

Recent Advances in the Field of Amino Acid-Conjugated Aminoferrocenes—A Personal Perspective




Abstract
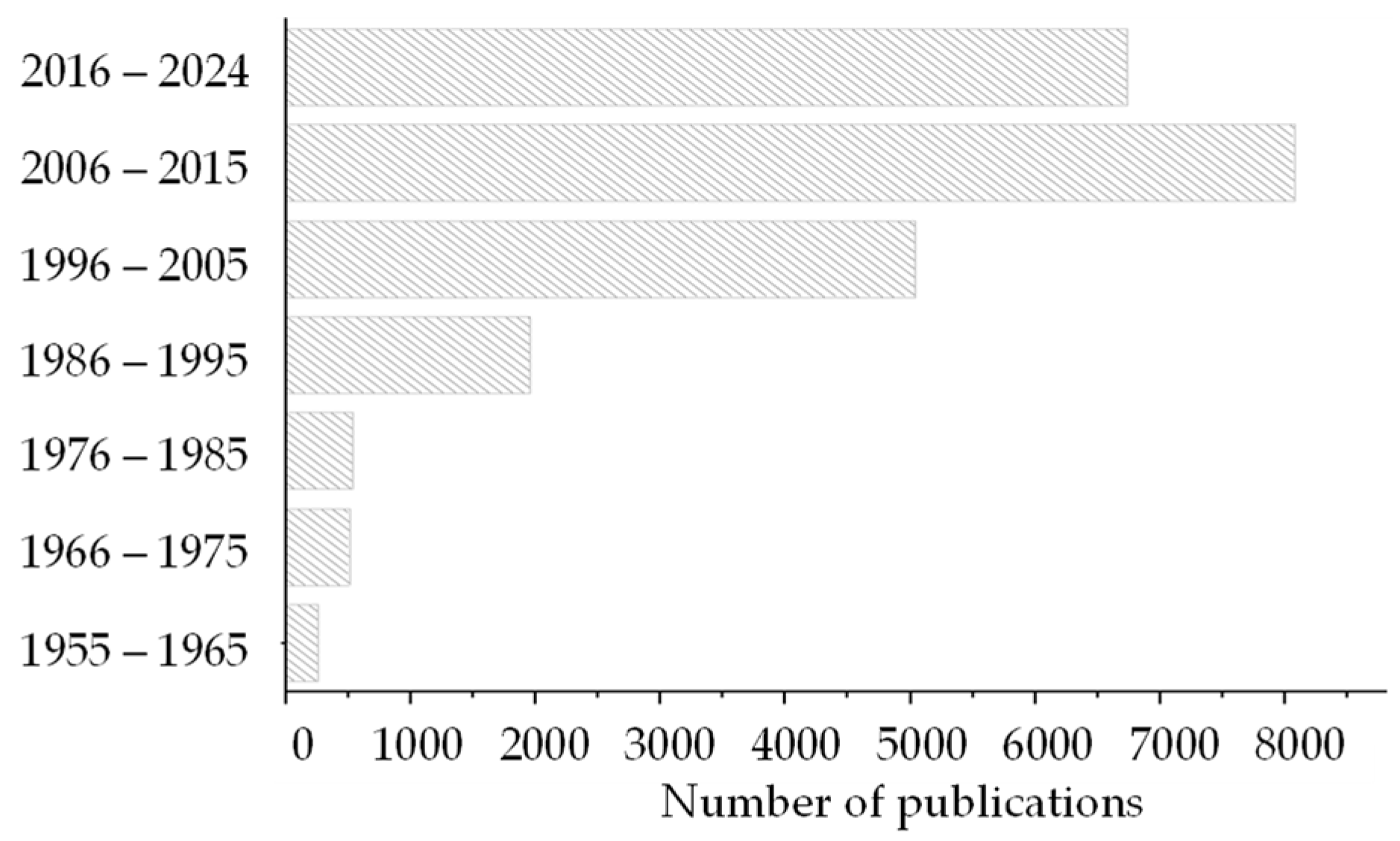
1. Introduction
2. The Turn-Inducing Capacity and Biological Activity of Conjugates 1–22
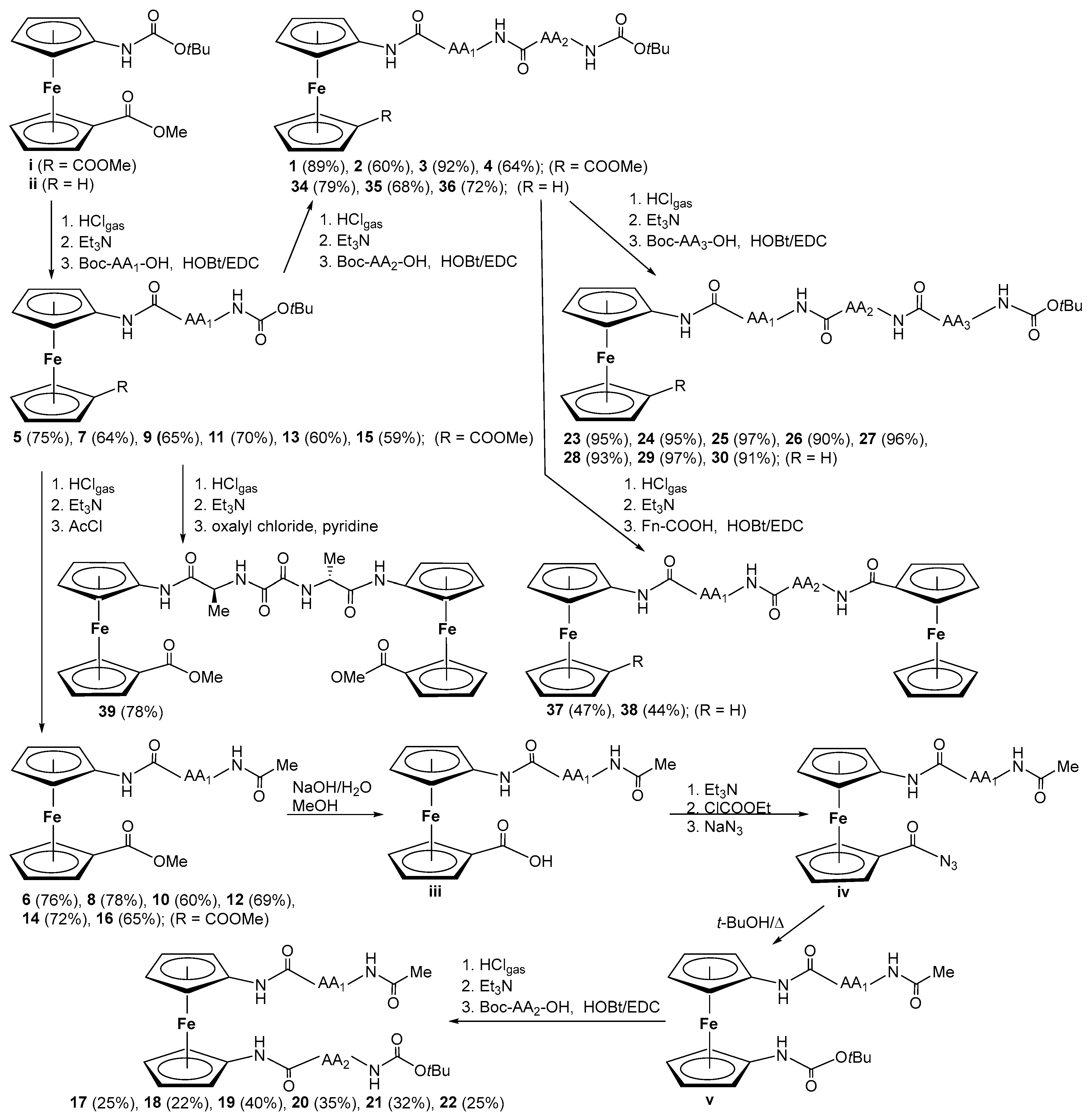
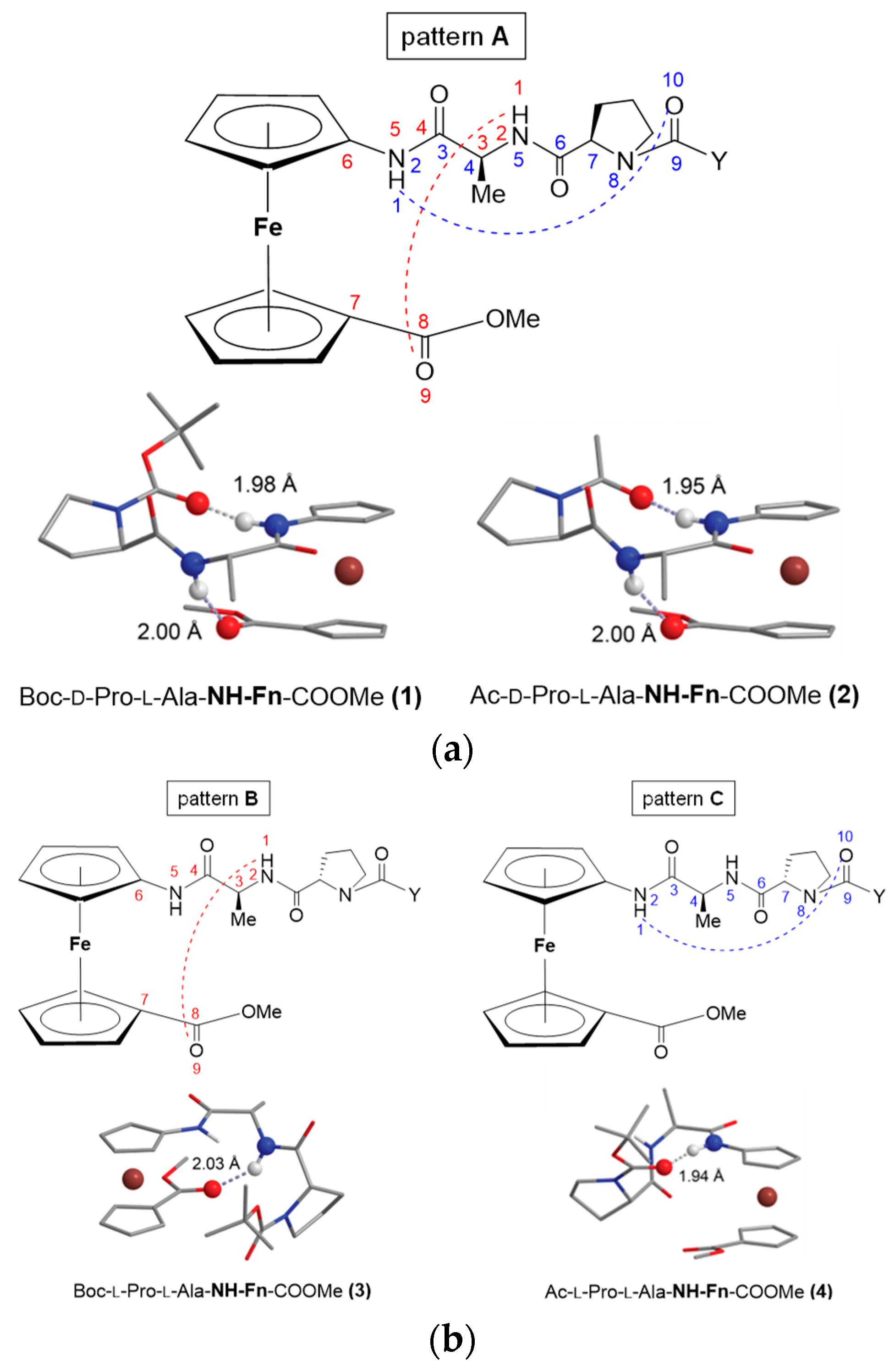
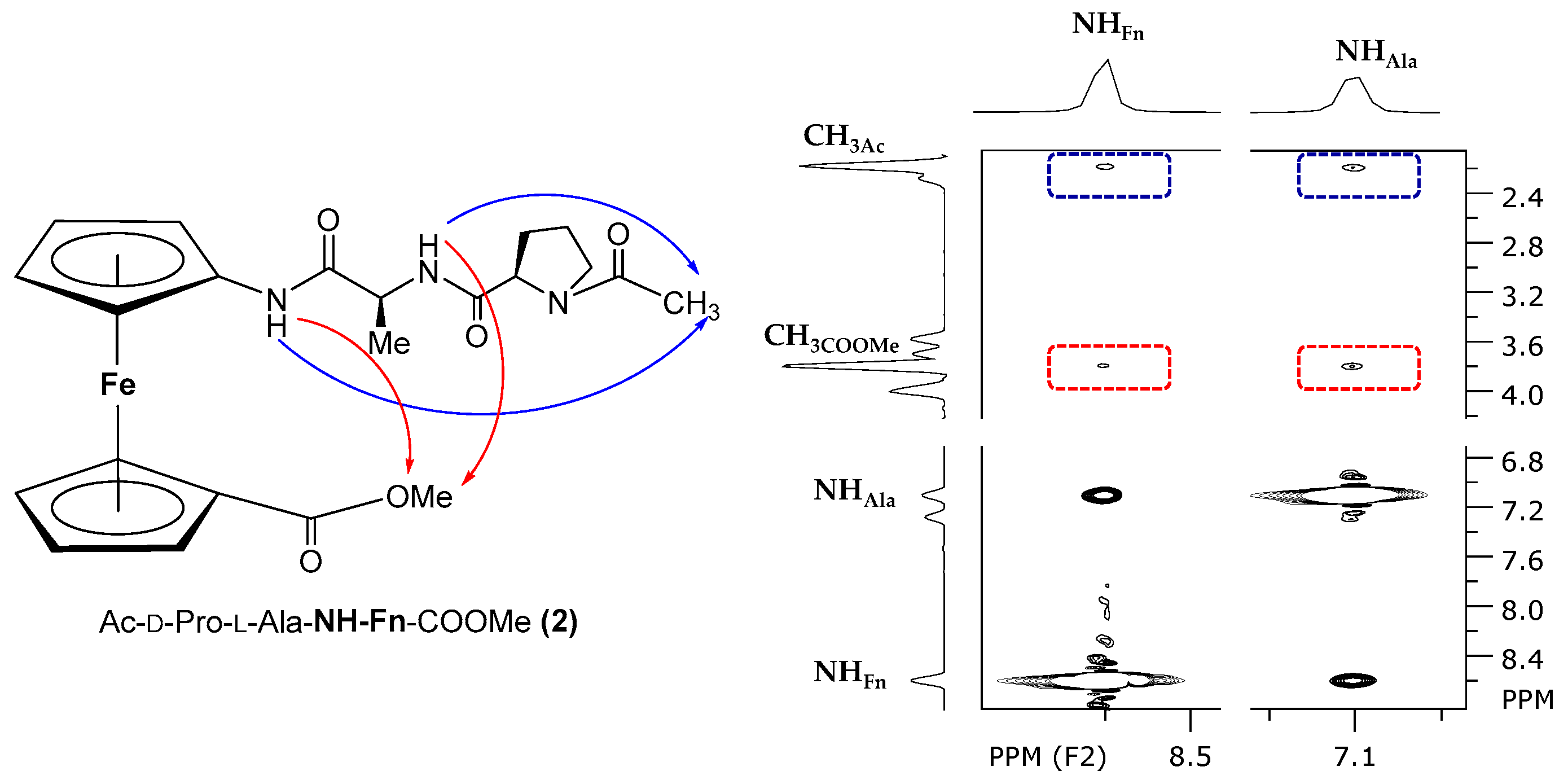
2.1. Conjugates of Amino-Functionalized Ferrocene with Homo- and Heterochiral Pro–Ala Dipeptides 1–4 [32]
2.1.1. Computational Study
2.1.2. Spectroscopic Study
- IR Spectroscopy
- 2.
- Concentration-dependent IR Spectroscopy
- 3.
- NMR Spectroscopy
- 4.
- Concentration-dependent NMR Spectroscopy
- 5.
- Solvent-dependent NMR Spectroscopy
- 6.
- Temperature-dependent NMR Spectroscopy
- 7.
- NOESY Spectroscopy
- 8.
- CD Spectroscopy
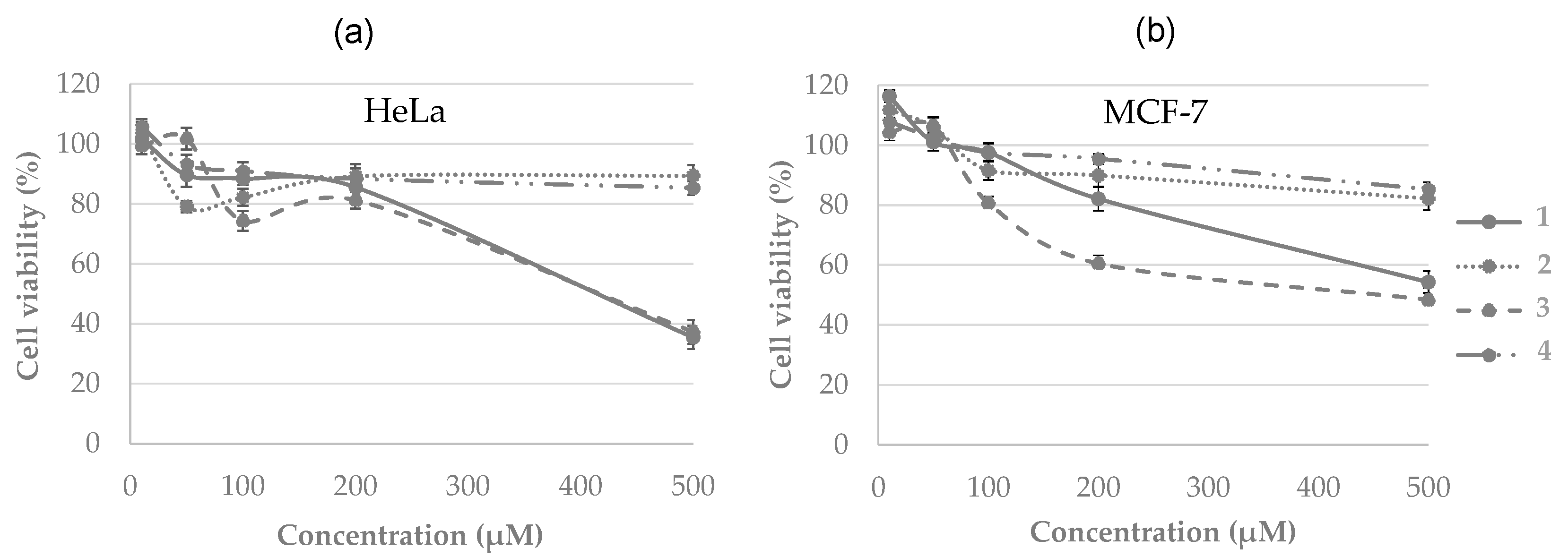
2.1.3. Biological Evaluation
- Antitumor activity
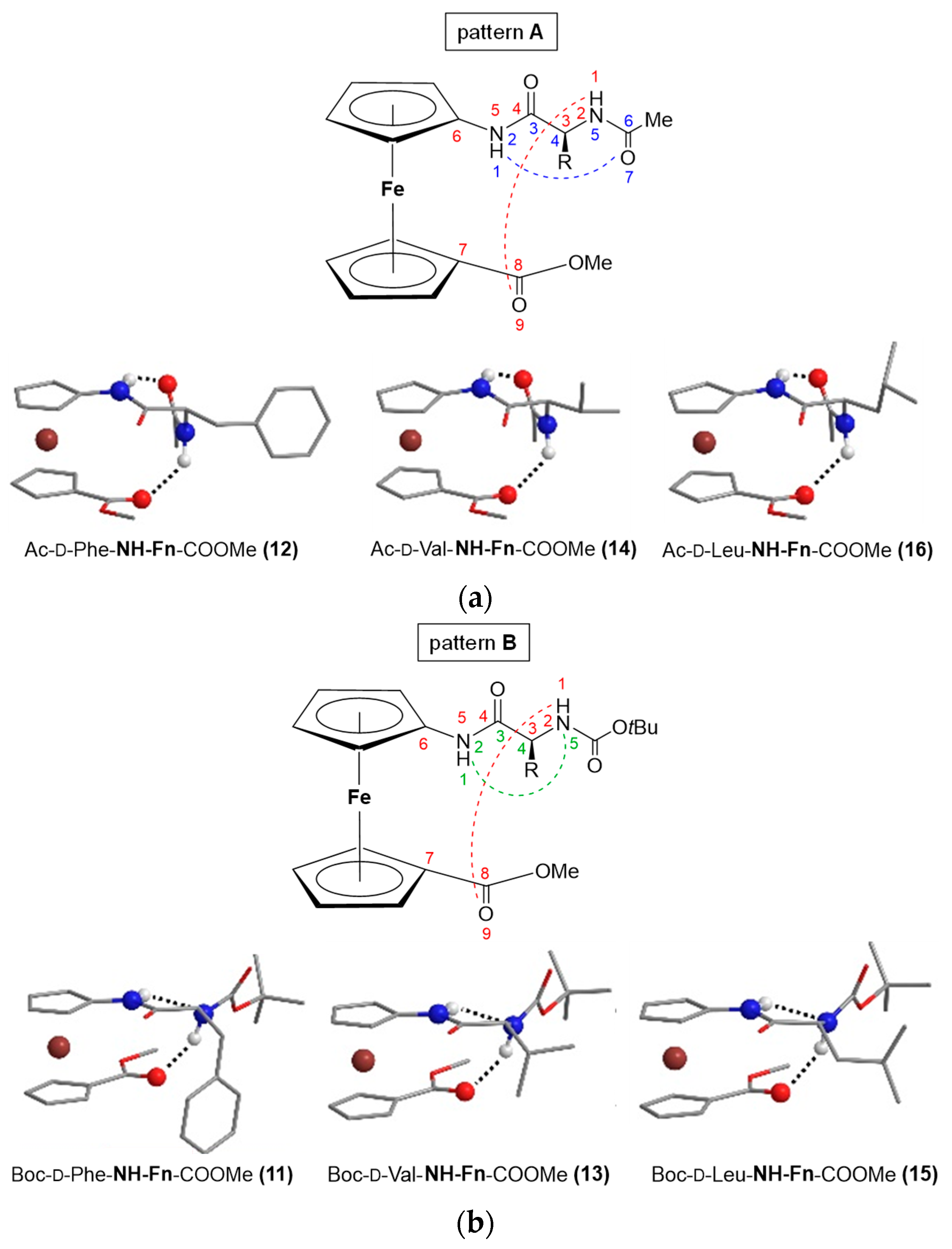
2.2. Unsymetrically Substituted Conjugates of Amino-Functionalized Ferrocene with Phe, Val, and Leu 5–16 [33]
2.2.1. Computational Study
2.2.2. Spectroscopic Study
- IR Spectroscopy
- 2.
- Concentration-dependent IR Spectroscopy
- 3.
- NMR Spectroscopy
- 4.
- Concentration-dependent NMR Spectroscopy
- 5.
- Solvent-dependent NMR Spectroscopy
- 6.
- Temperature-dependent NMR Spectroscopy
- 7.
- CD Spectroscopy
2.2.3. Biological Evaluation
- Antimicrobial activity
- 2.
- Antioxidant activity
- 3.
- Hydrophobicity
- 4.
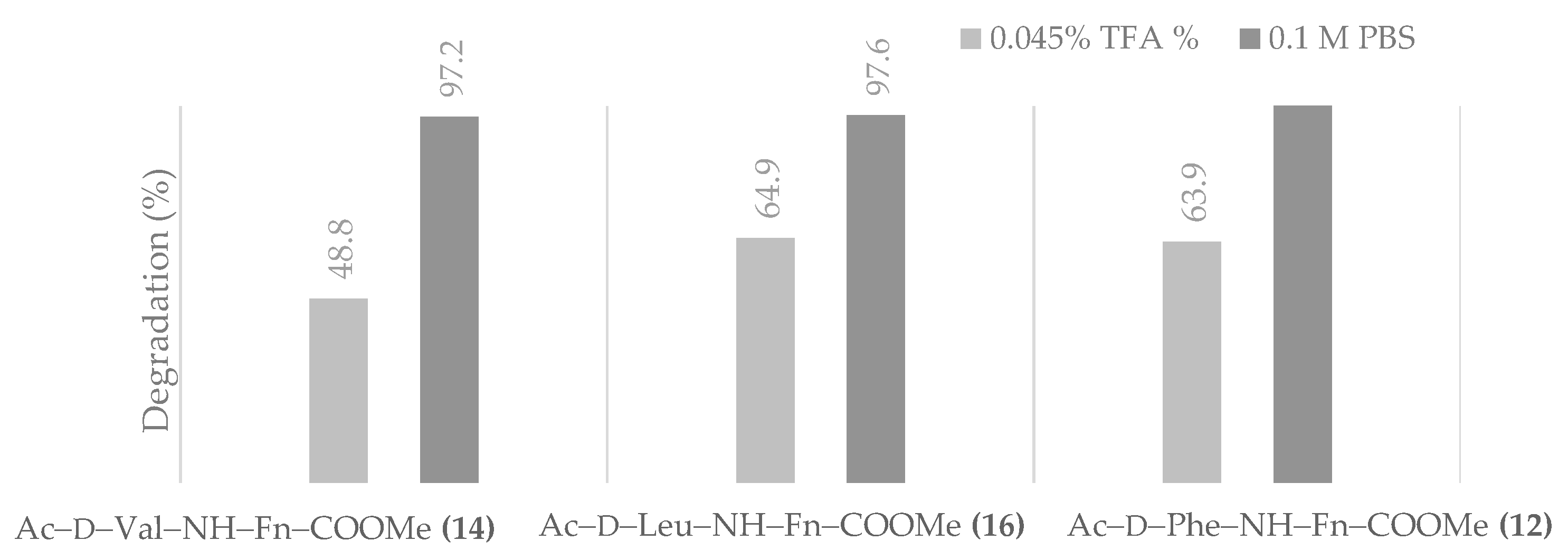
- Chemical and proteolytic stability
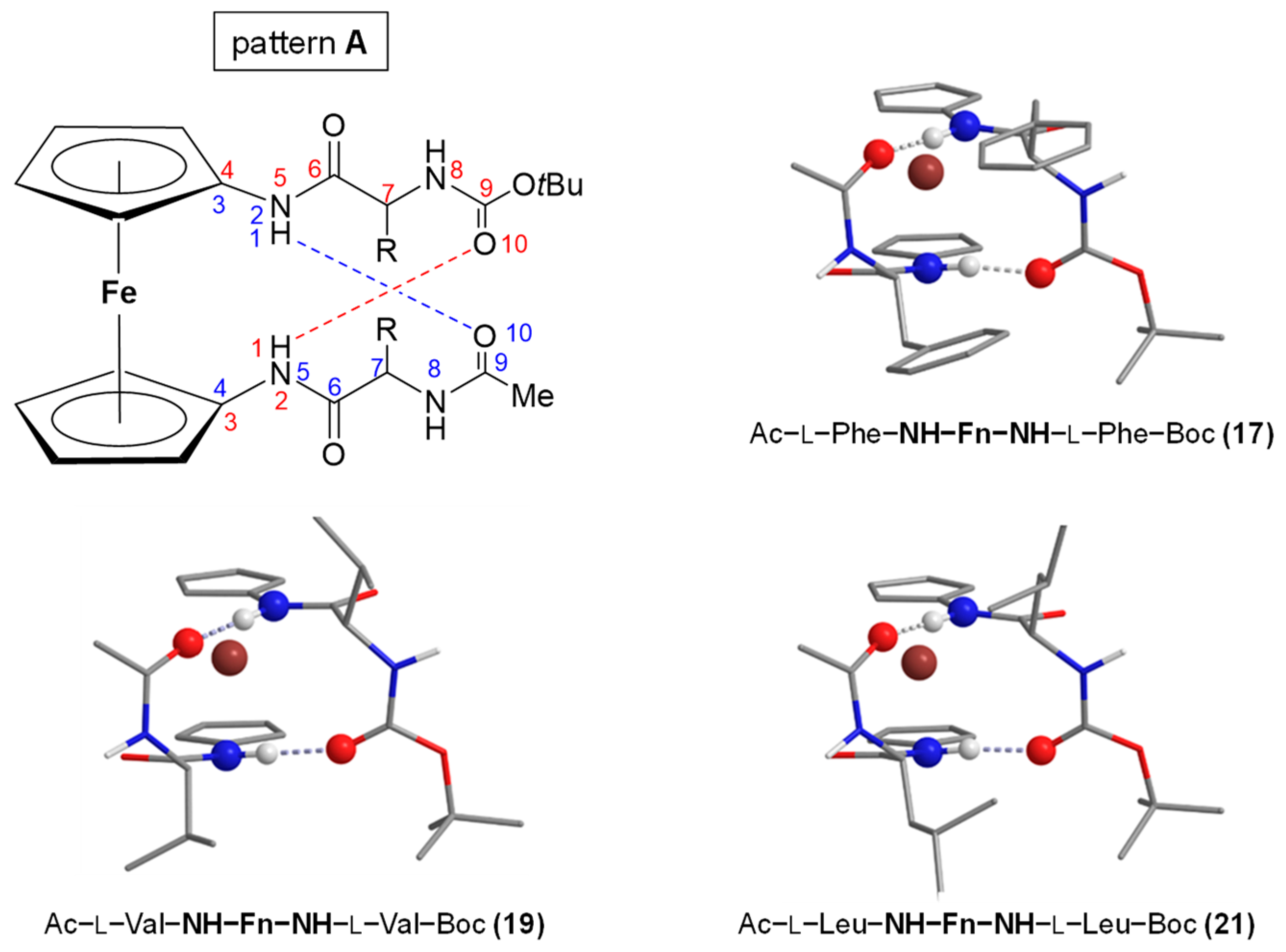
2.3. Symetrically Disubstituted Conjugates of Amino-Functionalized Ferrocene with Phe, Val and Leu 17–22 [34]
2.3.1. Computational Study
2.3.2. Spectroscopic Study
- IR Spectroscopy
- 2.
- Concentration-dependent IR Spectroscopy
- 3.
- NMR Spectroscopy
- 4.
- Concentration-dependent NMR Spectroscopy
- 5.
- Solvent-dependent NMR Spectroscopy
- 6.
- Temperature-dependent NMR Spectroscopy
- 7.
- NOESY Spectroscopy
- 8.
- CD Spectroscopy
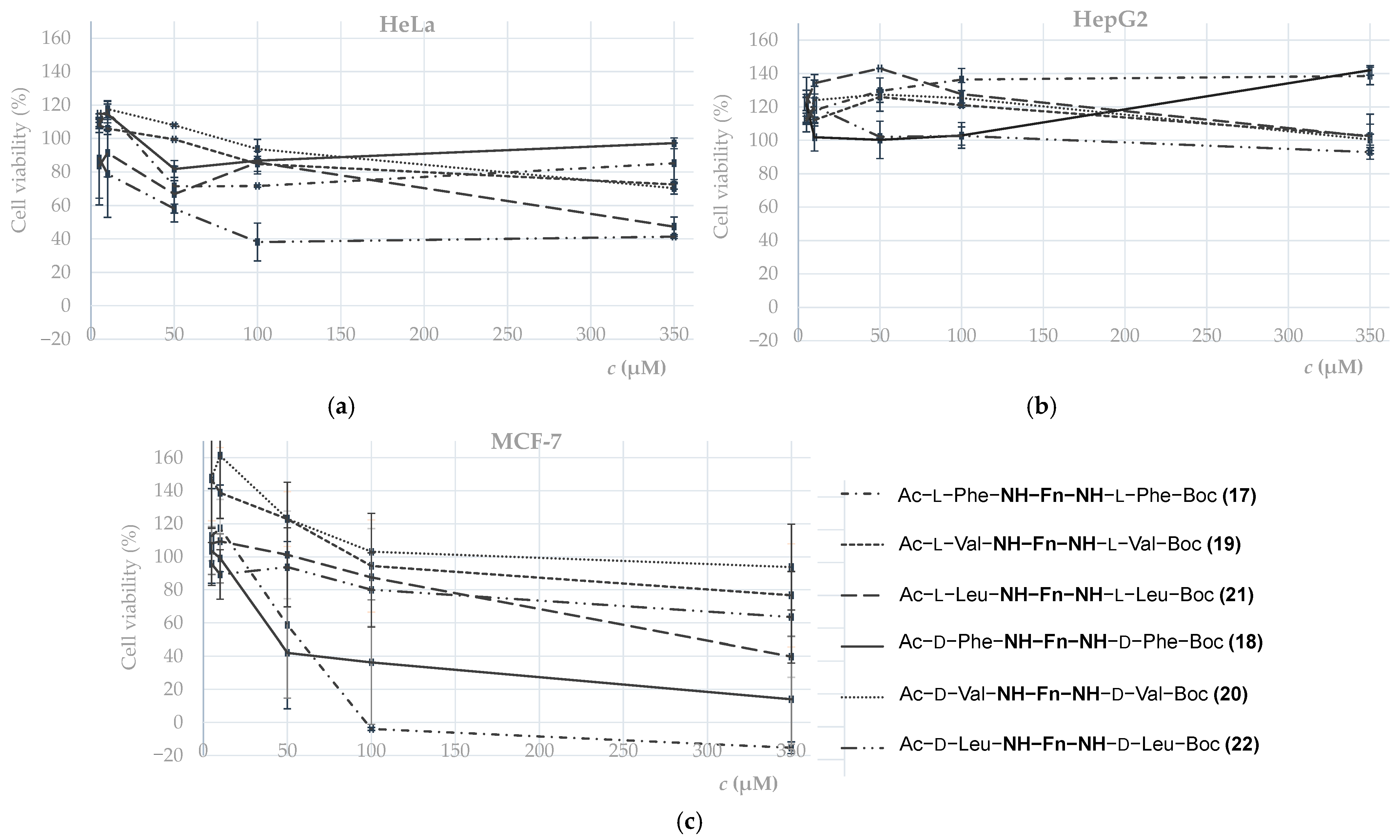
2.3.3. Biological Evaluation
- Antitumor activity
- 2.
- Antioxidant activity
3. Peptide Derivatives of Aminoferrocene—Origin of Chiroptical Activity
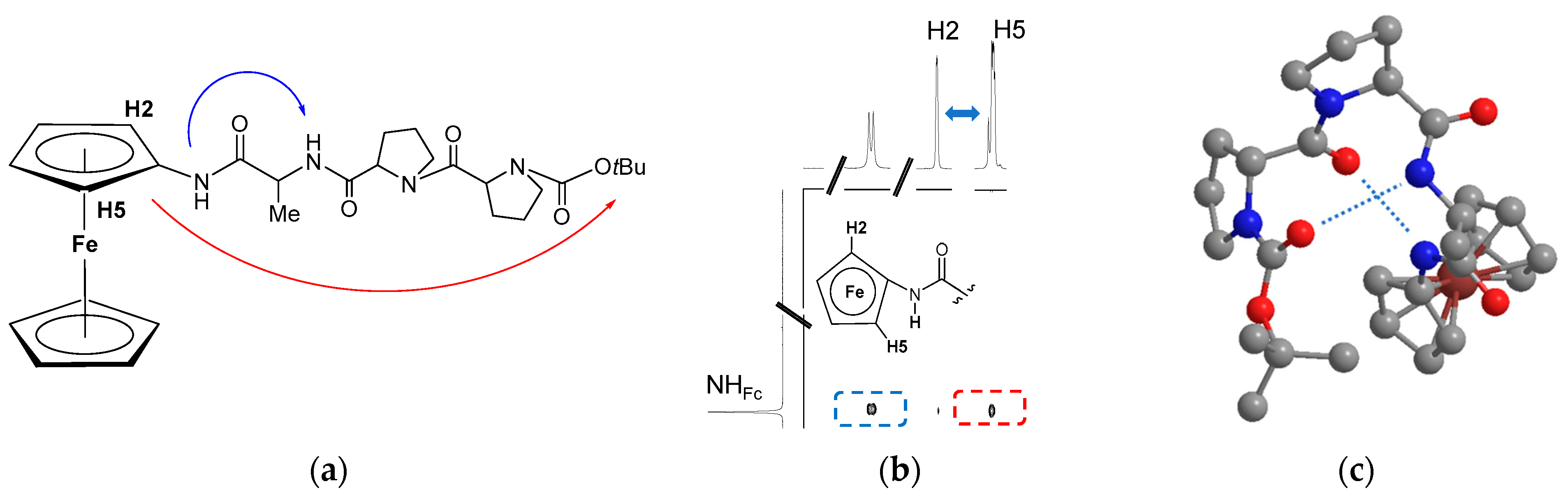
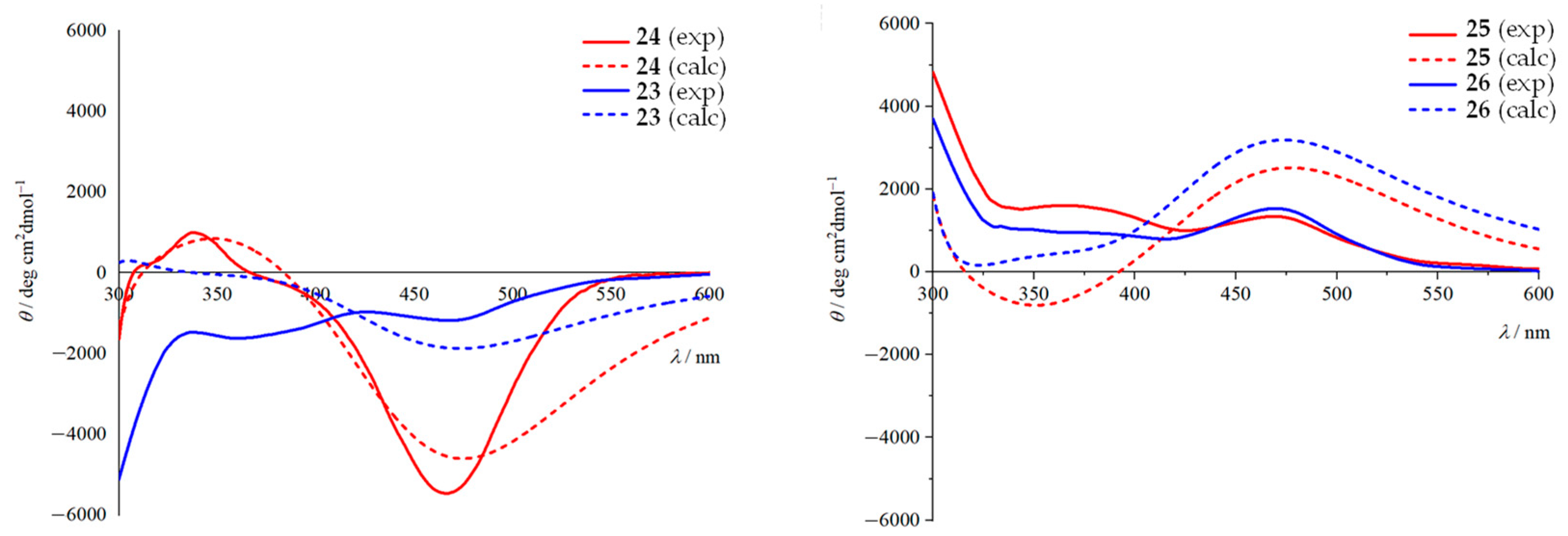
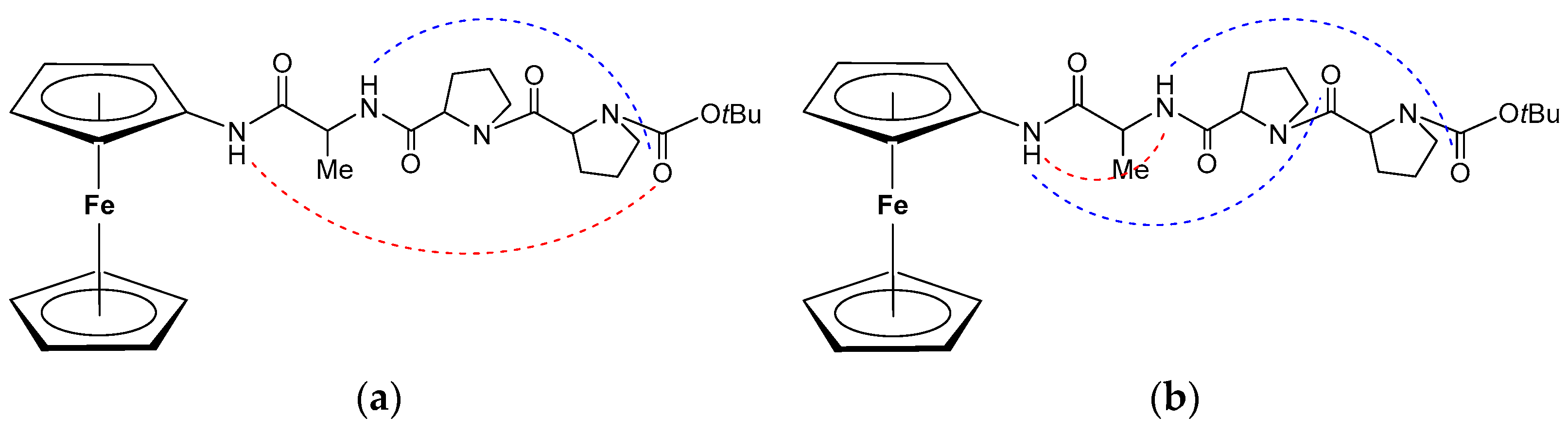
3.1. C-Terminal Ferrocene-Capped Tripeptides 23–30 [35]
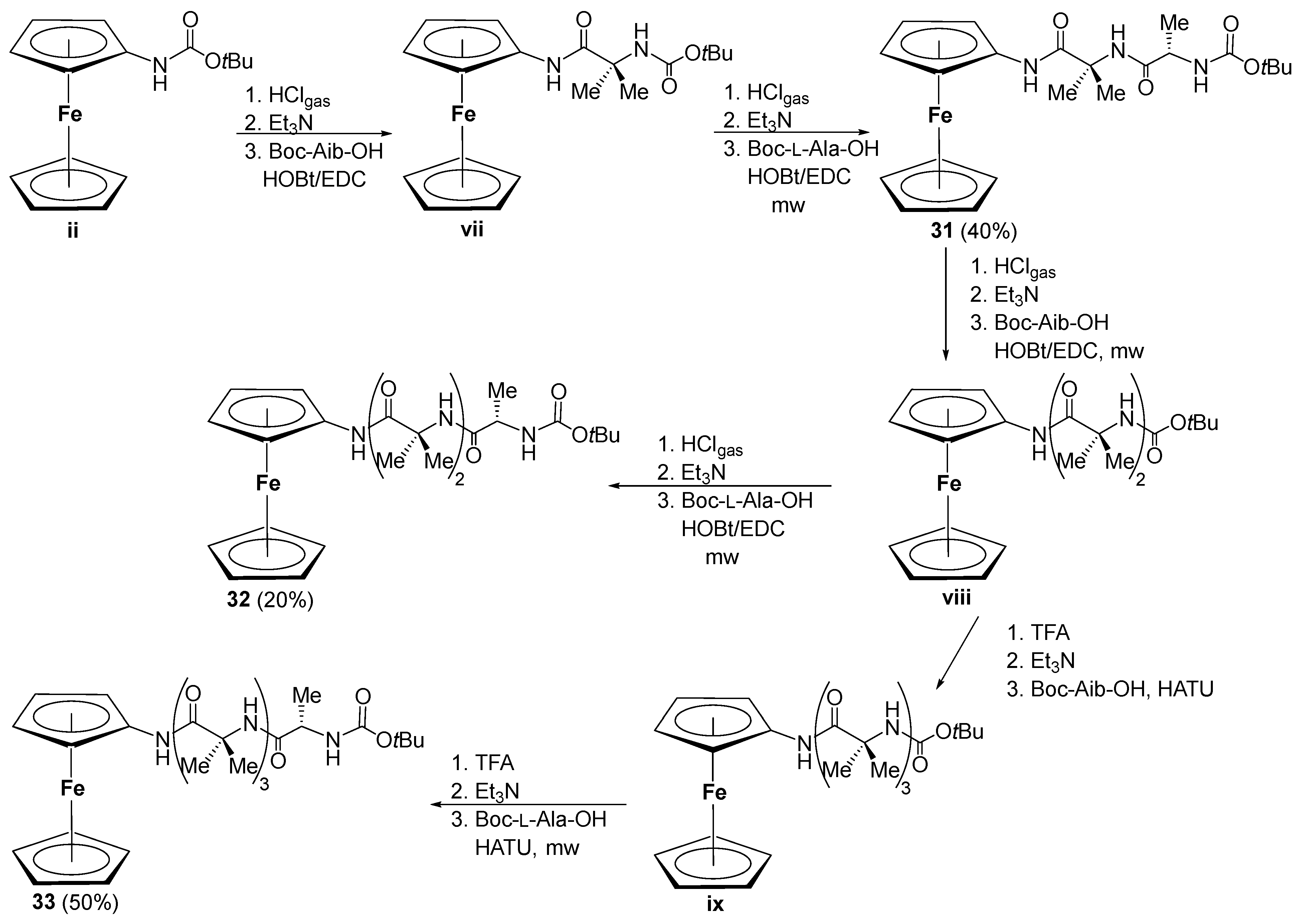
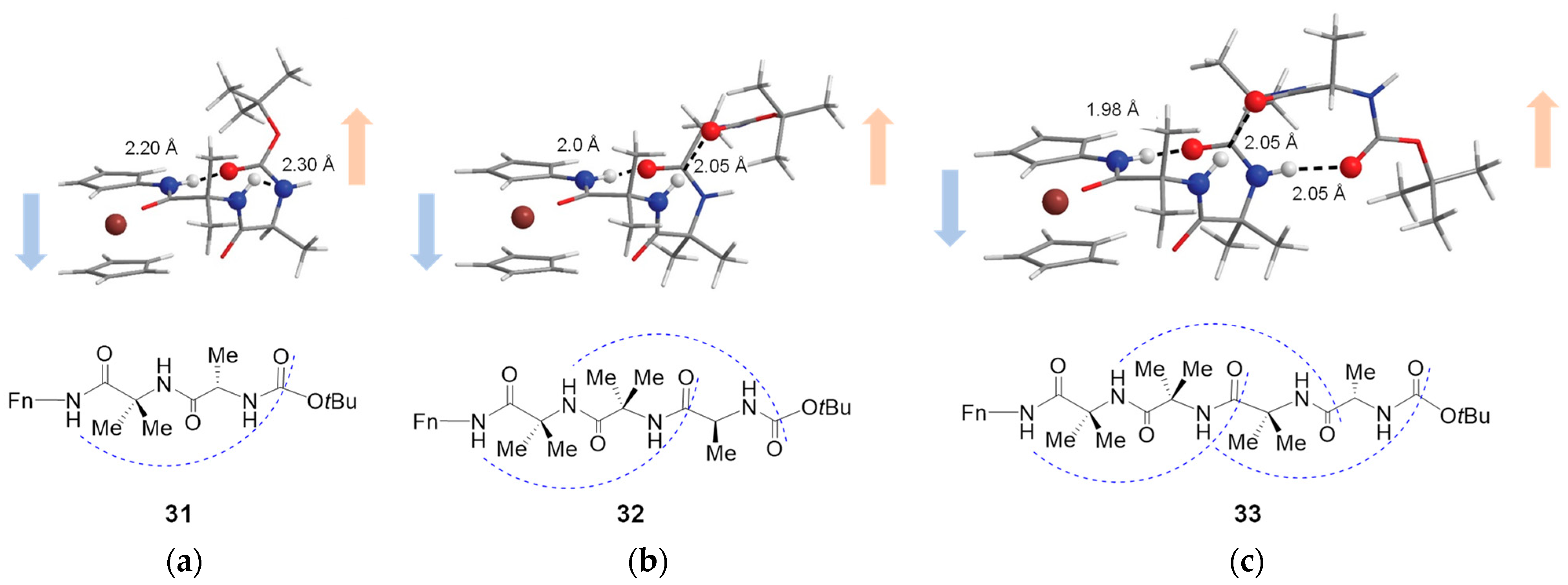
3.2. Transfer of Chiral Information in Aib Containing Aminoferrocene Peptides 31–33 [36]
3.3. Central-to-Helical-to-Axial Chirality Transfer in Ac6c Aminoferrocene Peptides 34–36 [37]
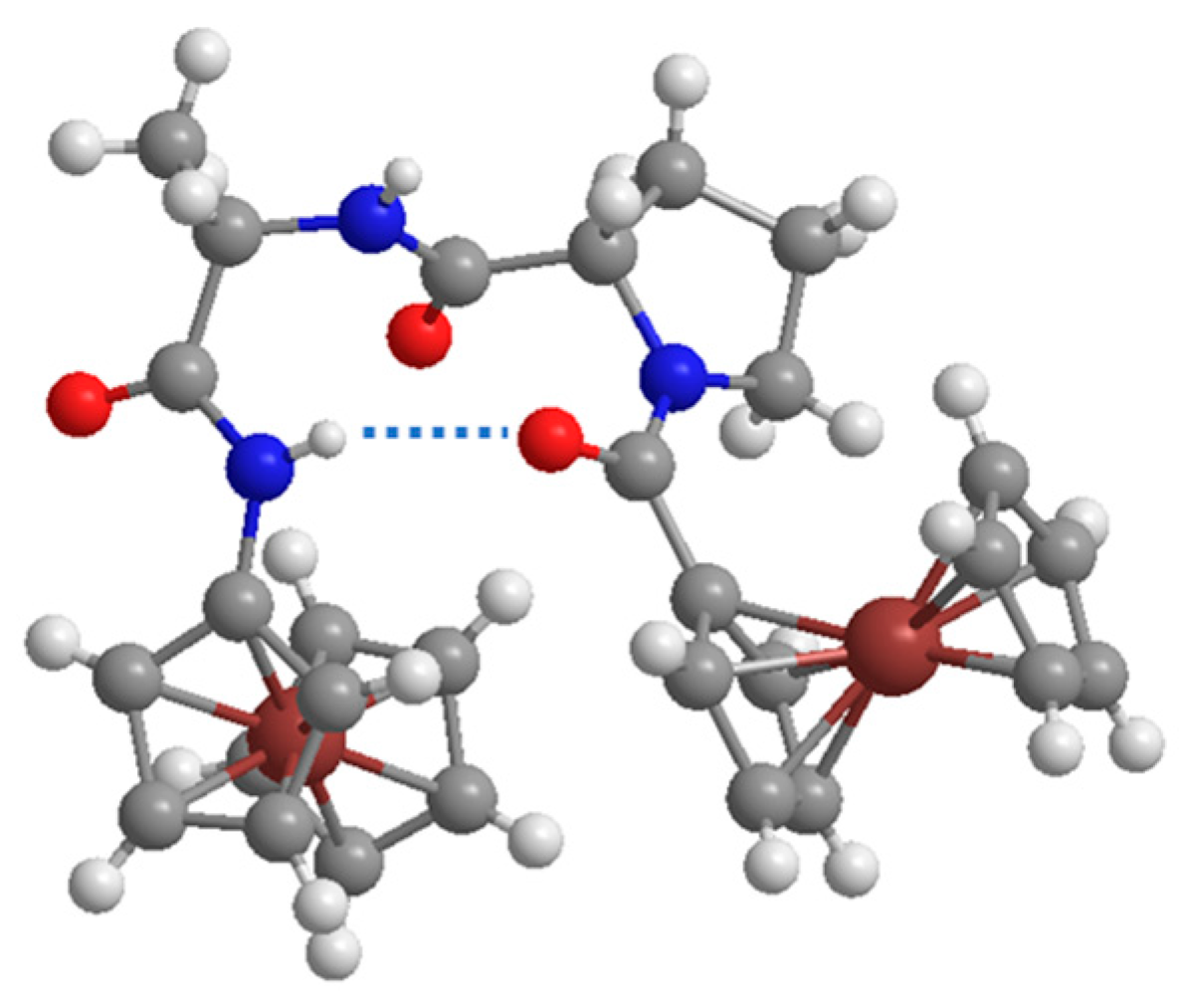
3.4. Dinuclear Ferrocene-Peptide Derivatives 37 and 38 [38]
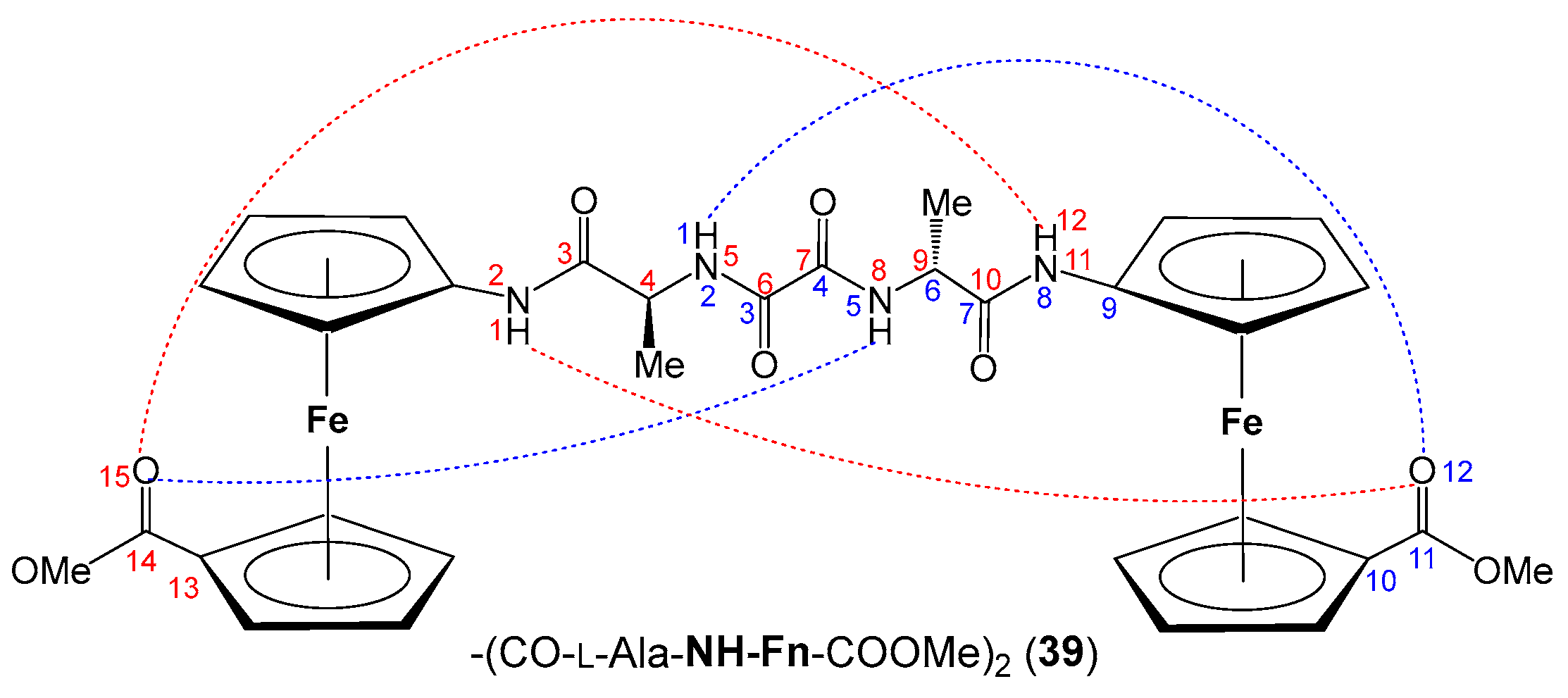
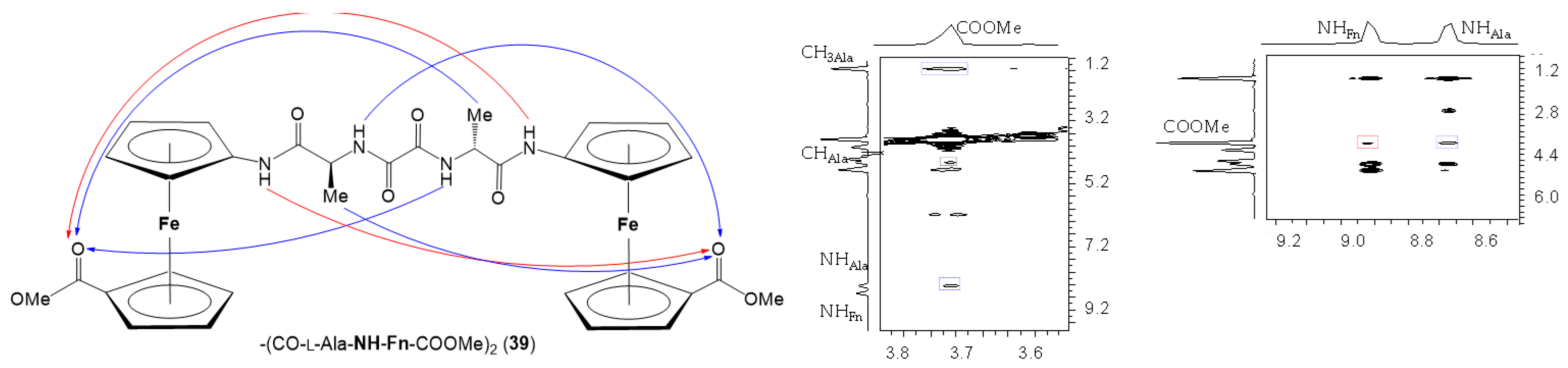
4. Oxalamide-Bridged Ferrocene 39 [39]
4.1. Computational Study
4.2. Spectroscopic Study
- IR Spectroscopy
- 2.
- Concentration-dependent IR Spectroscopy
- 3.
- NMR Spectroscopy
- 4.
- Concentration-dependent NMR Spectroscopy
- 5.
- Solvent-dependent NMR Spectroscopy
- 6.
- Temperature-dependent NMR Spectroscopy
- 7.
- NOESY Spectroscopy
- 8.
- CD Spectroscopy
4.3. Gelation Properties
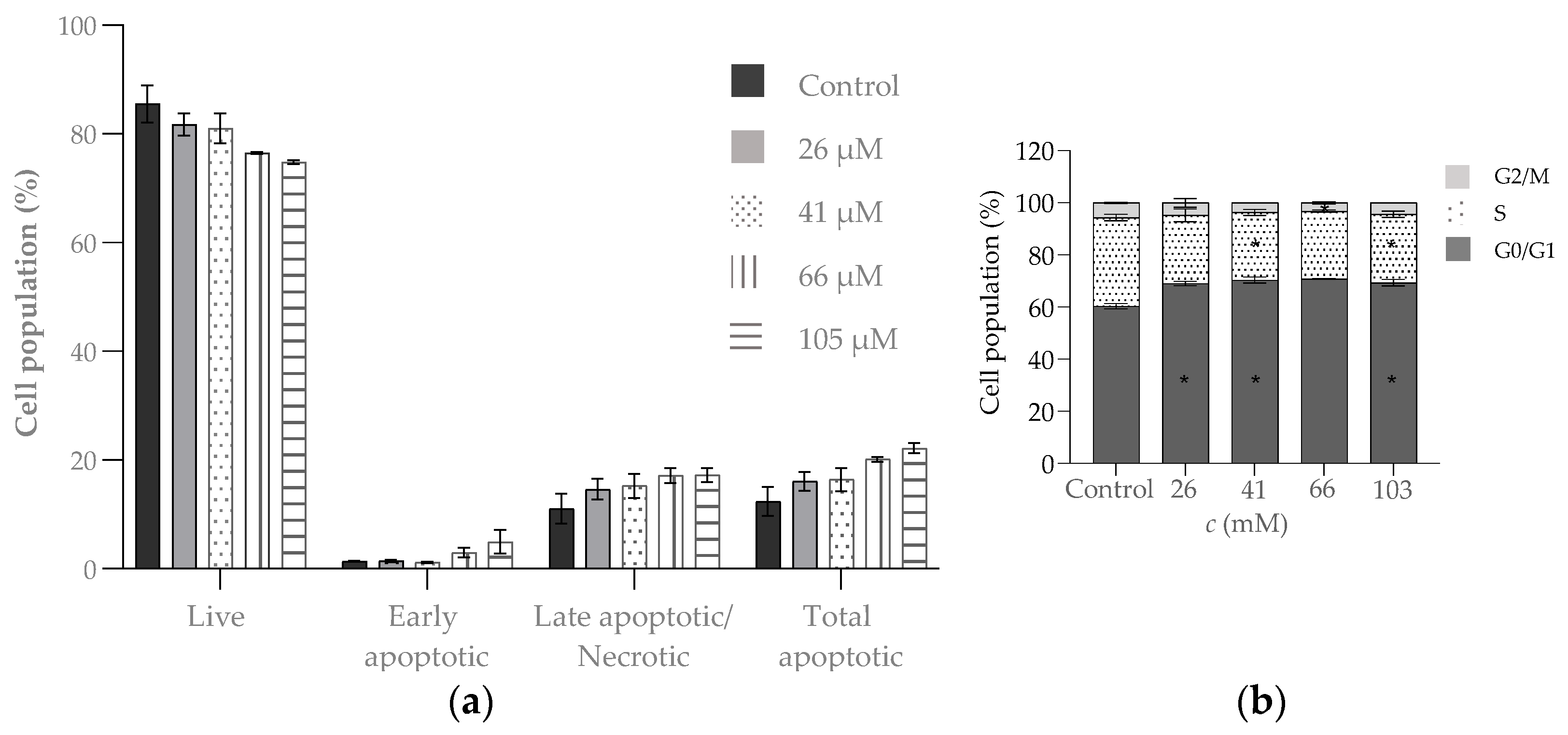
4.4. Biological Evaluation
- Antitumor activity
5. Conclusions and Future Perspective
- (i)
- The heterochirality of the peptide backbone contributes significantly to IHB-mediated folding into stable turn structures: the concentration-independent IR and concentration-, temperature- and solvent-independent NMR spectra of heterochiral conjugates Boc/Ac–d–Pro–l–Ala–NH–Fn–COOMe (1/2) indicate their participation in a strong intrastrand NHFn···O=CBoc/Ac HB corresponding to β-turn, and in an additional interstrand NHAla···O=CCOOMe HB forming a 9-membered ring. Their homochiral counterparts Boc/Ac–l–Pro–l–Ala–NH–Fn–COOMe (3/4) are involved in more flexible structures.
- (ii)
- Steric hindrance by the amino acid side chain significantly weakens conformational stability: the amide region of the IR spectra of the peptides Boc–AA–NH–Fn–COOMe (AA = d-Phe (11), d-Val (13), d-Leu (15)), Ac–AA–NH–Fn–COOMe (AA = d-Phe (12), d-Val (14), d-Leu (16)) is characterized by the dominance of the blue-shifted signals of the non-bonded NH groups, which is due to steric hindrance by the bulky and branched side chains of Phe, Val and Leu. In addition, the intensity ratios of the free and associated NH bands indicate that the bulky Boc-protecting groups in peptides 11, 13 and 15 hinder the involvement in hydrogen bonding more than the Ac-group in 12, 14 and 16. Accordingly, their chemical shifts showed a pronounced dependence on temperature and solvent, and only weak Cotton effects are observed in the CD spectra.
- (iii)
- The symmetrically disubstituted conjugates are involved in stronger IHBs and therefore adopt more stable conformations compared to asymmetrically monosubstituted peptides: while the asymmetrically monosubstituted conjugates 11–16 were found to be mostly non-involved in HBs, their higher symmetrically disubstituted homologues Ac–AA–NH–Fn–NH–AA–Boc (AA = l- and d-Phe (17/18), l- and d-Val (19/20), l- and d-Leu (21/22)) with additional hydrogen-bond-donor and acceptor sites were mainly involved in HBs. The most stable conformations were based on two 10-membered IHB rings, i.e., two β-turns connected by two hydrogen bonds between the peptide strands attached to the opposite cyclopentadienyl rings NHFn···OCBoc and NHFn···OCAc, respectively. The chemical shifts of the NH protons involved in HBs were temperature- and DMSO-independent. Moreover, the Cotton effects in their CD spectra were more than 20-fold stronger than those of peptides 11–16.
Author Contributions
Funding
Conflicts of Interest
References
- Kealy, T.J.; Pauson, P.L. New Type of Organo-Iron Compound. Nature 1951, 168, 1039–1040. [Google Scholar] [CrossRef]
- Miller, S.A.; Tebboth, J.A.; Tremaine, J.F. Dyclopentadienyliron. J. Chem. Soc. 1952, 632–635. [Google Scholar] [CrossRef]
- Available online: https://www.chemistryworld.com/podcasts/ferrocene/6134.article (accessed on 11 January 2024).
- Patra, M.; Gasser, G. The medicinal chemistry of ferrocene and its derivatives. Nat. Rev. Chem. 2017, 1, 0066. [Google Scholar] [CrossRef]
- Astruc, D. Why is Ferrocene so Exceptional? Eur. J. Inorg. Chem. 2017, 2017, 6–29. [Google Scholar] [CrossRef]
- Wang, R.; Chen, H.; Yan, W.; Zheng, M.; Zhang, T.; Zhang, Y. Ferrocene-containing hybrids as potential anticancer agents: Current developments, mechanisms of action and structure-activity relationships. Eur. J. Med. Chem. 2020, 190, 112109–112130. [Google Scholar] [CrossRef] [PubMed]
- Rauf, U.; Shabir, G.; Bukhari, S.; Albericio, F.; Saeed, A. Contemporary Developments in Ferrocene Chemistry: Physical, Chemical, Biological and Industrial Aspects. Molecules 2023, 28, 5765. [Google Scholar] [CrossRef] [PubMed]
- Ornelas, C.; Astruc, D. Ferrocene-Based Drugs, Delivery Nanomaterials and Fenton Mechanism: State of the Art, Recent Developments and Prospects. Pharmaceutics 2023, 15, 2044. [Google Scholar] [CrossRef] [PubMed]
- Čakić Semenčić, M.; Barišić, L. Ferrocene Bioconjugates. Croat. Chem. Acta 2017, 90, 537–569. [Google Scholar] [CrossRef]
- Schlotter, K.; Boeckler, F.; Hübner, H.; Gmeiner, P. Fancy Bioisosteres: Metallocene-Derived G-Protein-Coupled Receptor Ligands with Subnanomolar Binding Affinity and Novel Selectivity Profiles. J. Med. Chem. 2005, 48, 3696–3699. [Google Scholar] [CrossRef]
- Jaouen, G.; Vessières, A.; Top, S. Ferrocifen type anti cancer drugs. Chem. Soc. Rev. 2015, 44, 8802–8817. [Google Scholar] [CrossRef]
- Chavain, N.; Vezin, H.; Dive, D.; Touati, N.; Paul, J.F.; Buisine, E.; Biot, C. Investigation of the redox behavior of ferroquine, a new antimalarial. Mol. Pharm. 2008, 5, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Sethi, S.; Kumar Das, P.; Behera, N. The chemistry of aminoferrocene, Fe{(η5-C5H4NH2)(η5-Cp)}: Synthesis, reactivity and applications. J. Organomet. Chem. 2016, 824, 140–165. [Google Scholar] [CrossRef]
- Kumaravel, S.; Balamurugan, T.S.T.; Jia, S.-H.; Lin, H.-Y.; Huang, S.-T. Ratiometric electrochemical molecular switch for sensing hypochlorous acid: Applicable in food analysis and real-time in-situ monitoring. Anal. Chim. Acta 2020, 1106, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lu, J.; Hao, L.; Yang, H.; Song, X.; Si, F. Electrochemical detection of alkaline phosphatase activity through enzyme-catalyzed reaction using aminoferrocene as an electroactive probe. Anal. Bioanal. Chem. 2021, 413, 1827–1836. [Google Scholar] [CrossRef] [PubMed]
- Kumaravel, S.; Jian, S.-E.; Huang, S.-T.; Huang, C.-H.; Hong, W.-Z. Convenient and ultrasensitive detection of live Salmonella using ratiometric electrochemical molecular substrates. Anal. Chim. Acta 2022, 1190, 339244. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hong, Y.; Wang, M.; Zhu, Y. Multifunctional Nanolabels Based on Polydopamine Nanospheres for Sensitive Alpha Fetoprotein Electrochemical Detection. ACS Appl. Nano Mater. 2022, 5, 1588–1599. [Google Scholar] [CrossRef]
- Sariga; Varghese, A. The Renaissance of Ferrocene-Based Electrocatalysts: Properties, Synthesis Strategies, and Applications. Top. Curr. Chem. 2023, 381, 32–124. [Google Scholar] [CrossRef] [PubMed]
- Hagen, H.; Marzenell, P.; Jentzsch, E.; Wenz, F.; Veldwijk, M.R.; Mokhir, A. Aminoferrocene-based prodrugs activated by reactive oxygen species. J. Med. Chem. 2012, 55, 924–934. [Google Scholar] [CrossRef]
- Peiro Cadahia, J.; Previtali, V.; Troelsen, N.S.; Clausen, M.H. Prodrug strategies for targeted therapy triggered by reactive oxygen species. MedChemComm 2019, 10, 1531–1549. [Google Scholar] [CrossRef]
- Daum, S.; Toms, J.; Reshetnikov, V.; Gizem Özkan, H.; Hampel, F.; Maschauer, S.; Hakimioun, A.; Beierlein, F.; Sellner, L.; Schmitt, M.; et al. Identification of Boronic Acid Derivatives as an Active Form of N-Alkylaminoferrocene-Based Anticancer Prodrugs and Their Radiolabeling with 18F. Bioconjug. Chem. 2019, 30, 1077–1086. [Google Scholar] [CrossRef]
- Gizem Özkan, H.; Toms, J.; Maschauer, S.; Prante, O.; Mokhir, A. Aminoferrocene-Based Anticancer Prodrugs Labelled with Cyanine Dyes for in vivo Imaging. Eur. J. Inorg. Chem. 2021, 2021, 5096–5102. [Google Scholar] [CrossRef]
- Xu, H.G.; Schikora, M.; Sisa, M.; Daum, S.; Klemt, I.; Janko, C.; Alexiou, C.; Bila, G.; Bilyy, R.; Gong, W.; et al. An Endoplasmic Reticulum Specific Pro-amplifier of Reactive Oxygen Species in Cancer Cells. Angew. Chem. Int. Ed. 2021, 60, 11158–11162. [Google Scholar] [CrossRef]
- Gizem Özkan, H.; Thakor, V.; Xu, H.G.; Bila, G.; Bilyy, R.; Bida, D.; Böttcher, M.; Mougiakakos, D.; Tietze, R.; Mokhir, A. Anticancer Aminoferrocene Derivatives Inducing Production of Mitochondrial Reactive Oxygen Species. Chem. Eur. J. 2022, 28, e202104420. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tan, J.; Song, C.; Zhang, G.; Hu, X.; Liu, S. Self-Immolative Amphiphilic Poly(ferrocenes) for Synergistic Amplification of Oxidative Stress in Tumor Therapy. Angew. Chem. Int. Ed. 2023, 62, e202303829. [Google Scholar] [CrossRef] [PubMed]
- Wilde, M.; Arzur, D.; Baratte, B.; Lefebvre, D.; Robert, T.; Roisnel, T.; Le Jossic-Corcos, C.; Bach, S.; Corcos, L.; Erb, W. Regorafenib analogues and their ferrocenic counterparts: Synthesis and biological evaluation. New J. Chem. 2020, 44, 19723–19733. [Google Scholar] [CrossRef]
- Čakić Semenčić, M.; Kodrin, I.; Barišić, L.; Nuskol, M.; Meden, A. Synthesis and Conformational Study of Monosubstituted Aminoferrocene-Based Peptides Bearing Homo- and Heterochiral Pro-Ala Sequences. Eur. J. Inorg. Chem. 2017, 2017, 306–307. [Google Scholar] [CrossRef]
- Kovačević, M.; Kodrin, I.; Cetina, M.; Kmetič, I.; Murati, T.; Čakić Semenčić, M.; Roca, S.; Barišić, L. The conjugates of ferrocene-1,1′-diamine and amino acids. A novel synthetic approach and conformational analysis. Dalton Trans. 2015, 44, 16405–16420. [Google Scholar] [CrossRef] [PubMed]
- Kovačević, M.; Kodrin, I.; Roca, S.; Molčanov, K.; Shen, Y.; Adhikari, B.; Kraatz, H.-B.; Barišić, L. Helically Chiral Peptides That Contain Ferrocene-1,1′-diamine Scaffolds as a Turn Inducer. Chem.—Eur. J. 2017, 23, 10372–10395. [Google Scholar] [CrossRef] [PubMed]
- Kovač, V.; Radošević, K.; Bebek, A.; Makarević, J.; Štefanić, Z.; Barišić, L.; Žinić, M.; Rapić, V. The first oxalamide-bridged ferrocene: Facile synthesis, preliminary conformational analysis and biological evaluation. Appl. Organometal. Chem. 2017, 31, e3653. [Google Scholar] [CrossRef]
- Tomasini, C.; Castellucci, N. Peptides and peptidomimetics that behave as low molecular weight gelators. Chem. Soc. Rev. 2013, 42, 156–172. [Google Scholar] [CrossRef]
- Kovačević, M.; Čakić Semenčić, M.; Radošević, K.; Molčanov, K.; Roca, S.; Šimunović, L.; Kodrin, I.; Barišić, L. Conformational Preferences and Antiproliferative Activity of Peptidomimetics Containing Methyl 1′-Aminoferrocene-1-carboxylate and Turn-Forming Homo- and Heterochiral Pro-Ala Motifs. Int. J. Mol. Sci. 2021, 22, 13532. [Google Scholar] [CrossRef] [PubMed]
- Kovačević, M.; Čakić Semenčić, M.; Kodrin, I.; Roca, S.; Perica, J.; Mrvčić, J.; Stanzer, D.; Molčanov, K.; Milašinović, V.; Brkljačić, L.; et al. Biological Evaluation and Conformational Preferences of Ferrocene Dipeptides with Hydrophobic Amino Acids. Inorganics 2023, 11, 29. [Google Scholar] [CrossRef]
- Kovačević, M.; Markulin, D.; Zelenika, M.; Marjanović, M.; Lovrić, M.; Polančec, D.; Ivančić, M.; Mrvčić, J.; Molčanov, K.; Milašinović, V.; et al. Hydrogen Bonding Drives Helical Chirality via 10-Membered Rings in Dipeptide Conjugates of Ferrocene-1,1′-Diamine. Int. J. Mol. Sci. 2022, 23, 12233. [Google Scholar] [CrossRef]
- Nuskol, M.; Šutalo, P.; Đaković, M.; Kovačević, M.; Kodrin, I.; Čakić Semenčić, M. Testing the Potential of the Ferrocene Chromophore as a Circular Dichroism Probe for the Assignment of the Screw-Sense Preference of Tripeptides. Organometallics 2021, 40, 1351–1362. [Google Scholar] [CrossRef]
- Nuskol, M.; Šutalo, P.; Kodrin, I.; Čakić Semenčić, M. Sensing of the Induced Helical Chirality by the Chiroptical Response of the Ferrocene Chromophore. Eur. J. Inorg. Chem. 2022, 2022, e202100880. [Google Scholar] [CrossRef]
- Nuskol, M.; Šutalo, P.; Kovačević, M.; Kodrin, I.; Čakić Semenčić, M. Central-to-Helical-to-Axial Chirality Transfer in Chiroptical Sensing with Ferrocene Chromophore. Inorganics 2023, 11, 225. [Google Scholar] [CrossRef]
- Nuskol, M.; Studen, B.; Meden, A.; Kodrin, I.; Čakić Semenčić, M. Tight turn in dipeptide bridged ferrocenes: Synthesis, X-ray structural, theoretical and spectroscopic studies. Polyhedron 2019, 161, 137–144. [Google Scholar] [CrossRef]
- Kovač, V.; Kodrin, I.; Radošević, K.; Molčanov, K.; Adhikari, B.; Kraatz, H.-B.; Barišić, L. Oxalamide-Bridged Ferrocenes: Conformational and Gelation Properties and In Vitro Antitumor Activity. Organometallics 2022, 41, 920–936. [Google Scholar] [CrossRef]
- Barišić, L.; Rapić, V.; Kovač, V. Ferrocene Compounds. XXIX. Efficient Syntheses of 1′-Aminoferrocene-1-carboxylic Acid Derivatives. Croat. Chem. Acta 2002, 75, 199–210. [Google Scholar]
- Barišić, L.; Čakić, M.; Mahmoud, K.A.; Liu, Y.-N.; Kraatz, H.-B.; Pritzkow, H.; Kirin, S.I.; Metzler-Nolte, N.; Rapić, V. Helically Chiral Ferrocene Peptides Containing 1′-Aminoferrocene-1-Carboxylic Acid Subunits as Turn Inducers. Chem. Eur. J. 2006, 12, 4965–4980. [Google Scholar] [CrossRef]
- Wang, X.; Ni, D.; Liu, Y.; Lu, S. Rational Design of Peptide-Based Inhibitors Disrupting Protein-Protein Interactions. Front Chem. 2021, 9, 682675. [Google Scholar] [CrossRef] [PubMed]
- Seychell, B.C.; Beck, T. Molecular basis for protein-protein interactions. Beilstein J. Org. Chem. 2021, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Zhou, Q.; He, J.; Jiang, Z.; Peng, C.; Tong, R.; Shi, J. Recent advances in the development of protein-protein interactions modulators: Mechanisms and clinical trials. Signal Transduct. Target Ther. 2020, 5, 213–235. [Google Scholar] [CrossRef] [PubMed]
- Ovek, D.; Abali, Z.; Zeylan, M.E.; Keskin, O.; Gursoy, A.; Tuncbag, N. Artificial intelligence based methods for hot spot prediction. Curr. Opin. Struct. Biol. 2022, 72, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.N.; Hill, T.A.; Ruiz-Gómez, G.; Diness, F.; Mason, J.M.; Wu, C.; Abbenante, G.; Shepherd, N.E.; Fairlie, D.P. Twists or turns: Stabilising alpha vs. beta turns in tetrapeptides. Chem. Sci. 2019, 10, 10595–10600. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Jung, J.; Tung, T.T.; Park, S.B. β-Turn mimetic-based stabilizers of protein-protein interactions for the study of the non-canonical roles of leucyl-tRNA synthetase. Chem. Sci. 2016, 7, 2753–2761. [Google Scholar] [CrossRef]
- Laxio Arenas, J.; Kaffy, J.; Ongeri, S. Peptides and peptidomimetics as inhibitors of protein-protein interactions involving β-sheet secondary structures. Curr. Opin. Chem. Biol. 2019, 52, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Vagner, J.; Qu, H.; Hruby, V.J. Peptidomimetics, a synthetic tool of drug discovery. Curr. Opin. Chem. Biol. 2008, 12, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Lenci, E.; Trabocchi, A. Peptidomimetic toolbox for drug discovery. Chem. Soc. Rev. 2020, 49, 3262–3277. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Kundu, R. Cationic Amphiphilic Peptides: Synthetic Antimicrobial Agents Inspired by Nature. ChemMedChem 2020, 15, 1887–1896. [Google Scholar] [CrossRef]
- Liscano, Y.; Oñate-Garzón, J.; Delgado, J.P. Peptides with Dual Antimicrobial–Anticancer Activity: Strategies to Overcome Peptide Limitations and Rational Design of Anticancer Peptides. Molecules 2020, 25, 4245. [Google Scholar] [CrossRef]
- Giannis, A.; Rübsam, F. Peptidomimetics in Drug Design. In Advances in Drug Research, 1st ed.; Testa, B., Meyer, U.A., Eds.; Academic Press: Cambridge, MA, USA, 1997; Volume 29, pp. 1–78. [Google Scholar] [CrossRef]
- Nair, R.V.; Baravkar, S.B.; Ingole, T.S.; Sanjayan, G.J. Synthetic turn mimetics and hairpin nucleators. Chem. Commun. 2014, 50, 13874–13884. [Google Scholar] [CrossRef]
- Barišić, L.; Dropučić, M.; Rapić, V.; Pritzkow, H.; Kirin, S.I.; Metzler-Nolte, N. The first oligopeptide derivative of 1′-aminoferrocene-1-carboxylic acid shows helical chirality with antiparallel strands. Chem. Commun. 2004, 17, 2004–2005. [Google Scholar] [CrossRef] [PubMed]
- Moriuchi, T.; Ohmura, S.D.; Moriuchi-Kawakami, T. Chirality Induction in Bioorganometallic Conjugates. Inorganics 2018, 6, 111. [Google Scholar] [CrossRef]
- Byun, B.J.; Song, I.K.; Chung, Y.J.; Ryu, K.H.; Kang, Y.K. Conformational preferences of X-Pro sequences: Ala-Pro and Aib-Pro motifs. J. Phys. Chem. B 2010, 114, 14077–14086. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Legrand, B.; Vezenkov, L.L.; Berthet, M.; Subra, G.; Calmès, M.; Bantignies, J.-L.; Martinez, J.; Amblard, M. Turning Peptide Sequences into Ribbon Foldamers by a Straightforward Multicyclization Reaction. Angew. Chem. Int. Ed. 2015, 54, 13966–13970. [Google Scholar] [CrossRef]
- Metrano, A.J.; Abascal, N.C.; Mercado, B.Q.; Paulson, E.K.; Hurtley, A.E.; Miller, S.J. Diversity of Secondary Structure in Catalytic Peptides with β-Turn-Biased Sequences. J. Am. Chem. Soc. 2017, 139, 492–516. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, E. MacroModel; Version 10.3; Schrödinger: New York, NY, USA, 2014. [Google Scholar]
- Mohamadi, F.; Richards, N.G.J.; Guida, W.C.; Liskamp, R.; Lipton, M.; Caufield, C.; Chang, G.; Hendrickson, T.; Still, W.C. Macromodel—An integrated software system for modeling organic and bioorganic molecules using molecular mechanics. J. Comput. Chem. 1990, 11, 440–467. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16, Revision, C.01; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Marenich, A.V.; Cramer, C.J.; Truhlar, D.G. Universal Solvation Model Based on Solute Electron Density and on a Continuum Model of the Solvent Defined by the Bulk Dielectric Constant and Atomic Surface Tensions. J. Phys. Chem. B 2009, 113, 6378–6396. [Google Scholar] [CrossRef]
- Koch, U.; Popelier, P.L.A. Characterization of C-H-O Hydrogen Bonds on the Basis of the Charge Density. J. Phys. Chem. 1995, 99, 9747–9754. [Google Scholar] [CrossRef]
- Popelier, P.L.A. Characterization of a dihydrogen bond on the basis of the electron density. J. Phys. Chem. A 1998, 102, 1873–1878. [Google Scholar] [CrossRef]
- Caramiello, A.M.; Bellucci, M.C.; Cristina, G.; Castellano, C.; Meneghetti, F.; Mori, M.; Secundo, F.; Viani, F.; Sacchetti, A.; Volonterio, A. Synthesis and Conformational Analysis of Hydantoin-Based Universal Peptidomimetics. J. Org. Chem. 2023, 88, 10381–10402. [Google Scholar] [CrossRef]
- Kong, J.; Yu, S. Fourier transform infrared spectroscopic analysis of protein secondary structures. Acta Biochim. Biophys. Sin. 2007, 39, 549–559. [Google Scholar] [CrossRef]
- Vincenzi, M.; Mercurio, F.A.; Leone, M. NMR Spectroscopy in the Conformational Analysis of Peptides: An Overview. Curr. Med. Chem. 2021, 28, 2729–2782. [Google Scholar] [CrossRef]
- Vass, E.; Hollósi, M.; Besson, F.; Buchet, R. Vibrational spectroscopic detection of beta- and gamma-turns in synthetic and natural peptides and proteins. Chem. Rev. 2003, 103, 1917–1954. [Google Scholar] [CrossRef]
- Ramos, S.; Thielges, M.C. Site-Specific 1D and 2D IR Spectroscopy to Characterize the Conformations and Dynamics of Protein Molecular Recognition. J. Phys. Chem B. 2019, 123, 3551–3566. [Google Scholar] [CrossRef]
- Arunan, E.; Desiraju, G.R.; Klein, R.A.; Sadlej, J.; Scheiner, S.; Alkorta, I.; Clary, D.C.; Crabtree, R.H.; Dannenberg, J.J.; Hobza, P.; et al. Defining the hydrogen bond: An account (IUPAC Technical Report). Pure Appl. Chem. 2011, 83, 1619–1636. [Google Scholar] [CrossRef]
- Fornaro, T.; Burini, D.; Biczysko, M.; Barone, V. Hydrogen-bonding effects on infrared spectra from anharmonic computations: Uracil-water complexes and uracil dimers. J. Phys. Chem. A 2015, 119, 4224–4236. [Google Scholar] [CrossRef]
- Wagner, G.; Pardi, A.; Wuethrich, K. Hydrogen bond length and proton NMR chemical shifts in proteins. J. Am. Chem. Soc. 1983, 105, 5948–5949. [Google Scholar] [CrossRef]
- Newberry, R.W.; Raines, R.T. Secondary Forces in Protein Folding. ACS Chem. Biol. 2019, 14, 1677–1686. [Google Scholar] [CrossRef]
- Llinás, M.; Klein, M.P. Solution conformation of the ferrichromes. VI. Charge relay at the peptide bond. Proton magnetic resonance study of solvation effects on the amide electron density distribution. J. Am. Chem. Soc. 1975, 97, 4731–4737. [Google Scholar] [CrossRef]
- Stevens, E.S.; Sugawara, N.; Bonora, G.M.; Toniolo, C. Conformational analysis of linear peptides. 3. Temperature dependence of NH chemical shifts in chloroform. J. Am. Chem. Soc. 1980, 102, 7048–7050. [Google Scholar] [CrossRef]
- Kessler, H. Conformation and Biological Activity of cyclic Peptides. Angew. Chem. Int. Ed. 1982, 21, 512–523. [Google Scholar] [CrossRef]
- Iqbal, M.; Balaram, P. Aggregation of apolar peptides in organic solvents. Concentration dependence of 1H-nmr parameters for peptide NH groups in 310 helical decapeptide fragment of suzukacillin. Biopolymers 1982, 21, 1427–1433. [Google Scholar] [CrossRef]
- Vijayakumar, E.K.S.; Balaram, P. Stereochemistry of a-aminoisobutyric acid peptides in solution. Helical conformations of protected decapeptides with repeating Aib-L-Ala and Aib-L-Val sequences. Biopolymers 1983, 22, 2133–2140. [Google Scholar] [CrossRef]
- Andersen, N.H.; Neidigh, J.W.; Harris, S.M.; Lee, G.M.; Liu, Z.; Tong, H. Extracting Information from the Temperature Gradients of Polypeptide NH Chemical Shifts. 1. The Importance of Conformational Averaging. J. Am. Chem. Soc. 1997, 119, 8547–8561. [Google Scholar] [CrossRef]
- Baxter, N.J.; Williamson, M.P. Temperature dependence of 1H chemical shifts in proteins. J. Biomol. NMR 1997, 9, 359–369. [Google Scholar] [CrossRef]
- Cierpicki, T.; Otlewski, J. Amide proton temperature coefficients as hydrogen bond indicators in proteins. J. Biomol. NMR 2001, 21, 249–261. [Google Scholar] [CrossRef]
- Cierpicki, T.; Zhukov, I.; Byrd, R.A.; Otlewski, J. Hydrogen bonds in human ubiquitin reflected in temperature coefficients of amide protons. J. Magn. Reson. 2002, 157, 178–180. [Google Scholar] [CrossRef]
- Pignataro, M.F.; Herrera, M.G.; Dodero, V.I. Evaluation of Peptide/Protein Self-Assembly and Aggregation by Spectroscopic Methods. Molecules 2020, 25, 4854. [Google Scholar] [CrossRef]
- Rogers, D.M.; Jasim, S.B.; Dyer, N.T.; Auvray, F.; Réfrégiers, M.; Hirst, J.D. Electronic Circular Dichroism Spectroscopy of Proteins. Chem 2019, 5, 2751–2774. [Google Scholar] [CrossRef]
- Moriuchi, T. Helical Chirality of Ferrocene Moieties in Cyclic Ferrocene-Peptide Conjugates. Chem. Eur. J. 2022, 2022, e202100902. [Google Scholar] [CrossRef]
- Barth, N.D.; Subiros-Funosas, R.; Mendive-Tapia, L.; Duffin, R.; Shields, M.A.; Cartwright, J.A.; Henriques, S.T.; Sot, J.; Goñi, F.M.; Lavilla, R.; et al. A fluorogenic cyclic peptide for imaging and quantification of drug-induced apoptosis. Nat. Commun. 2020, 11, 4027. [Google Scholar] [CrossRef]
- Li, L.; Thomas, R.M.M.; Suzuki, H.; de Brabander, J.K.; Wang, X.; Harran, P.G. A small molecule Smac mimic potentiates TRAIL- and TNFalpha-mediated cell death. Science 2004, 305, 1471–1474. [Google Scholar] [CrossRef]
- Lyu, Z.; Yang, P.; Lei, J.; Zhao, J. Biological Function of Antimicrobial Peptides on Suppressing Pathogens and Improving Host Immunity. Antibiotics 2023, 12, 1037. [Google Scholar] [CrossRef]
- Iqbal, M.; Balaram, P. The 310 helical conformation of the amino terminal decapeptide of suzukacillin. 270 MHz hydrogen-1 NMR evidence for eight intramolecular hydrogen bonds. J. Am. Chem. Soc. 1981, 103, 5548–5552. [Google Scholar] [CrossRef]
- Venkatachalapathi, Y.V.; Prasad, B.V.V.; Balaram, P. Conformational analysis of small disulfide loops. Spectroscopic and theoretical studies on a synthetic cyclic tetrapeptide containing cystine. Biochemistry 1982, 2, 5502–5509. [Google Scholar] [CrossRef]
- Albada, B.; Metzler-Nolte, N. Highly Potent Antibacterial Organometallic Peptide Conjugates. Acc. Chem. Res. 2017, 50, 2510–2518. [Google Scholar] [CrossRef] [PubMed]
- Soulère, L.; Bernard, J. Design, solid phase synthesis and evaluation of cationic ferrocenoyl peptide bioconjugates as potential antioxidant enzyme mimics. Bioorganic Med. Chem. Lett. 2009, 19, 1173–1176. [Google Scholar] [CrossRef]
- Xianjiao, M.; Shengling, L.; Wenbing, M.; Jianlong, W.; Zhiyong, H.; Duanlin, C. Synthesis and Antioxidant Activities of Ferrocenyl-containing Curcumin Analogues. Lett. Drug Des. Discov. 2018, 15, 1252–1258. [Google Scholar] [CrossRef]
- Kataoka, H. Sample preparation for liquid chromatography. In Liquid chromatography: Applications, 2nd ed.; Fanali, S., Haddad, P., Poole, C., Riekkola, M.-L., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–37. [Google Scholar] [CrossRef]
- Fekete, S.; Veuthey, J.L.; Guillarme, D. New trends in reversed-phase liquid chromatographic separations of therapeutic peptides and proteins: Theory and applications. J. Pharm. Biomed. Anal. 2012, 69, 9–27. [Google Scholar] [CrossRef]
- Neuhaus, C.S.; Gabernet, G.; Steuer, C.; Hiss, J.A.; Zenobi, R.; Schneider, G. Simulated Molecular Evolution for Anticancer Peptide Design. Angew. Chem. Int. Ed. 2019, 58, 1674–1678. [Google Scholar] [CrossRef]
- Shibue, M.; Mant, C.T.; Hodges, R.S. Effect of anionic ion-pairing reagent hydrophobicity on selectivity of peptide separations by reversed-phase liquid chromatography. J. Chromatogr. A 2005, 1080, 68–75. [Google Scholar] [CrossRef]
- Boyd, M.R.; Kenneth, D.P. Some practical considerations and applications of the national cancer institute in vitro anticancer drug discovery screen. Drug Dev. Res. 1995, 34, 91–109. [Google Scholar] [CrossRef]
- Pascal, F.; Bedouet, L.; Baylatry, M.; Namur, J.; Laurent, A. Comparative Chemosensitivity of VX2 and HCC Cell Lines to Drugs Used in TACE. Anticancer Res. 2015, 35, 6497–6503. [Google Scholar]
- Kraatz, H.-B.; Leek, D.M.; Houmam, A.; Enright, G.D.; Lusztyk, J.; Wayner, D.D.M. The ferrocene moiety as a structural probe: Redox and structural properties of ferrocenoyl-oligoprolines Fc-Pron-OBzl (n = 1–4) and Fc-Pro2-Phe-Obzl. J. Organomet. Chem. 1999, 589, 38–49. [Google Scholar] [CrossRef]
- Moriuchi, T.; Fujiwara, T.; Hirao, T. β-Turn-structure-assembled palladium complexes by complexation-induced self-organization of ferrocene–dipeptide conjugates. Dalton Trans. 2009, 22, 4286–4288. [Google Scholar] [CrossRef]
- Zong, Z.; Zhang, H.; Hao, A.; Xing, P. The origin of supramolecular chirality in 1-ferrocenyl amino acids. Dalton Trans. 2021, 50, 9695–9699. [Google Scholar] [CrossRef]
- Kovač, V.; Čakić Semenčić, M.; Kodrin, I.; Roca, S.; Rapić, V. Ferrocene-dipeptide conjugates derived from aminoferrocene and 1-acetyl-1′-aminoferrocene: Synthesis and conformational studies. Tetrahedron 2013, 69, 10497–10506. [Google Scholar] [CrossRef]
- Toniolo, C.; Formaggio, F.; Crisma, M.; Schoemaker, H.E.; Kamphuis, J. The p-bromobenzamido chromophore as a circular dichroic probe for the assignment of the screw sense of helical peptides. Tetrahedron Asymmetry 1994, 5, 507–510. [Google Scholar] [CrossRef]
- Diemer, V.; Maury, J.; Le Bailly, B.A.F.; Webb, S.J.; Clayden, J. Dibenzazepinyl ureas as dual NMR and CD probes of helical screw-sense preference in conformationally equilibrating dynamic foldamers. Chem. Commun. 2017, 53, 10768–10771. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.V.; Balaram, P. The stereochemistry of peptides containing alpha-aminoisobutyric acid. CRC Crit. Rev. Biochem. 1984, 16, 307–348. [Google Scholar] [CrossRef] [PubMed]
- Wüthrich, K. NMR of Proteins and Nucleic Acids; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Gellman, S.H.; Dado, G.P.; Liang, G.-B.; Adams, B.R. Conformation-directing effects of a single intramolecular amide-amide hydrogen bond: Variable-temperature NMR and IR studies on a homologous diamide series. J. Am. Chem. Soc. 1991, 113, 1164–1173. [Google Scholar] [CrossRef]
- Pardi, A.; Wagner, G.; Wuthrich, K. Protein conformation and proton nuclear-magnetic-resonance chemical shifts. Eur. J. Biochem. 1983, 137, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Omar, M.M.; Abd El-Halim, H.F.; Khalil, E.A.M. Synthesis, characterization, and biological and anticancer studies of mixed ligand complexes with Schiff base and 2,2′-bipyridine. Appl. Organomet. Chem. 2017, 31, e3724. [Google Scholar] [CrossRef]
- Steeneck, C.; Kinzel, O.; Anderhub, S.; Hornberger, M.; Pinto, S.; Morschhaeuser, B.; Albers, M.; Sonnek, C.; Czekańska, M.; Hoffmann, T. Discovery and optimization of substituted oxalamides as novel heme-displacing IDO1 inhibitors. Bioorg. Med. Chem. Lett. 2021, 33, 127744. [Google Scholar] [CrossRef]
- Saito, F.; Schreiner, P.R. Determination of the Absolute Configurations of Chiral Alkanes-An Analysis of the Available Tools. Eur. J. Org. Chem. 2020, 2020, 6328–6339. [Google Scholar] [CrossRef]
- Wolf, C.; Bentleya, K.W. Chirality sensing using stereodynamic probes with distinct electronic circular dichroism output. Chem. Soc. Rev. 2013, 42, 5408–5424. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | NHfree (ῠmax/cm−1) | NHassoc. (ῠmax/cm−1) | INHfree/INHassoc. |
|---|---|---|---|
| Boc–d–Pro–l–Ala–NH–Fn–COOMe (1) | 3418 | 3325 | 1:2.4 |
| Ac–d–Pro–l–Ala–NH–Fn–COOMe (2) | 3424 | 3303 | 1:2.3 |
| Boc–l–Pro–l–Ala–NH–Fn–COOMe (3) | 3418 | 3310 | 1:1.4 |
| Ac–l–Pro–l–Ala–NH–Fn–COOMe (4) | 3420 | 3309 | 1:1.6 |
| Compound | NHFn | NHAla | NHFn | NHAla | NHFn | NHAla |
|---|---|---|---|---|---|---|
| δ (ppm) | Δδ (ppm) | Δδ/ΔT (ppb K–1) | ||||
| Boc–d–Pro–l–Ala–NH–Fn–COOMe (1) | 8.37 | 7.24 | 0.45 | 0.94 | −7.42 | −6.14 |
| Ac–d–Pro–l–Ala–NH–Fn–COOMe (2) | 8.58 | 7.08 | 0.3 | 1.46 | −5.42 | −3.85 |
| Boc–l–Pro–l–Ala–NH–Fn–COOMe (3) | 8.15 | 6.85 | 0.9 | 1.12 | −8.57 | −0.08 |
| Ac–l–Pro–l–Ala–NH–Fn–COOMe (4) | 7.99 | 7.22 | 0.98 | 0.83 | −6.14 | −4.14 |
| Compound | Mθ (deg cm2 dmol−1) |
|---|---|
| Boc–d–Pro–l–Ala–NH–Fn–COOMe (1) | +4230 |
| Ac–d–Pro–l–Ala–NH–Fn–COOMe (2) | +4766 |
| Boc–l–Pro–l–Ala–NH–Fn–COOMe (3) | −2481 |
| Ac–l–Pro–l–Ala–NH–Fn–COOMe (4) | +205 |
| IC50 (μM) | ||
|---|---|---|
| Compound | HeLa Cells | MCF-7 Cells |
| Boc–d–Pro–l–Ala–NH–Fn–COOMe (1) | 436.1959 | n.d. 1 |
| Ac–d–Pro–l–Ala–NH–Fn–COOMe (2) | n.d. 1 | n.d. 1 |
| Boc–l–Pro–l–Ala–NH–Fn–COOMe (3) | 370.3969 | 270.6925 |
| Ac–l–Pro–l–Ala–NH–Fn–COOMe (4) | n.d. 1 | n.d. 1 |
| Compound | NHfree (ῠmax/cm−1) | NHassoc. (ῠmax/cm−1) | INHfree/INHassoc. |
|---|---|---|---|
| Boc–d–Phe–NH–Fn–COOMe (11) | 3412 | 3336 | 4.4:1 |
| Ac–d–Phe–NH–Fn–COOMe (12) | 3417 | 3295 | 1.5:1 |
| Boc–d–Val–NH–Fn–COOMe (13) | 3421 | 3328 | 3:1 |
| Ac–d–Val–NH–Fn–COOMe (14) | 3420 | 3289 | 1.4:1 |
| Boc–d–Leu–NH–Fn–COOMe (15) | 3422 | 3333 | 2.8:1 |
| Ac–d–Leu–NH–Fn–COOMe (16) | 3423 | 3289 | 1.4:1 |
| Compound | NHFn | NHAA | NHFn | NHAA | NHFn | NHAA |
|---|---|---|---|---|---|---|
| δ (ppm) | Δδ (ppm) | Δδ/ΔT (ppb K–1) | ||||
| Boc–d–Phe–NH–Fn–COOMe (11) | 7.32 | 5.15 | ||||
| Ac–d–Phe–NH–Fn–COOMe (12) | 7.73 | 6.44 | 1.72 | 1.54 | −14.14 | −9.14 |
| Boc–d–Val–NH–Fn–COOMe (13) | 7.38 | 5.15 | ||||
| Ac–d–Val–NH–Fn–COOMe (14) | 8.19 | 6.46 | 1.19 | 0.9 | −16.86 | −7.86 |
| Boc–d–Leu–NH–Fn–COOMe (15) | 7.62 | 5.03 | ||||
| Ac–d–Leu–NH–Fn–COOMe (16) | 8.15 | 6.38 | 1.23 | 1.37 | −15.43 | −11.28 |
| Compound | Mθ (deg cm2 dmol−1) |
|---|---|
| Boc–l–Phe–NH–Fn–COOMe (5) | 679 |
| Boc–d–Phe–NH–Fn–COOMe (11) | −679 |
| Ac–l–Phe–NH–Fn–COOMe (6) | 672 |
| Ac–d–Phe–NH–Fn–COOMe (12) | −667 |
| Boc–l–Val–NH–Fn–COOMe (7) | 1228 |
| Boc–d–Val–NH–Fn–COOMe (13) | −1216 |
| Ac–l–Val–NH–Fn–COOMe (8) | 675 |
| Ac–d–Val–NH–Fn–COOMe (14) | −687 |
| Boc–l–Leu–NH–Fn–COOMe (9) | 960 |
| Boc–d–Leu–NH–Fn–COOMe (15) | −957 |
| Ac–l–Leu–NH–Fn–COOMe (10) | 456 |
| Ac–d–Leu–NH–Fn–COOMe (16) | −469 |
| Compound | Test Microorganisms | Molarity of Solutions for Disk Diffusion Method (mM) | Inhibition Zone (mm) |
|---|---|---|---|
| Ac–d–Phe–NH–Fn–COOMe (12) | S. aureus B. subtilis P. aeruginosa | 224 | 14 ± 1 |
| 14 ± 1 | |||
| 14 ± 1 | |||
| Ac–d–Val–NH–Fn–COOMe (14) | S. aureus B. subtilis P. aeruginosa | 251 | n.d. 1 |
| 7 ± 1 | |||
| n.d. 1 | |||
| Ac–d–Leu–NH–Fn–COOMe (16) | S. aureus B. subtilis P. aeruginosa | 243 | 16 ± 1 |
| 16 ± 1 | |||
| 19 ± 2 | |||
| Kanamycin | S. aureus B. subtilis P. aeruginosa | 50 µg disk | 25 ± 0 |
| 25 ± 0 | |||
| 28 ± 1 |
| Compound (1 mM) | DPPH (% Inhibition) | DPPH (mM Trolox Equivalent) | FRAP (mM Trolox) |
|---|---|---|---|
| Boc–l–Phe–NH–Fn–COOMe (5) | 5.50 ± 3.87 | 0.067 | 1.62 ± 0.001 |
| Boc–d–Phe–NH–Fn–COOMe (11) | 3.99 ± 0.48 | 0.051 | 2.00 ± 0.001 |
| Ac–l–Phe–NH–Fn–COOMe (6) | 3.65 ± 2.26 | 0.046 | 1.09 ± 0.030 |
| Ac–d–Phe–NH–Fn–COOMe (12) | 5.25 ± 2.26 | 0.067 | 2.22 ± 0.006 |
| Boc–l–Val–NH–Fn–COOMe (7) | 11.75 ± 2.10 | 0.150 | 2.16 ± 0.013 |
| Boc–d–Val–NH–Fn–COOMe (13) | 7.76 ± 0.65 | 0.099 | 2.17 ± 0.015 |
| Ac–l–Val–NH–Fn–COOMe (8) | 4.57 ± 0.64 | 0.058 | 0.58 ± 0.009 |
| Ac–d–Val–NH–Fn–COOMe (14) | 10.62 ± 1.13 | 0.135 | 2.29 ± 0.003 |
| Boc–l–Leu–NH–Fn–COOMe (9) | 16.55 ± 1.77 | 0.211 | 2.27 ± 0.006 |
| Boc–d–Leu–NH–Fn–COOMe (15) | 8.79 ± 2.10 | 0.112 | 2.21 ± 0.003 |
| Ac–l–Leu–NH–Fn–COOMe (10) | 3.19 ± 1.29 | 0.041 | 0.72 ± 0.004 |
| Ac–d–Leu–NH–Fn–COOMe (16) | 6.96 ± 0.81 | 0.089 | 1.77 ± 0.001 |
| Standard (0.5 mM Trolox) | 39.27 ± 8.40 | 0.500 |
| Compound | tR (min) | % ACN |
|---|---|---|
| Boc–l–Phe–NH–Fn–COOMe (5) | 8.3 | 87.9 |
| Boc–d–Phe–NH–Fn–COOMe (11) | 8.3 | 87.9 |
| Ac–l–Phe–NH–Fn–COOMe (6) | 3.6 | 68.0 |
| Ac–d–Phe–NH–Fn–COOMe (12) | 3.6 | 68.0 |
| Boc–l–Val–NH–Fn–COOMe (7) | 7.1 | 82.6 |
| Boc–d–Val–NH–Fn–COOMe (13) | 7.1 | 82.6 |
| Ac–l–Val–NH–Fn–COOMe (8) | 2.6 | 63.7 |
| Ac–D–Val–NH–Fn–COOMe (14) | 2.6 | 63.7 |
| Boc–l–Leu–NH–Fn–COOMe (9) | 8.3 | 87.5 |
| Boc–d–Leu–NH–Fn–COOMe (15) | 8.3 | 87.5 |
| Ac–l–Leu–NH–Fn–COOMe (10) | 3.3 | 66.6 |
| Ac–d–Leu–NH–Fn–COOMe (16) | 3.3 | 66.6 |
| Compound | NHfree (ῠmax/cm−1) | NHassoc. (ῠmax/cm−1) | INHfree/INHassoc. |
|---|---|---|---|
| Ac–l–Phe–NH–Fn–NH–l–Phe–Boc (17) | 3430 | 3302, 3266, 3217 | 0.7:1 |
| Ac–l–Val–NH–Fn–NH–l–Val–Boc (19) | 3434 | 3305, 3249 | 0.5:1 |
| Ac–l–Leu–NH–Fn–NH–l–Leu–Boc (21) | 3434 | 3301, 3253 | 0.5:1 |
| Compound | NHFna | NHFnb | NHAc | NHBoc |
|---|---|---|---|---|
| δ (ppm) | ||||
| Ac–l–Phe–NHa–Fn–NHb–l–Phe–Boc (17) | 9.15 | 9.21 | 7.24 | 5.32 |
| Ac–l–Val–NHa–Fn–NHb–l–Val–Boc (19) | 9.02 | 9.29 | 6.74 | 5.29 |
| Ac–l–Leu–NHa–Fn–NHb–l–Leu–Boc (21) | 9.13 | 9.36 | 7.28 | 5.14 |
| NHFna | NHFnb | NHAc | NHBoc | |
| Δδ (ppm) | ||||
| Ac–l–Phe–NHa–Fn–NHb–l–Phe–Boc (17) | 0.16 | 0.07 | 1.44 | 1.77 |
| Ac–l–Val–NHa–Fn–NHb–l–Val–Boc (19) | 0.11 | 0.15 | 1.29 | 1.52 |
| Ac–l–Leu–NHa–Fn–NHb–l–Leu–Boc (21) | 0 | 0.05 | 0.96 | 1.9 |
| NHFna | NHFnb | NHAc | NHBoc | |
| Δδ/ΔT (ppb K–1) | ||||
| Ac–l–Phe–NHa–Fn–NHb–l–Phe–Boc (17) | −8.71 | −8.0 | −18.42 | −2.29 |
| Ac–l–Val–NHa–Fn–NHb–l–Val–Boc (19) | −7.14 | −5.71 | −11.57 | −1.42 |
| Ac–l–Leu–NHa–Fn–NHb–l–Leu–Boc (21) | −6.57 | −6.57 | −17.71 | −1.57 |
| Compound | Mθ (deg cm2 dmol−1) |
|---|---|
| Ac–l–Phe–NH–Fn–NH–l–Phe–Boc (17) | 19.542 |
| Ac–d–Phe–NH–Fn–NH–d–Phe–Boc (18) | −20.197 |
| Ac–l–Val–NH–Fn–NH–l–Val–Boc (19) | 25.725 |
| Ac–d–Val–NH–Fn–NH–d–Val–Boc (20) | −26.442 |
| Ac–l–Leu–NH–Fn–NH–l–Leu–Boc (21) | 25.357 |
| Ac–d–Leu–NH–Fn–NH–d–Leu–Boc (22) | −28.407 |
| IC50 (μM) | |||
|---|---|---|---|
| Compound | HeLa | HepG2 | MCF-7 |
| Ac–l–Phe–NH–Fn–NH–l–Phe–Boc (17) | >350 | >350 | 53.1 ± 23 |
| Ac–d–Phe–NH–Fn–NH–d–Phe–Boc (18) | >350 | >350 | 32.7 ± 6.89 |
| Ac–l–Val–NH–Fn–NH–l–Val–Boc (19) | >350 | >350 | >350 |
| Ac–d–Val–NH–Fn–NH–d–Val–Boc (20) | >350 | >350 | >350 |
| Ac–l–Leu–NH–Fn–NH–l–Leu–Boc (21) | 331 ± 39 | >350 | 261 ± 97 |
| Ac–d–Leu–NH–Fn–NH–d–Leu–Boc (22) | 80.8 ± 15 | >350 | >350 |
| Cisplatin | 46.14 a | 15.9 b | 97.86 a |
| Compound (c = 1 mM) | FRAP (mM Trolox) | DPPH (% Inhibition of 0.1 mM Trolox) |
|---|---|---|
| Ac–l–Phe–NH–Fn–NH–l–Phe–Boc (17) | 0.63 ± 0.21 | 113.77 ± 4.02 |
| Ac–d–Phe–NH–Fn–NH–d–Phe–Boc (18) | 0.58 ± 0.20 | 116.80 ± 18.19 |
| Ac–l–Val–NH–Fn–NH–l–Val–Boc (19) | 0.52 ± 0.06 | 102.62 ± 20.66 |
| Ac–d–Val–NH–Fn–NH–d–Val–Boc (20) | 0.62 ± 0.15 | 88.68 ± 24.52 |
| Ac–l–Leu–NH–Fn–NH–l–Leu–Boc (21) | 0.56 ± 0.17 | 81.98 ± 7.04 |
| Ac–d–Leu–NH–Fn–NH–d–Leu–Boc (22) | 0.60 ± 0.10 | 122.75 ± 2.17 |
| Trolox 0.1 mM | - | 100 ± 0.0 |
| Compound | NHFn | NHAla | NHFn | NHAla | NHFn | NHAla |
|---|---|---|---|---|---|---|
| δ (ppm) | Δδ (ppm) | Δδ/ΔT (ppb K–1) | ||||
| Boc–l–Pro–l–Pro–l–Ala–NH–Fc (23) | 8.26 | 7.67 | 0.07 | 0.07 | −4 | −4.8 |
| Boc–d–Pro–l–Pro–l–Ala–NH–Fn (24) | 8.26 | 7.11 | 0.36 | 0.03 | −2.4 | −0.8 |
| Boc–l–Pro–d–Pro–l–Ala–NH–Fn (25) | 8.04 | 7.52 | 0.55 | 0.8 | −2.8 | −2.8 |
| Boc–d–Pro–d–Pro–l–Ala–NH–Fn (26) | 8.58 | 6.50 | 0.34 | 1.96 | −6.4 | 0.8 |
| Compound | NHFn | NHAib1 | NHAib2 | NHAib3 | NHAla |
|---|---|---|---|---|---|
| δ (ppm) | |||||
| Boc–l–Ala–Aib–NH–Fn (31) | 8.38 | 6.45 | 5.11 | ||
| Boc–l–Ala–Aib–Aib–NH–Fn (32) | 8.33 | 7.04 | 6.55 | 5.10 | |
| Boc–l–Ala–Aib–Aib–Aib–NH–Fn (33) | 8.51 | 7.46 | 7.39 | 6.81 | 5.50 |
| NHFn | NHAib1 | NHAib2 | NHAib3 | NHAla | |
| Δδ (ppm) | |||||
| Boc–l–Ala–Aib–NH–Fn (31) | 0.1 | 0.8 | 1.6 | ||
| Boc–l–Ala–Aib–Aib–NH–Fn (32) | 0.2 | 0.7 | 0.6 | 1.6 | |
| Boc–l–Ala–Aib–Aib–Aib–NH–Fn (33) | 0.1 | 0.4 | 0.1 | 1.0 | 1.5 |
| NHFn | NHAib1 | NHAib2 | NHAib3 | NHAla | |
| Δδ/ΔT (ppb K–1) | |||||
| Boc–l–Ala–Aib–NH–Fn (31) | −3.2 | −0.1 | −2.1 | ||
| Boc–l–Ala–Aib–Aib–NH–Fn (32) | −2.2 | −4.8 | −0.6 | −1.9 | |
| Boc–l–Ala–Aib–Aib–Aib–NH–Fn (33) | −2.8 | −5.8 | −6.3 | −0.8 | −0.8 |
| Compound | NHfree (ῠmax/cm−1) | NHassoc. (ῠmax/cm−1) | |
|---|---|---|---|
| –(CO–Ala–NH–Fn–COOMe)2 (39) | THF | / | 3382, 3294, 3252 |
| toluene | / | 3305, 3268 |
| Compound | NHFn | NHAla | NHFn | NHAA | NHFn | NHAA | |
|---|---|---|---|---|---|---|---|
| δ (ppm) | Δδ (ppm) | Δδ/ΔT (ppb K–1) | |||||
| –(CO–Ala–NH–Fn–COOMe)2 (39) | THF | 8.67 | 8.44 | 0.9 | 0.15 | −7.7 | −5 |
| toluene | 7.90 | 8.36 | / | / | −7 | −12 | |
| IC50 (μM) | ||
|---|---|---|
| –(CO–Ala–NH–Fn–COOMe)2 (39) | –(CO–NH–Fn–COOMe)2 | |
| HEK293T | 94.88 | n.d. a,b |
| MCF-7 | 104.45 | / |
| HepG2 | 140.34 | n.d. a,b |
| HeLa | 110.20 | 252.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čakić Semenčić, M.; Kovačević, M.; Barišić, L. Recent Advances in the Field of Amino Acid-Conjugated Aminoferrocenes—A Personal Perspective. Int. J. Mol. Sci. 2024, 25, 4810. https://doi.org/10.3390/ijms25094810
Čakić Semenčić M, Kovačević M, Barišić L. Recent Advances in the Field of Amino Acid-Conjugated Aminoferrocenes—A Personal Perspective. International Journal of Molecular Sciences. 2024; 25(9):4810. https://doi.org/10.3390/ijms25094810
Chicago/Turabian StyleČakić Semenčić, Mojca, Monika Kovačević, and Lidija Barišić. 2024. "Recent Advances in the Field of Amino Acid-Conjugated Aminoferrocenes—A Personal Perspective" International Journal of Molecular Sciences 25, no. 9: 4810. https://doi.org/10.3390/ijms25094810
APA StyleČakić Semenčić, M., Kovačević, M., & Barišić, L. (2024). Recent Advances in the Field of Amino Acid-Conjugated Aminoferrocenes—A Personal Perspective. International Journal of Molecular Sciences, 25(9), 4810. https://doi.org/10.3390/ijms25094810