, A.W.; Quelle, F.W.; Sigmund, C.D. PPARγ and Retinol Binding Protein 7 Form a Regulatory Hub Promoting Antioxidant Properties of the Endothelium. Physiol. Genom. 2017, 49, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Zizola, C.F.; Schwartz, G.J.; Vogel, S. Cellular Retinol-Binding Protein Type III Is a PPARγ Target Gene and Plays a Role in Lipid Metabolism. Am. J. Physiol. Metab. 2008, 295, E1358–E1368. [Google Scholar] [CrossRef]
- Berry, D.C.; Noy, N. Signaling by Vitamin A and Retinol-Binding Protein in Regulation of Insulin Responses and Lipid Homeostasis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2012, 1821, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Librizzi, M.; Naselli, F.; Abruscato, G.; Luparello, C.; Caradonna, F. Parathyroid Hormone Related Protein (PTHrP)-Associated Molecular Signatures in Tissue Differentiation and Non-Tumoral Diseases. Biology 2023, 12, 950. [Google Scholar] [CrossRef] [PubMed]
- Massfelder, T.; Helwig, J.-J. The Parathyroid Hormone-Related Protein System: More Data but More Unsolved Questions. Curr. Opin. Nephrol. Hypertens. 2003, 12, 35–42. [Google Scholar] [CrossRef]
- Martin, T.J.; Johnson, R.W. Multiple Actions of Parathyroid Hormone-Related Protein in Breast Cancer Bone Metastasis. Br. J. Pharmacol. 2021, 178, 1923–1935. [Google Scholar] [CrossRef]
- Kir, S.; White, J.P.; Kleiner, S.; Kazak, L.; Cohen, P.; Baracos, V.E.; Spiegelman, B.M. Tumour-Derived PTH-Related Protein Triggers Adipose Tissue Browning and Cancer Cachexia. Nature 2014, 513, 100–104. [Google Scholar] [CrossRef]
- Hu, W.; Xiong, H.; Ru, Z.; Zhao, Y.; Zhou, Y.; Xie, K.; Xiao, W.; Xiong, Z.; Wang, C.; Yuan, C.; et al. Extracellular Vesicles-Released Parathyroid Hormone-Related Protein from Lewis Lung Carcinoma Induces Lipolysis and Adipose Tissue Browning in Cancer Cachexia. Cell Death Dis. 2021, 12, 134. [Google Scholar] [CrossRef] [PubMed]
- Ardura, J.A.; Sanz, A.B.; Ortiz, A.; Esbrit, P. Parathyroid Hormone-Related Protein Protects Renal Tubuloepithelial Cells from Apoptosis by Activating Transcription Factor Runx2. Kidney Int. 2013, 83, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Ardura, J.A.; Rayego-Mateos, S.; Rámila, D.; Ruiz-Ortega, M.; Esbrit, P. Parathyroid Hormone-Related Protein Promotes Epithelial-Mesenchymal Transition. J. Am. Soc. Nephrol. 2010, 21, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Ortega, A.; Izquierdo, A.; López-Luna, P.; Bosch, R.J. Parathyroid Hormone-Related Protein Induces Hypertrophy in Podocytes via TGF-Β1 and P27Kip1: Implications for Diabetic Nephropathy. Nephrol. Dial. Transplant. 2010, 25, 2447–2457. [Google Scholar] [CrossRef]
- Ortega, A.; Romero, M.; Izquierdo, A.; Troyano, N.; Arce, Y.; Ardura, J.A.; Arenas, M.I.; Bover, J.; Esbrit, P.; Bosch, R.J. Parathyroid Hormone-Related Protein Is a Hypertrophy Factor for Human Mesangial Cells: Implications for Diabetic Nephropathy. J. Cell. Physiol. 2012, 227, 1980–1987. [Google Scholar] [CrossRef]
- Kramann, R.; Schneider, R.K. Parathyroid Hormone-Related Protein and Regulation of Cell Survival in the Kidney. Kidney Int. 2013, 83, 777–779. [Google Scholar] [CrossRef]
- Santos, S.; Bosch, R.J.; Ortega, A.; Largo, R.; Gazapo, R.; Jesu’, J.; Egido, J.; Esbrit, P. Up-Regulation of Parathyroid Hormone-Related Protein in Folic Acid-Induced Acute Renal Failure. Kidney Int. 2001, 60, 982–995. [Google Scholar] [CrossRef] [PubMed]
- Bosch, R.J.; Ortega, A.; Izquierdo, A.; Arribas, I.; Bover, J.; Esbrit, P. A Transgenic Mouse Model for Studying the Role of the Parathyroid Hormone-Related Protein System in Renal Injury. J. Biomed. Biotechnol. 2011, 2011, 290874. [Google Scholar] [CrossRef]
- Tang, V.A.S.; Villanueva, A.R. Utility of N-Terminal Parathyroid Hormone-Related Protein in Evaluating Hypercalcemia in Patients with CKD. J. Am. Soc. Nephrol. 2024, 35. [Google Scholar] [CrossRef]
- Romero, M.; Ortega, A.; Olea, N.; Arenas, M.I.; Izquierdo, A.; Bover, J.; Esbrit, P.; Bosch, R.J. Novel Role of Parathyroid Hormone-Related Protein in the Pathophysiology of the Diabetic Kidney: Evidence from Experimental and Human Diabetic Nephropathy. J. Diabetes Res. 2013, 2013, 162846. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, A.; López-Luna, P.; Ortega, A.; Romero, M.; Gutiérrez-Tarrés, M.A.; Arribas, I.; Álvarez, M.J.R.; Esbrit, P.; Bosch, R.J. The Parathyroid Hormone-Related Protein System and Diabetic Nephropathy Outcome in Streptozotocin-Induced Diabetes. Kidney Int. 2006, 69, 2171–2178. [Google Scholar] [CrossRef]
- Izquierdo-Lahuerta, A. The Parathyroid Hormone-Related Protein/Parathyroid Hormone 1 Receptor Axis in Adipose Tissue. Biomolecules 2021, 11, 1570. [Google Scholar] [CrossRef]
- Martinez-Garcia, C.; Izquierdo, A.; Velagapudi, V.; Vivas, Y.; Velasco, I.; Campbell, M.; Burling, K.; Cava, F.; Ros, M.; Oresic, M.; et al. Accelerated Renal Disease Is Associated with the Development of Metabolic Syndrome in a Glucolipotoxic Mouse Model. Dis. Model. Mech. 2012, 5, 636–648. [Google Scholar] [CrossRef]
- Kim, J.H.; Yamaori, S.; Tanabe, T.; Takagi, M.; Matsubara, T.; Okamoto, M.; Kimura, S.; Gonzalez, F.J. Lack of Epithelial PPARγ Causes Cystic Adenomatoid Malformations in Mouse Fetal Lung. Biochem. Biophys. Res. Commun. 2017, 491, 271–276. [Google Scholar] [CrossRef]
- Chan, G.K.; Deckelbaum, R.A.; Bolivar, I.; Goltzman, D.; Karaplis, A.C. PTHrP Inhibits Adipocyte Differentiation by Down-Regulating PPARγ Activity via a MAPK-Dependent Pathway. Endocrinology 2001, 142, 4900–4909. [Google Scholar] [CrossRef]
- Kawashima-Ohya, Y.; Kuruta, Y.; Yan, W.; Kawamoto, T.; Noshiro, M.; Kato, Y. Retinol-Binding Protein Is Produced by Rabbit Chondrocytes and Responds to Parathyroid Hormone (PTH)/PTH-Related Peptide-Cyclic Adenosine Monophosphate Pathway. Endocrinology 1999, 140, 1075–1081. [Google Scholar] [CrossRef]
- Kir, S.; Komaba, H.; Garcia, A.P.; Economopoulos, K.P.; Liu, W.; Lanske, B.; Hodin, R.A.; Spiegelman, B.M. PTH/PTHrP Receptor Mediates Cachexia in Models of Kidney Failure and Cancer. Cell Metab. 2016, 23, 315–323. [Google Scholar] [CrossRef]
- Chan, G.K.; Miao, D.; Deckelbaum, R.; Bolivar, I.; Karaplis, A.; Goltzman, D. Parathyroid Hormone-Related Peptide Interacts with Bone Morphogenetic Protein 2 to Increase Osteoblastogenesis and Decrease Adipogenesis in Pluripotent C3H10T1/2 Mesenchymal Cells. Endocrinology 2003, 144, 5511–5520. [Google Scholar] [CrossRef]
- Amizuka, N.; Karaplis, A.C.; Henderson, J.E.; Warshawsky, H.; Lipman, M.L.; Matsuki, Y.; Ejiri, S.; Tanaka, M.; Izumi, N.; Ozawa, H.; et al. Haploinsufficiency of Parathyroid Hormone-Related Peptide (PTHrP) Results in Abnormal Postnatal Bone Development. Dev. Biol. 1996, 175, 166–176. [Google Scholar] [CrossRef]
- Roca-Rodríguez, M.M.; El Bekay, R.; Garrido-Sanchez, L.; Gómez-Serrano, M.; Coin-Aragüez, L.; Oliva-Olivera, W.; Lhamyani, S.; Clemente-Postigo, M.; García-Santos, E.; De Diaz, R.L.; et al. Parathyroid Hormone-Related Protein, Human Adipose-Derived Stem Cells Adipogenic Capacity and Healthy Obesity. J. Clin. Endocrinol. Metab. 2015, 100, E826–E835. [Google Scholar] [CrossRef]
- Martínez-García, C.; Izquierdo-Lahuerta, A.; Vivas, Y.; Velasco, I.; Yeo, T.-K.; Chen, S.; Medina-Gomez, G. Renal Lipotoxicity-Associated Inflammation and Insulin Resistance Affects Actin Cytoskeleton Organization in Podocytes. PLoS ONE 2015, 10, e0142291. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Body Weight (g) | Kidney Weight (g) | Glucose (mg/dL) | Proteinuria (mg/24 h) |
|---|---|---|---|---|
| Control | 33.9 ± 0.1 | 0.23 ± 0.00 | 119.2 ± 4.7 | 1.9 ± 0.4 |
| Diabetic | 31.7 ± 0.2 | 0.30 ± 0.03 * | 606 ± 60 ** | 10.5 ± 1.2 * |
| Diabetic with insulin | 34.5 ± 1.0 | 0.29 ± 0.01 | 232 ± 17.1 * | 9.0 ± 3.0 |
| Tg PTHrP | 32.8 ± 2.2 | 0.21 ± 0.01 | 133.3 ± 13.5 | 2.2 ± 0.4 |
| Tg PTHrP diabetic | 28 ± 1.1 | 0.33 ± 0.01 * | 620 ± 58 ** | 12.3 ± 1.2 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nieto-Bona, M.P.; Carrasco, A.G.; Medina-Gomez, G.; Bosch, R.J.; Izquierdo-Lahuerta, A. PTHrP Promotes RBP4 Expression Under the Control of PPARγ in the Kidney. Int. J. Mol. Sci. 2025, 26, 142. https://doi.org/10.3390/ijms26010142
Nieto-Bona MP, Carrasco AG, Medina-Gomez G, Bosch RJ, Izquierdo-Lahuerta A. PTHrP Promotes RBP4 Expression Under the Control of PPARγ in the Kidney. International Journal of Molecular Sciences. 2025; 26(1):142. https://doi.org/10.3390/ijms26010142
Chicago/Turabian StyleNieto-Bona, María Paz, Almudena G. Carrasco, Gema Medina-Gomez, Ricardo J. Bosch, and Adriana Izquierdo-Lahuerta. 2025. "PTHrP Promotes RBP4 Expression Under the Control of PPARγ in the Kidney" International Journal of Molecular Sciences 26, no. 1: 142. https://doi.org/10.3390/ijms26010142
APA StyleNieto-Bona, M. P., Carrasco, A. G., Medina-Gomez, G., Bosch, R. J., & Izquierdo-Lahuerta, A. (2025). PTHrP Promotes RBP4 Expression Under the Control of PPARγ in the Kidney. International Journal of Molecular Sciences, 26(1), 142. https://doi.org/10.3390/ijms26010142