
Unveiling the Hidden Regulators: The Impact of lncRNAs on Zoonoses



Abstract
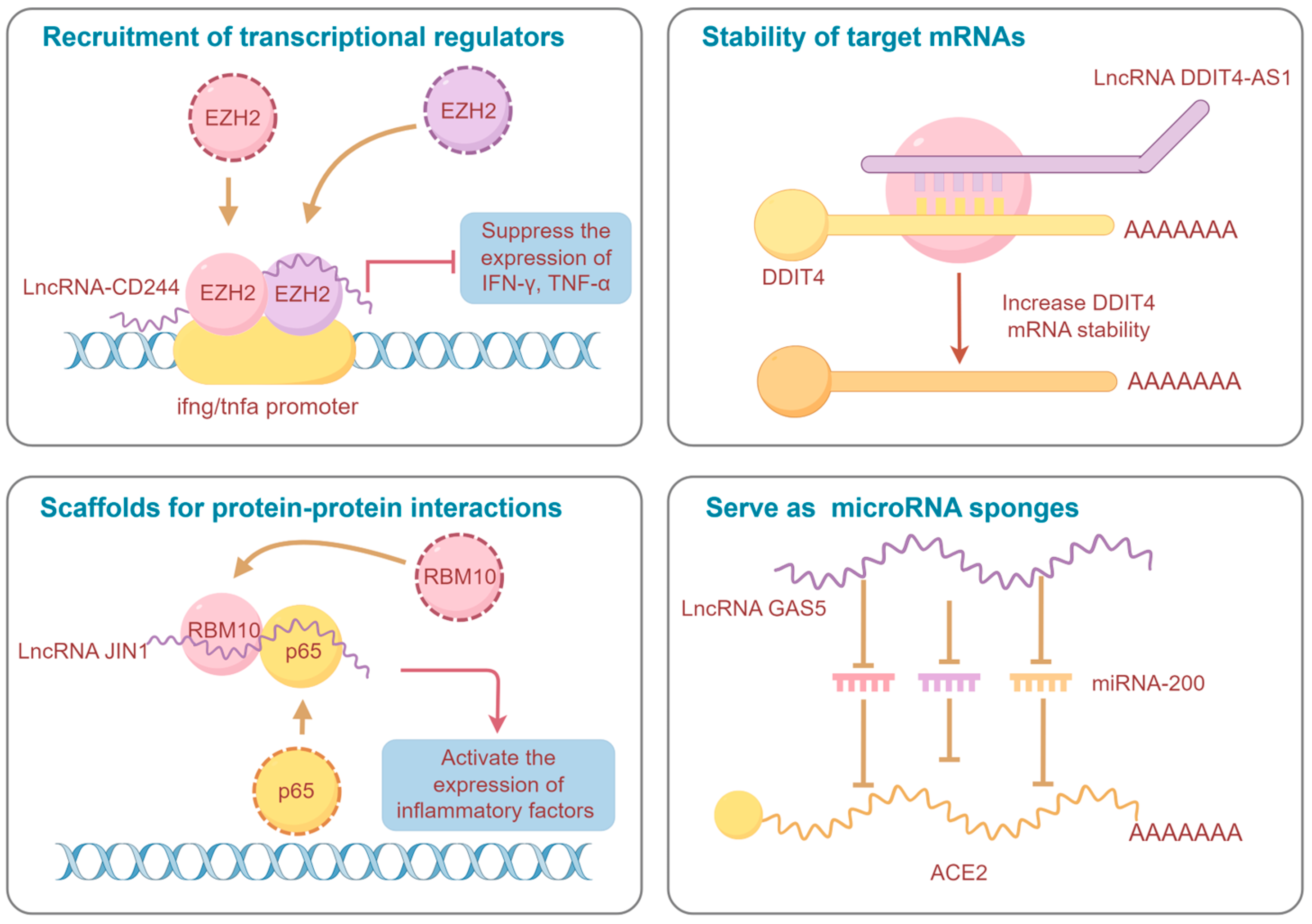
1. Introduction
2. LncRNAs in Bacterial Zoonoses
2.1. Tuberculosis
2.2. Colibacillosis
2.3. Brucellosis
2.4. Salmonellosis
2.5. Pseudomonas aeruginosa Infection
2.6. Listeriosis
2.7. Staphylococcosis

{kind=link}
{kind=link}
| Pathogen | LncRNA | Category | Function or Mechanism | Reference |
|---|---|---|---|---|
| Mtb | lncRNA-CD244 | Host antisense lncRNA | Regulate T-cell responses against TB infection | [36] |
| NEAT1 | Host intergenic lncRNA | Regulate the inflammatory responses in macrophages | [37,93] | |
| XLOC_012582 | Host intergenic lncRNA | Regulate the expression of SOCS3 | [38] | |
| PCED1B-AS1 | Host antisense lncRNA | Modulate macrophage apoptosis and autophagy by targeting miR-155 | [39] | |
| lincRNA-EPS | Host intergenic lncRNA | Regulate apoptosis and autophagy of macrophages via JNK/MAPK signaling | [94] | |
| lincRNA-Cox2 | Host intergenic lncRNA | Regulate macrophage apoptosis | [95] | |
| lnc-EST12 | Host intergenic lncRNA | Regulate anti-Mtb innate immunity through FUBP3 | [40] | |
| DANCR | Host intergenic lncRNA | Restrain intracellular survival of Mtb via miR-1301-3p and miR-5194 | [41] | |
| XIST | Host intergenic lncRNA | Promote the polarization of macrophages to the M1 phenotype via miR-125b-5p/A20/NF-κB axis | [42] | |
| MIAT | Host intergenic lncRNA | Regulate autophagy and antimicrobial responses | [43] | |
| MIR99AHG | Host intergenic lncRNA | Promote Mtb growth by regulating inflammation and macrophage polarization | [44] | |
| LINC00870 | Host intergenic lncRNA | Biomarker | [45] | |
| CCAT1 | Host intergenic lncRNA | Biomarker | [45] | |
| LOC152742 | Host intergenic lncRNA | Biomarker | [45] | |
| MIR3945HG | Host intergenic lncRNA | Biomarker | [46] | |
| E. coli | FUT3-AS1 | Host antisense lncRNA | Modulates E. coli susceptibility via histone H4 modifications | [49] |
| lncRSPH9-4 | Host sense overlapping lncRNA | Disrupt endothelial barrier via miR-17-5p/MMP3 axis | [52] | |
| lncC11orf54-1 | Host intronic lncRNA | Modulate neuroinflammation responses | [51] | |
| DDIT-AS1 | Host antisense lncRNA | Modulate DDIT4 expression and promote neuroinflammation responses | [53] | |
| XIST | Host intergenic lncRNA | Regulate NF-κB/NLRP3 inflammasome pathway | [54] | |
| HOTAIR | Host antisense lncRNA | Promote kidney injury in sepsis | [55] | |
| SOX2OT | Host sense overlapping lncRNA | Mitigate LPS-induced injuries in cardiomyocytes | [56] | |
| MALAT1 | Host intergenic lncRNA | Regulate macrophage polarization | [57] | |
| Brucella | lnc_000428 | Host antisense lncRNA | Regulate Brucella intracellular replication | [60] |
| Gm28309 | Host intronic lncRNA | Regulate inflammatory and anti-Brucella responses via NF-κB/NLRP3 signaling | [61] | |
| linc-MAF-4 | Host intergenic lncRNA | Biomarker | [62] | |
| IFNG-AS1 | Host intergenic lncRNA | Biomarker | [63] | |
| S. typhimurium | LNCGM1082 | Host intergenic lncRNA | Activate NLRC4 and induce resistance to S. typhimurium | [66] |
| NeST (IFNG-AS1) | Host intergenic lncRNA | Modulate host susceptibility to pathogens by altering epigenetic marking of IFNγ-encoding chromatin | [67] | |
| TVX1 | Host intergenic lncRNA | Attenuated S. typhimurium-induced microglial inflammation | [68] | |
| NEAT1 | Host intergenic lncRNA | Biomarker | [65] | |
| P. aeruginosa | MEG3 | Host intergenic lncRNA | Influence the proliferation of P. aeruginosa by miR-138/IL-1β axis | [71] |
| NRIR | Host intergenic lncRNA | Affect the maturation of dendritic cell and the activation of T cell | [72] | |
| MEG9 | Host intergenic lncRNA | Biomarker | [70] | |
| BLACAT1 | Host intronic lncRNA | Biomarker | [70] | |
| L. monocytogenes | lincRNA-EPS | Host intergenic lncRNA | Impair the host defense against L. monocytogenes infection | [79,80] |
| lincRNA-Cox2 | Host intergenic lncRNA | Regulate migration and phagocytosis of macrophages | [81] | |
| AS-IL-1α | Host antisense lncRNA | A regulator of innate immune response by regulating IL-1α transcription | [82] | |
| SROS1 | Host intergenic lncRNA | Promote IFN-γ-STAT1-mediated innate immunity | [83] | |
| lasRNAs | Pathogen-derived lncRNA | Represent a regulatory pattern that connect adjacent genes with opposing functions | [84] | |
| S. aureus | BMNCR | Host intronic lncRNA | Influence the proliferation and apoptosis of epithelial cells | [91] |
| LRRC75A-AS | Host antisense lncRNA | Regulate the expression of tight junctions and affect inflammation | [92] | |
| SSR42 | Pathogen-derived lncRNA | Modulate the expression of several virulence factors | [89,90] |
3. LncRNAs in Viral Zoonoses
3.1. Rabies
3.2. Ebola Virus Disease
3.3. Flavivirus Infection
3.4. AIDS
3.5. Influenza
3.6. Herpesvirus Infection
3.7. Coronavirus Disease
| Pathogen | LncRNA | Category | Function or Mechanism | Reference |
|---|---|---|---|---|
| RABV | EDAL | Host intergenic lncRNA | Inhibit the replication of neurotropic virus | [102,103] |
| DENV (Flaviviridae) | NEAT1 | Host intergenic lncRNA | Affect antiviral response and viral replication in dengue infection | [113] |
| ERGAL | Host intergenic lncRNA | Promote stability and integrity of vascular endothelial barrier during DENV infection | [114] | |
| JEV (Flaviviridae) | SUSAJ1 | Host sense overlapping lncRNA | Inhibit JEV proliferation and replication | [115,116] |
| Flaviviridae | JINR1 | Host intergenic lncRNA | Regulate viral replication and cell death | [117] |
| ZAP-IT1 | Host intronic lncRNA | Exert antiviral effect in an IFN-independent manner | [118] | |
| MALAT1 | Host intergenic lncRNA | Potential antiviral function | [119] | |
| Gm20559 | Host intergenic lncRNA | Modulate the expression of various pro-inflammatory cytokines during flavivirus infection | [120] | |
| sfRNAs/ xrRNAs | Pathogen-derived lncRNA | Impact viral replication | [121] | |
| HIV | RUNXOR | Host sense overlapping lncRNA | Regulate multiple immunosuppressive signaling molecules | [126,164] |
| HOTAIRM1 | Host intergenic lncRNA | Increase levels of immunosuppressive molecules | [127] | |
| GAS5 | Host antisense lncRNA | Control HIV replication, regulate the activity and longevity of CD4 T cells | [128,129] | |
| NKILA | Host antisense lncRNA | Inhibit HIV-1 replication by suppressing HIV-1 LTR promoter activity | [130] | |
| Influenza viruses | PSMB8-AS1 | Host antisense lncRNA | Promotes influenza virus replication | [137,138] |
| LINC01191 (VIN) | Host intergenic lncRNA | Regulate viral protein synthesis | [137,140] | |
| DANCR | Host intergenic lncRNA | Involved in respiratory infections and regulate inflammation | [137,139] | |
| BCAR4 | Host intergenic lncRNA | Biomarker | [137] | |
| LncRNA#61 | Host sense overlapping lncRNA | Suppress viral replication, mediate host immune responses | [141] | |
| LncRNA#45 | Host intronic lncRNA | Function as a broad-spectrum antiviral factor | [142] | |
| CHROMR | Host antisense lncRNA | Restrict influenza virus replication by sequestering IRF2/IRF2BP2 complex | [143] | |
| lncNSPL | Host intergenic lncRNA | Influence influenza immune escape by modulating IFN-I expression | [144] | |
| RDUR | Host intergenic lncRNA | Regulate innate immunity against virus by controlling IFN-β and ISGs | [145] | |
| PRV (Orthoherpesviridae) | lnc_000641 | Host intergenic lncRNA | Influence PRV replication through JAK-STAT1 pathway | [149] |
| lncA02830 | Host intronic lncRNA | Affect PRV replication in a IFN-dependent manner | [150] | |
| NOIR1/NOIR2 | Pathogen-derived lncRNA | Locate in the IR region of the PRV | [152] | |
| PTO/PTO-US1 | Pathogen-derived lncRNA | Overlap with the oriS region of the PRV | [152] | |
| CTO-S/CTO-L | Pathogen-derived lncRNA | Function as TATA boxes in herpesviruses | [152] | |
| AZURE | Pathogen-derived lncRNA | Locate in the IR-US overlapping region of the PRV | [152] | |
| EBV (Orthoherpesviridae) | H19 | Host intergenic lncRNA | Biomarker | [151] |
| SARS-CoV-2 | GAS5 | Host antisense lncRNA | Affect SARS-CoV-2 invasion via GAS5/miRNA-200/ACE2 axis | [159] |
| SNHG15 | Host intergenic lncRNA | Aid SARS-CoV-2 entry through RABL2A, facilitate memory CD8+ T cell production | [160] | |
| PIRAT | Host intergenic lncRNA | Modulate systemic antiviral responses to SARS-CoV-2 | [161] | |
| LUCAT1 | Host intergenic lncRNA | Modulate systemic antiviral responses to SARS-CoV-2 | [161] | |
| XIST | Host intergenic lncRNA | Biomarker | [162] | |
| ZFY-AS1 | Host antisense lncRNA | Biomarker | [162] | |
| TTTY14 | Host intergenic lncRNA | Biomarker | [162] | |
| MALAT1 | Host intergenic lncRNA | Biomarker | [163] | |
| MEG3 | Host intergenic lncRNA | Biomarker | [163] |
4. LncRNAs in Parasitic Zoonoses
4.1. Malaria
4.2. Schistosomiasis
4.3. Cryptosporidiosis
4.4. Toxoplasmosis
4.5. Echinococcosis
| Pathogen | LncRNA | Category | Function or Mechanism | Reference |
|---|---|---|---|---|
| Plasmodium | GDV1 | Pathogen-derived lncRNA | Regulate sexual development | [171,172] |
| TARE-3-lncRNA/TARE-6-lncRNA | Pathogen-derived lncRNA | Affect the intra-erythrocytic developmental cycle of Plasmodium | [172,173,174] | |
| Var-specific lncRNA | Pathogen-derived lncRNA | Enhance the virulence of P. falciparum by modulating var | [175] | |
| LncRNA-ch14 | Pathogen-derived lncRNA | Regulate gametocyte development | [170] | |
| ENMSUSG00000111521.1 | Host antisense lncRNA | Regulate host immunity by TGF-β/Smad2/3 signaling | [176] | |
| XLOC_038009 | Host intergenic lncRNA | Regulate host immunity by TGF-β/Smad2/3 signaling | [176] | |
| XLOC_058629 | Host intergenic lncRNA | Regulate host immunity by TGF-β/Smad2/3 signaling | [176] | |
| XLOC_065676 | Host intergenic lncRNA | Regulate host immunity by TGF-β/Smad2/3 signaling | [176] | |
| MALAT1 | Host intergenic lncRNA | Function as a negative regulator of cellular immune response | [177] | |
| Schistosoma | SmLINC101519 | Pathogen-derived lncRNA | Regulate the motility of adult worms | [179] |
| SmLINC175062 | Pathogen-derived lncRNA | Regulate the motility of adult worms | [179] | |
| SmLINC110998 | Pathogen-derived lncRNA | Regulate the motility of adult worms | [179] | |
| Gm16685 | Host antisense lncRNA | Promote M1 macrophage polarization by regulating miR-205-5p | [183] | |
| H19 | Host intergenic lncRNA | Influence S. japonica infection via H19/miR-130b-3p/Cyp4a14 axis | [184] | |
| Cryptosporidium | U90926 | Host intergenic lncRNA (peptide coding) | Regulate cell autonomous antiparasitic defense in a pro-parasitic manner | [186] |
| Nostrill | Host intergenic lncRNA | Promote antiparasitic defense through regulating NF-κB p65 | [187] | |
| NR_045064 | Host intergenic lncRNA | Promote host defense against Cryptosporidium by modulating NOS2/CSF2 | [188] | |
| XR_001779380 | Host intergenic lncRNA | Relevant to anti-Cryptosporidium defense in a IFN- dependent manner | [189] | |
| NR_033736 | Host intergenic lncRNA | Contribute to host innate defense against Cryptosporidium | [190] | |
| Toxoplasma | Csf1-lnc | Host sense overlapping lncRNA | Controlled by secretory kinase ROP16 | [192] |
| Socs2-lnc | Host sense overlapping lncRNA | Controlled by secretory kinase ROP16 | [192] | |
| lncRNA147410.3 | Host antisense lncRNA | Affect microglial proliferation, differentiation and apoptosis by targeting Hoxb3 | [193] | |
| lncRNA-11496 | Host sense overlapping lncRNA | Affect microglial proliferation, differentiation and apoptosis by targeting Mef2c | [194] | |
| NONSHAT022487 | Host antisense lncRNA | Suppress the expression of the immune-related molecule UNC93B1 | [195] | |
| Echinococcus | lncRNA028466 | Host intergenic lncRNA | Be involved in cytokine expression of Th1 and Th2 | [200] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tomori, O.; Oluwayelu, D.O. Domestic Animals as Potential Reservoirs of Zoonotic Viral Diseases. Annu. Rev. Anim. Biosci. 2023, 11, 33–55. [Google Scholar] [CrossRef]
- Kalawat, U.; Mohan, A. Endemic, Emerging and Re-Emerging Zoonotic Diseases: The Way Forward! J. Clin. Sci. Res. 2023, 12, 79–80. [Google Scholar] [CrossRef]
- Head, J.R.; Bumburidi, Y.; Mirzabekova, G.; Rakhimov, K.; Dzhumankulov, M.; Salyer, S.J.; Knust, B.; Berezovskiy, D.; Kulatayeva, M.; Zhetibaev, S.; et al. Risk Factors for and Seroprevalence of Tickborne Zoonotic Diseases among Livestock Owners, Kazakhstan. Emerg. Infect. Dis. 2020, 26, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global Trends in Emerging Infectious Diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to Zoonotic Spillover. Nat. Rev. Microbiol. 2017, 15, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.R.; Holmes, E.C.; Morens, D.M.; Park, E.-C.; Burke, D.S.; Calisher, C.H.; Laughlin, C.A.; Saif, L.J.; Daszak, P. Cross-Species Virus Transmission and the Emergence of New Epidemic Diseases. Microbiol. Mol. Biol. Rev. 2008, 72, 457–470. [Google Scholar] [CrossRef] [PubMed]
- The ENCODE Project Consortium. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long Non-Coding RNAs: Definitions, Functions, Challenges and Recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Ponting, C.P.; Haerty, W. Genome-Wide Analysis of Human Long Noncoding RNAs: A Provocative Review. Annu. Rev. Genom. Hum. Genet. 2022, 23, 153–172. [Google Scholar] [CrossRef]
- Volders, P.-J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. LNCipedia 5: Towards a Reference Set of Human Long Non-Coding RNAs. Nucleic Acids Res. 2019, 47, D135–D139. [Google Scholar] [CrossRef]
- Li, Z.; Liu, L.; Feng, C.; Qin, Y.; Xiao, J.; Zhang, Z.; Ma, L. LncBook 2.0: Integrating Human Long Non-Coding RNAs with Multi-Omics Annotations. Nucleic Acids Res. 2022, 51, D186–D191. [Google Scholar] [CrossRef]
- Peng, Z.; Liu, C.; Wu, M. New Insights into Long Noncoding RNAs and Their Roles in Glioma. Mol. Cancer 2018, 17, 61. [Google Scholar] [CrossRef]
- St Laurent, G.; Wahlestedt, C.; Kapranov, P. The Landscape of Long Non-Coding RNA Classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef]
- Cabili, M.N.; Dunagin, M.C.; McClanahan, P.D.; Biaesch, A.; Padovan-Merhar, O.; Regev, A.; Rinn, J.L.; Raj, A. Localization and Abundance Analysis of Human lncRNAs at Single-Cell and Single-Molecule Resolution. Genome Biol. 2015, 16, 20. [Google Scholar] [CrossRef] [PubMed]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA Localization and Function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef] [PubMed]
- Guh, C.-Y.; Hsieh, Y.-H.; Chu, H.-P. Functions and Properties of Nuclear lncRNAs-from Systematically Mapping the Interactomes of lncRNAs. J. Biomed. Sci. 2020, 27, 44. [Google Scholar] [CrossRef] [PubMed]
- Vance, K.W.; Ponting, C.P. Transcriptional Regulatory Functions of Nuclear Long Noncoding RNAs. Trends Genet. 2014, 30, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yan, Z.; Fu, C.; Wen, X.; Jia, L.; Zhou, L.; Du, Z.; Wang, C.; Wang, Y.; Chen, J.; et al. LncRNA Osilr9 Coordinates Promoter DNA Demethylation and the Intrachromosomal Loop Structure Required for Maintaining Stem Cell Pluripotency. Mol. Ther. J. Am. Soc. Gene Ther. 2023, 31, 1791–1806. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Xu, J.; Sun, R.; Mumbach, M.R.; Carter, A.C.; Chen, Y.G.; Yost, K.E.; Kim, J.; He, J.; Nevins, S.A.; et al. Promoter of lncRNA Gene PVT1 Is a Tumor-Suppressor DNA Boundary Element. Cell 2018, 173, 1398–1412.e22. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Sirey, T.M.; Roberts, K.; Haerty, W.; Bedoya-Reina, O.; Rogatti-Granados, S.; Tan, J.Y.; Li, N.; Heather, L.C.; Carter, R.N.; Cooper, S.; et al. The Long Non-Coding RNA Cerox1 Is a Post Transcriptional Regulator of Mitochondrial Complex I Catalytic Activity. eLife 2019, 8, e45051. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, Y.; Zhang, Y.; Yang, J.; Zhang, M.; Li, S.; Xue, G.; Li, X.; Zhang, X.; Yang, J.; et al. Cytoplasmic Sequestration of P53 by lncRNA-CIRPILalleviates Myocardial Ischemia/Reperfusion Injury. Commun. Biol. 2022, 5, 716. [Google Scholar] [CrossRef]
- Slavoff, S.A.; Mitchell, A.J.; Schwaid, A.G.; Cabili, M.N.; Ma, J.; Levin, J.Z.; Karger, A.D.; Budnik, B.A.; Rinn, J.L.; Saghatelian, A. Peptidomic Discovery of Short Open Reading Frame-Encoded Peptides in Human Cells. Nat. Chem. Biol. 2013, 9, 59–64. [Google Scholar] [CrossRef]
- Matsumoto, A.; Pasut, A.; Matsumoto, M.; Yamashita, R.; Fung, J.; Monteleone, E.; Saghatelian, A.; Nakayama, K.I.; Clohessy, J.G.; Pandolfi, P.P. mTORC1 and Muscle Regeneration Are Regulated by the LINC00961-Encoded SPAR Polypeptide. Nature 2017, 541, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-Z.; Chen, M.; Chen, D.; Gao, X.-C.; Zhu, S.; Huang, H.; Hu, M.; Zhu, H.; Yan, G.-R. A Peptide Encoded by a Putative lncRNA HOXB-AS3 Suppresses Colon Cancer Growth. Mol. Cell 2017, 68, 171–184.e6. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zhang, C.; Ji, Y.; Ge, M.-K.; Yu, Y.; Zhang, N.; Yang, S.; Yu, J.-X.; Shen, S.-M.; Chen, G.-Q. Epithelial Cells-Enriched lncRNA SNHG8 Regulates Chromatin Condensation by Binding to Histone H1s. Cell. Death. Differ. 2022, 29, 1569–1581. [Google Scholar] [CrossRef]
- Qin, W.; Qi, X.; Xie, Y.; Wang, H.; Wu, S.; Sun, M.; Bao, W. LncRNA446 Regulates Tight Junctions by Inhibiting the Ubiquitinated Degradation of Alix after Porcine Epidemic Diarrhea Virus Infection. J. Virol. 2023, 97, e01884-22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, M.; Zhang, L.; Wang, W.; Hua, S.; Zhou, C.; Sun, X. Long Non-Coding RNAs and Immune Cells: Unveiling the Role in Viral Infections. Biomed. Pharmacother. Biomed. Pharmacother. 2023, 170, 115978. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Chen, H.; Luo, F.; Zhou, H.; Li, Z. Roles of Long Noncoding RNAs in Bacterial Infection. Life Sci. 2020, 263, 118579. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhou, Y.; Li, H. LncRNA, miRNA and lncRNA-miRNA Interaction in Viral Infection. Virus Res. 2018, 257, 25–32. [Google Scholar] [CrossRef]
- McDaniel, C.J.; Cardwell, D.M.; Moeller, R.B.; Gray, G.C. Humans and Cattle: A Review of Bovine Zoonoses. Vector Borne Zoonotic Dis. 2014, 14, 1–19. [Google Scholar] [CrossRef]
- Rahman, M.T.; Sobur, M.A.; Islam, M.S.; Ievy, S.; Hossain, M.J.; Zowalaty, M.E.E.; Rahman, A.T.; Ashour, H.M. Zoonotic Diseases: Etiology, Impact, and Control. Microorganisms 2020, 8, 1405. [Google Scholar] [CrossRef]
- Olaru, I.D.; Walther, B.; Schaumburg, F. Zoonotic Sources and the Spread of Antimicrobial Resistance from the Perspective of Low and Middle-Income Countries. Infect. Dis. Poverty 2023, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Howard, N.C.; Khader, S.A. Immunometabolism during Mycobacterium tuberculosis Infection. Trends Microbiol. 2020, 28, 832–850. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.D.; Mott, D.; Sutiwisesak, R.; Lu, Y.-J.; Raso, F.; Stowell, B.; Babunovic, G.H.; Lee, J.; Carpenter, S.M.; Way, S.S.; et al. Mycobacterium tuberculosis-Specific CD4+ and CD8+ T Cells Differ in Their Capacity to Recognize Infected Macrophages. PLoS Pathog. 2018, 14, e1007060. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhong, H.; Xie, X.; Chen, C.Y.; Huang, D.; Shen, L.; Zhang, H.; Chen, Z.W.; Zeng, G. Long Noncoding RNA Derived from CD244 Signaling Epigenetically Controls CD8+ T-Cell Immune Responses in Tuberculosis Infection. Proc. Natl. Acad. Sci. USA 2015, 112, E3883–E3892. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tan, Y.; Zhang, X.; Cheng, M.; Hu, J.; Liu, J.; Chen, X.; Zhu, J. Comprehensive Identification of Immuno-Related Transcriptional Signature for Active Pulmonary Tuberculosis by Integrated Analysis of Array and Single Cell RNA-Seq. J. Infect. 2022, 85, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Xu, X.; Xue, J.; Duan, W.; Yi, Z. Deregulated lncRNAs in B Cells from Patients with Active Tuberculosis. PLoS ONE 2017, 12, e0170712. [Google Scholar] [CrossRef]
- Li, M.; Cui, J.; Niu, W.; Huang, J.; Feng, T.; Sun, B.; Yao, H. Long Non-Coding PCED1B-AS1 Regulates Macrophage Apoptosis and Autophagy by Sponging miR-155 in Active Tuberculosis. Biochem. Biophys. Res. Commun. 2019, 509, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Xie, Y.; Xu, D.; Qu, Z.; Wu, J.; Zhou, Y.; Wei, Y.; Xiong, H.; Zhang, X.-L. Lnc-EST12, Which Is Negatively Regulated by Mycobacterial EST12, Suppresses Antimycobacterial Innate Immunity through Its Interaction with FUBP3. Cell. Mol. Immunol. 2022, 19, 883–897. [Google Scholar] [CrossRef]
- Qu, Y.; Jiang, D.; Liu, M.; Wang, H.; Xu, T.; Zhou, H.; Huang, M.; Shu, W.; Xu, G. LncRNA DANCR Restrained the Survival of Mycobacterium tuberculosis H37Ra by Sponging miR-1301-3p/miR-5194. Front. Microbiol. 2023, 14, 1119629. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.-B.; Li, L.-T.; Xi, J.-C.; Liu, H.-T.; Liu, Z.; Yu, L.; Tang, P.-F. Negative Pressure Promotes Macrophage M1 Polarization after Mycobacterium tuberculosis Infection via the lncRNA XIST/microRNA-125b-5p/A20/NF-κB Axis. Ann. N. Y. Acad. Sci. 2022, 1514, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Lou, J.; Zheng, X.-M.; Yang, X.-Y. LncRNA MIAT Regulates Autophagy and Apoptosis of Macrophage Infected by Mycobacterium tuberculosis through the miR-665/ULK1 Signaling Axis. Mol. Immunol. 2021, 139, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Gcanga, L.; Tamgue, O.; Ozturk, M.; Pillay, S.; Jacobs, R.; Chia, J.E.; Mbandi, S.K.; Davids, M.; Dheda, K.; Schmeier, S.; et al. Host-Directed Targeting of LincRNA-MIR99AHG Suppresses Intracellular Growth of Mycobacterium tuberculosis. Nucleic Acid Ther. 2022, 32, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Liu, Y.; Ma, Y.; Yang, F.; Ruan, Y.; Xu, J.-F.; Pi, J. Advances of Long Non-Coding RNAs as Potential Biomarkers for Tuberculosis: New Hope for Diagnosis? Pharmaceutics 2023, 15, 2096. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, J.; Wang, J.; Wen, Q.; Wang, H.; He, J.; Hu, S.; He, W.; Du, X.; Liu, S.; et al. Microarray Analysis of Long Noncoding RNA and mRNA Expression Profiles in Human Macrophages Infected with Mycobacterium tuberculosis. Sci. Rep. 2016, 6, 38963. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, X.; Tong, P.; Zheng, B.; Zhu, M.; Peng, B.; Wang, J.; Liu, Y. RNA-Seq Analysis of Long Non-Coding RNA in Human Intestinal Epithelial Cells Infected by Shiga Toxin-Producing Escherichia coli. Cytokine 2024, 173, 156421. [Google Scholar] [CrossRef]
- Yang, R.; Wang, X.; Liu, H.; Chen, J.; Tan, C.; Chen, H.; Wang, X. Egr-1 Is a Key Regulator of the Blood-Brain Barrier Damage Induced by Meningitic Escherichia coli. Cell Commun. Signal. 2024, 22, 44. [Google Scholar] [CrossRef]
- Wu, Z.; Fan, H.; Jin, J.; Gao, S.; Huang, R.; Wu, S.; Bao, W. Insight into Mechanisms of Pig lncRNA FUT3-AS1 Regulating E. coli F18-Bacterial Diarrhea. PLoS Pathog. 2022, 18, e1010584. [Google Scholar] [CrossRef]
- Yang, B.; Yin, P.; Yang, R.; Xu, B.; Fu, J.; Zhi, S.; Dai, M.; Tan, C.; Chen, H.; Wang, X. Holistic Insights into Meningitic Escherichia coli Infection of Astrocytes Based on Whole Transcriptome Profiling. Epigenomics 2020, 12, 1611–1632. [Google Scholar] [CrossRef]
- Xu, B.; Yang, R.; Yang, B.; Li, L.; Chen, J.; Fu, J.; Qu, X.; Huo, D.; Tan, C.; Chen, H.; et al. Long Non-Coding RNA lncC11orf54-1 Modulates Neuroinflammatory Responses by Activating NF-κB Signaling during Meningitic Escherichia Coli Infection. Mol. Brain 2022, 15, 4. [Google Scholar] [CrossRef]
- Xu, B.; Yang, R.; Fu, J.; Yang, B.; Chen, J.; Tan, C.; Chen, H.; Wang, X. LncRSPH9-4 Facilitates Meningitic Escherichia Coli-Caused Blood-Brain Barrier Disruption via miR-17-5p/MMP3 Axis. Int. J. Mol. Sci. 2021, 22, 6343. [Google Scholar] [CrossRef]
- Yang, B.; Xu, B.; Yang, R.; Fu, J.; Li, L.; Huo, D.; Chen, J.; Yang, X.; Tan, C.; Chen, H.; et al. Long Non-Coding Antisense RNA DDIT4-AS1 Regulates Meningitic Escherichia Coli-Induced Neuroinflammation by Promoting DDIT4 mRNA Stability. Mol. Neurobiol. 2022, 59, 1351–1365. [Google Scholar] [CrossRef]
- Ma, M.; Pei, Y.; Wang, X.; Feng, J.; Zhang, Y.; Gao, M.-Q. LncRNA XIST Mediates Bovine Mammary Epithelial Cell Inflammatory Response via NF-κB/NLRP3 Inflammasome Pathway. Cell Prolif. 2019, 52, e12525. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, J.; Jiang, X.; Wang, H.; Pan, G. LncRNA HOX Transcript Antisense RNA Accelerated Kidney Injury Induced by Urine-Derived Sepsis through the miR-22/High Mobility Group Box 1 Pathway. Life Sci. 2018, 210, 185–191. [Google Scholar] [CrossRef]
- Zhu, W.; Peng, F.; Cui, X.; Li, J.; Sun, C. LncRNA SOX2OT Facilitates LPS-Induced Inflammatory Injury by Regulating Intercellular Adhesion Molecule 1 (ICAM1) via Sponging miR-215-5p. Clin. Immunol. 2022, 238, 109006. [Google Scholar] [CrossRef]
- Ahmad, I.; Naqvi, R.A.; Valverde, A.; Naqvi, A.R. LncRNA MALAT1/microRNA-30b Axis Regulates Macrophage Polarization and Function. Front. Immunol. 2023, 14, 1214810. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.B.; Lee, W.J.; Shin, M.K.; Jung, M.H.; Shin, S.W.; Yoo, A.N.; Kim, J.W.; Yoo, H.S. Early Transcriptional Responses of Internalization Defective Brucella Abortus Mutants in Professional Phagocytes, RAW 264.7. BMC Genom. 2013, 14, 426. [Google Scholar] [CrossRef] [PubMed]
- von Bargen, K.; Gorvel, J.P.; Salcedo, S.P. Internal Affairs: Investigating the Brucella Intracellular Lifestyle. FEMS Microbiol. Rev. 2012, 36, 533–562. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Hu, H.; Tian, M.; Zhuang, H.; Ding, C.; Yu, S. Differentially Expressed Long Noncoding RNAs in RAW264.7 Macrophages during Brucella Infection and Functional Analysis on the Bacterial Intracellular Replication. Sci. Rep. 2022, 12, 21320. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Guo, J.; Sun, Z.; Liu, L.; Zhao, T.; Li, J.; Tang, G.; Zhu, D.; Tao, T.; Zhang, H. Brucella-Induced Downregulation of lncRNA Gm28309 Triggers Macrophages Inflammatory Response Through the miR-3068-5p/NF-κB Pathway. Front. Immunol. 2020, 11, 805275. [Google Scholar] [CrossRef] [PubMed]
- Gheitasi, R.; Keramat, F.; Solgi, G.; Hajilooi, M. Investigation of Linc-MAF-4 Expression as an Effective Marker in Brucellosis. Mol. Immunol. 2020, 123, 60–63. [Google Scholar] [CrossRef]
- Gheitasi, R.; Jourghasemi, S.; Pakzad, I.; Hosseinpour Sarmadi, V.; Samieipour, Y.; Sekawi, Z.; Azizi Jalilian, F. A Potential Marker in Brucellosis, Long Non Coding RNA IFNG-AS1. Mol. Biol. Rep. 2019, 46, 6495–6500. [Google Scholar] [CrossRef] [PubMed]
- Poppe, C.; Smart, N.; Khakhria, R.; Johnson, W.; Spika, J.; Prescott, J. Salmonella typhimurium DT104: A Virulent and Drug-Resistant Pathogen. Can. Vet. J. 1998, 39, 559–565. [Google Scholar] [PubMed]
- Imamura, K.; Takaya, A.; Ishida, Y.; Fukuoka, Y.; Taya, T.; Nakaki, R.; Kakeda, M.; Imamachi, N.; Sato, A.; Yamada, T.; et al. Diminished Nuclear RNA Decay upon Salmonella Infection Upregulates Antibacterial Noncoding RNAs. EMBO J. 2018, 37, e97723. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, Y.; Wei, J.; Yue, J.; Wang, Y.; Zhang, Q.; Jin, M.; Wang, R.; Yang, X.; Zhang, J.; et al. LNCGM1082-Mediated NLRC4 Activation Drives Resistance to Bacterial Infection. Cell. Mol. Immunol. 2023, 20, 475–488. [Google Scholar] [CrossRef]
- Monack, D.M.; Chang, H.Y.; Brahic, M.; Kirkegaard, K. NeST, a Long Noncoding RNA, Controls Microbial Susceptibility and Epigenetic Activation of the Ifng Locus. Cell 2013, 152, 743. [Google Scholar] [CrossRef]
- Zou, W.; Zhang, J.; Zhang, K.; Peng, Z.; Xin, R.; Wang, L.; Li, J. Asiatic Acid Attenuates Inflammation Induced by Salmonella via Upregulating LncRNA TVX1 in Microglia. Int. J. Mol. Sci. 2022, 23, 10978. [Google Scholar] [CrossRef]
- Saleh, L.A.; Boyd, A.; Aragon, I.V.; Koloteva, A.; Spadafora, D.; Mneimneh, W.; Barrington, R.A.; Richter, W. Ablation of PDE4B Protects from Pseudomonas aeruginosa-Induced Acute Lung Injury in Mice by Ameliorating the Cytostorm and Associated Hypothermia. FASEB J. 2021, 35, e21797. [Google Scholar] [CrossRef]
- Balloy, V.; Koshy, R.; Perra, L.; Corvol, H.; Chignard, M.; Guillot, L.; Scaria, V. Bronchial Epithelial Cells from Cystic Fibrosis Patients Express a Specific Long Non-Coding RNA Signature upon Pseudomonas aeruginosa Infection. Front. Cell. Infect. Microbiol. 2017, 7, 218. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Fang, L.; Pu, Q.; Bu, H.; Zhu, P.; Chen, Z.; Yu, M.; Li, X.; Weiland, T.; Bansal, A.; et al. MEG3-4 Is a miRNA Decoy That Regulates IL-1β Abundance to Initiate and Then Limit Inflammation to Prevent Sepsis during Lung Infection. Sci. Signal. 2018, 11, eaao2387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Y.; Lu, Y.; Li, S.; Liu, J.; Zhang, Y.; Wang, L.; Li, M.; Luo, Y.; Zhang, W.; et al. N-3-(Oxododecanoyl)-L-Homoserine Lactone Suppresses Dendritic Cell Maturation by Upregulating the Long Noncoding RNA NRIR. J. Biosci. 2021, 46, 65. [Google Scholar] [CrossRef]
- Mogotu, M.W.; Abong, G.O.; Mburu, J.; Ndambi, O.A. Assessment of Hygiene Practices and Microbial Safety of Milk Supplied by Smallholder Farmers to Processors in Selected Counties in Kenya. Trop. Anim. Health Prod. 2022, 54, 220. [Google Scholar] [CrossRef] [PubMed]
- Indramohan, M.; Sieve, A.N.; Break, T.J.; Berg, R.E. Inflammatory Monocyte Recruitment Is Regulated by Interleukin-23 during Systemic Bacterial Infection. Infect. Immun. 2012, 80, 4099. [Google Scholar] [CrossRef] [PubMed]
- Dussurget, O.; Bierne, H.; Cossart, P. The Bacterial Pathogen Listeria monocytogenes and the Interferon Family: Type I, Type II and Type III Interferons. Front. Cell. Infect. Microbiol. 2014, 4, 50. [Google Scholar] [CrossRef]
- Quereda, J.J.; Dussurget, O.; Nahori, M.-A.; Ghozlane, A.; Volant, S.; Dillies, M.-A.; Regnault, B.; Kennedy, S.; Mondot, S.; Villoing, B.; et al. Bacteriocin from Epidemic Listeria Strains Alters the Host Intestinal Microbiota to Favor Infection. Proc. Natl. Acad. Sci. USA 2016, 113, 5706–5711. [Google Scholar] [CrossRef] [PubMed]
- Lam, G.Y.; Cemma, M.; Muise, A.M.; Higgins, D.E.; Brumell, J.H. Host and Bacterial Factors That Regulate LC3 Recruitment to Listeria monocytogenes during the Early Stages of Macrophage Infection. Autophagy 2013, 9, 985–995. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, D.; Chen, G.; Li, G.; Dou, S.; Wang, R.; Xiao, H.; Hou, C.; Li, Y.; Feng, J.; et al. Tim-3 Inhibits Macrophage Control of Listeria Monocytogenes by Inhibiting Nrf2. Sci. Rep. 2017, 7, 42095. [Google Scholar] [CrossRef]
- Atianand, M.K.; Hu, W.; Satpathy, A.T.; Shen, Y.; Ricci, E.P.; Alvarez-Dominguez, J.R.; Bhatta, A.; Schattgen, S.A.; McGowan, J.D.; Blin, J.; et al. A Long Noncoding RNA lincRNA-EPS Acts as a Transcriptional Brake to Restrain Inflammation. Cell 2016, 165, 1672–1685. [Google Scholar] [CrossRef]
- Agliano, F.; Fitzgerald, K.A.; Vella, A.T.; Rathinam, V.A.; Medvedev, A.E. Long Non-Coding RNA LincRNA-EPS Inhibits Host Defense Against Listeria monocytogenes Infection. Front. Cell. Infect. Microbiol. 2019, 9, 481. [Google Scholar] [CrossRef]
- Zhu, Y.; Lu, Y.; Yuan, L.; Ling, W.; Jiang, X.; Chen, S.; Hu, B. LincRNA-Cox2 Regulates IL6/JAK3/STAT3 and NF-κB P65 Pathway Activation in Listeria monocytogenes-Infected RAW264.7 Cells. Int. J. Med. Microbiol. 2021, 311, 151515. [Google Scholar] [CrossRef]
- Chan, J.; Atianand, M.; Jiang, Z.; Carpenter, S.; Aiello, D.; Elling, R.; Fitzgerald, K.A.; Caffrey, D.R. Cutting Edge: A Natural Antisense Transcript, AS-IL1α, Controls Inducible Transcription of the Proinflammatory Cytokine IL-1α. J. Immunol. 2015, 195, 1359–1363. [Google Scholar] [CrossRef]
- Xu, H.; Jiang, Y.; Xu, X.; Su, X.; Liu, Y.; Ma, Y.; Zhao, Y.; Shen, Z.; Huang, B.; Cao, X. Inducible Degradation of lncRNA Sros1 Promotes IFN-γ-Mediated Activation of Innate Immune Responses by Stabilizing Stat1 mRNA. Nat. Immunol. 2019, 20, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Wurtzel, O.; Sesto, N.; Mellin, J.R.; Karunker, I.; Edelheit, S.; Bécavin, C.; Archambaud, C.; Cossart, P.; Sorek, R. Comparative Transcriptomics of Pathogenic and Non-Pathogenic Listeria Species. Mol. Syst. Biol. 2012, 8, 583. [Google Scholar] [CrossRef]
- Foster, T. Staphylococcus. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 978-0-9631172-1-2. [Google Scholar]
- Algammal, A.M.; Hetta, H.F.; Elkelish, A.; Alkhalifah, D.H.H.; Hozzein, W.N.; Batiha, G.E.-S.; El Nahhas, N.; Mabrok, M.A. Methicillin-Resistant Staphylococcus aureus (MRSA): One Health Perspective Approach to the Bacterium Epidemiology, Virulence Factors, Antibiotic-Resistance, and Zoonotic Impact. Infect. Drug Resist. 2020, 13, 3255–3265. [Google Scholar] [CrossRef]
- Wilke, G.A.; Wardenburg, J.B. Role of a Disintegrin and Metalloprotease 10 in Staphylococcus Aureus α-Hemolysin–Mediated Cellular Injury. Proc. Natl. Acad. Sci. USA 2010, 107, 13473–13478. [Google Scholar] [CrossRef]
- Duan, J.; Li, M.; Hao, Z.; Shen, X.; Liu, L.; Jin, Y.; Wang, S.; Guo, Y.; Yang, L.; Wang, L.; et al. Subinhibitory Concentrations of Resveratrol Reduce Alpha-Hemolysin Production in Staphylococcus Aureus Isolates by Downregulating saeRS. Emerg. Microbes Infect. 2018, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Horn, J.; Klepsch, M.; Manger, M.; Wolz, C.; Rudel, T.; Fraunholz, M. Long Noncoding RNA SSR42 Controls Staphylococcus Aureus Alpha-Toxin Transcription in Response to Environmental Stimuli. J. Bacteriol. 2018, 200, e00252-18. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.M.; Miller, E.W.; Benson, M.A.; Alonzo, F.; Yoong, P.; Torres, V.J.; Hinrichs, S.H.; Dunman, P.M. Characterization of SSR42, a Novel Virulence Factor Regulatory RNA That Contributes to the Pathogenesis of a Staphylococcus Aureus USA300 Representative. J. Bacteriol. 2012, 194, 2924–2938. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhao, T.; Ma, Y.; Wu, X.; Chu, S.; Yang, Z. Multiple Roles of LncRNA-BMNCR on Cell Proliferation and Apoptosis by Targeting miR-145/CBFB Axis in BMECs. Vet. Q. 2023, 43, 1–11. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Zhang, R.; Li, D.; Gao, M.-Q. LRRC75A Antisense lncRNA1 Knockout Attenuates Inflammatory Responses of Bovine Mammary Epithelial Cells. Int. J. Biol. Sci. 2020, 16, 251. [Google Scholar] [CrossRef] [PubMed]
- Aldakheel, F.M.; Syed, R.; Ahmed, M.; Xu, T. Modulation of lncRNA NEAT1 Overturns the Macrophages Based Immune Response in M. tuberculosis Infected Patients via miR-373 Regulation. J. Appl. Genet. 2023. [Google Scholar] [CrossRef]
- Ke, Z.; Lu, J.; Zhu, J.; Yang, Z.; Jin, Z.; Yuan, L. Down-Regulation of lincRNA-EPS Regulates Apoptosis and Autophagy in BCG-Infected RAW264.7 Macrophages via JNK/MAPK Signaling Pathway. Infect. Genet. Evol. 2020, 77, 104077. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yu, J.; Ma, C.; Gong, Z.; Wu, X.; Deng, G. Impact of Knockdown LincRNA-Cox2 on Apoptosis of Macrophage Infected with Bacillus Calmette-Guérin. Mol. Immunol. 2021, 130, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Heeney, J.L. Zoonotic Viral Diseases and the Frontier of Early Diagnosis, Control and Prevention. J. Intern. Med. 2006, 260, 399–408. [Google Scholar] [CrossRef]
- Reed, K.D. Viral Zoonoses. In Reference Module in Biomedical Sciences; Elsevier: Madison, WI, USA, 2018; ISBN 978-0-12-801238-3. [Google Scholar]
- Gan, H.; Hou, X.; Wang, Y.; Xu, G.; Huang, Z.; Zhang, T.; Lin, R.; Xue, M.; Hu, H.; Liu, M.; et al. Global Burden of Rabies in 204 Countries and Territories, from 1990 to 2019: Results from the Global Burden of Disease Study 2019. Int. J. Infect. Dis. 2023, 126, 136–144. [Google Scholar] [CrossRef]
- Diallo, M.K.; Diallo, A.O.; Dicko, A.; Richard, V.; Espié, E. Human Rabies Post Exposure Prophylaxis at the Pasteur Institute of Dakar, Senegal: Trends and Risk Factors. BMC Infect. Dis. 2019, 19, 321. [Google Scholar] [CrossRef]
- Zhao, P.; Guo, S.; Zhong, Z.; Yang, S.; Xia, X. Quantitative Characterization of the B Cell Receptor Repertoires of Human Immunized with Commercial Rabies Virus Vaccine. Hum. Vaccines Immunother. 2021, 17, 2538–2546. [Google Scholar] [CrossRef]
- Potratz, M.; Zaeck, L.M.; Weigel, C.; Klein, A.; Freuling, C.M.; Müller, T.; Finke, S. Neuroglia Infection by Rabies Virus after Anterograde Virus Spread in Peripheral Neurons. Acta Neuropathol. Commun. 2020, 8, 199. [Google Scholar] [CrossRef]
- Sui, B.; Chen, D.; Liu, W.; Wu, Q.; Tian, B.; Li, Y.; Hou, J.; Liu, S.; Xie, J.; Jiang, H.; et al. A Novel Antiviral lncRNA, EDAL, Shields a T309 O-GlcNAcylation Site to Promote EZH2 Lysosomal Degradation. Genome Biol. 2020, 21, 228. [Google Scholar] [CrossRef]
- Sui, B.; Zhao, J.; Zheng, J.; Zhou, M.; Chen, H.; Fu, Z.F.; Zhao, L. lncRNA EDAL Restricts Rabies Lyssavirus Replication in a Cell-Specific and Infection Route-Dependent Manner. J. Gen. Virol. 2022, 103, 001725. [Google Scholar] [CrossRef]
- Marí Saéz, A.; Weiss, S.; Nowak, K.; Lapeyre, V.; Zimmermann, F.; Düx, A.; Kühl, H.S.; Kaba, M.; Regnaut, S.; Merkel, K.; et al. Investigating the Zoonotic Origin of the West African Ebola Epidemic. EMBO Mol. Med. 2015, 7, 17–23. [Google Scholar] [CrossRef]
- Jacob, S.T.; Crozier, I.; Fischer, W.A.; Hewlett, A.; Kraft, C.S.; Vega, M.A.D.L.; Soka, M.J.; Wahl, V.; Griffiths, A.; Bollinger, L.; et al. Ebola Virus Disease. Nat. Rev. Dis. Primer 2020, 6, 13. [Google Scholar] [CrossRef]
- Santus, L.; Sopena-Rios, M.; García-Pérez, R.; Lin, A.E.; Adams, G.C.; Barnes, K.G.; Siddle, K.J.; Wohl, S.; Reverter, F.; Rinn, J.L.; et al. Single-Cell Profiling of lncRNA Expression during Ebola Virus Infection in Rhesus Macaques. Nat. Commun. 2023, 14, 3866. [Google Scholar] [CrossRef]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef]
- Brinton, M.A.; Basu, M. Functions of the 3′ and 5′ Genome RNA Regions of Members of the Genus Flavivirus. Virus Res. 2015, 206, 108–119. [Google Scholar] [CrossRef]
- Murugesan, A.; Manoharan, M. Dengue Virus. Emerg. Reemerging Viral Pathog. 2020, 1, 281–359. [Google Scholar] [CrossRef]
- Kumar, S.; Nyodu, R.; Maurya, V.K.; Saxena, S.K.; Kumar, S.; Nyodu, R.; Maurya, V.K.; Saxena, S.K. Pathogenesis and Host Immune Response during Japanese Encephalitis Virus Infection. In Innate Immunity in Health and Disease; IntechOpen: London, UK, 2021; ISBN 978-1-83880-766-5. [Google Scholar]
- Hsieh, J.T.; St. John, A.L. Japanese Encephalitis Virus and Its Mechanisms of Neuroinvasion. PLoS Pathog. 2020, 16, e1008260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Qi, L.; Li, T.; Li, X.; Yang, D.; Cao, S.; Ye, J.; Wei, B. PD1+CCR2+CD8+ T Cells Infiltrate the Central Nervous System during Acute Japanese Encephalitis Virus Infection. Virol. Sin. 2019, 34, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.; Thapa, U.; Bandyopadhyay, B.; Vrati, S.; Banerjee, A. Knockdown of NEAT1 Restricts Dengue Virus Replication by Augmenting Interferon Alpha-Inducible Protein 27 via the RIG-I Pathway. J. Gen. Virol. 2023, 104. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Wang, H.; Cui, G.; Guo, Q.; Si, L.; Yan, H.; Fang, D.; Jiang, L.; Jiang, Z.; Zhou, J. ERG-Associated lncRNA (ERGAL) Promotes the Stability and Integrity of Vascular Endothelial Barrier During Dengue Viral Infection via Interaction With miR-183-5p. Front. Cell. Infect. Microbiol. 2020, 10, 477. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yuan, Q.; Zhang, C.; Dai, Z.; Du, C.; Wang, H.; Li, X.; Yang, S.; Zhao, A. Inhibition of Japanese Encephalitis Virus Proliferation by Long Non-Coding RNA SUSAJ1 in PK-15 Cells. Virol. J. 2021, 18, 29. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Fan, J.; Wang, H.; Li, X.; Yang, S.; Zhao, A.; Zhou, X. LncRNA-SUSAJ1 Activates the ER Stress Pathway Inhibiting JEV Proliferation by Promoting PK15 Cells Apoptosis. Front. Biosci. Landmark Ed. 2022, 27, 260. [Google Scholar] [CrossRef]
- Tripathi, S.; Sengar, S.; Shree, B.; Mohapatra, S.; Basu, A.; Sharma, V. An RBM10 and NF-κB Interacting Host lncRNA Promotes JEV Replication and Neuronal Cell Death. J. Virol. 2023, 97, e0118323. [Google Scholar] [CrossRef]
- Huang, Y.; Su, Y.; Shen, L.; Huo, Z.; Chen, C.; Sun, T.; Tian, X.; Li, N.; Yang, C. A Novel IFNbeta-Induced Long Non-Coding RNA ZAP-IT1 Interrupts Zika Virus Replication in A549 Cells. Virol. Sin. 2022, 37, 904. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Vrati, S. The Malat1 Long Non-Coding RNA Is Upregulated by Signalling through the PERK Axis of Unfolded Protein Response during Flavivirus Infection. Sci. Rep. 2015, 5, 17794. [Google Scholar] [CrossRef]
- Mohapatra, S.; Tripathi, S.; Sharma, V.; Basu, A. Regulation of Microglia-Mediated Inflammation by Host lncRNA Gm20559 upon Flaviviral Infection. Cytokine 2023, 172, 156383. [Google Scholar] [CrossRef]
- Charley, P.A.; Wilusz, J. Standing Your Ground to Exoribonucleases: Function of Flavivirus Long Non-Coding RNAs. Virus Res. 2016, 212, 70–77. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Trends in Sexual Risk Behavior among High School Students—United States, 1990, 1991, and 1993. MMWR Morb. Mortal. Wkly. Rep. 1995, 44, 124–125, 131–132. [Google Scholar]
- van Schalkwyk, C.; Mahy, M.; Johnson, L.F.; Imai-Eaton, J.W. Updated Data and Methods for the 2023 UNAIDS HIV Estimates. J. Acquir. Immune Defic. Syndr. 2024, 95, e1–e4. [Google Scholar] [CrossRef]
- Sharp, P.M.; Hahn, B.H. Origins of HIV and the AIDS Pandemic. Cold Spring Harb. Perspect. Med. 2011, 1, a006841. [Google Scholar] [CrossRef] [PubMed]
- Le Goaster, J.; Bouree, P.; El Sissy, F.N.; Phuong Bui, F.; Pokossy Epee, J.; Rollin, P.; Tangy, F.; Haenni, A.-L. HSV-1/HSV-2 Infection-Related Cancers in Bantu Populations Driving HIV-1 Prevalence in Africa: Tracking the Origin of AIDS at the Onset of the 20th Century. Case Rep. Oncol. 2016, 9, 815–825. [Google Scholar] [CrossRef]
- Zhang, J.; Thakuri, B.K.C.; Zhao, J.; Nguyen, L.N.; Nguyen, L.N.T.; Khanal, S.; Cao, D.; Dang, X.; Schank, M.; Lu, Z.; et al. Long Noncoding RNA RUNXOR Promotes Myeloid-Derived Suppressor Cell Expansion and Functions via Enhancing Immunosuppressive Molecule Expressions during Latent HIV Infection. J. Immunol. 2021, 206, 2052–2060. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Thakuri, B.K.C.; Zhao, J.; Nguyen, L.N.; Nguyen, L.N.T.; Cao, D.; Dang, X.; Khanal, S.; Schank, M.; Lu, Z.; et al. Long Noncoding RNA HOTAIRM1 Promotes Myeloid-Derived Suppressor Cell Expansion and Suppressive Functions through up-Regulating HOXA1 Expression during Latent HIV Infection. AIDS Lond. Engl. 2020, 34, 2211–2221. [Google Scholar] [CrossRef]
- Chen, L.; Chen, L.; Zuo, L.; Gao, Z.; Shi, Y.; Yuan, P.; Han, S.; Yin, J.; Peng, B.; He, X.; et al. Short Communication: Long Noncoding RNA GAS5 Inhibits HIV-1 Replication Through Interaction with miR-873. AIDS Res. Hum. Retroviruses 2018, 34, 544–549. [Google Scholar] [CrossRef]
- Nguyen, L.N.T.; Nguyen, L.N.; Zhao, J.; Schank, M.; Dang, X.; Cao, D.; Khanal, S.; Chand Thakuri, B.K.; Lu, Z.; Zhang, J.; et al. Long Non-Coding RNA GAS5 Regulates T Cell Functions via miR21-Mediated Signaling in People Living With HIV. Front. Immunol. 2021, 12, 601298. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Huan, C.; Yang, J.; Li, Z.; Zheng, B.; Wang, Y.; Zhang, W. NF-κB-Interacting Long Noncoding RNA Regulates HIV-1 Replication and Latency by Repressing NF-κB Signaling. J. Virol. 2020, 94, e01057-20. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Abe, J.; Adkins, S.; Alkhovsky, S.V.; Avšič-Županc, T.; Ayllón, M.A.; Bahl, J.; Balkema-Buschmann, A.; Ballinger, M.J.; Baranwal, V.K.; et al. Annual (2023) Taxonomic Update of RNA-Directed RNA Polymerase-Encoding Negative-Sense RNA Viruses (Realm Riboviria: Kingdom Orthornavirae: Phylum Negarnaviricota). J. Gen. Virol. 2023, 104, 001864. [Google Scholar] [CrossRef]
- Mostafa, A.; Abdelwhab, E.M.; Mettenleiter, T.C.; Pleschka, S. Zoonotic Potential of Influenza A Viruses: A Comprehensive Overview. Viruses 2018, 10, 497. [Google Scholar] [CrossRef]
- Goneau, L.W.; Mehta, K.; Wong, J.; L’Huillier, A.G.; Gubbay, J.B. Zoonotic Influenza and Human Health-Part 1: Virology and Epidemiology of Zoonotic Influenzas. Curr. Infect. Dis. Rep. 2018, 20, 37. [Google Scholar] [CrossRef]
- Janke, B.H. Influenza A Virus Infections in Swine: Pathogenesis and Diagnosis. Vet. Pathol. 2014, 51, 410–426. [Google Scholar] [CrossRef]
- Wille, M.; Holmes, E.C. The Ecology and Evolution of Influenza Viruses. Cold Spring Harb. Perspect. Med. 2020, 10, a038489. [Google Scholar] [CrossRef]
- Richard, M.; Fouchier, R.A.M. Influenza A Virus Transmission via Respiratory Aerosols or Droplets as It Relates to Pandemic Potential. FEMS Microbiol. Rev. 2016, 40, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Thatai, A.K.S.; Ammankallu, S.; Devasahayam Arokia Balaya, R.; Soman, S.P.; Nisar, M.; Babu, S.; John, L.; George, A.; Anto, C.K.; Sanjeev, D.; et al. VirhostlncR: A Comprehensive Database to Explore lncRNAs and Their Targets in Viral Infections. Comput. Biol. Med. 2023, 164, 107279. [Google Scholar] [CrossRef] [PubMed]
- More, S.; Zhu, Z.; Lin, K.; Huang, C.; Pushparaj, S.; Liang, Y.; Sathiaseelan, R.; Yang, X.; Liu, L. Long Non-Coding RNA PSMB8-AS1 Regulates Influenza Virus Replication. RNA Biol. 2019, 16, 340–353. [Google Scholar] [CrossRef] [PubMed]
- Meydan, C.; Madrer, N.; Soreq, H. The Neat Dance of COVID-19: NEAT1, DANCR, and Co-Modulated Cholinergic RNAs Link to Inflammation. Front. Immunol. 2020, 11, 590870. [Google Scholar] [CrossRef] [PubMed]
- Winterling, C.; Koch, M.; Koeppel, M.; Garcia-Alcalde, F.; Karlas, A.; Meyer, T.F. Evidence for a Crucial Role of a Host Non-Coding RNA in Influenza A Virus Replication. RNA Biol. 2014, 11, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, L.; Zheng, X.; Wang, G.; Chen, X.; Hu, Z.; Chen, Y.; Wang, X.; Gu, M.; Hu, S.; et al. Long Noncoding RNA #61 Exerts a Broad Anti-Influenza a Virus Effect by Its Long Arm Rings. Antivir. Res. 2023, 215, 105637. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zheng, X.; Li, J.; Wang, G.; Hu, Z.; Chen, Y.; Wang, X.; Gu, M.; Gao, R.; Hu, S.; et al. Long Noncoding RNA#45 Exerts Broad Inhibitory Effect on Influenza a Virus Replication via Its Stem Ring Arms. Virulence 2021, 12, 2443–2460. [Google Scholar] [CrossRef] [PubMed]
- van Solingen, C.; Cyr, Y.; Scacalossi, K.R.; de Vries, M.; Barrett, T.J.; de Jong, A.; Gourvest, M.; Zhang, T.; Peled, D.; Kher, R.; et al. Long Noncoding RNA CHROMR Regulates Antiviral Immunity in Humans. Proc. Natl. Acad. Sci. USA 2022, 119, e2210321119. [Google Scholar] [CrossRef]
- Jiang, J.; Li, Y.; Sun, Z.; Gong, L.; Li, X.; Shi, F.; Yao, J.; Meng, Y.; Meng, X.; Zhang, Q.; et al. LncNSPL Facilitates Influenza A Viral Immune Escape by Restricting TRIM25-Mediated K63-Linked RIG-I Ubiquitination. iScience 2022, 25, 104607. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, J.; Liu, S.; Chen, B.; Xiao, M.; Li, Y.; Liao, Y.; Rai, K.R.; Zhao, Z.; Ouyang, J.; et al. RDUR, a lncRNA, Promotes Innate Antiviral Responses and Provides Feedback Control of NF-κB Activation. Front. Immunol. 2021, 12, 672165. [Google Scholar] [CrossRef]
- Gatherer, D.; Depledge, D.P.; Hartley, C.A.; Szpara, M.L.; Vaz, P.K.; Benkő, M.; Brandt, C.R.; Bryant, N.A.; Dastjerdi, A.; Doszpoly, A.; et al. ICTV Virus Taxonomy Profile: Herpesviridae 2021. J. Gen. Virol. 2021, 102, 001673. [Google Scholar] [CrossRef]
- Woźniakowski, G.; Samorek-Salamonowicz, E. Animal Herpesviruses and Their Zoonotic Potential for Cross-Species Infection. Ann. Agric. Environ. Med. AAEM 2015, 22, 191–194. [Google Scholar] [CrossRef]
- Yang, L.; Maruo, S.; Takada, K. CD21-Mediated Entry and Stable Infection by Epstein-Barr Virus in Canine and Rat Cells. J. Virol. 2000, 74, 10745–10751. [Google Scholar] [CrossRef][Green Version]
- Fang, L.; Gao, Y.; Liu, X.; Bai, J.; Jiang, P.; Wang, X. Long Non-Coding RNA LNC_000641 Regulates Pseudorabies Virus Replication. Vet. Res. 2021, 52, 52. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhang, K.; Huang, W.; Tang, W.; Li, H.; Dong, W.; Gu, J.; Zhou, J. Identification of Functional lncRNAs in Pseudorabies Virus Type II Infected Cells. Vet. Microbiol. 2020, 242, 108564. [Google Scholar] [CrossRef] [PubMed]
- Hotoboc, I.E.; Fudulu, A.; Grigore, R.; Bertesteanu, S.; Huica, I.; Iancu, I.V.; Botezatu, A.; Bleotu, C.; Anton, G. The Association between lncRNA H19 and EZH2 Expression in Patients with EBV-Positive Laryngeal Carcinoma. Acta Otorhinolaryngol. Ital. 2021, 41, 537–543. [Google Scholar] [CrossRef]
- Tombácz, D.; Csabai, Z.; Oláh, P.; Balázs, Z.; Likó, I.; Zsigmond, L.; Sharon, D.; Snyder, M.; Boldogkői, Z. Full-Length Isoform Sequencing Reveals Novel Transcripts and Substantial Transcriptional Overlaps in a Herpesvirus. PLoS ONE 2016, 11, e0162868. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; de Groot, R.J.; Haagmans, B.; Lau, S.K.P.; Neuman, B.W.; Perlman, S.; Sola, I.; van der Hoek, L.; Wong, A.C.P.; Yeh, S.-H. ICTV Virus Taxonomy Profile: Coronaviridae 2023. J. Gen. Virol. 2023, 104, 001843. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Zhao, J.; Li, L.; Huang, X.; Yang, H.; Cheng, J.; Liu, C.; Zhang, G. Pathogenic Characteristics of a QX-like Infectious Bronchitis Virus Strain SD in Chickens Exposed at Different Ages and Protective Efficacy of Combining Live Homologous and Heterologous Vaccination. Vet. Res. 2020, 51, 86. [Google Scholar] [CrossRef]
- Alluwaimi, A.M.; Alshubaith, I.H.; Al-Ali, A.M.; Abohelaika, S. The Coronaviruses of Animals and Birds: Their Zoonosis, Vaccines, and Models for SARS-CoV and SARS-CoV2. Front. Vet. Sci. 2020, 7, 582287. [Google Scholar] [CrossRef]
- Graham, R.L.; Baric, R.S. Recombination, Reservoirs, and the Modular Spike: Mechanisms of Coronavirus Cross-Species Transmission. J. Virol. 2010, 84, 3134–3146. [Google Scholar] [CrossRef]
- Ye, Z.-W.; Yuan, S.; Yuen, K.-S.; Fung, S.-Y.; Chan, C.-P.; Jin, D.-Y. Zoonotic Origins of Human Coronaviruses. Int. J. Biol. Sci. 2020, 16, 1686–1697. [Google Scholar] [CrossRef]
- Tortorici, M.A.; Walls, A.C.; Joshi, A.; Park, Y.-J.; Eguia, R.T.; Miranda, M.C.; Kepl, E.; Dosey, A.; Stevens-Ayers, T.; Boeckh, M.J.; et al. Structure, Receptor Recognition, and Antigenicity of the Human Coronavirus CCoV-HuPn-2018 Spike Glycoprotein. Cell 2022, 185, 2279–2291.e17. [Google Scholar] [CrossRef] [PubMed]
- Ayeldeen, G.; Shaker, O.G.; Amer, E.; Zaafan, M.A.; Herzalla, M.R.; Keshk, M.A.; Abdelhamid, A.M. The Impact of lncRNA-GAS5/miRNA-200/ACE2 Molecular Pathway on the Severity of COVID-19. Curr. Med. Chem. 2024, 31, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Pushparaj, S.; Gandikota, C.; Vaddadi, K.; Liang, Y.; Liu, L. SNHG15 Aids SARS-CoV-2 Entry via RABL2A. RNA Biol. 2023, 20, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Aznaourova, M.; Schmerer, N.; Janga, H.; Zhang, Z.; Pauck, K.; Bushe, J.; Volkers, S.M.; Wendisch, D.; Georg, P.; Ntini, E.; et al. Single-Cell RNA Sequencing Uncovers the Nuclear Decoy lincRNA PIRAT as a Regulator of Systemic Monocyte Immunity during COVID-19. Proc. Natl. Acad. Sci. USA 2022, 119, e2120680119. [Google Scholar] [CrossRef] [PubMed]
- Askari, N.; Hadizadeh, M.; Rashidifar, M. A New Insight into Sex-Specific Non-Coding RNAs and Networks in Response to SARS-CoV-2. Infect. Genet. Evol. 2022, 97, 105195. [Google Scholar] [CrossRef] [PubMed]
- Genena, S.E.S.R.; Fadhil, M.M.; Mansour, M.M.; Attwa, A.H.M.; Khalil, M.M.I.M. Expression Pattern of Long Non-Coding RNAs MALAT1 and MEG3 in COVID-19 Patients. J. Gene Med. 2023, 25, e3532. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Zhou, L.; Wang, H.; Chen, N.; Jia, L.; Wang, C.; Wang, Y.; Chen, J.; Wen, X.; Niu, C.; et al. Profiling the Epigenetic Interplay of lncRNA RUNXOR and Oncogenic RUNX1 in Breast Cancer Cells by Gene in Situ Cis-Activation. Am. J. Cancer Res. 2019, 9, 1635–1649. [Google Scholar] [PubMed]
- Mathison, B.A.; Pritt, B.S. The Landscape of Parasitic Infections in the United States. Mod. Pathol. 2023, 36, 100217. [Google Scholar] [CrossRef] [PubMed]
- Alonso, P.L. The Role of Mass Drug Administration of Antimalarials. Am. J. Trop. Med. Hyg. 2020, 103, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Olajide, J.S.; Olopade, B.; Cai, J. Functional Intricacy and Symmetry of Long Non-Coding RNAs in Parasitic Infections. Front. Cell. Infect. Microbiol. 2021, 11, 751523. [Google Scholar] [CrossRef] [PubMed]
- Leija-Montoya, A.G.; González-Ramírez, J.; Martínez-Coronilla, G.; Mejía-León, M.E.; Isiordia-Espinoza, M.; Sánchez-Muñoz, F.; Chávez-Cortez, E.G.; Pitones-Rubio, V.; Serafín-Higuera, N. Roles of microRNAs and Long Non-Coding RNAs Encoded by Parasitic Helminths in Human Carcinogenesis. Int. J. Mol. Sci. 2022, 23, 8173. [Google Scholar] [CrossRef] [PubMed]
- Garrigós, M.; Ylla, G.; Martínez-de la Puente, J.; Figuerola, J.; Ruiz-López, M.J. Two Avian Plasmodium Species Trigger Different Transcriptional Responses on Their Vector Culex Pipiens. Mol. Ecol. 2023, 00, e17240. [Google Scholar] [CrossRef]
- Batugedara, G.; Lu, X.M.; Hristov, B.; Abel, S.; Chahine, Z.; Hollin, T.; Williams, D.; Wang, T.; Cort, A.; Lenz, T.; et al. Novel Insights into the Role of Long Non-Coding RNA in the Human Malaria Parasite, Plasmodium falciparum. Nat. Commun. 2023, 14, 5086. [Google Scholar] [CrossRef]
- Filarsky, M.; Fraschka, S.A.; Niederwieser, I.; Brancucci, N.M.B.; Carrington, E.; Carrió, E.; Moes, S.; Jenoe, P.; Bártfai, R.; Voss, T.S. GDV1 Induces Sexual Commitment of Malaria Parasites by Antagonizing HP1-Dependent Gene Silencing. Science 2018, 359, 1259–1263. [Google Scholar] [CrossRef]
- Simantov, K.; Goyal, M.; Dzikowski, R. Emerging Biology of Noncoding RNAs in Malaria Parasites. PLoS Pathog. 2022, 18, e1010600. [Google Scholar] [CrossRef]
- Broadbent, K.M.; Park, D.; Wolf, A.R.; Van Tyne, D.; Sims, J.S.; Ribacke, U.; Volkman, S.; Duraisingh, M.; Wirth, D.; Sabeti, P.C.; et al. A Global Transcriptional Analysis of Plasmodium falciparum Malaria Reveals a Novel Family of Telomere-Associated lncRNAs. Genome Biol. 2011, 12, R56. [Google Scholar] [CrossRef]
- Thompson, T.A.; Chahine, Z.; Roch, K.G.L. The Role of Long Noncoding RNAs in Malaria Parasites. Trends Parasitol. 2023, 39, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Amit-Avraham, I.; Pozner, G.; Eshar, S.; Fastman, Y.; Kolevzon, N.; Yavin, E.; Dzikowski, R. Antisense Long Noncoding RNAs Regulate Var Gene Activation in the Malaria Parasite Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2015, 112, E982–E991. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, S.-C.; Fan, X.-Y.; Jin, Y.-L.; Li, X.; Du, Y.-T. Plasmodium Manipulates the Expression of Host Long Non-Coding RNA during Red Blood Cell Intracellular Infection. Parasit. Vectors 2022, 15, 182. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, J.P.; West, K.A.; James, K.R.; Rani, G.F.; Dey, N.; Romano, A.; Brown, N.; Teichmann, S.A.; Kaye, P.M.; Lagos, D. Malat1 Suppresses Immunity to Infection through Promoting Expression of Maf and IL-10 in Th Cells. J. Immunol. 2020, 204, 2949–2960. [Google Scholar] [CrossRef] [PubMed]
- McManus, D.P.; Dunne, D.W.; Sacko, M.; Utzinger, J.; Vennervald, B.J.; Zhou, X.-N. Schistosomiasis. Nat. Rev. Dis. Primer 2018, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Silveira, G.O.; Coelho, H.S.; Pereira, A.S.A.; Miyasato, P.A.; Santos, D.W.; Maciel, L.F.; Olberg, G.G.G.; Tahira, A.C.; Nakano, E.; Oliveira, M.L.S.; et al. Long Non-Coding RNAs Are Essential for Schistosoma mansoni Pairing-Dependent Adult Worm Homeostasis and Fertility. PLoS Pathog. 2023, 19, e1011369. [Google Scholar] [CrossRef]
- Amaral, M.S.; Maciel, L.F.; Silveira, G.O.; Olberg, G.G.O.; Leite, J.V.P.; Imamura, L.K.; Pereira, A.S.A.; Miyasato, P.A.; Nakano, E.; Verjovski-Almeida, S. Long Non-Coding RNA Levels Can Be Modulated by 5-Azacytidine in Schistosoma mansoni. Sci. Rep. 2020, 10, 21565. [Google Scholar] [CrossRef]
- Liao, Q.; Zhang, Y.; Zhu, Y.; Chen, J.; Dong, C.; Tao, Y.; He, A.; Liu, J.; Wu, Z. Identification of Long Noncoding RNAs in Schistosoma mansoni and Schistosoma japonicum. Exp. Parasitol. 2018, 191, 82–87. [Google Scholar] [CrossRef]
- Xia, T.; Giri, B.R.; Liu, J.; Du, P.; Li, X.; Li, X.; Li, S.; Cheng, G. RNA Sequencing Analysis of Altered Expression of Long Noncoding RNAs Associated with Schistosoma japonicum Infection in the Murine Liver and Spleen. Parasit. Vectors 2020, 13, 601. [Google Scholar] [CrossRef]
- Zhao, R.; Tang, X.; Lin, H.; Xing, C.; Xu, N.; Dai, B.; Wang, P.; Shao, W.; Liu, M.; Shen, J.; et al. Knocking Down Gm16685 Decreases Liver Granuloma in Murine Schistosomiasis japonica. Microorganisms 2023, 11, 796. [Google Scholar] [CrossRef]
- Ma, R.; Liu, Q.; Liu, Z.; Sun, X.; Jiang, X.; Hou, J.; Zhang, Y.; Wu, Y.; Cheng, M.; Dong, Z. H19/Mir-130b-3p/Cyp4a14 Potentiate the Effect of Praziquantel on Liver in the Treatment of Schistosoma japonicum Infection. Acta Trop. 2023, 247, 107012. [Google Scholar] [CrossRef] [PubMed]
- Kurniawan, A.; Dwintasari, S.W.; Connelly, L.; Nichols, R.A.B.; Yunihastuti, E.; Karyadi, T.; Djauzi, S. Cryptosporidium Species from Human Immunodeficiency-Infected Patients with Chronic Diarrhea in Jakarta, Indonesia. Ann. Epidemiol. 2013, 23, 720–723. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.L.; Li, M.; Gong, A.-Y.; Deng, S.; Jin, K.; Wang, S.; Chen, X.-M. Cryptosporidium parvum Hijacks a Host’s Long Noncoding RNA U90926 to Evade Intestinal Epithelial Cell-Autonomous Antiparasitic Defense. Front. Immunol. 2023, 14, 1205468. [Google Scholar] [CrossRef] [PubMed]
- Mathy, N.W.; Deng, S.; Gong, A.-Y.; Li, M.; Wang, Y.; Burleigh, O.; Kochvar, A.; Whiteford, E.R.; Shibata, A.; Chen, X.-M. The Long Non-Coding RNA Nostrill Regulates Transcription of Irf7 Through Interaction With NF-κB P65 to Enhance Intestinal Epithelial Defense against Cryptosporidium parvum. Front. Immunol. 2022, 13, 863957. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Gong, A.-Y.; Zhang, X.-T.; Wang, Y.; Mathy, N.W.; Martins, G.A.; Strauss-Soukup, J.K.; Chen, X.-M. Induction of a Long Noncoding RNA Transcript, NR_045064, Promotes Defense Gene Transcription and Facilitates Intestinal Epithelial Cell Responses against Cryptosporidium Infection. J. Immunol. 2018, 201, 3630–3640. [Google Scholar] [CrossRef] [PubMed]
- Gong, A.-Y.; Wang, Y.; Li, M.; Zhang, X.-T.; Deng, S.; Chen, J.M.; Lu, E.; Mathy, N.W.; Martins, G.A.; Strauss-Soukup, J.K.; et al. LncRNA XR_001779380 Primes Epithelial Cells for IFN-γ-Mediated Gene Transcription and Facilitates Age-Dependent Intestinal Antimicrobial Defense. mBio 2021, 12, e0212721. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jin, K.; Li, M.; Mathy, N.W.; Gong, A.-Y.; Deng, S.; Martins, G.A.; Sun, M.; Strauss-Soukup, J.K.; Chen, X.-M. A Host Cell Long Noncoding RNA NR_033736 Regulates Type I Interferon-Mediated Gene Transcription and Modulates Intestinal Epithelial Anti-Cryptosporidium Defense. PLoS Pathog. 2021, 17, e1009241. [Google Scholar] [CrossRef] [PubMed]
- Ben-Harari, R.R.; Goodwin, E.; Casoy, J. Adverse Event Profile of Pyrimethamine-Based Therapy in Toxoplasmosis: A Systematic Review. Drugs R&D 2017, 17, 523–544. [Google Scholar] [CrossRef]
- Menard, K.L.; Haskins, B.E.; Colombo, A.P.; Denkers, E.Y. Toxoplasma gondii Manipulates Expression of Host Long Noncoding RNA during Intracellular Infection. Sci. Rep. 2018, 8, 15017. [Google Scholar] [CrossRef]
- Wang, Y.; Han, R.; Xu, Z.; Sun, X.; Zhou, C.; Han, B.; He, S.; Cong, H. Upregulation of lncRNA147410.3 in the Brain of Mice with Chronic Toxoplasma Infection Promoted Microglia Apoptosis by Regulating Hoxb3. Front. Cell. Neurosci. 2021, 15, 648047. [Google Scholar] [CrossRef]
- Sun, X.; Wang, T.; Wang, Y.; Ai, K.; Pan, G.; Li, Y.; Zhou, C.; He, S.; Cong, H. Downregulation of lncRNA-11496 in the Brain Contributes to Microglia Apoptosis via Regulation of Mef2c in Chronic T. gondii Infection Mice. Front. Mol. Neurosci. 2020, 13, 77. [Google Scholar] [CrossRef]
- Liu, W.; Huang, L.; Wei, Q.; Zhang, Y.; Zhang, S.; Zhang, W.; Cai, L.; Liang, S. Microarray Analysis of Long Non-Coding RNA Expression Profiles Uncovers a Toxoplasma-Induced Negative Regulation of Host Immune Signaling. Parasit. Vectors 2018, 11, 174. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Vuitton, L.; Tuxun, T.; Li, J.; Vuitton, D.A.; Zhang, W.; McManus, D.P. Echinococcosis: Advances in the 21st Century. Clin. Microbiol. Rev. 2019, 32, e00075-18. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, H.; Li, Y.; Wang, L.; Chen, G.; Pu, G.; Guo, X.; Cho, W.C.; Fasihi Harandi, M.; Zheng, Y.; et al. Integrative Analysis of RNA Expression and Regulatory Networks in Mice Liver Infected by Echinococcus multilocularis. Front. Cell Dev. Biol. 2022, 10, 798551. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Wang, Y.; Yin, J.; Zhang, J.; Cao, S.; Cao, J.; Shen, Y. Microarray Analysis of Long Non-Coding RNA Expression Profiles in Monocytic Myeloid-Derived Suppressor Cells in Echinococcus granulosus-Infected Mice. Parasit. Vectors 2018, 11, 327. [Google Scholar] [CrossRef]
- Yang, R.; Qu, X.; Zhi, S.; Wang, J.; Fu, J.; Tan, C.; Chen, H.; Wang, X. Exosomes Derived from Meningitic Escherichia Coli-Infected Brain Microvascular Endothelial Cells Facilitate Astrocyte Activation. Mol. Neurobiol. 2024. [Google Scholar] [CrossRef]
- Wang, C.; Yang, S.-H.; Niu, N.; Tao, J.; Du, X.-C.; Yang, J.-H.; Zhu, M.-X.; Wang, Y.-N.; Zhao, W. lncRNA028466 Regulates Th1/Th2 Cytokine Expression and Associates with Echinococcus granulosus Antigen P29 Immunity. Parasit. Vectors 2021, 14, 295. [Google Scholar] [CrossRef]
- Yang, R.; Wang, J.; Wang, F.; Zhang, H.; Tan, C.; Chen, H.; Wang, X. Blood-Brain Barrier Integrity Damage in Bacterial Meningitis: The Underlying Link, Mechanisms, and Therapeutic Targets. Int. J. Mol. Sci. 2023, 24, 2852. [Google Scholar] [CrossRef]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-Borne Virus Diversity, Spillover and Emergence. Nat. Rev. Microbiol. 2020, 18, 461–471. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, B.; He, Y.; Yang, R.; Li, J.; Wang, X. Unveiling the Hidden Regulators: The Impact of lncRNAs on Zoonoses. Int. J. Mol. Sci. 2024, 25, 3539. https://doi.org/10.3390/ijms25063539
Xu B, He Y, Yang R, Li J, Wang X. Unveiling the Hidden Regulators: The Impact of lncRNAs on Zoonoses. International Journal of Molecular Sciences. 2024; 25(6):3539. https://doi.org/10.3390/ijms25063539
Chicago/Turabian StyleXu, Bojie, Yujuan He, Ruicheng Yang, Junmin Li, and Xiangru Wang. 2024. "Unveiling the Hidden Regulators: The Impact of lncRNAs on Zoonoses" International Journal of Molecular Sciences 25, no. 6: 3539. https://doi.org/10.3390/ijms25063539
APA StyleXu, B., He, Y., Yang, R., Li, J., & Wang, X. (2024). Unveiling the Hidden Regulators: The Impact of lncRNAs on Zoonoses. International Journal of Molecular Sciences, 25(6), 3539. https://doi.org/10.3390/ijms25063539