
Lectin-Based Immunophenotyping and Whole Proteomic Profiling of CT-26 Colon Carcinoma Murine Model

 , , ,
, , , 

Abstract
1. Introduction
2. Results
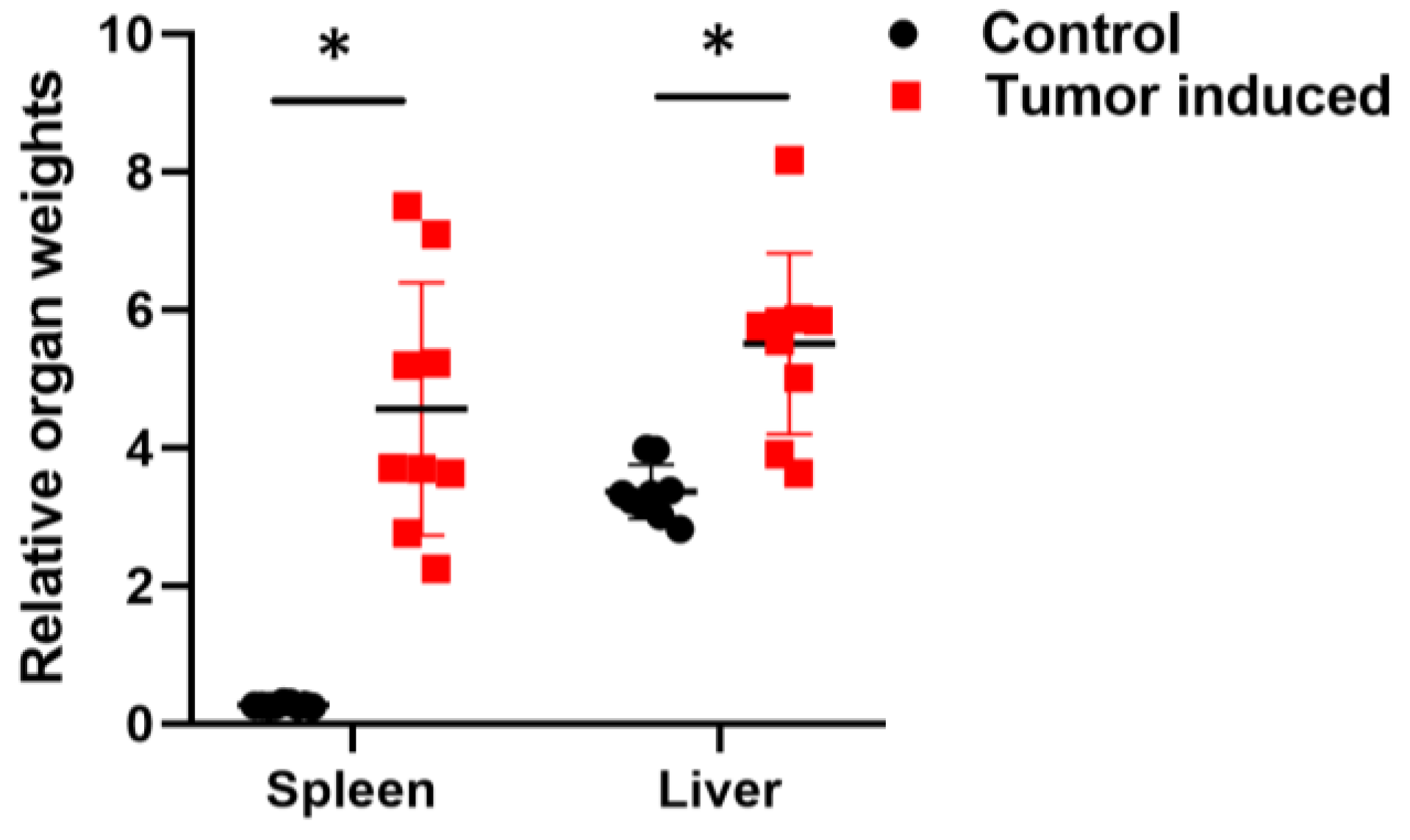
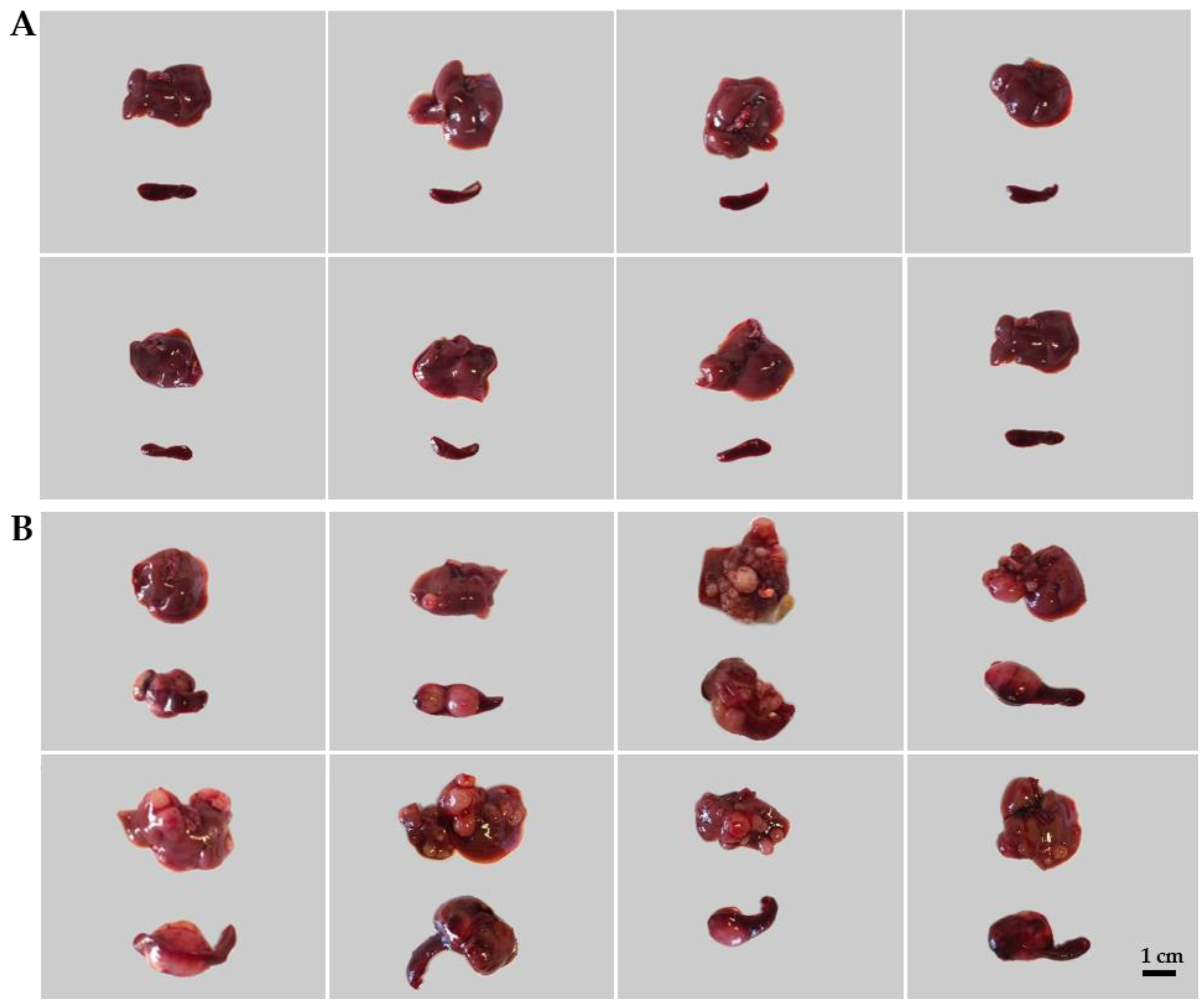
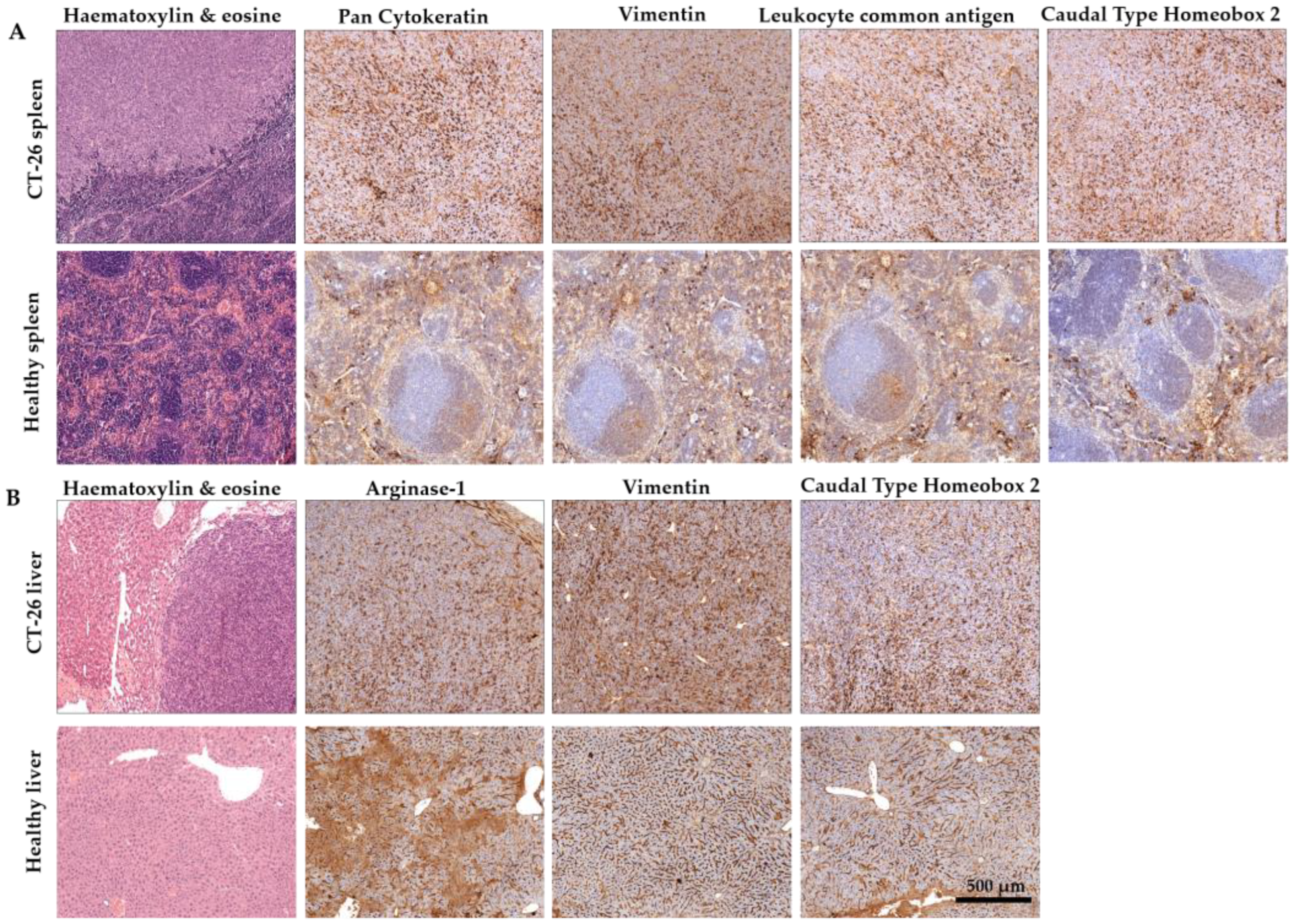
2.1. Establishment of Murine CRC
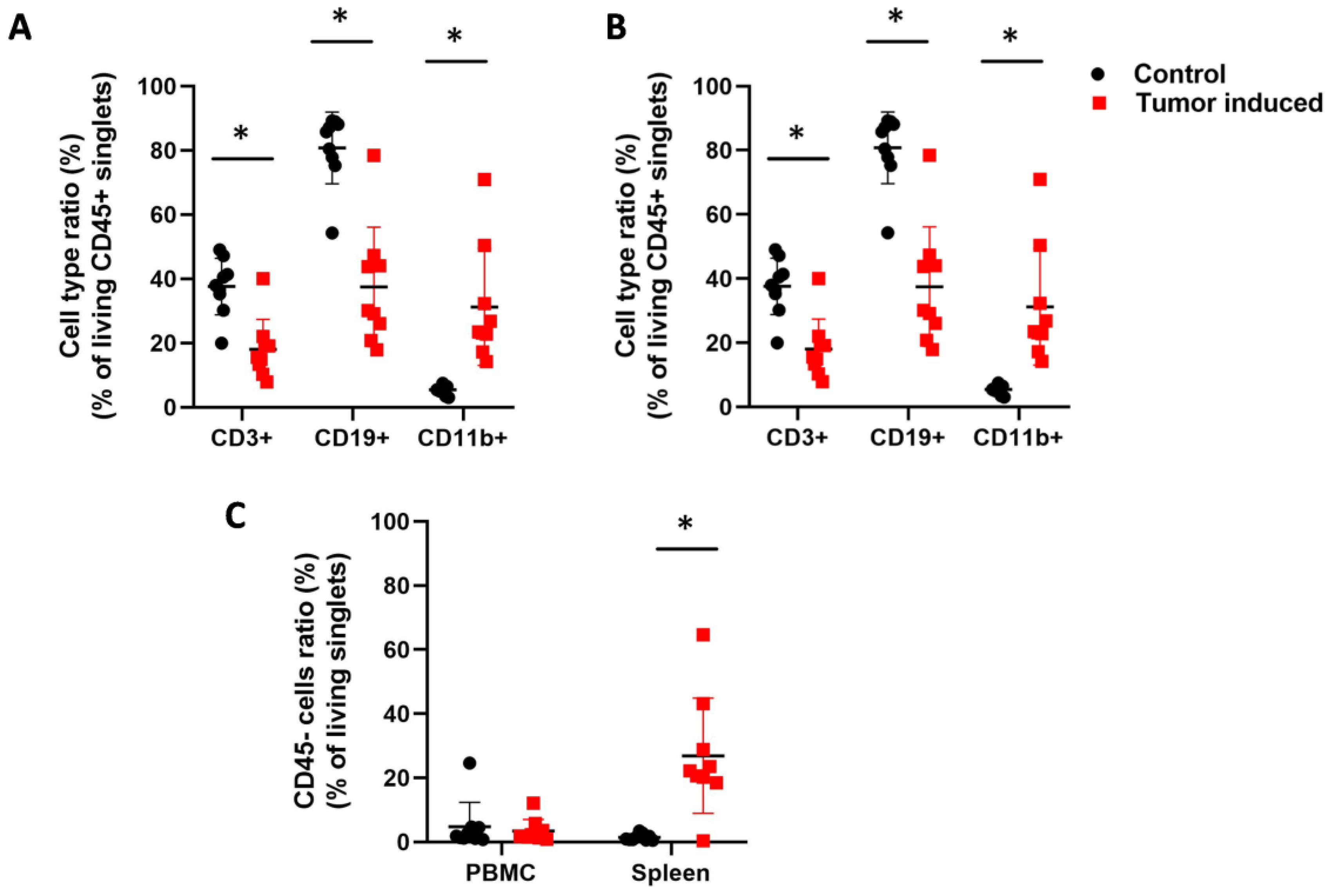
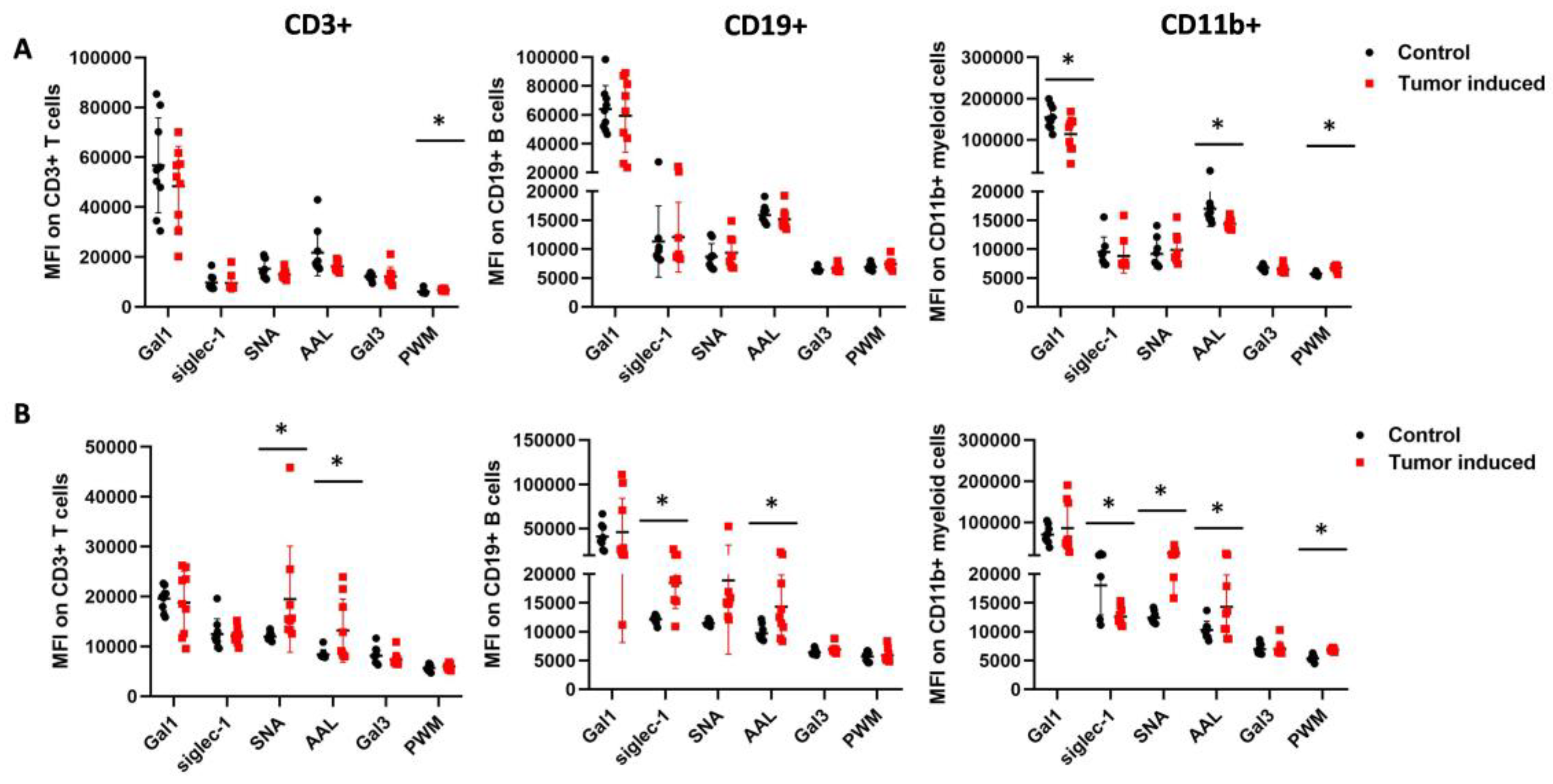
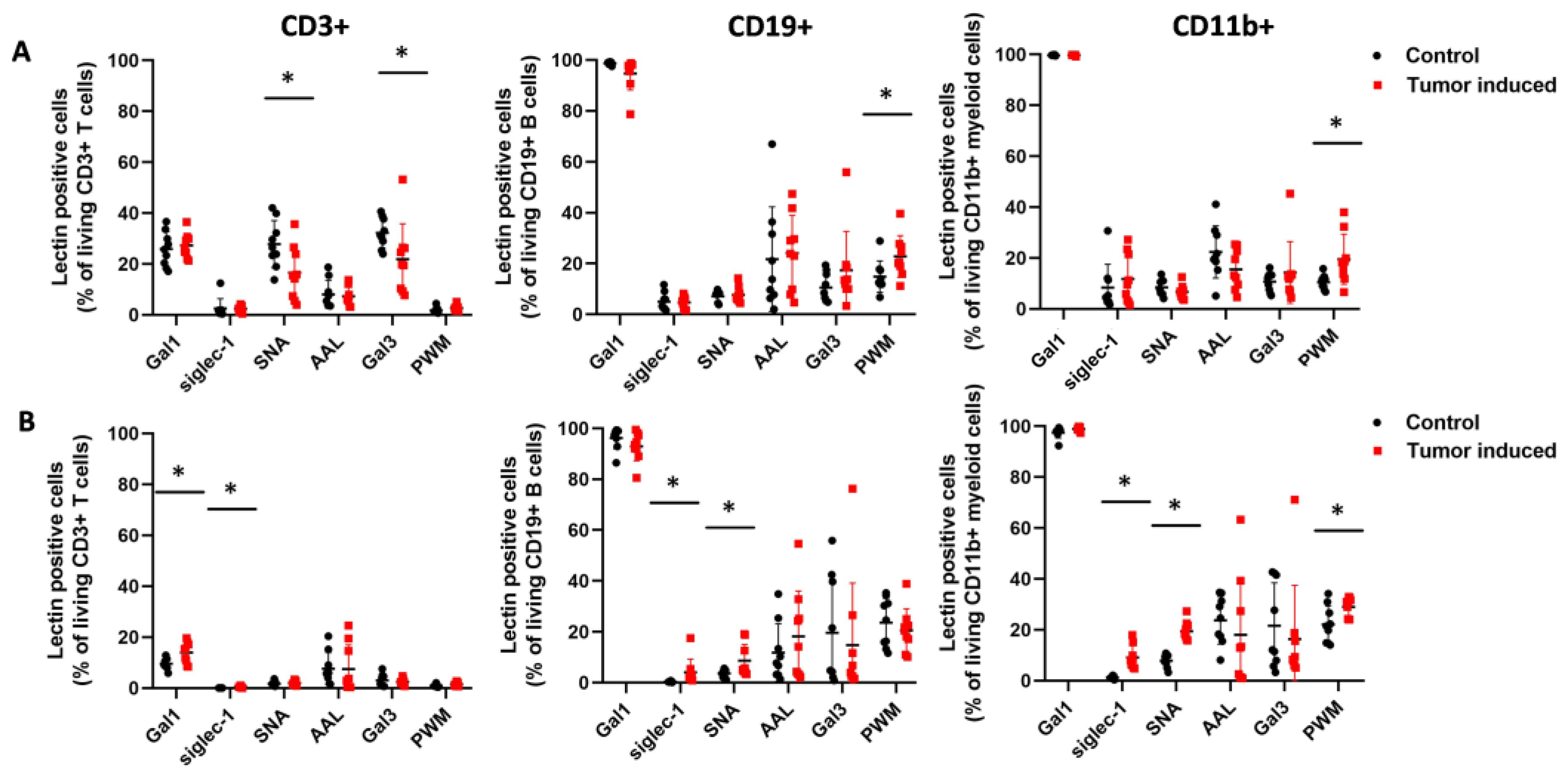
2.2. Flow Cytometric Immunophenotyping of the Leukocytes in CRC-Bearing Mice Using Fluorescently Labeled Lectins
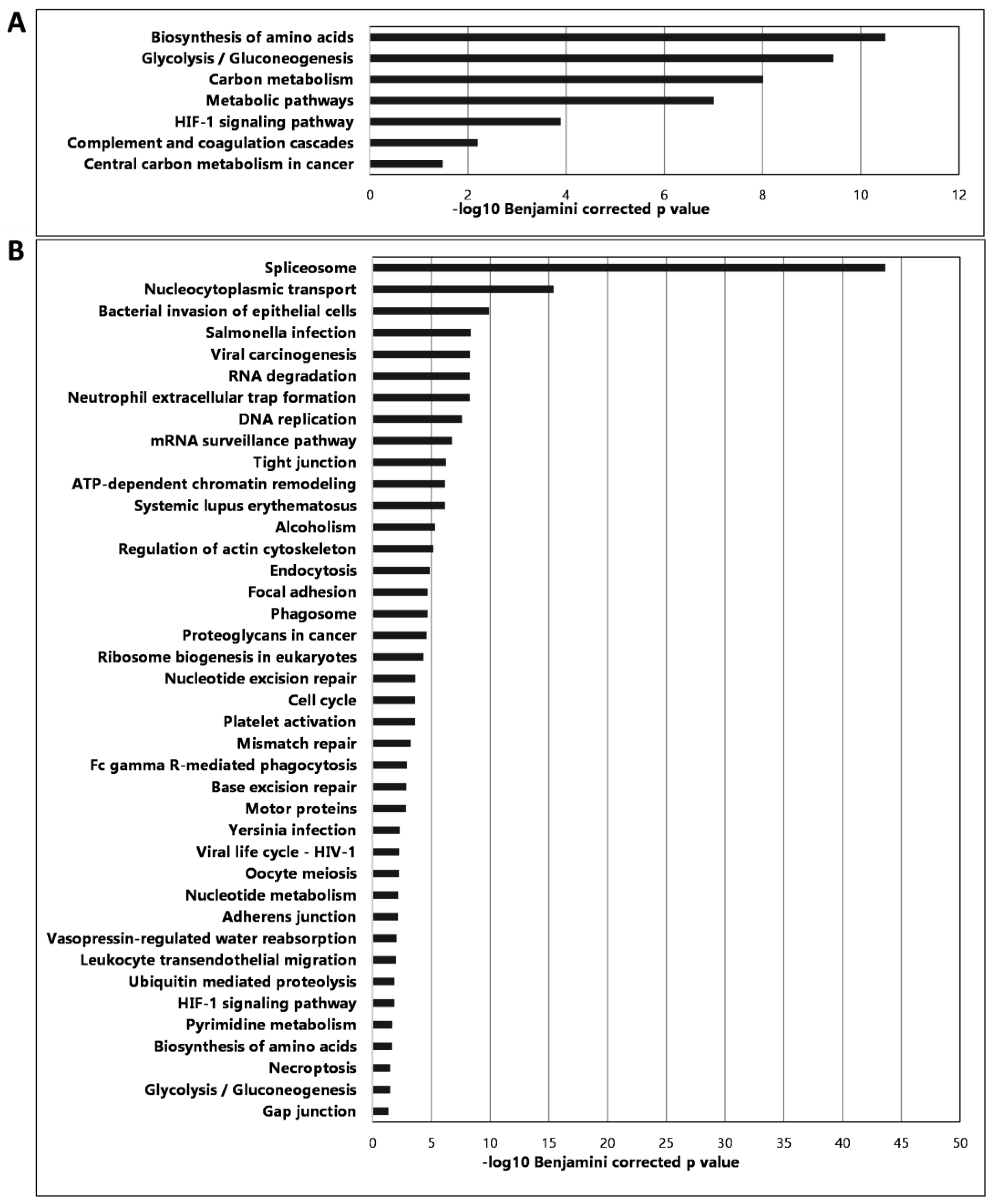
2.3. Proteomic Analysis of the Established Murine CT26 Tumor Tissues
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Lines and Tissue Culture Conditions
4.3. Spleen Injection
4.4. Immunohistochemistry
4.5. Peripheral White Blood Cell Isolation
4.6. Spleen Cell Isolation
4.7. Lectin Labeling
4.8. Lectin Agglutination Test
4.9. Labeling and Flow Cytometry
4.10. Proteomic Characterization
4.11. Data Visualization and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Wagle, N.S.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 233–254. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Juul, J.S.; Hornung, N.; Andersen, B.; Laurberg, S.; Olesen, F.; Vedsted, P. The value of using the faecal immunochemical test in general practice on patients presenting with non-alarm symptoms of colorectal cancer. Br. J. Cancer 2018, 119, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, S.; Lepage, C.; Hatem, C.; Coatmeur, O.; Faivre, J.; Bouvier, A.M. Epidemiology and management of liver metastases from colorectal cancer. Ann. Surg. 2006, 244, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Burtin, F.; Mullins, C.S.; Linnebacher, M. Mouse models of colorectal cancer: Past, present and future perspectives. World J. Gastroenterol. 2020, 26, 1394–1426. [Google Scholar] [CrossRef] [PubMed]
- Mlecnik, B.; Tosolini, M.; Kirilovsky, A.; Berger, A.; Bindea, G.; Meatchi, T.; Bruneval, P.; Trajanoski, Z.; Fridman, W.H.; Pages, F.; et al. Histopathologic-based prognostic factors of colorectal cancers are associated with the state of the local immune reaction. J. Clin. Oncol. 2011, 29, 610–618. [Google Scholar] [CrossRef]
- Edin, S.; Kaprio, T.; Hagstrom, J.; Larsson, P.; Mustonen, H.; Bockelman, C.; Strigard, K.; Gunnarsson, U.; Haglund, C.; Palmqvist, R. The Prognostic Importance of CD20(+) B lymphocytes in Colorectal Cancer and the Relation to Other Immune Cell subsets. Sci. Rep. 2019, 9, 19997. [Google Scholar] [CrossRef] [PubMed]
- Coppola, A.; Arriga, R.; Lauro, D.; Del Principe, M.I.; Buccisano, F.; Maurillo, L.; Palomba, P.; Venditti, A.; Sconocchia, G. NK Cell Inflammation in the Clinical Outcome of Colorectal Carcinoma. Front. Med. 2015, 2, 33. [Google Scholar] [CrossRef]
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting: Integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef]
- Szebeni, G.J.; Vizler, C.; Nagy, L.I.; Kitajka, K.; Puskas, L.G. Pro-Tumoral Inflammatory Myeloid Cells as Emerging Therapeutic Targets. Int. J. Mol. Sci. 2016, 17, 1958. [Google Scholar] [CrossRef]
- Szebeni, G.J.; Vizler, C.; Kitajka, K.; Puskas, L.G. Inflammation and Cancer: Extra- and Intracellular Determinants of Tumor-Associated Macrophages as Tumor Promoters. Mediat. Inflamm. 2017, 2017, 9294018. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Myers, J.S.; Wang, F.; Wang, K.; Lucas, J.; Rosfjord, E.; Lucas, J.; Hooper, A.T.; Yang, S.; Lemon, L.A.; et al. Comparison of the molecular and cellular phenotypes of common mouse syngeneic models with human tumors. BMC Genom. 2020, 21, 2. [Google Scholar] [CrossRef] [PubMed]
- Castle, J.C.; Loewer, M.; Boegel, S.; de Graaf, J.; Bender, C.; Tadmor, A.D.; Boisguerin, V.; Bukur, T.; Sorn, P.; Paret, C.; et al. Immunomic, genomic and transcriptomic characterization of CT26 colorectal carcinoma. BMC Genom. 2014, 15, 190. [Google Scholar] [CrossRef]
- Kollmar, O.; Schilling, M.K.; Menger, M.D. Experimental liver metastasis: Standards for local cell implantation to study isolated tumor growth in mice. Clin. Exp. Metastasis 2004, 21, 453–460. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; van Kooyk, Y.; Cobb, B.A. Glycobiology of immune responses. Ann. N. Y. Acad. Sci. 2012, 1253, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Szabo, E.; Hornung, A.; Monostori, E.; Bocskai, M.; Czibula, A.; Kovacs, L. Altered Cell Surface N-Glycosylation of Resting and Activated T Cells in Systemic Lupus Erythematosus. Int. J. Mol. Sci. 2019, 20, 4455. [Google Scholar] [CrossRef] [PubMed]
- Mody, R.; Joshi, S.; Chaney, W. Use of lectins as diagnostic and therapeutic tools for cancer. J. Pharmacol. Toxicol. Methods 1995, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tuccillo, F.M.; de Laurentiis, A.; Palmieri, C.; Fiume, G.; Bonelli, P.; Borrelli, A.; Tassone, P.; Scala, I.; Buonaguro, F.M.; Quinto, I.; et al. Aberrant glycosylation as biomarker for cancer: Focus on CD43. Biomed. Res. Int. 2014, 2014, 742831. [Google Scholar] [CrossRef]
- Chen, C.Y.; Jan, Y.H.; Juan, Y.H.; Yang, C.J.; Huang, M.S.; Yu, C.J.; Yang, P.C.; Hsiao, M.; Hsu, T.L.; Wong, C.H. Fucosyltransferase 8 as a functional regulator of nonsmall cell lung cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 630–635. [Google Scholar] [CrossRef]
- Moriwaki, K.; Miyoshi, E. Fucosylation and gastrointestinal cancer. World J. Hepatol. 2010, 2, 151–161. [Google Scholar] [CrossRef]
- Dobie, C.; Skropeta, D. Insights into the role of sialylation in cancer progression and metastasis. Br. J. Cancer 2021, 124, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Bull, C.; Stoel, M.A.; den Brok, M.H.; Adema, G.J. Sialic acids sweeten a tumor’s life. Cancer Res. 2014, 74, 3199–3204. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, G.J.; Kriston-Pal, E.; Blazso, P.; Katona, R.L.; Novak, J.; Szabo, E.; Czibula, A.; Fajka-Boja, R.; Hegyi, B.; Uher, F.; et al. Identification of galectin-1 as a critical factor in function of mouse mesenchymal stromal cell-mediated tumor promotion. PLoS ONE 2012, 7, e41372. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Patwa, T.H.; Xu, L.; Shedden, K.; Misek, D.E.; Tuck, M.; Jin, G.; Ruffin, M.T.; Turgeon, D.K.; Synal, S.; et al. Plasma glycoprotein profiling for colorectal cancer biomarker identification by lectin glycoarray and lectin blot. J. Proteome Res. 2008, 7, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Setayesh, T.; Colquhoun, S.D.; Wan, Y.Y. Overexpression of Galectin-1 and Galectin-3 in hepatocellular carcinoma. Liver Res. 2020, 4, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, X.; Chen, M.; Zhang, F.; Liu, X.; Yu, J.; Zhong, Y.; Shu, J.; Chen, W.; Du, H.; et al. Integrated Glycome Strategy for Characterization of Aberrant LacNAc Contained N-Glycans Associated With Gastric Carcinoma. Front. Oncol. 2019, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.A.; Zidell, R.H.; Perry, R.W. Relationships between organ weight and body/brain weight in the rat: What is the best analytical endpoint? Toxicol. Pathol. 2004, 32, 448–466. [Google Scholar] [CrossRef] [PubMed]
- Sellers, R.S.; Morton, D.; Michael, B.; Roome, N.; Johnson, J.K.; Yano, B.L.; Perry, R.; Schafer, K. Society of Toxicologic Pathology position paper: Organ weight recommendations for toxicology studies. Toxicol. Pathol. 2007, 35, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Piao, Y.; Liu, Y.; Xie, X. Change trends of organ weight background data in sprague dawley rats at different ages. J. Toxicol. Pathol. 2013, 26, 29–34. [Google Scholar] [CrossRef]
- Moll, R.; Franke, W.W.; Schiller, D.L.; Geiger, B.; Krepler, R. The catalog of human cytokeratins: Patterns of expression in normal epithelia, tumors and cultured cells. Cell 1982, 31, 11–24. [Google Scholar] [CrossRef]
- Kleist, B.; Xu, L.; Li, G.; Kersten, C. Expression of the adult intestinal stem cell marker Lgr5 in the metastatic cascade of colorectal cancer. Int. J. Clin. Exp. Pathol. 2011, 4, 327–335. [Google Scholar] [PubMed]
- Jepsen, R.K.; Klarskov, L.L.; Lippert, M.F.; Novotny, G.W.; Hansen, T.P.; Christensen, I.J.; Hogdall, E.; Riis, L.B. Digital image analysis of pan-cytokeratin stained tumor slides for evaluation of tumor budding in pT1/pT2 colorectal cancer: Results of a feasibility study. Pathol. Res. Pract. 2018, 214, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, G.J.; Alfoldi, R.; Nagy, L.I.; Neuperger, P.; Gemes, N.; Balog, J.A.; Tiszlavicz, L.; Puskas, L.G. Introduction of an Ultraviolet C-Irradiated 4T1 Murine Breast Cancer Whole-Cell Vaccine Model. Vaccines 2023, 11, 1254. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhu, G.; Lin, C.; Lin, P.; Chen, H.; He, R.; Huang, Y.; Yang, S.; Ye, J. Vimentin affects colorectal cancer proliferation, invasion, and migration via regulated by activator protein 1. J. Cell. Physiol. 2021, 236, 7591–7604. [Google Scholar] [CrossRef] [PubMed]
- Al-Maghrabi, J. Vimentin immunoexpression is associated with higher tumor grade, metastasis, and shorter survival in colorectal cancer. Int. J. Clin. Exp. Pathol. 2020, 13, 493–500. [Google Scholar]
- Park, S.Y.; Kim, J.Y.; Jang, G.B.; Choi, J.H.; Kim, J.H.; Lee, C.J.; Lee, S.; Baek, J.H.; Park, K.K.; Kim, J.M.; et al. Aberrant activation of the CD45-Wnt signaling axis promotes stemness and therapy resistance in colorectal cancer cells. Theranostics 2021, 11, 8755–8770. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.B.; Qiao, L.N.; Sun, X.J.; Qi, J.; Ren, H.L.; Wei, G.B.; Zhou, P.H.; Yao, J.F.; Zhang, L.; Jia, P.B. Overexpression of caudal-related homeobox transcription factor 2 inhibits the growth of transplanted colorectal tumors in nude mice. Mol. Med. Rep. 2015, 12, 3409–3415. [Google Scholar] [CrossRef] [PubMed]
- Aasebo, K.; Dragomir, A.; Sundstrom, M.; Mezheyeuski, A.; Edqvist, P.H.; Eide, G.E.; Ponten, F.; Pfeiffer, P.; Glimelius, B.; Sorbye, H. CDX2: A Prognostic Marker in Metastatic Colorectal Cancer Defining a Better BRAF Mutated and a Worse KRAS Mutated Subgroup. Front. Oncol. 2020, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Hinoi, T.; Loda, M.; Fearon, E.R. Silencing of CDX2 expression in colon cancer via a dominant repression pathway. J. Biol. Chem. 2003, 278, 44608–44616. [Google Scholar] [CrossRef]
- Ma, Z.; Lian, J.; Yang, M.; Wuyang, J.; Zhao, C.; Chen, W.; Liu, C.; Zhao, Q.; Lou, C.; Han, J.; et al. Overexpression of Arginase-1 is an indicator of poor prognosis in patients with colorectal cancer. Pathol. Res. Pract. 2019, 215, 152383. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Y.; Wang, R.; Dai, N.; Zhang, W.; Li, F. The significance of arginase-1 expression in the diagnosis of liver cancer: A protocol for a systematic review. Medicine 2020, 99, e19159. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, M.; Kwok, S.; Yano, H.; Pai, R.K. Arginase-1 is a more sensitive marker of hepatic differentiation than HepPar-1 and glypican-3 in fine-needle aspiration biopsies. Cancer Cytopathol. 2012, 120, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.A.; Drake, V.; Huang, H.S.; Chiu, S.; Zheng, L. Reprogramming the tumor microenvironment: Tumor-induced immunosuppressive factors paralyze T cells. Oncoimmunology 2015, 4, e1016700. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wei, G.; Liu, D. CD19: A biomarker for B cell development, lymphoma diagnosis and therapy. Exp. Hematol. Oncol. 2012, 1, 36. [Google Scholar] [CrossRef] [PubMed]
- Balog, J.A.; Hackler, L., Jr.; Kovacs, A.K.; Neuperger, P.; Alfoldi, R.; Nagy, L.I.; Puskas, L.G.; Szebeni, G.J. Single Cell Mass Cytometry Revealed the Immunomodulatory Effect of Cisplatin Via Downregulation of Splenic CD44+, IL-17A+ MDSCs and Promotion of Circulating IFN-gamma+ Myeloid Cells in the 4T1 Metastatic Breast Cancer Model. Int. J. Mol. Sci. 2019, 21, 170. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hou, W.; Zhong, M.; Wu, B. Comprehensive Proteomic Analysis of Colon Cancer Tissue Revealed the Reason for the Worse Prognosis of Right-Sided Colon Cancer and Mucinous Colon Cancer at the Protein Level. Curr. Oncol. 2021, 28, 3554–3572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Davis, C.; Ryan, J.; Janney, C.; Pena, M.M. Development and characterization of a reliable mouse model of colorectal cancer metastasis to the liver. Clin. Exp. Metastasis 2013, 30, 903–918. [Google Scholar] [CrossRef] [PubMed]
- Luster, A.D.; Alon, R.; von Andrian, U.H. Immune cell migration in inflammation: Present and future therapeutic targets. Nat. Immunol. 2005, 6, 1182–1190. [Google Scholar] [CrossRef]
- Wild, M.K.; Luhn, K.; Marquardt, T.; Vestweber, D. Leukocyte adhesion deficiency II: Therapy and genetic defect. Cells Tissues Organs 2002, 172, 161–173. [Google Scholar] [CrossRef]
- Jellusova, J.; Nitschke, L. Regulation of B cell functions by the sialic acid-binding receptors siglec-G and CD22. Front. Immunol. 2011, 2, 96. [Google Scholar] [CrossRef]
- Tateno, H.; Toyota, M.; Saito, S.; Onuma, Y.; Ito, Y.; Hiemori, K.; Fukumura, M.; Matsushima, A.; Nakanishi, M.; Ohnuma, K.; et al. Glycome diagnosis of human induced pluripotent stem cells using lectin microarray. J. Biol. Chem. 2011, 286, 20345–20353. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Chammas, R.; Bellis, S.L. Sialylation of beta1 integrins blocks cell adhesion to galectin-3 and protects cells against galectin-3-induced apoptosis. J. Biol. Chem. 2008, 283, 22177–22185. [Google Scholar] [CrossRef]
- Le Minh, G.; Esquea, E.M.; Young, R.G.; Huang, J.; Reginato, M.J. On a sugar high: Role of O-GlcNAcylation in cancer. J. Biol. Chem. 2023, 299, 105344. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wen, T.; Zhu, M.; Li, L.; Wei, J.; Wu, X.; Guo, M.; Liu, S.; Zhao, H.; Xia, S.; et al. Glycoproteomic analysis of tissues from patients with colon cancer using lectin microarrays and nanoLC-MS/MS. Mol. Biosyst. 2013, 9, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
- Ishida, H.; Togayachi, A.; Sakai, T.; Iwai, T.; Hiruma, T.; Sato, T.; Okubo, R.; Inaba, N.; Kudo, T.; Gotoh, M.; et al. A novel beta1,3-N-acetylglucosaminyltransferase (beta3Gn-T8), which synthesizes poly-N-acetyllactosamine, is dramatically upregulated in colon cancer. FEBS Lett. 2005, 579, 71–78. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, T.K.; Honing, H.; Franke, N.; van Remoortere, A.; Schiphorst, W.E.; Liu, F.T.; Deelder, A.M.; Cummings, R.D.; Hokke, C.H.; van Die, I. LacdiNAc-glycans constitute a parasite pattern for galectin-3-mediated immune recognition. J. Immunol. 2004, 173, 1902–1907. [Google Scholar] [CrossRef] [PubMed]
- de Freitas Junior, J.C.; Morgado-Diaz, J.A. The role of N-glycans in colorectal cancer progression: Potential biomarkers and therapeutic applications. Oncotarget 2016, 7, 19395–19413. [Google Scholar] [CrossRef]
- Vukobrat-Bijedic, Z.; Husic-Selimovic, A.; Sofic, A.; Bijedic, N.; Bjelogrlic, I.; Gogov, B.; Mehmedovic, A. Cancer Antigens (CEA and CA 19-9) as Markers of Advanced Stage of Colorectal Carcinoma. Med. Arch. 2013, 67, 397–401. [Google Scholar] [CrossRef]
- Leviatan Ben-Arye, S.; Yu, H.; Chen, X.; Padler-Karavani, V. Profiling Anti-Neu5Gc IgG in Human Sera with a Sialoglycan Microarray Assay. J. Vis. Exp. 2017, 13, 56094. [Google Scholar] [CrossRef]
- Chang, I.J.; He, M.; Lam, C.T. Congenital disorders of glycosylation. Ann. Transl. Med. 2018, 6, 477. [Google Scholar] [CrossRef]
- Palmieri, C.; Caley, M.P.; Purshouse, K.; Fonseca, A.V.; Rodriguez-Teja, M.; Kogianni, G.; Woodley, L.; Odendaal, J.; Elliott, K.; Waxman, J.; et al. Endo180 modulation by bisphosphonates and diagnostic accuracy in metastatic breast cancer. Br. J. Cancer 2013, 108, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Tupa, V.; Drahosova, S.; Grendar, M.; Adamkov, M. Expression and association of carbonic anhydrase IX and cyclooxygenase-2 in colorectal cancer. Pathol. Res. Pract. 2019, 215, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Korkeila, E.; Talvinen, K.; Jaakkola, P.M.; Minn, H.; Syrjanen, K.; Sundstrom, J.; Pyrhonen, S. Expression of carbonic anhydrase IX suggests poor outcome in rectal cancer. Br. J. Cancer 2009, 100, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.M.; Lee, H.J.; Kim, S.H.; Kim, H.K.; Mok, Y.J.; Park, Y.T.; Choi, J.S.; Moon, H.Y. Expression of protein S100A4 is a predictor of recurrence in colorectal cancer. World J. Gastroenterol. 2010, 16, 3897–3904. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Wu, T.; Wang, K.; Luo, S.; Chen, Z.; Fan, M.; Xue, D.; Lu, H.; Zhuang, Q.; Xu, X. Prognostic significance of galectin-1 expression in patients with cancer: A meta-analysis. Cancer Cell Int. 2018, 18, 108. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.Y.; Jiang, S.S.; Lee, Y.W.; Tsai, F.Y.; Chang, C.C.; Chen, L.T.; Yen, B.L. Stromal Galectin-1 Promotes Colorectal Cancer Cancer-Initiating Cell Features and Disease Dissemination Through SOX9 and beta-Catenin: Development of Niche-Based Biomarkers. Front. Oncol. 2021, 11, 716055. [Google Scholar] [CrossRef]
- Aureli, A.; Del Corno, M.; Marziani, B.; Gessani, S.; Conti, L. Highlights on the Role of Galectin-3 in Colorectal Cancer and the Preventive/Therapeutic Potential of Food-Derived Inhibitors. Cancers 2022, 15, 52. [Google Scholar] [CrossRef]
- Luo, Z.W.; Zhu, M.G.; Zhang, Z.Q.; Ye, F.J.; Huang, W.H.; Luo, X.Z. Increased expression of Ki-67 is a poor prognostic marker for colorectal cancer patients: A meta analysis. BMC Cancer 2019, 19, 123. [Google Scholar] [CrossRef]
- Suren, D.; Yildirim, M.; Demirpence, O.; Kaya, V.; Alikanoglu, A.S.; Bulbuller, N.; Yildiz, M.; Sezer, C. The role of high mobility group box 1 (HMGB1) in colorectal cancer. Med. Sci. Monit. 2014, 20, 530–537. [Google Scholar] [CrossRef]
- Puskas, L.G.; Man, I.; Szebeni, G.; Tiszlavicz, L.; Tsai, S.; James, M.A. Novel Anti-CRR9/CLPTM1L Antibodies with Antitumorigenic Activity Inhibit Cell Surface Accumulation, PI3K Interaction, and Survival Signaling. Mol. Cancer Ther. 2016, 15, 985–997. [Google Scholar] [CrossRef]
- Neuperger, P.; Balog, J.A.; Tiszlavicz, L.; Furak, J.; Gemes, N.; Kotogany, E.; Szalontai, K.; Puskas, L.G.; Szebeni, G.J. Analysis of the Single-Cell Heterogeneity of Adenocarcinoma Cell Lines and the Investigation of Intratumor Heterogeneity Reveals the Expression of Transmembrane Protein 45A (TMEM45A) in Lung Adenocarcinoma Cancer Patients. Cancers 2021, 14, 144. [Google Scholar] [CrossRef] [PubMed]
- Kotogany, E.; Balog, J.A.; Nagy, L.I.; Alfoldi, R.; Bertagnolo, V.; Brugnoli, F.; Demjen, A.; Kovacs, A.K.; Batar, P.; Mezei, G.; et al. Imidazo[1,2-b]pyrazole-7-Carboxamide Derivative Induces Differentiation-Coupled Apoptosis of Immature Myeloid Cells Such as Acute Myeloid Leukemia and Myeloid-Derived Suppressor Cells. Int. J. Mol. Sci. 2020, 21, 5135. [Google Scholar] [CrossRef] [PubMed]
- Honfi, D.; Gemes, N.; Szabo, E.; Neuperger, P.; Balog, J.A.; Nagy, L.I.; Toldi, G.; Puskas, L.G.; Szebeni, G.J.; Balog, A. Comparison of Homologous and Heterologous Booster SARS-CoV-2 Vaccination in Autoimmune Rheumatic and Musculoskeletal Patients. Int. J. Mol. Sci. 2022, 23, 1411. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, G.J.; Gemes, N.; Honfi, D.; Szabo, E.; Neuperger, P.; Balog, J.A.; Nagy, L.I.; Szekanecz, Z.; Puskas, L.G.; Toldi, G.; et al. Humoral and Cellular Immunogenicity and Safety of Five Different SARS-CoV-2 Vaccines in Patients With Autoimmune Rheumatic and Musculoskeletal Diseases in Remission or With Low Disease Activity and in Healthy Controls: A Single Center Study. Front. Immunol. 2022, 13, 846248. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number (Uniprot) | Description | Fold Change (Tumor/Control) |
|---|---|---|
| P19437 | B-lymphocyte antigen CD20 | −38.46 |
| Q91WN4 | Kynurenine 3-monooxygenase | −31.25 |
| Q62507 | Cochlin | −26.32 |
| P42209 | Septin-1 | −25.64 |
| P11911 | B-cell antigen receptor complex-associated protein alpha chain | −24.39 |
| P04224 | H-2 class II histocompatibility antigen, E-K alpha chain | −22.22 |
| Q8BV49 | Pyrin and HIN domain-containing protein 1 | −21.74 |
| Q99JY3 | GTPase IMAP family member 4 | −20.41 |
| Q9ESM6 | Glycerophosphoinositol inositolphosphodiesterase GDPD2 | −20 |
| P11032 | Granzyme A | −19.61 |
| Q8VHB5 | Carbonic anhydrase 9 | 26.16 |
| P06837 | Neuromodulin | 27.13 |
| P30204 | Macrophage scavenger receptor types I and II | 27.82 |
| P22935 | Cellular retinoic acid-binding protein 2 | 35.85 |
| P0DOG8 | Glyco-Gag protein | 43.60 |
| P52927 | High mobility group protein HMGI-C | 46.88 |
| P70202 | Latexin | 56.80 |
| P52624 | Uridine phosphorylase 1 | 65.19 |
| E9Q7T7 | Chondroadherin-like protein | 68.25 |
| Q64437 | All-trans-retinol dehydrogenase [NAD(+)] ADH7 | 100 |
| Accession Number (Uniprot) | Description | Fold Change (Tumor/Control) |
|---|---|---|
| P11589 | Major urinary protein 2 | −55.56 |
| P20852 | Cytochrome P450 2A5 | −27.03 |
| P04939 | Major urinary protein 3 | −25.64 |
| P00186 | Cytochrome P450 1A2 | −21.74 |
| O09158 | Cytochrome P450 3A25 | −21.74 |
| Q64176 | Carboxylesterase 1E | −21.74 |
| Q64459 | Cytochrome P450 3A11 | −17.24 |
| Q8R1S9 | Sodium-coupled neutral amino acid transporter 4 | −16.95 |
| Q64458 | Cytochrome P450 2C29 | −15.87 |
| Q571F8 | Glutaminase liver isoform, mitochondrial | −15.15 |
| Q9R1Q7 | Proteolipid protein 2 | 68.83 |
| P16110 | Galectin-3 | 73.50 |
| P97310 | DNA replication licensing factor MCM2 | 81.89 |
| Q8K009 | Mitochondrial 10-formyltetrahydrofolate dehydrogenase | 82.70 |
| Q61753 | D-3-phosphoglycerate dehydrogenase | 100 |
| P25206 | DNA replication licensing factor MCM3 | 100 |
| P52927 | High mobility group protein HMGI-C | 100 |
| P14069 | Protein S100-A6 | 100 |
| P70202 | Latexin | 100 |
| Q64437 | All-trans-retinol dehydrogenase [NAD(+)] ADH7 | 100 |
| Lectin | Abbreviations | Preferred Sugar Specificity | Fluorescent Dye |
|---|---|---|---|
| Phytolacca americana lectin | PWM/PWA | N-Acetylglucosamine | Alexa Fluor 700 |
| Aleuria aurantia lectin | AAL | Fucose, Arabinose | APC |
| Sambucus nigra lectin | SNA/EBL I | α 2-6 Sialic Acid | PE/Cy7 |
| Galectin-1 | Gal1 | N-acetyllactosamine | Alexa Fluor 488 |
| Galectin-3 | Gal3 | N-acetyllactosamine | APC/Cy7 |
| Sialoadhesin | siglec-1/CD | α 2-3 Sialic Acid | PE/Texas Red |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faragó, A.; Zvara, Á.; Tiszlavicz, L.; Hunyadi-Gulyás, É.; Darula, Z.; Hegedűs, Z.; Szabó, E.; Surguta, S.E.; Tóvári, J.; Puskás, L.G.; et al. Lectin-Based Immunophenotyping and Whole Proteomic Profiling of CT-26 Colon Carcinoma Murine Model. Int. J. Mol. Sci. 2024, 25, 4022. https://doi.org/10.3390/ijms25074022
Faragó A, Zvara Á, Tiszlavicz L, Hunyadi-Gulyás É, Darula Z, Hegedűs Z, Szabó E, Surguta SE, Tóvári J, Puskás LG, et al. Lectin-Based Immunophenotyping and Whole Proteomic Profiling of CT-26 Colon Carcinoma Murine Model. International Journal of Molecular Sciences. 2024; 25(7):4022. https://doi.org/10.3390/ijms25074022
Chicago/Turabian StyleFaragó, Anna, Ágnes Zvara, László Tiszlavicz, Éva Hunyadi-Gulyás, Zsuzsanna Darula, Zoltán Hegedűs, Enikő Szabó, Sára Eszter Surguta, József Tóvári, László G. Puskás, and et al. 2024. "Lectin-Based Immunophenotyping and Whole Proteomic Profiling of CT-26 Colon Carcinoma Murine Model" International Journal of Molecular Sciences 25, no. 7: 4022. https://doi.org/10.3390/ijms25074022
APA StyleFaragó, A., Zvara, Á., Tiszlavicz, L., Hunyadi-Gulyás, É., Darula, Z., Hegedűs, Z., Szabó, E., Surguta, S. E., Tóvári, J., Puskás, L. G., & Szebeni, G. J. (2024). Lectin-Based Immunophenotyping and Whole Proteomic Profiling of CT-26 Colon Carcinoma Murine Model. International Journal of Molecular Sciences, 25(7), 4022. https://doi.org/10.3390/ijms25074022