
Odorant Receptors Expressing and Antennal Lobes Architecture Are Linked to Caste Dimorphism in Asian Honeybee, Apis cerana (Hymenoptera: Apidae)
 and
and
Abstract
1. Introduction
2. Results
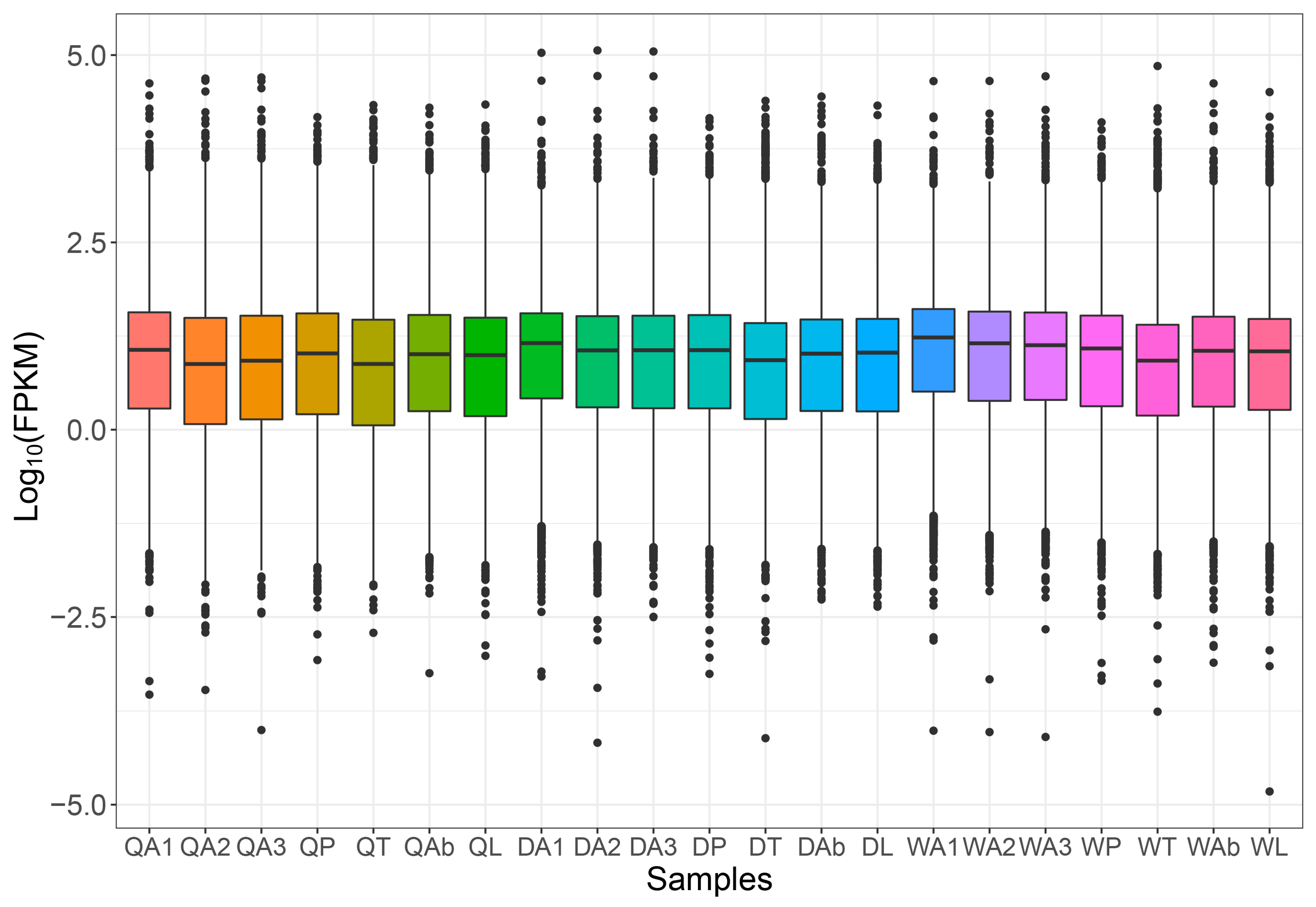
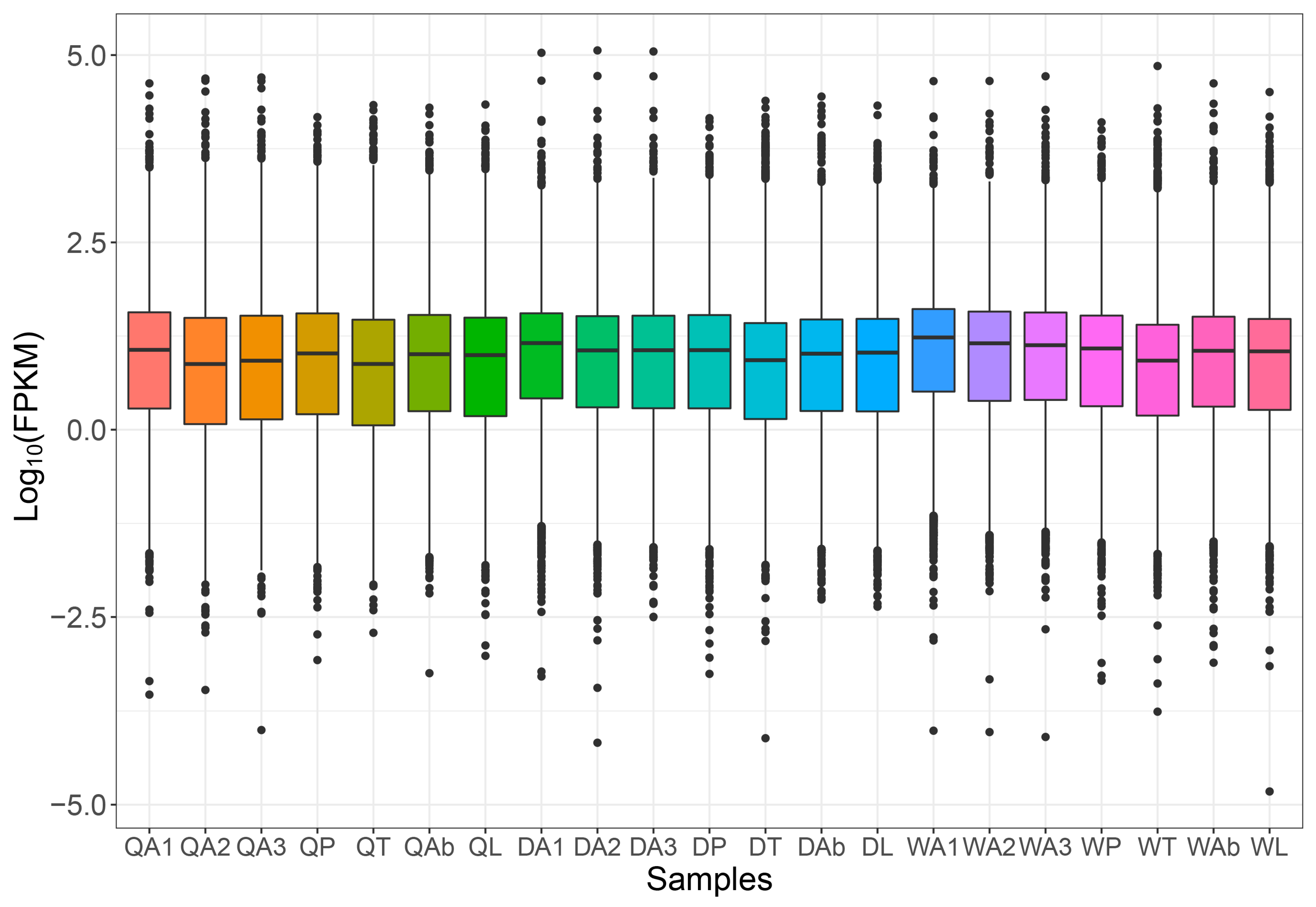
2.1. Overview of Transcriptome
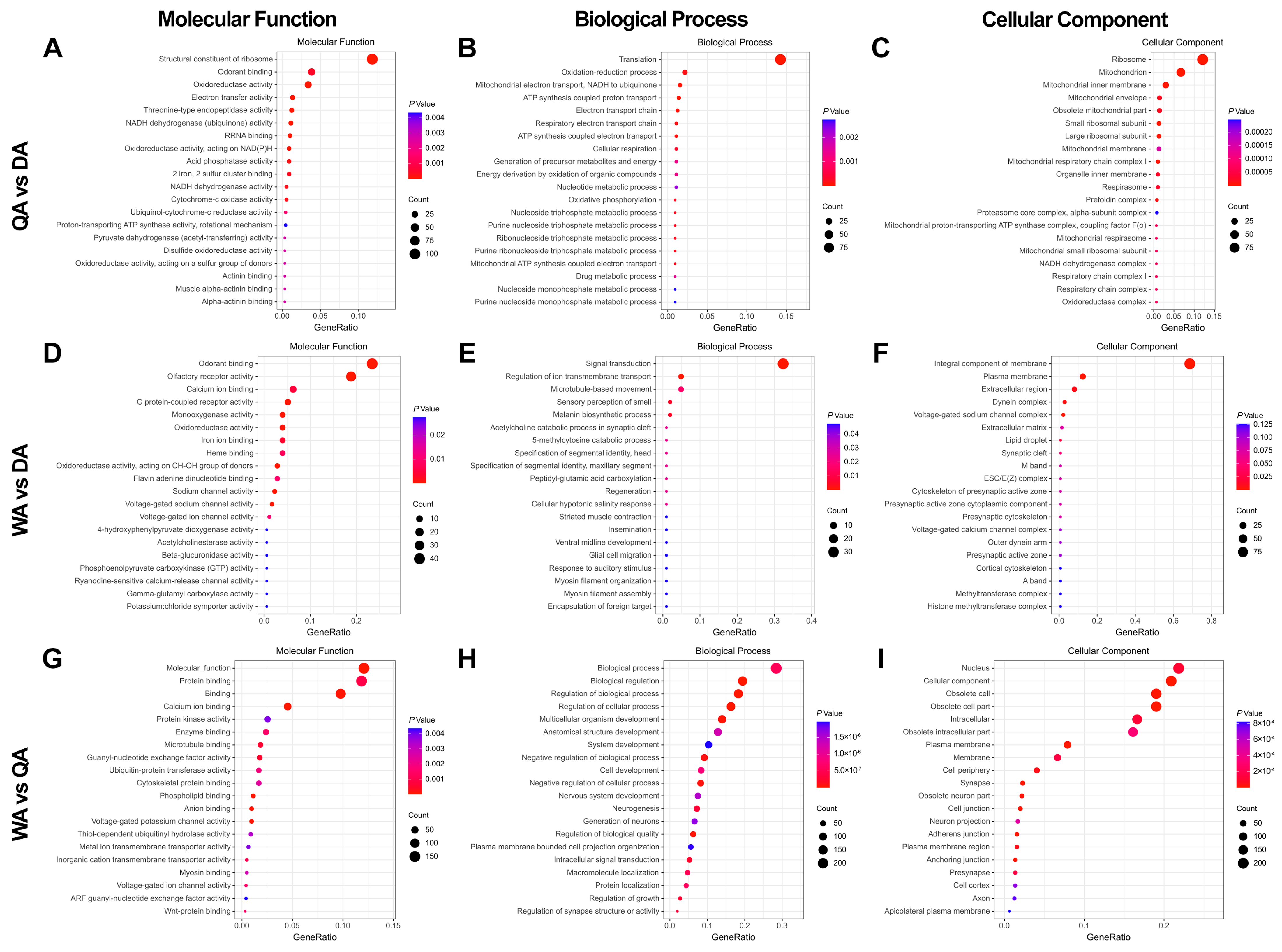
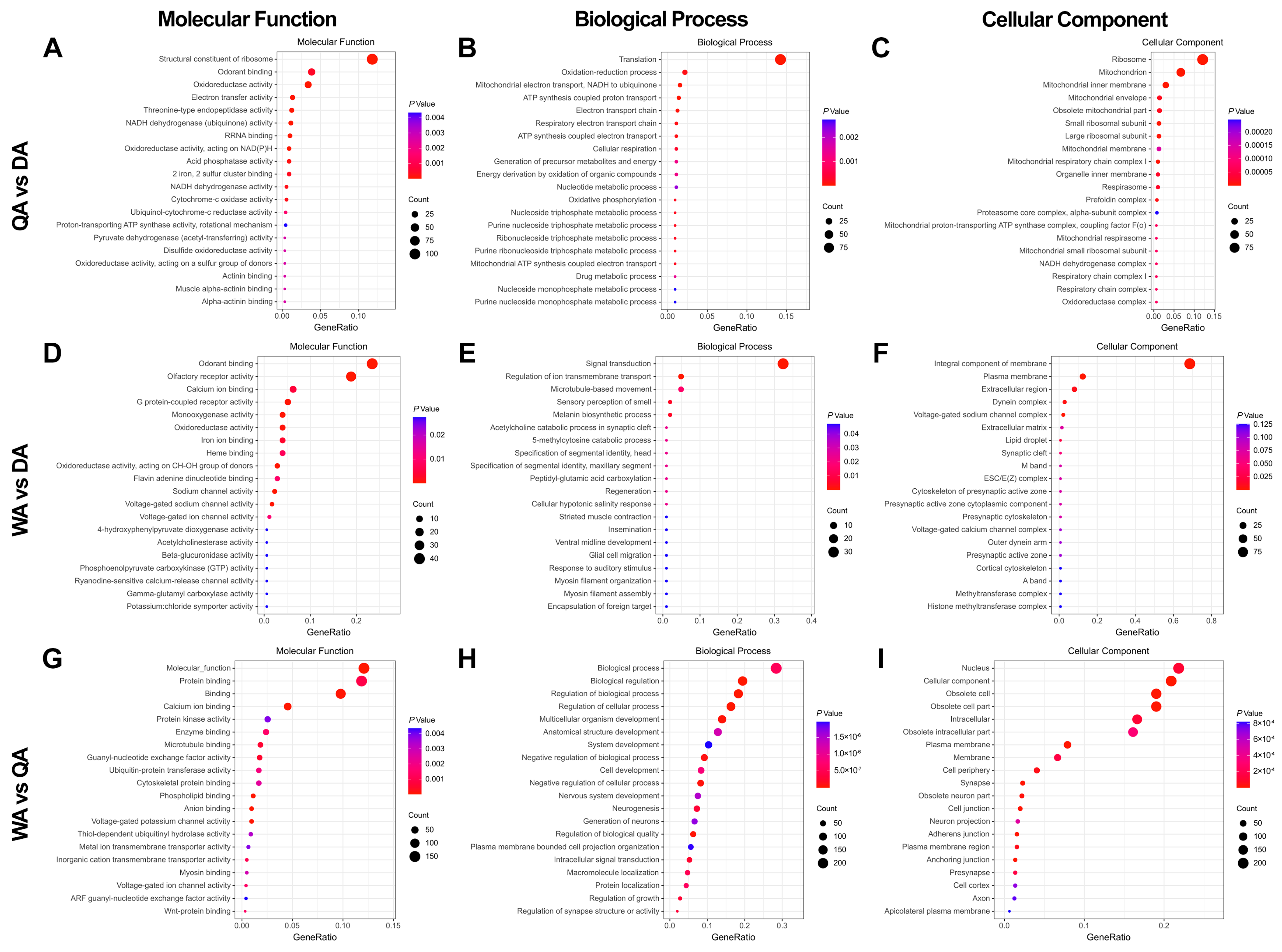
2.2. Differential Gene Analysis in Antenna
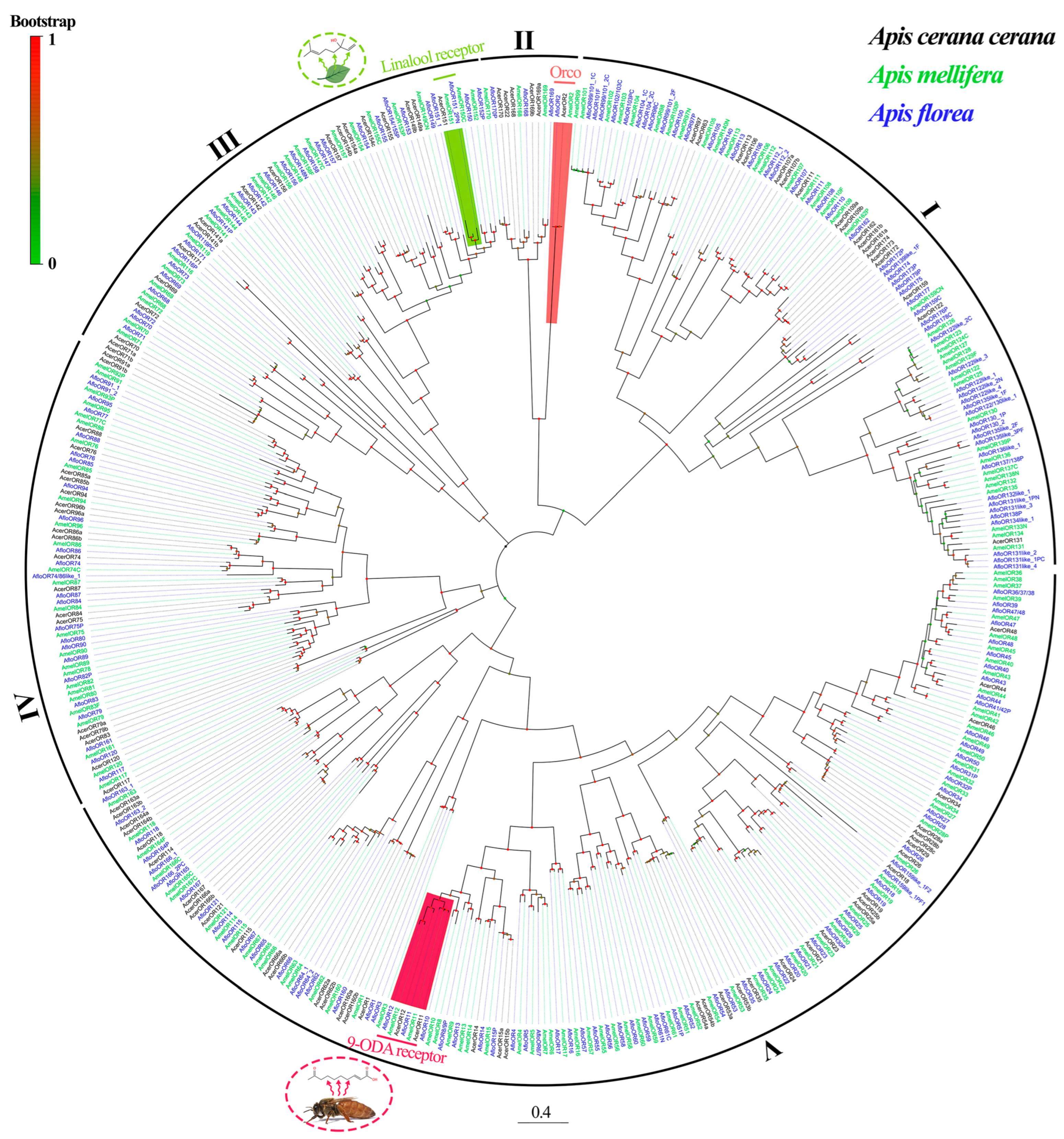
2.3. Identification and Homology Analysis of AcerORs
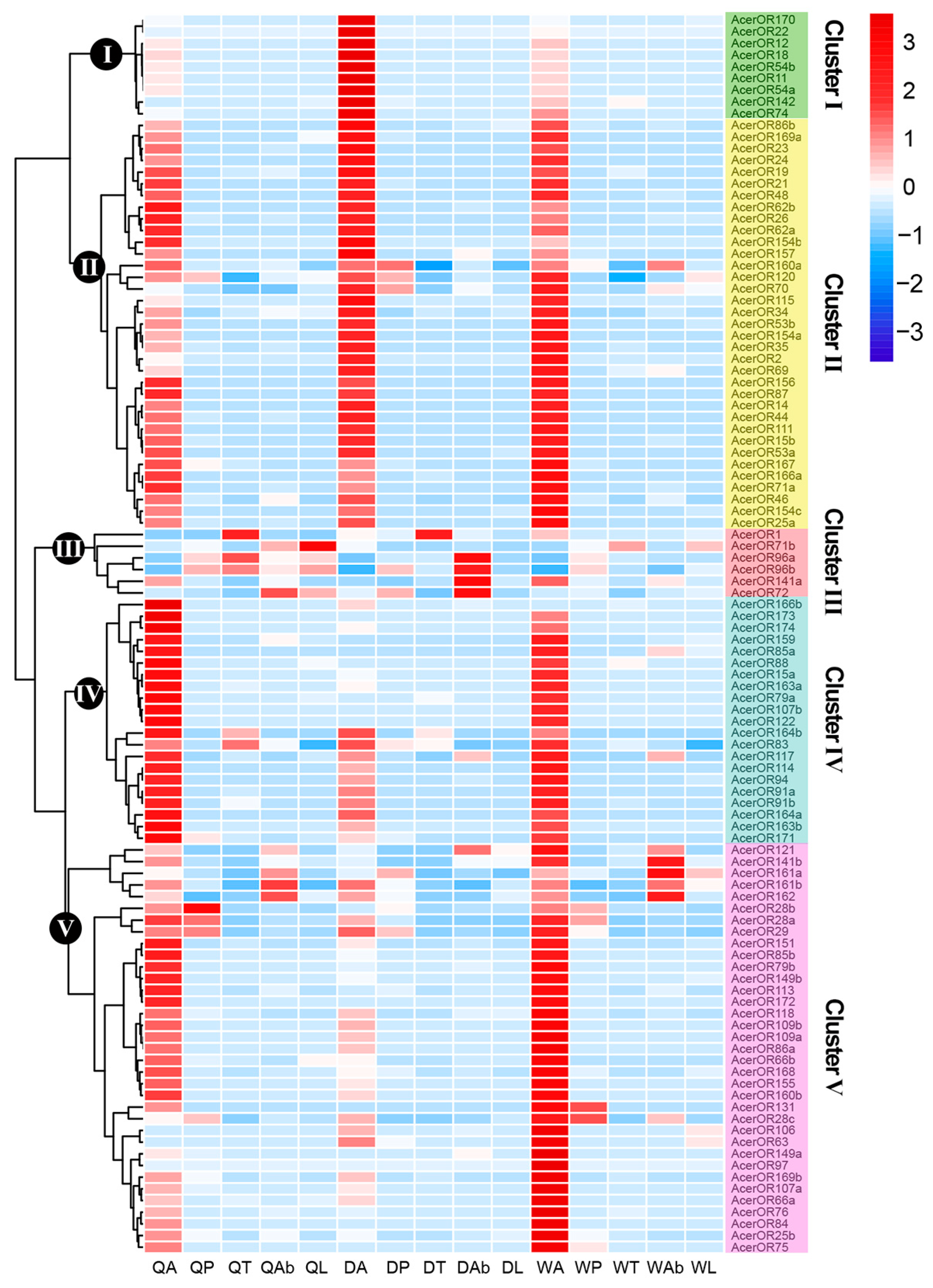
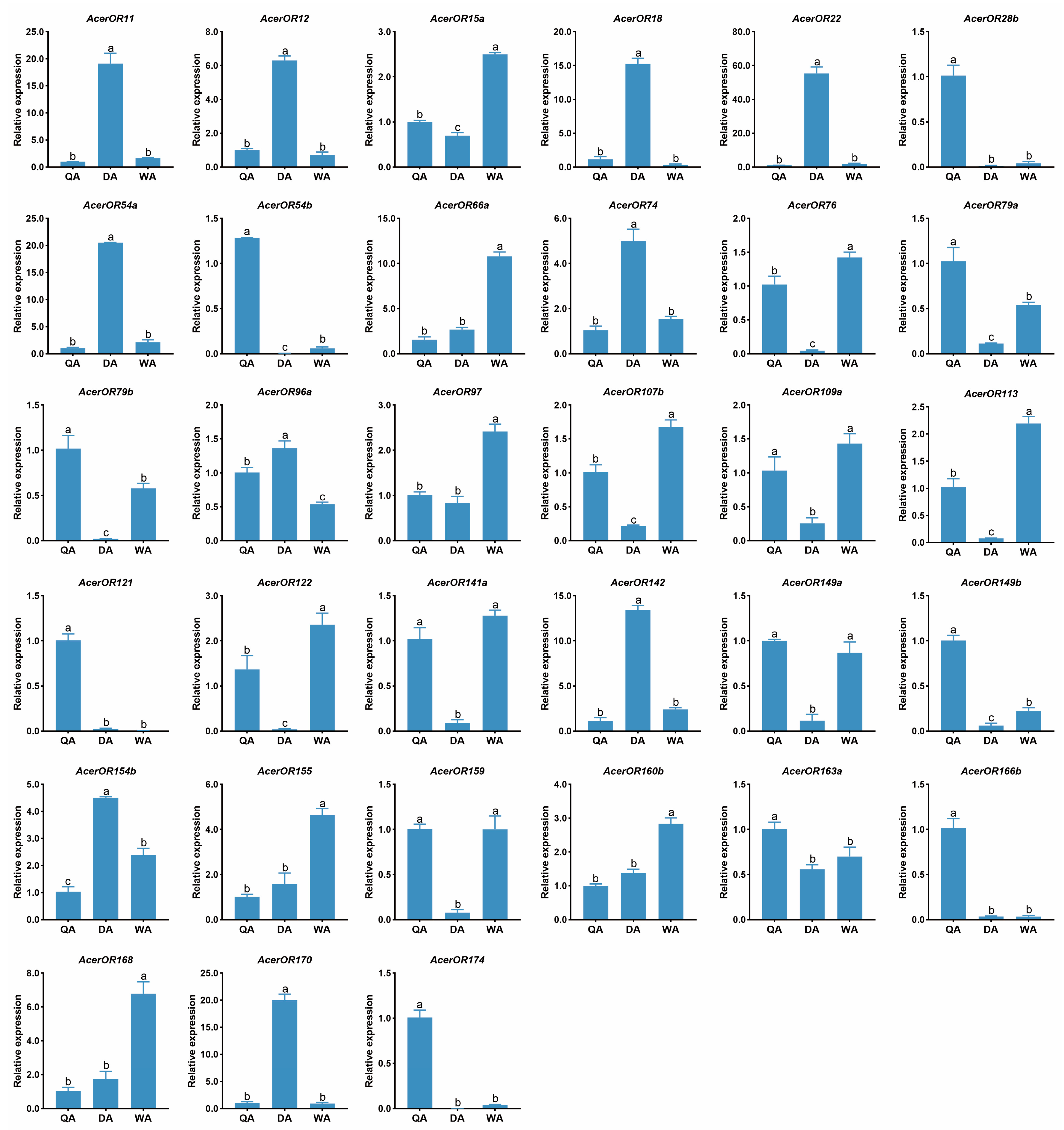
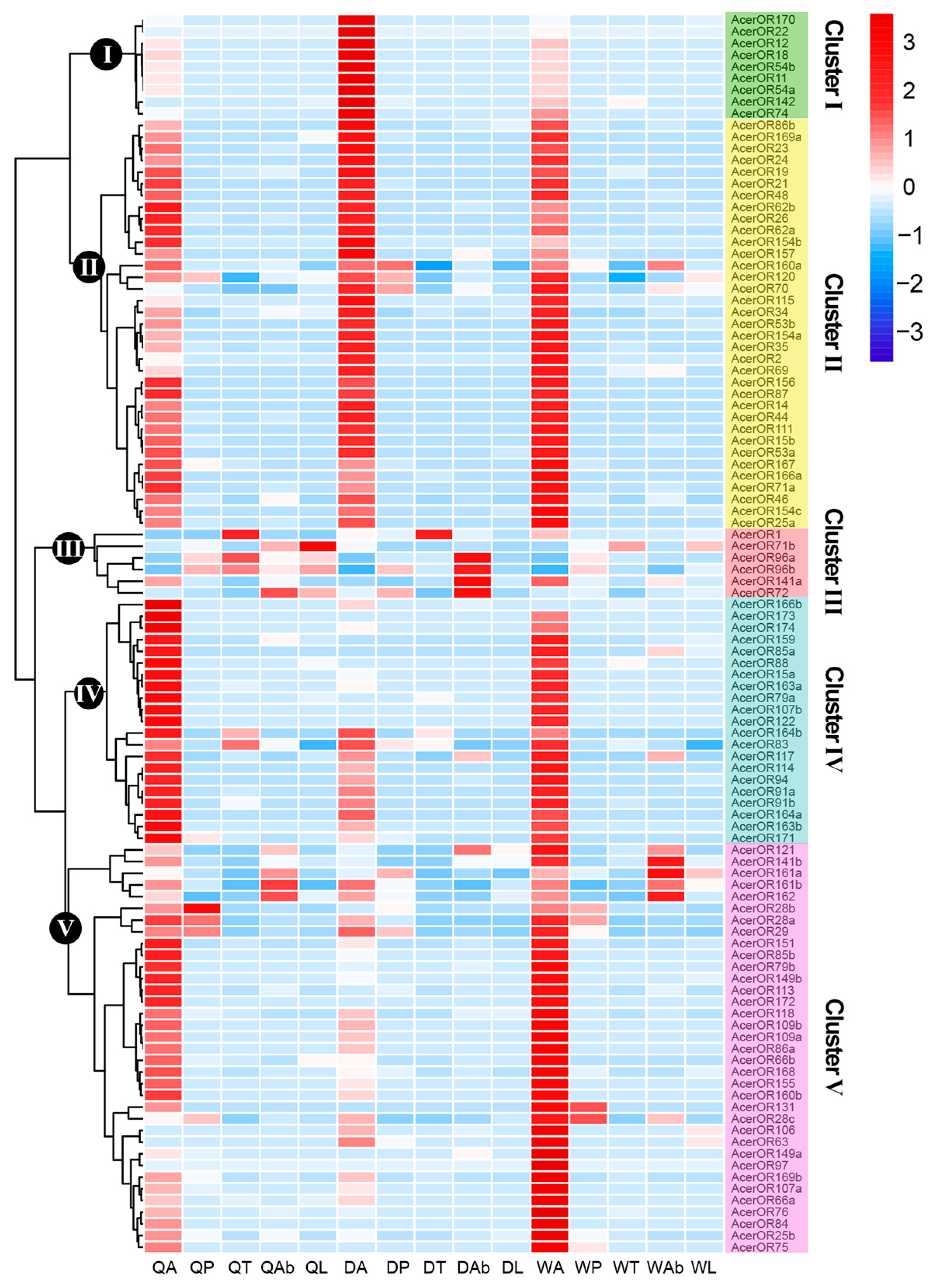
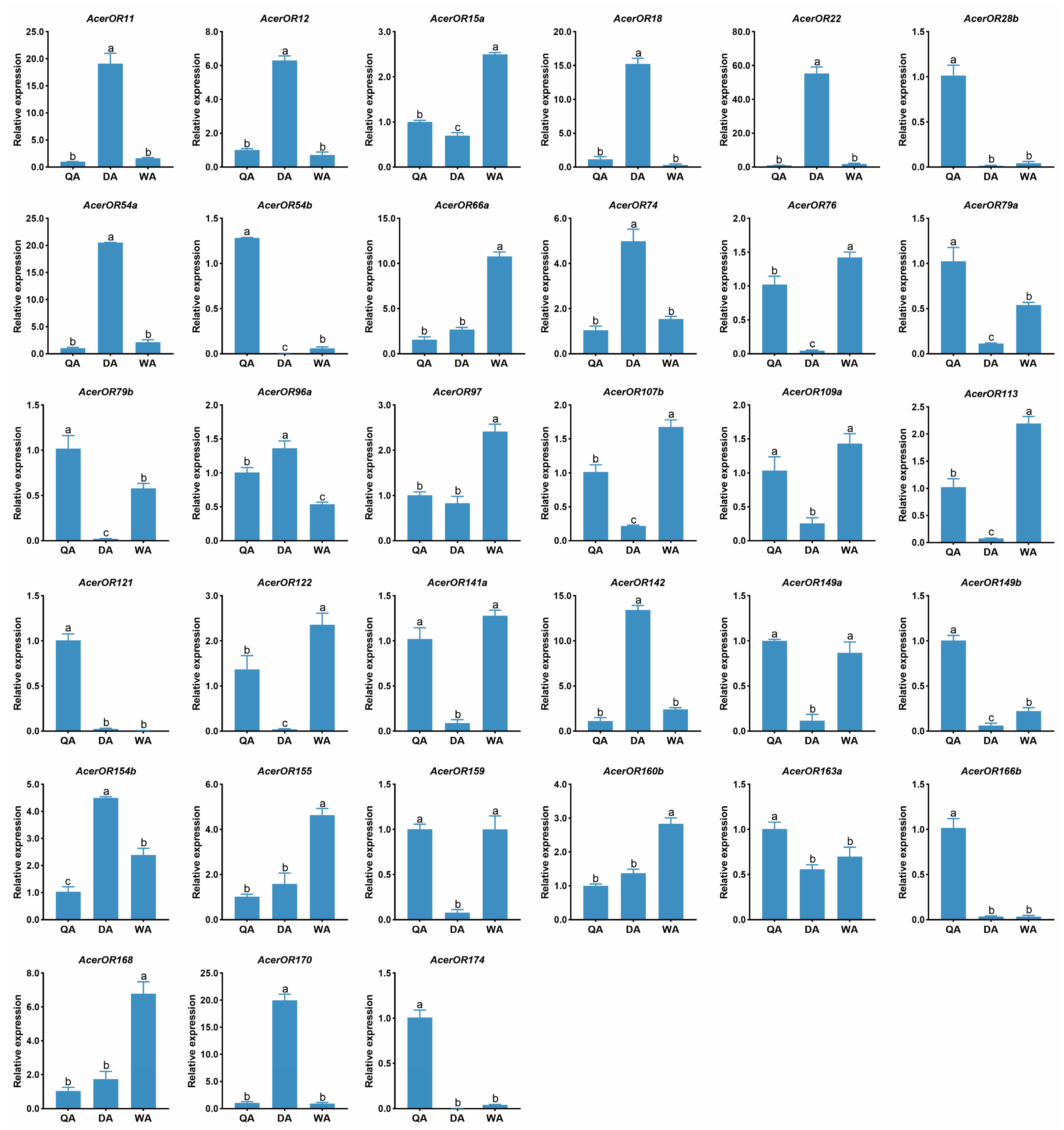
2.4. Expression Profiles of AcerORs
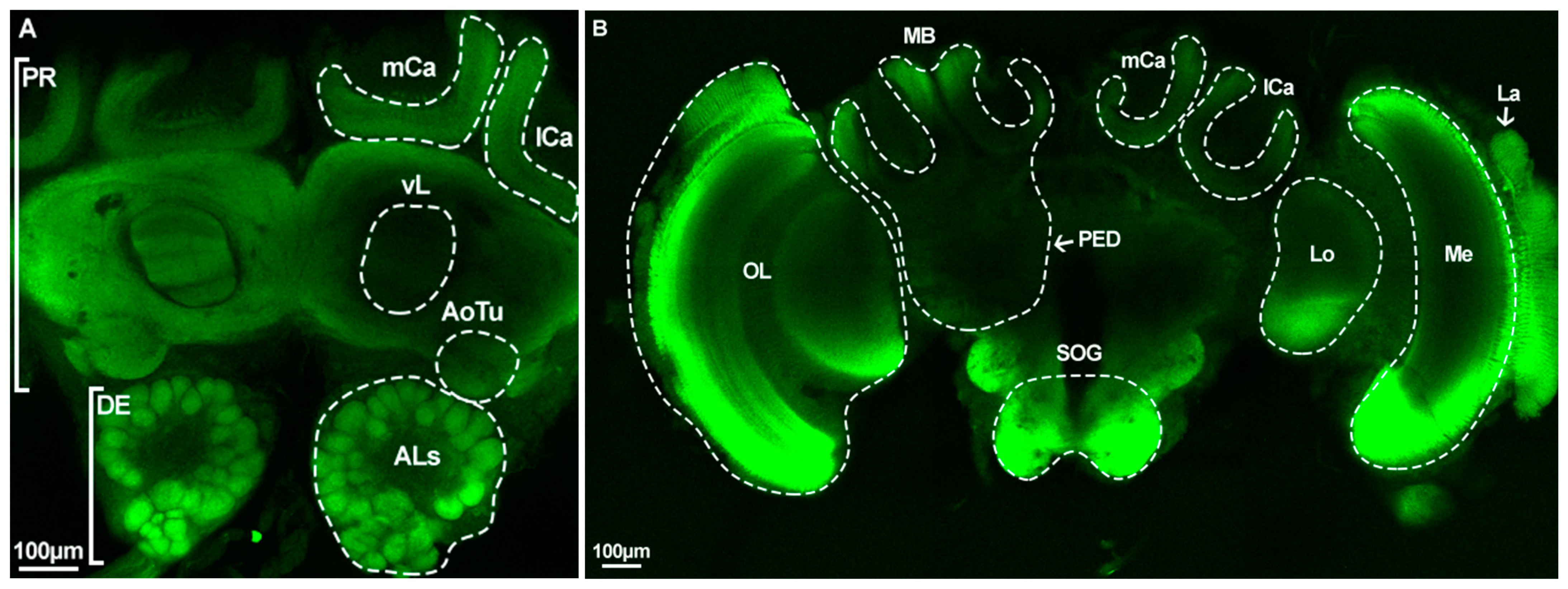
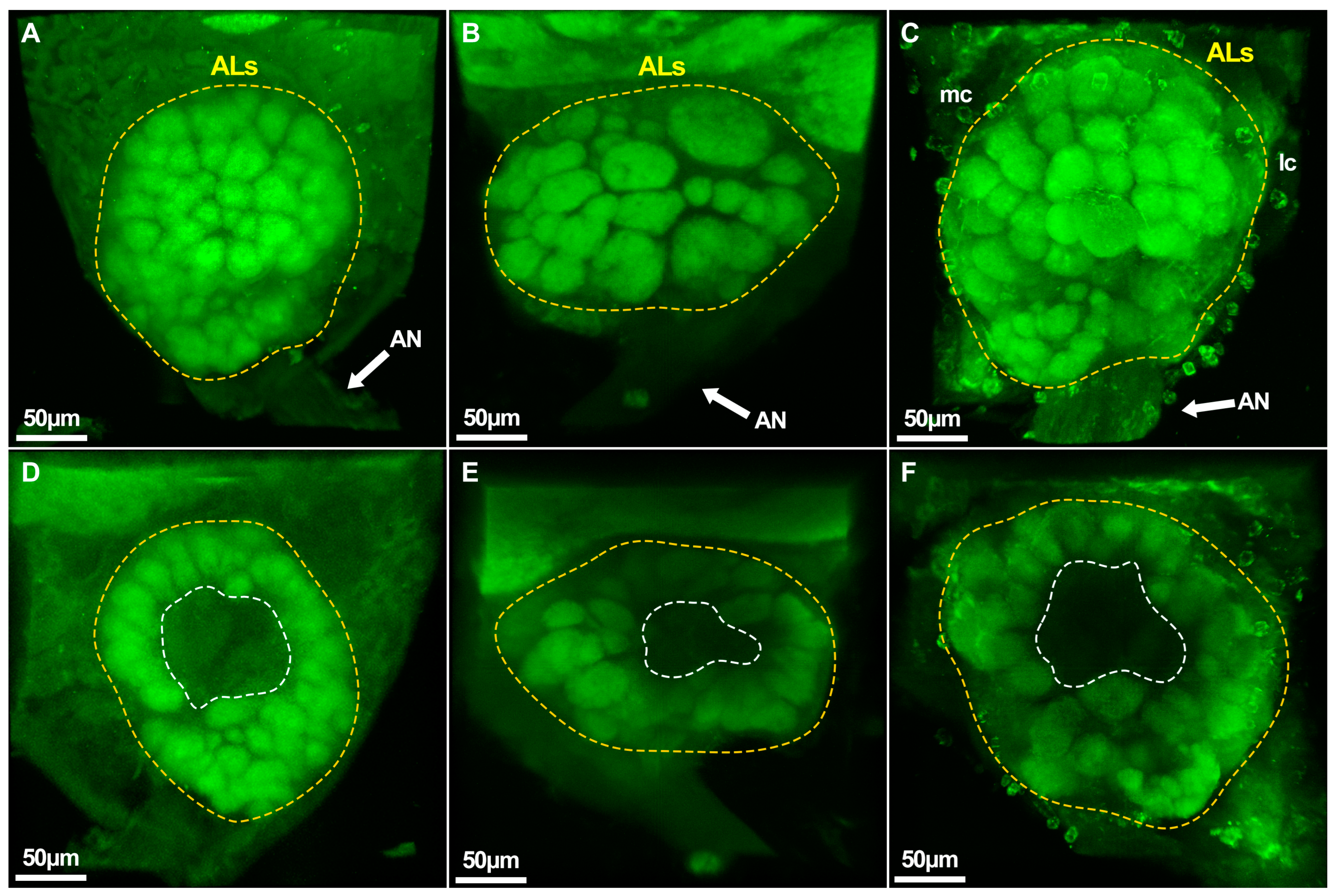
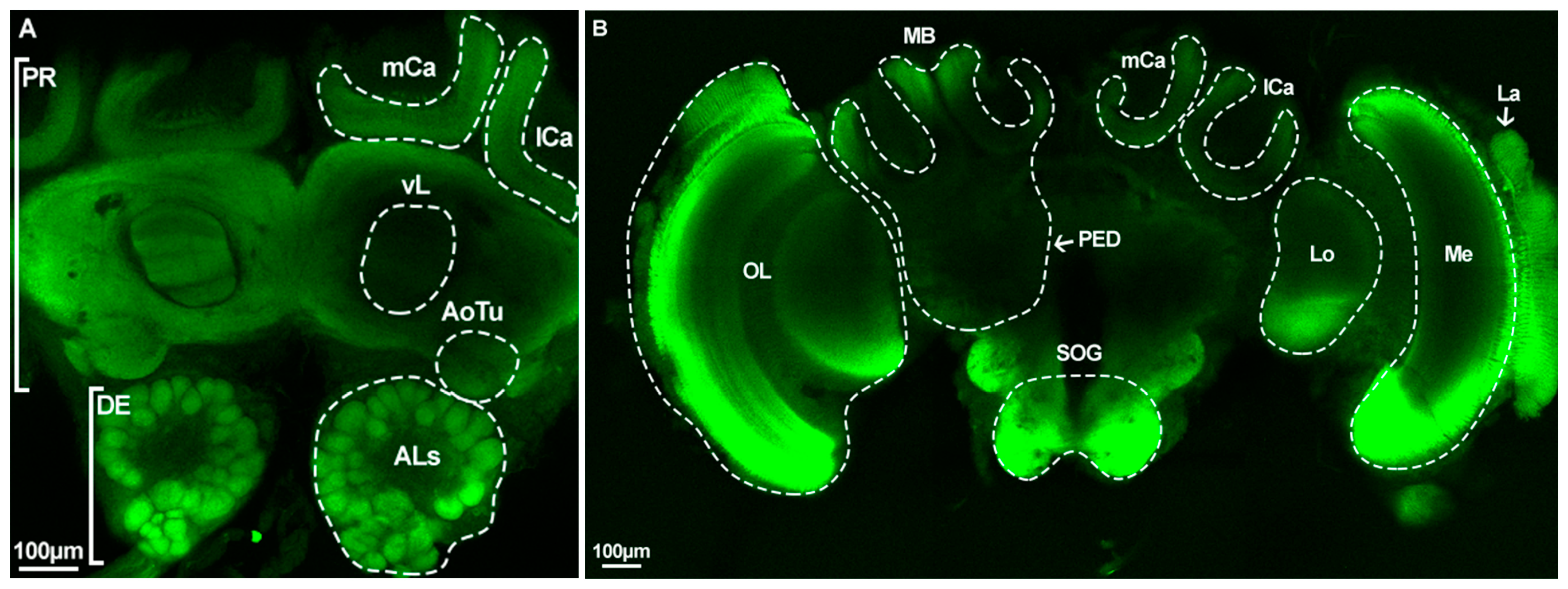
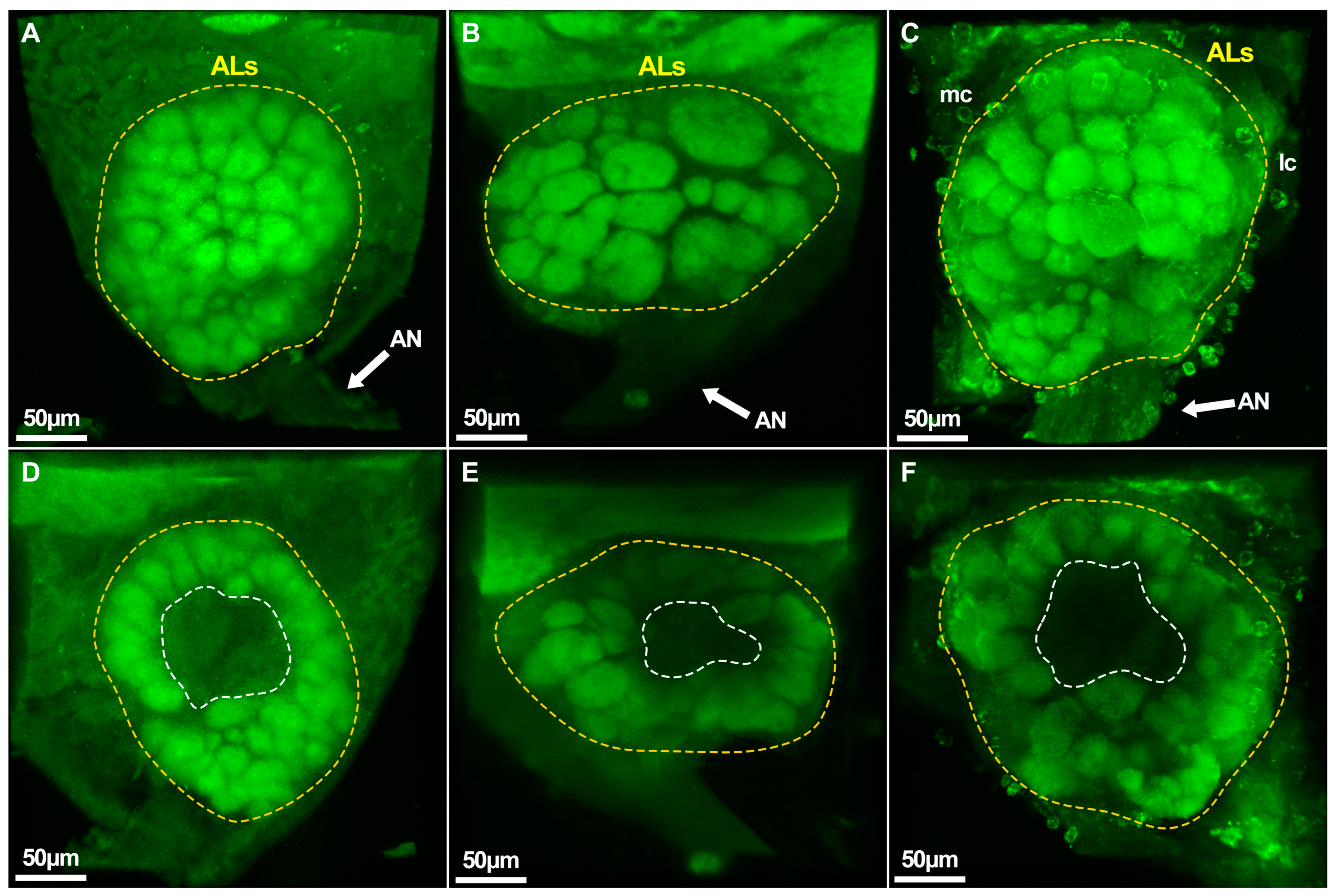
2.5. Caste-Specific Architecture of the Whole Brain and Antennal Lobes
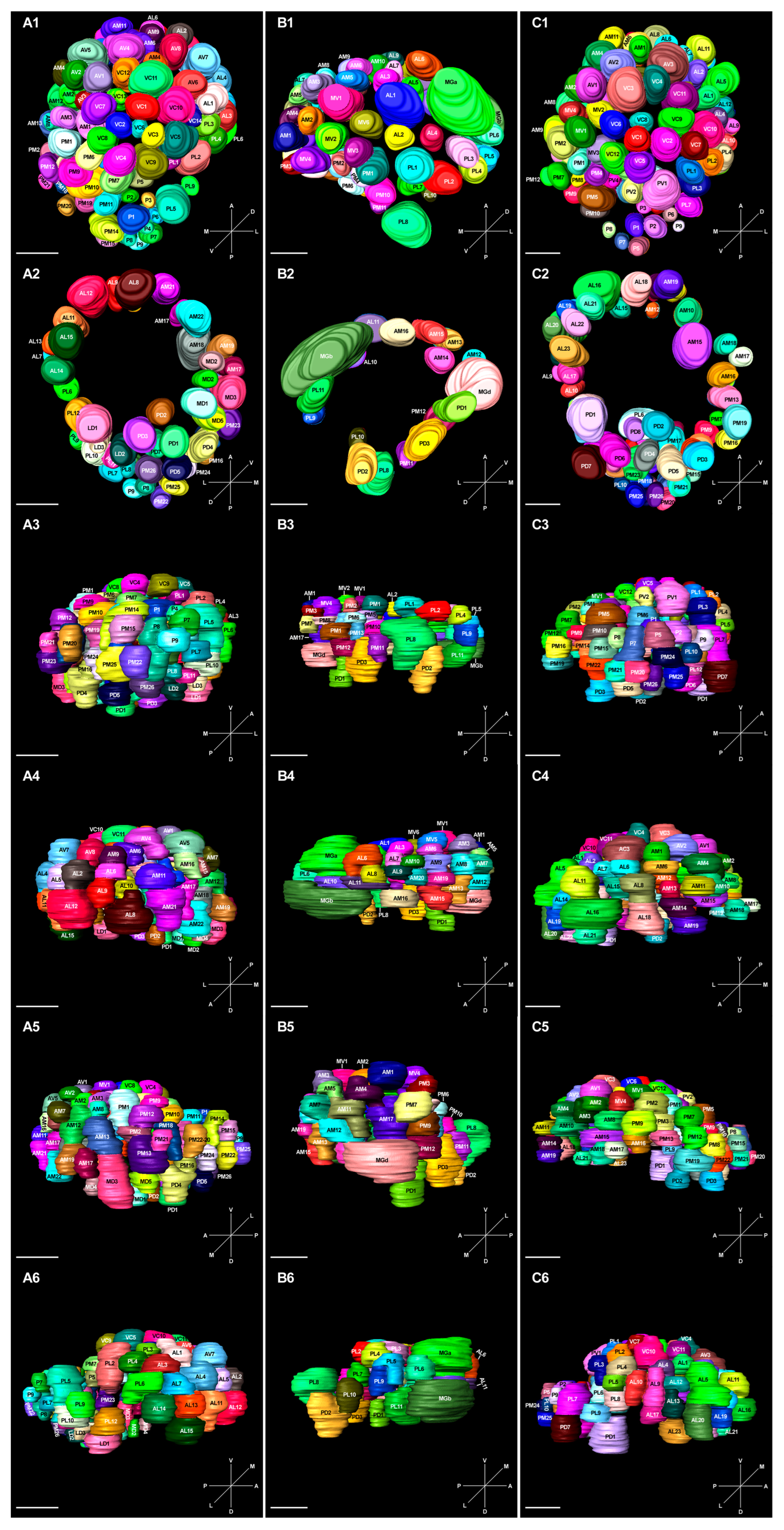
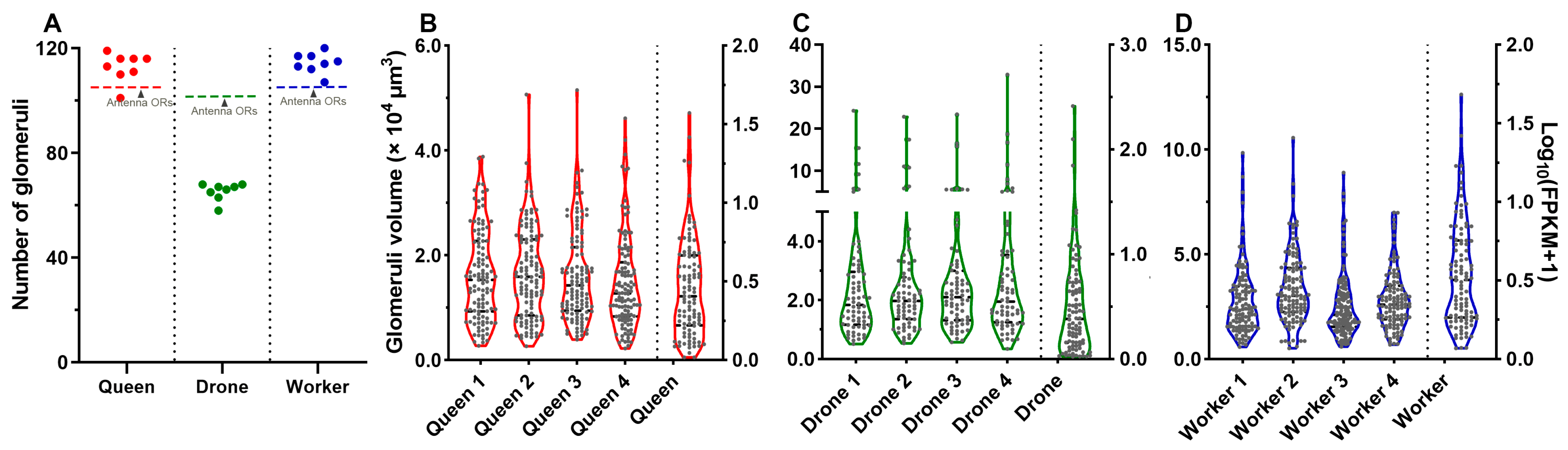
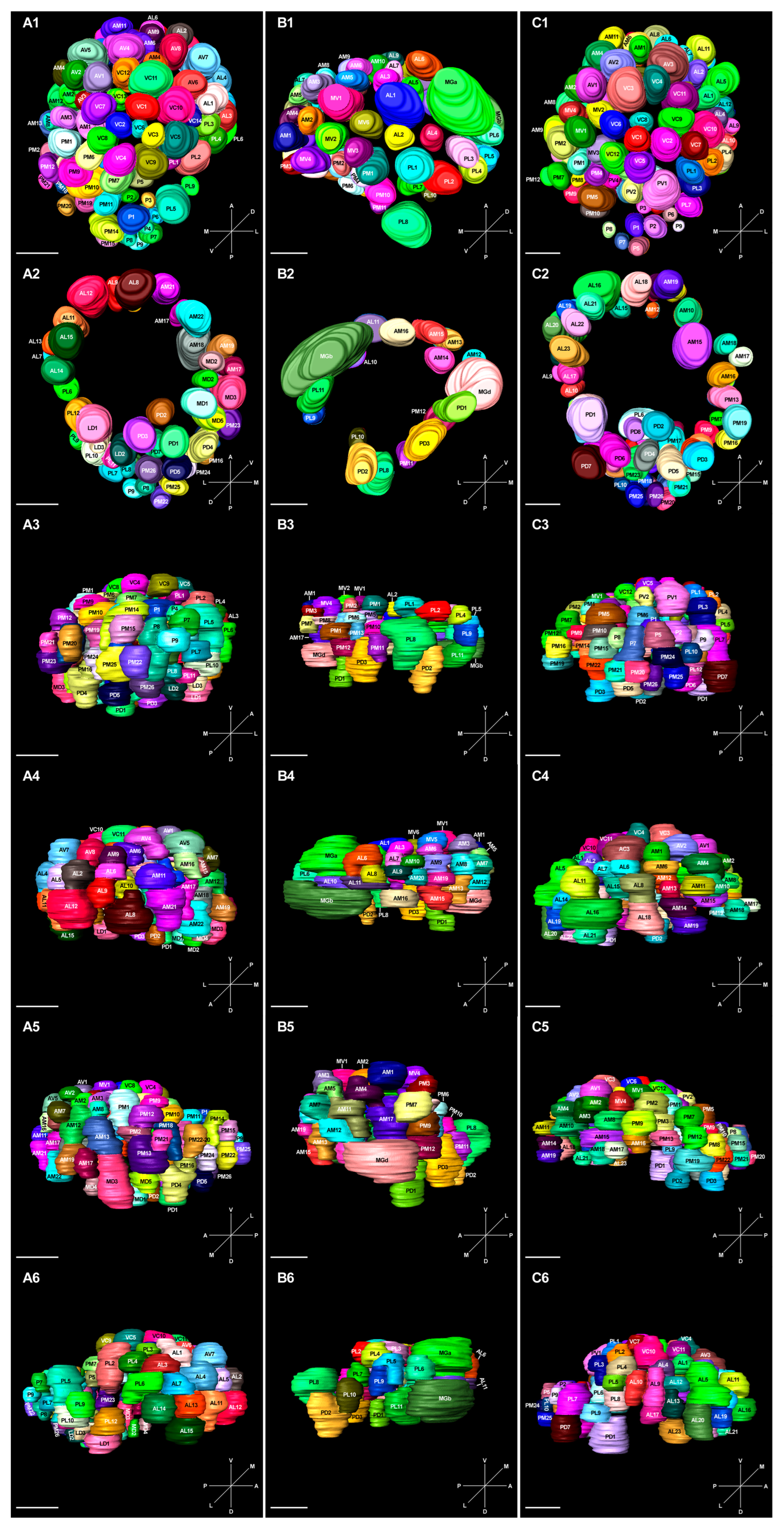
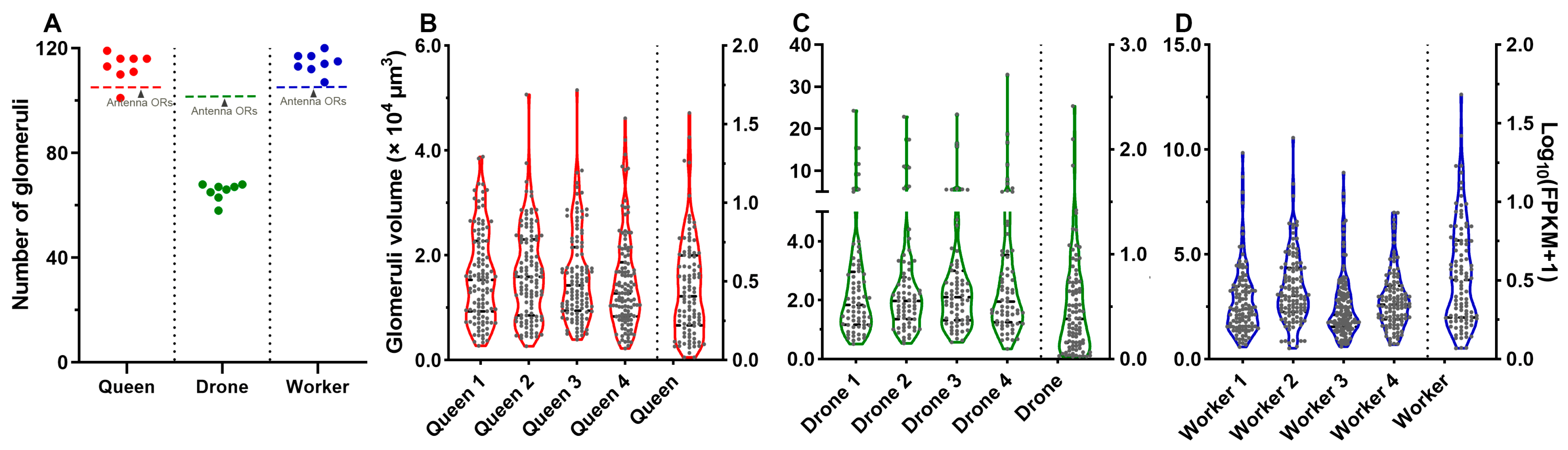
2.6. 3D Model and Characteristics of Glomeruli
2.7. Correlation between Glomeruli and ORs in A. cerana
3. Discussion
3.1. Caste Dimorphism Association with AcerORs
3.2. Caste Dimorphism Association with Als
3.3. Correlation between AcerORs and Glomeruli
4. Materials and Methods
4.1. Insects
4.2. RNA Sequencing and Differential Gene Expression Analysis
4.3. Sequence Homology and Expression Pattern Analysis of AcerORs
4.4. Real-Time Quantitative PCR (RT-qPCR) Validation of RNA-Seq Data
4.5. Immunohistochemical Staining
4.6. Confocal Laser Microscopy
4.7. 3D model Reconstruction
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bastin, F.; Chole, H.; Lafon, G.; Sandoz, J.C. Virgin queen attraction toward males in honey bees. Sci. Rep. 2017, 7, 6293. [Google Scholar] [CrossRef]
- Villar, G.; Wolfson, M.D.; Hefetz, A.; Grozinger, C.M. Evaluating the role of drone-produced chemical signals in mediating social interactions in honey bees (Apis mellifera). J. Chem. Ecol. 2018, 44, 1–8. [Google Scholar] [CrossRef]
- Paoli, M.; Galizia, G.C. Olfactory coding in honeybees. Cell Tissue Res. 2021, 383, 35–58. [Google Scholar] [CrossRef]
- Friedman, D.A.; Johnson, B.R.; Linksvayer, T.A. Distributed physiology and the molecular basis of social life in eusocial insects. Horm. Behav. 2020, 122, 104757. [Google Scholar] [CrossRef]
- Yan, H.; Liebig, J. Genetic basis of chemical communication in eusocial insects. Genes Dev. 2021, 35, 470–482. [Google Scholar] [CrossRef]
- Zhou, X.; Rokas, A.; Berger, S.L.; Liebig, J.; Ray, A.; Zwiebel, L.J. Chemoreceptor Evolution in Hymenoptera and Its Implications for the Evolution of Eusociality. Genome Biol. Evol. 2015, 7, 2407–2416. [Google Scholar] [CrossRef]
- Yin, X.; Iovinella, I.; Marangoni, R.; Cattonaro, F.; Flamini, G.; Sagona, S.; Zhang, L.; Pelosi, P.; Felicioli, A. Odorant-binding proteins and olfactory coding in the solitary bee Osmia cornuta. Cell Mol. Life Sci. 2013, 70, 3029–3039. [Google Scholar] [CrossRef]
- Godfrey, R.K.; Gronenberg, W. Brain evolution in social insects: Advocating for the comparative approach. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 2019, 205, 13–32. [Google Scholar] [CrossRef]
- McKenzie, S.K.; Fetter-Pruneda, I.; Ruta, V.; Kronauer, D.J. Transcriptomics and neuroanatomy of the clonal raider ant implicate an expanded clade of odorant receptors in chemical communication. Proc. Natl. Acad. Sci. USA 2016, 113, 14091–14096. [Google Scholar] [CrossRef]
- Robertson, H.M.; Wanner, K.W. The chemoreceptor superfamily in the honey bee, Apis mellifera: Expansion of the odorant, but not gustatory, receptor family. Genome Res. 2006, 16, 1395–1403. [Google Scholar] [CrossRef]
- Zhou, X.; Slone, J.D.; Rokas, A.; Berger, S.L.; Liebig, J.; Ray, A.; Reinberg, D.; Zwiebel, L.J. Phylogenetic and transcriptomic analysis of chemosensory receptors in a pair of divergent ant species reveals sex-specific signatures of odor coding. PLoS Genet. 2012, 8, e1002930. [Google Scholar] [CrossRef] [PubMed]
- Grob, R.; Heinig, N.; Grubel, K.; Rossler, W.; Fleischmann, P.N. Sex-specific and caste-specific brain adaptations related to spatial orientation in Cataglyphis ants. J. Comp. Neurol. 2021, 529, 3882–3892. [Google Scholar] [CrossRef] [PubMed]
- Brand, P.; Larcher, V.; Couto, A.; Sandoz, J.C.; Ramirez, S.R. Sexual dimorphism in visual and olfactory brain centers in the perfume-collecting orchid bee Euglossa dilemma (Hymenoptera, Apidae). J. Comp. Neurol. 2018, 526, 2068–2077. [Google Scholar] [CrossRef] [PubMed]
- Rozanski, A.N.; Cini, A.; Lopreto, T.E.; Gandia, K.M.; Hauber, M.E.; Cervo, R.; Uy, F.M.K. Differential investment in visual and olfactory brain regions is linked to the sensory needs of a wasp social parasite and its host. J. Comp. Neurol. 2022, 530, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Bastin, F.; Couto, A.; Larcher, V.; Phiancharoen, M.; Koeniger, G.; Koeniger, N.; Sandoz, J.C. Marked interspecific differences in the neuroanatomy of the male olfactory system of honey bees (genus Apis). J. Comp. Neurol. 2018, 526, 3020–3034. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Jung, J.W.; Choi, B.S.; Jayakodi, M.; Lee, J.; Lim, J.; Yu, Y.; Choi, Y.S.; Lee, M.L.; Park, Y.; et al. Uncovering the novel characteristics of Asian honey bee, Apis cerana, by whole genome sequencing. BMC Genom. 2015, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, G.; Guo, D.; Wang, Y.; Liu, Q.; Gao, Z.; Wang, H.; Liu, Z.; Guo, X.; Xu, B. Transcriptomic and metabolomic landscape of the molecular effects of glyphosate commercial formulation on Apis mellifera ligustica and Apis cerana cerana. Sci. Total Environ. 2020, 744, 140819. [Google Scholar] [CrossRef] [PubMed]
- Cremer, S. Social immunity in insects. Curr. Biol. 2019, 29, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.C.; Sheppard, W.S. Phylogenetic relationships of honey bees (Hymenoptera:Apinae:Apini) inferred from nuclear and mitochondrial DNA sequence data. Mol. Phylogenet. Evol. 2005, 37, 25–35. [Google Scholar] [CrossRef]
- Wang, Z.; Tan, K. Comparative analysis of olfactory learning of Apis cerana and Apis mellifera. Apidologie 2013, 45, 45–52. [Google Scholar] [CrossRef]
- Insolia, L.; Molinari, R.; Rogers, S.R.; Williams, G.R.; Chiaromonte, F.; Calovi, M. Honey bee colony loss linked to parasites, pesticides and extreme weather across the United States. Sci. Rep. 2023, 13, 1270. [Google Scholar] [CrossRef]
- Pettis, J.S.; vanEngelsdorp, D.; Johnson, J.; Dively, G. Pesticide exposure in honey bees results in increased levels of the gut pathogen Nosema. Naturwissenschaften 2012, 99, 153–158. [Google Scholar] [CrossRef]
- Yang, G. Protect and develop Apis cerana cerana to maintain ecological balance. Apic. China 2019, 70, 18–20. [Google Scholar]
- Obiero, G.F.; Pauli, T.; Geuverink, E.; Veenendaal, R.; Niehuis, O.; Grosse-Wilde, E. Chemoreceptor Diversity in Apoid Wasps and Its Reduction during the Evolution of the Pollen-Collecting Lifestyle of Bees (Hymenoptera: Apoidea). Genome Biol. Evol. 2021, 13, evaa269. [Google Scholar] [CrossRef]
- Karpe, S.D.; Dhingra, S.; Brockmann, A.; Sowdhamini, R. Computational genome-wide survey of odorant receptors from two solitary bees Dufourea novaeangliae (Hymenoptera: Halictidae) and Habropoda laboriosa (Hymenoptera: Apidae). Sci. Rep. 2017, 7, 10823. [Google Scholar] [CrossRef]
- Wang, Z.L.; Zhu, Y.Q.; Yan, Q.; Yan, W.Y.; Zheng, H.J.; Zeng, Z.J. A Chromosome-Scale Assembly of the Asian Honeybee Apis cerana Genome. Front. Genet. 2020, 11, 279. [Google Scholar] [CrossRef]
- Guo, L.; Zhao, H.; Xu, B.; Jiang, Y. Odorant receptor might be related to sperm DNA integrity in Apis cerana cerana. Anim. Reprod. Sci. 2018, 193, 33–39. [Google Scholar] [CrossRef]
- Pitts, R.J.; Liu, C.; Zhou, X.; Malpartida, J.C.; Zwiebel, L.J. Odorant receptor-mediated sperm activation in disease vector mosquitoes. Proc. Natl. Acad. Sci. USA 2014, 111, 2566–2571. [Google Scholar] [CrossRef]
- Claudianos, C.; Lim, J.; Young, M.; Yan, S.; Cristino, A.S.; Newcomb, R.D.; Gunasekaran, N.; Reinhard, J. Odor memories regulate olfactory receptor expression in the sensory periphery. Eur. J. Neurosci. 2014, 39, 1642–1654. [Google Scholar] [CrossRef]
- Arnold, G.; Masson, C.; Budharugsa, S. Comparative study of the antennal lobes and their afferent pathway in the worker bee and the drone (Apis mellifera). Cell Tissue Res. 1985, 242, 593–605. [Google Scholar] [CrossRef]
- Anton, S.; Rossler, W. Plasticity and modulation of olfactory circuits in insects. Cell Tissue Res. 2021, 383, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Groh, C.; Rossler, W. Caste-specific postembryonic development of primary and secondary olfactory centers in the female honeybee brain. Arthropod Struct. Dev. 2008, 37, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Streinzer, M.; Kelber, C.; Pfabigan, S.; Kleineidam, C.J.; Spaethe, J. Sexual dimorphism in the olfactory system of a solitary and a eusocial bee species. J. Comp. Neurol. 2013, 521, 2742–2755. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, S.C.; Liebig, J.; Rössler, W. Biogenic amines in the ponerine ant Harpegnathos saltator: Serotonin and dopamine immunoreactivity in the brain. Arthropod Struct. Dev. 2005, 34, 429–440. [Google Scholar] [CrossRef]
- Sandoz, J.C. Odour-evoked responses to queen pheromone components and to plant odours using optical imaging in the antennal lobe of the honey bee drone Apis mellifera L. J. Exp. Biol. 2006, 209, 3587–3598. [Google Scholar] [CrossRef] [PubMed]
- Keesey, I.W.; Zhang, J.; Depetris-Chauvin, A.; Obiero, G.F.; Gupta, A.; Gupta, N.; Vogel, H.; Knaden, M.; Hansson, B.S. Functional olfactory evolution in Drosophila suzukii and the subgenus Sophophora. iScience 2022, 25, 104212. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A.L.; Van der Goes van Naters, W.; Lessing, D.; Warr, C.G.; Carlson, J.R. Coexpression of two functional odor receptors in one neuron. Neuron 2005, 45, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Riabinina, O.; Task, D.; Marr, E.; Lin, C.C.; Alford, R.; O’Brochta, D.A.; Potter, C.J. Organization of olfactory centres in the malaria mosquito Anopheles gambiae. Nat. Commun. 2016, 7, 13010. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Liu, F.; Liu, N. Three-dimensional structure of the antennal lobe in the Southern house mosquito Culex quinquefasciatus. Insect Sci. 2021, 28, 93–102. [Google Scholar] [CrossRef]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef]
- Herre, M.; Goldman, O.V.; Lu, T.C.; Caballero-Vidal, G.; Qi, Y.; Gilbert, Z.N.; Gong, Z.; Morita, T.; Rahiel, S.; Ghaninia, M.; et al. Non-canonical odor coding in the mosquito. Cell 2022, 185, 3104–3123.e28. [Google Scholar] [CrossRef]
- Wanner, K.W.; Nichols, A.S.; Walden, K.K.; Brockmann, A.; Luetje, C.W.; Robertson, H.M. A honey bee odorant receptor for the queen substance 9-oxo-2-decenoic acid. Proc. Natl. Acad. Sci. USA 2007, 104, 14383–14388. [Google Scholar] [CrossRef]
- Karpe, S.D.; Jain, R.; Brockmann, A.; Sowdhamini, R. Identification of Complete Repertoire of Apis florea Odorant Receptors Reveals Complex Orthologous Relationships with Apis mellifera. Genome Biol. Evol. 2016, 8, 2879–2895. [Google Scholar] [CrossRef]
- Kazawa, T.; Namiki, S.; Fukushima, R.; Terada, M.; Soo, K.; Kanzaki, R. Constancy and variability of glomerular organization in the antennal lobe of the silkmoth. Cell Tissue Res. 2009, 336, 119–136. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Total DEGs | Upregulated Genes | Downregulated Genes |
|---|---|---|---|
| QA vs. DA | 3252 | 1594 | 1658 |
| WA vs. DA | 568 | 317 | 251 |
| WA vs. QA | 3866 | 2100 | 1766 |
| Caste | Total | Antenna | Proboscis | Thorax | Abdomen | Legs |
|---|---|---|---|---|---|---|
| Queen | 106 | 106 | 46 | 40 | 60 | 58 |
| Drone | 101 | 99 | 46 | 44 | 58 | 41 |
| Worker | 106 | 106 | 55 | 36 | 63 | 63 |
| Queens | Drones | Workers | |
|---|---|---|---|
| Total number of glomeruli | 116 ± 2.9 a | 68 ± 0.5 b | 117 ± 2.3 a |
| Total volume of ALs (×106 μm3) | 1.82 ± 0.06 b | 2.04 ± 0.14 b | 3.34 ± 0.35 a |
| Total surface area of ALs (×105 μm2) | 4.01 ± 0.09 b | 3.74 ± 0.08 b | 6.07 ± 0.43 a |
| AL type | 15 | 11 | 24 |
| AM type | 22 | 20 | 19 |
| AV type | 8 | - | 3 |
| MV type | - | 7 | 4 |
| LD type | 3 | - | - |
| MD type | 5 | - | - |
| PD type | 7 | 3 | 8 |
| P type | 9 | - | 9 |
| PL type | 14 | 16 | 17 |
| PM type | 28 | 14 | 26 |
| PV type | - | - | 4 |
| MG/G type | - | 4 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ke, H.; Chen, Y.; Zhang, B.; Duan, S.; Ma, X.; Ren, B.; Wang, Y. Odorant Receptors Expressing and Antennal Lobes Architecture Are Linked to Caste Dimorphism in Asian Honeybee, Apis cerana (Hymenoptera: Apidae). Int. J. Mol. Sci. 2024, 25, 3934. https://doi.org/10.3390/ijms25073934
Ke H, Chen Y, Zhang B, Duan S, Ma X, Ren B, Wang Y. Odorant Receptors Expressing and Antennal Lobes Architecture Are Linked to Caste Dimorphism in Asian Honeybee, Apis cerana (Hymenoptera: Apidae). International Journal of Molecular Sciences. 2024; 25(7):3934. https://doi.org/10.3390/ijms25073934
Chicago/Turabian StyleKe, Haoqin, Yu Chen, Baoyi Zhang, Shiwen Duan, Xiaomei Ma, Bingzhong Ren, and Yinliang Wang. 2024. "Odorant Receptors Expressing and Antennal Lobes Architecture Are Linked to Caste Dimorphism in Asian Honeybee, Apis cerana (Hymenoptera: Apidae)" International Journal of Molecular Sciences 25, no. 7: 3934. https://doi.org/10.3390/ijms25073934
APA StyleKe, H., Chen, Y., Zhang, B., Duan, S., Ma, X., Ren, B., & Wang, Y. (2024). Odorant Receptors Expressing and Antennal Lobes Architecture Are Linked to Caste Dimorphism in Asian Honeybee, Apis cerana (Hymenoptera: Apidae). International Journal of Molecular Sciences, 25(7), 3934. https://doi.org/10.3390/ijms25073934