
Microcurrent-Mediated Modulation of Myofibroblasts for Cardiac Repair and Regeneration

 , ,
, ,
Abstract
1. Introduction
2. Results
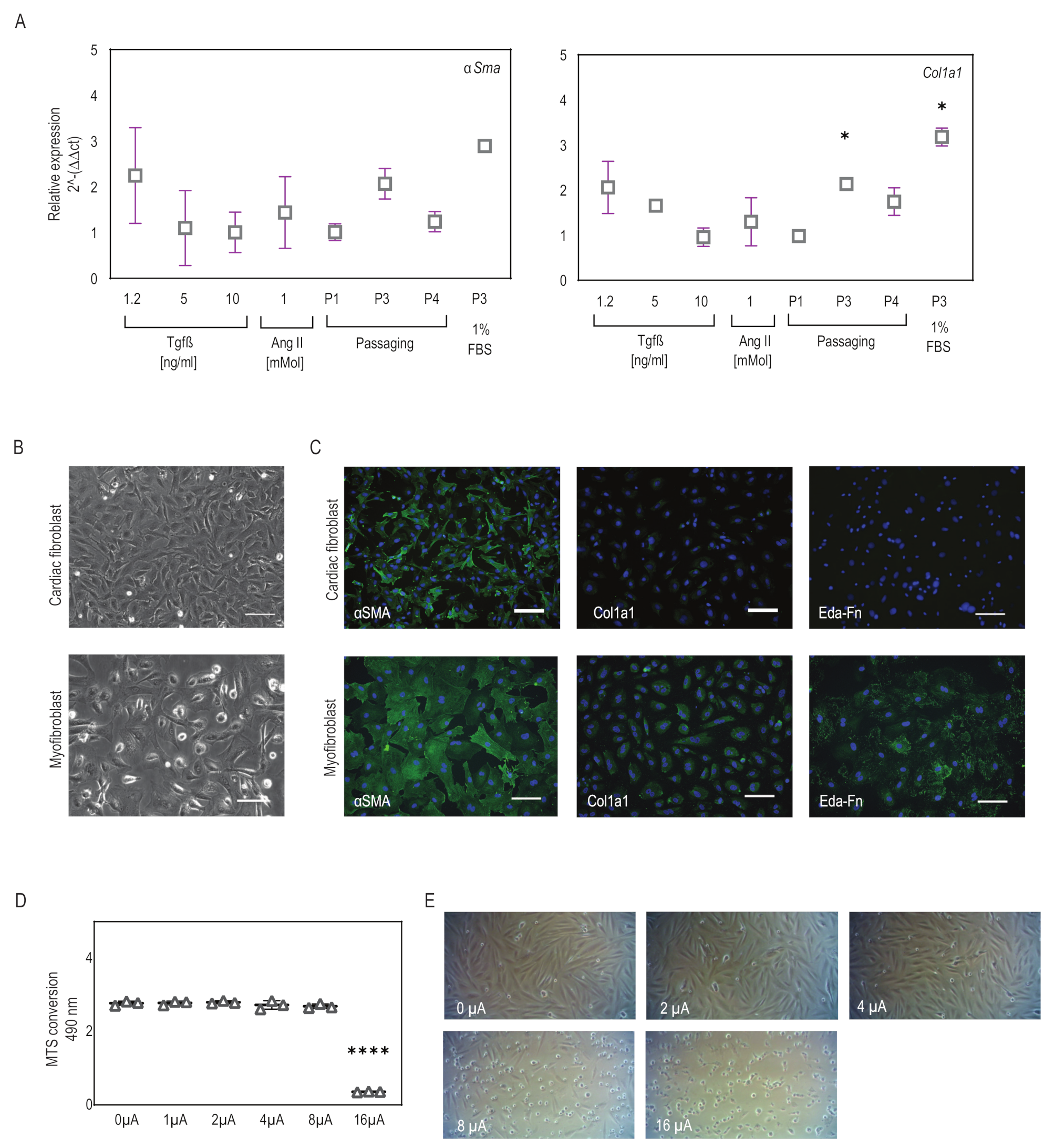
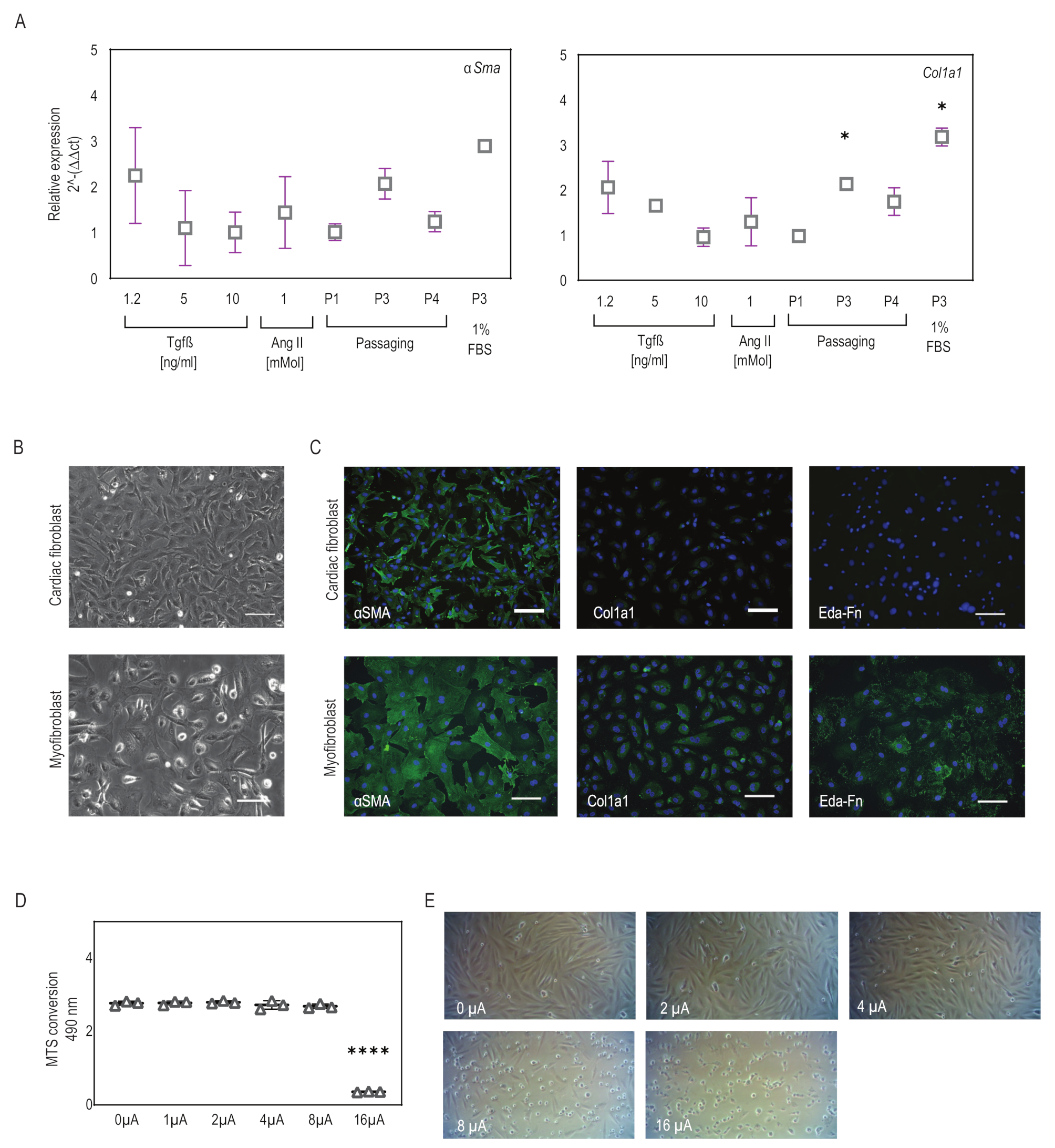
2.1. Myofibroblasts Differentiate from Fibroblasts Efficiently Using a Combination of Low-Serum Medium and Passaging
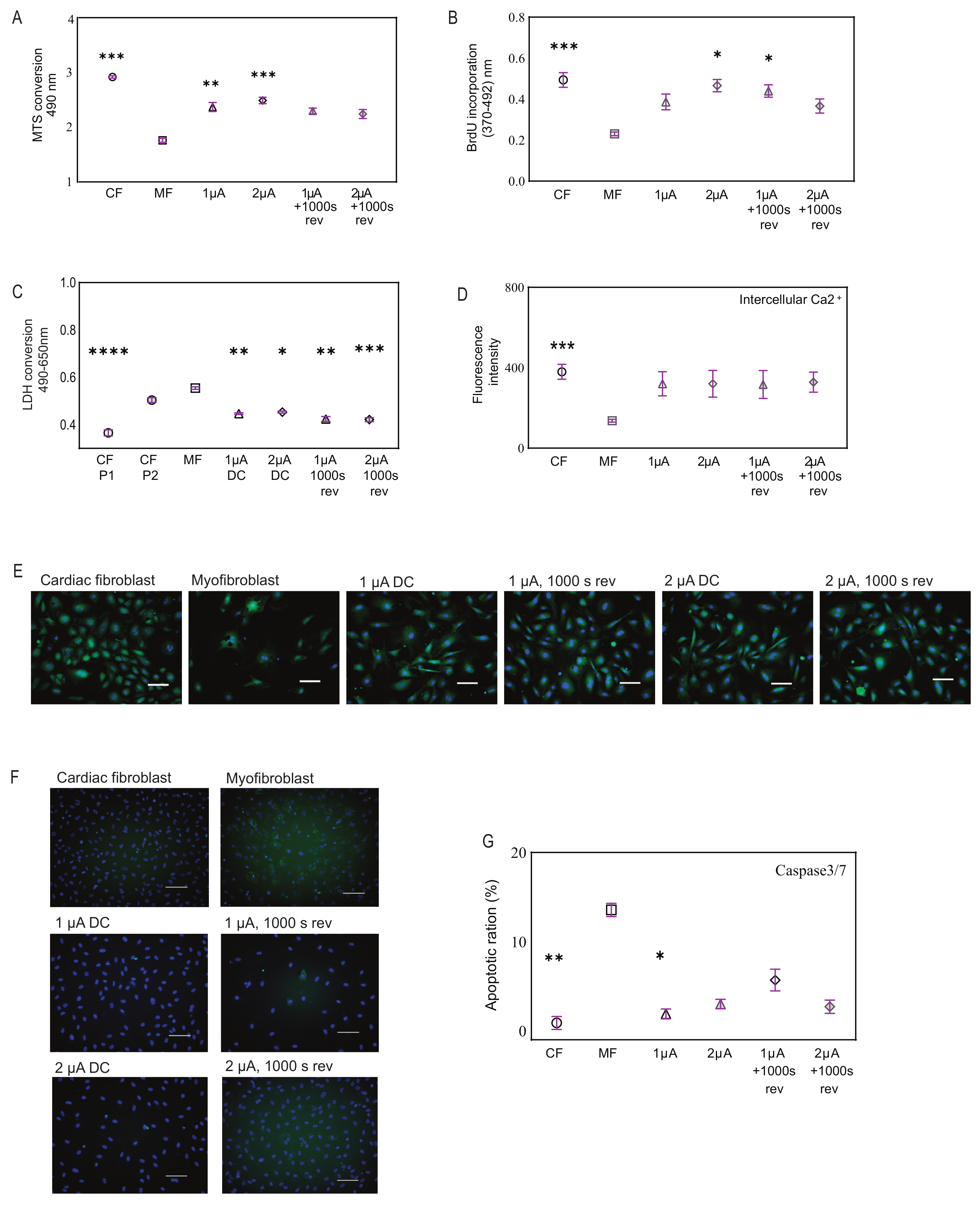
2.2. Microcurrent Treatment
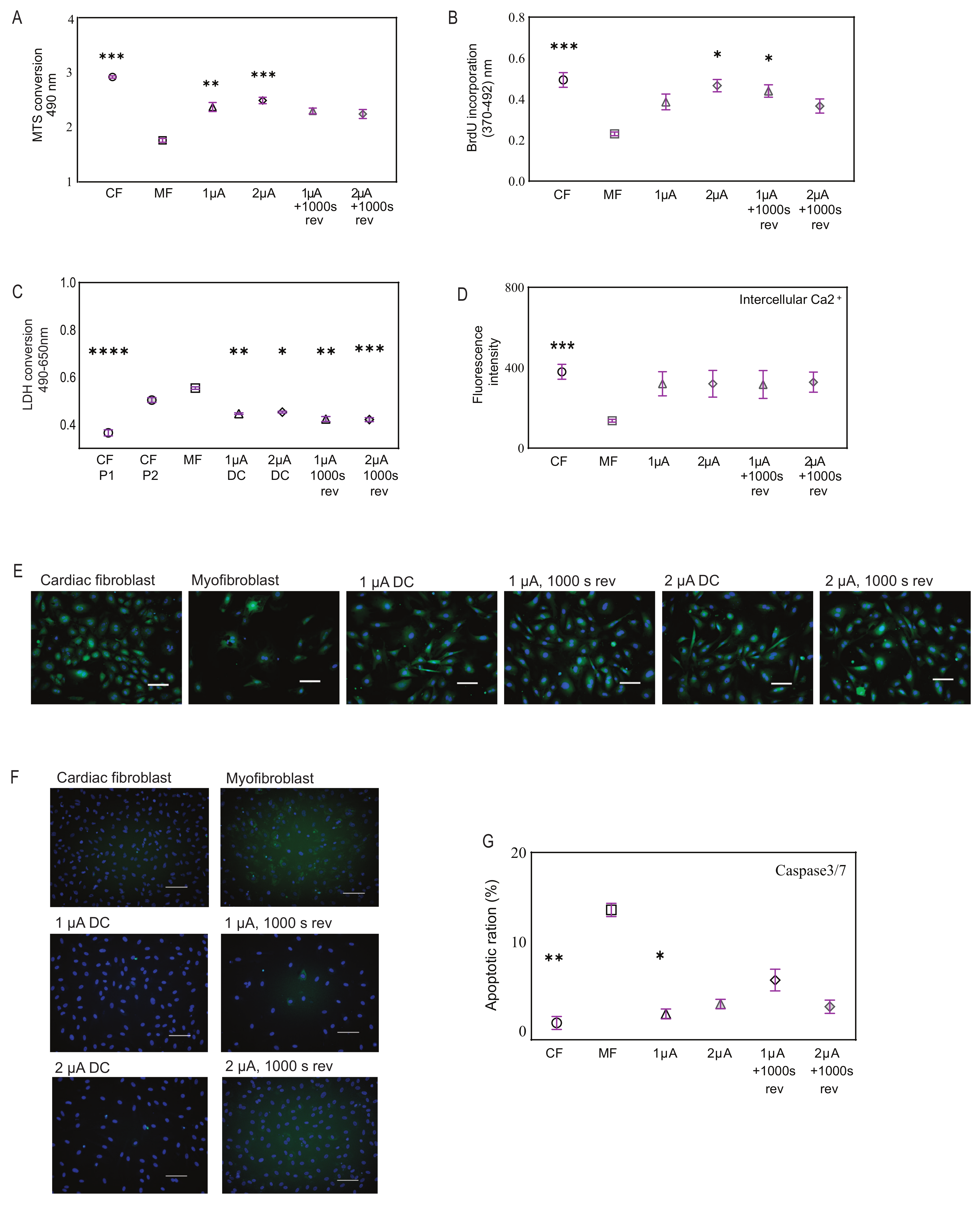
2.3. Microcurrent Treatment Enhances Cell Viability, Restores Calcium Signaling, and Modulates Apoptosis
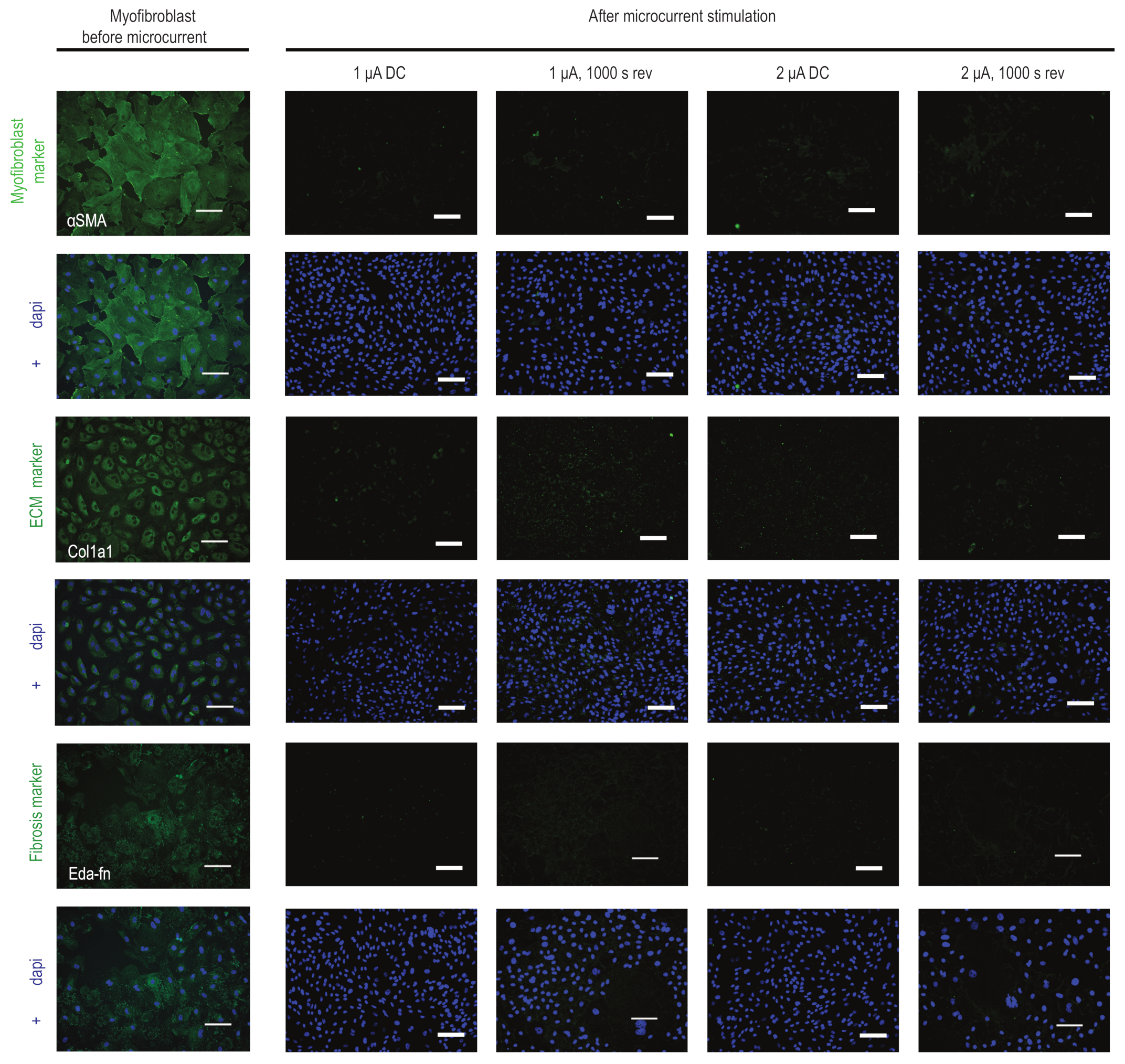
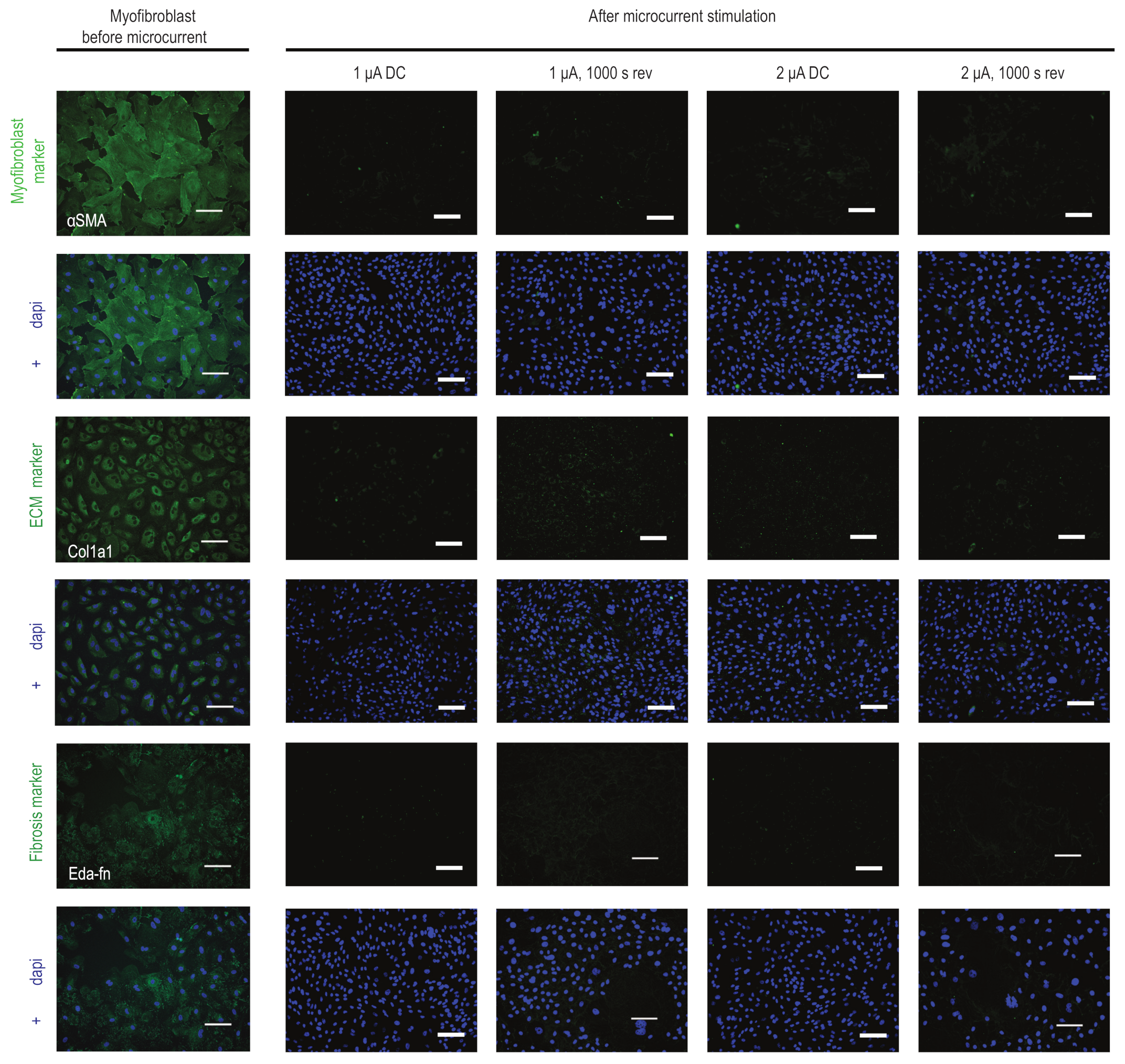
2.4. Microcurrent Treatment Alters the Myofibroblast Phenotype
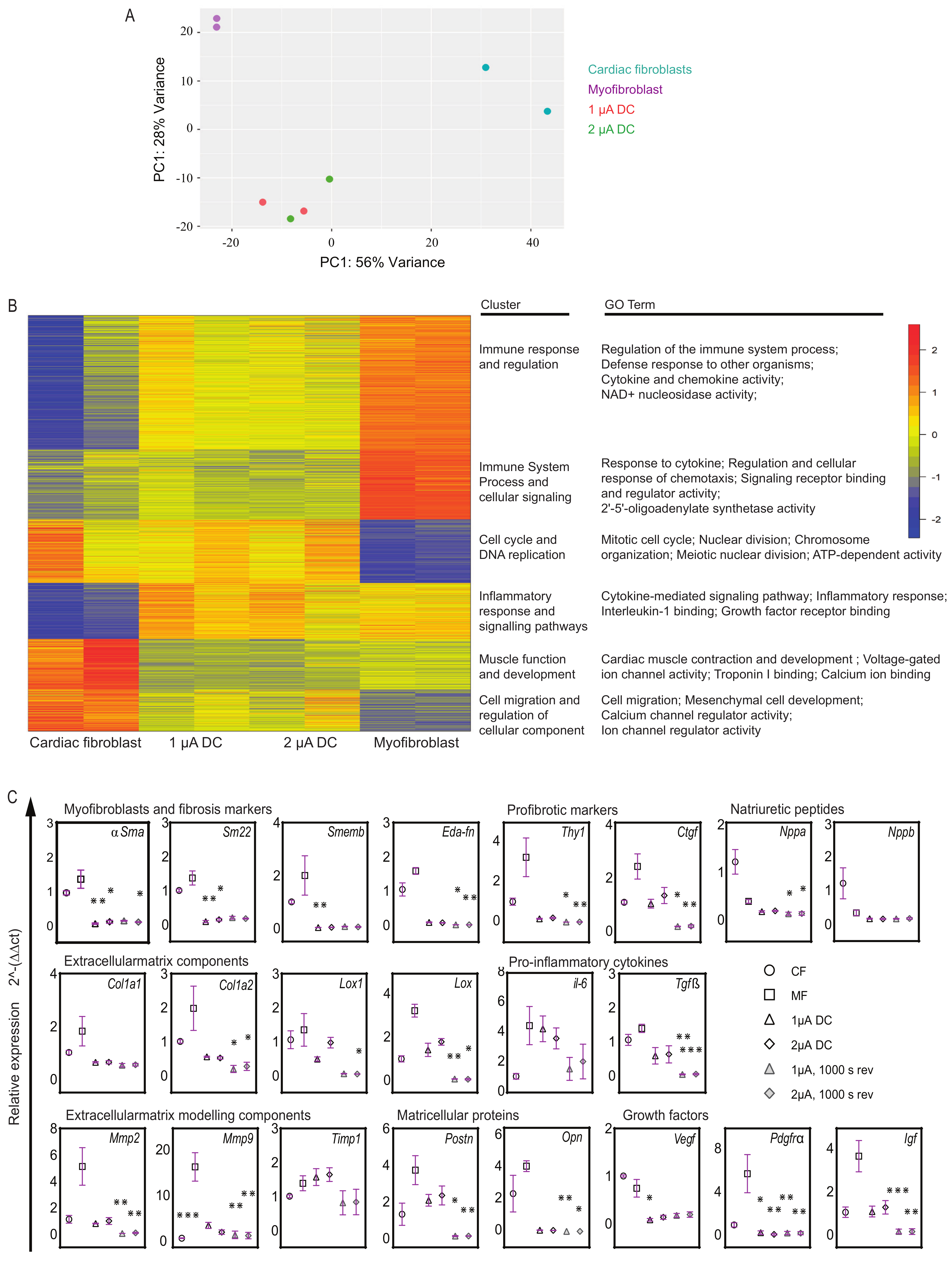
2.5. Transcriptome Analysis Reveals Unique Gene Expression Signatures and Cellular Responses in Microcurrent-Treated Cells
2.6. Microcurrent Treatment Modulates Gene Expression Profiles in Myofibroblasts
3. Materials and Methods
3.1. Cell Culture
3.2. Cardiac Fibroblast to Myofibroblast Differentiation
3.3. Microcurrent Equipment and Treatment
3.4. Cell Behavior
- (i).
- MTS assay
- (ii).
- BrdU Proliferation assay
- (iii).
- LDH cytotoxicity assay
- (iv).
- Calcium analysis
- (v).
- Caspase3/7 activity assay
3.5. Immunocytology of Adherent Cells and Fluorescence Microscopy
3.6. RNA Isolation, RNA Sequencing, and Bioinformatics Analysis
3.7. Reverse Transcription-Quantitative PCR
3.8. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Konstam, M.A.; Kramer, D.G.; Patel, A.R.; Maron, M.S.; Udelson, J.E. Left ventricular remodeling in heart failure: Current concepts in clinical significance and assessment. JACC Cardiovasc. Imaging 2011, 4, 98–108. [Google Scholar] [CrossRef]
- Baum, J.; Duffy, H.S. Fibroblasts and myofibroblasts: What are we talking about? J. Cardiovasc. Pharmacol. 2011, 57, 376–379. [Google Scholar] [CrossRef]
- Hinz, B. The role of myofibroblasts in wound healing. Curr. Res. Transl. Med. 2016, 64, 171–177. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Q.; Kong, W. Extracellular matrix remodeling and cardiac fibrosis. Matrix Biol. 2018, 68–69, 490–506. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Cardiac fibrosis: Cell biological mechanisms, molecular pathways and therapeutic opportunities. Mol. Asp. Med. 2019, 65, 70–99. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Olson, E.N.; Bassel-Duby, R. Therapeutic approaches for cardiac regeneration and repair. Nat. Rev. Cardiol. 2018, 15, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Murry, C.E. Function Follows Form—A Review of Cardiac Cell Therapy. Circ. J. 2019, 83, 2399–2412. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, Y.; Liu, J.; Qian, L. Direct cell reprogramming: Approaches, mechanisms and progress. Nat. Rev. Mol. Cell Biol. 2021, 22, 410–424. [Google Scholar] [CrossRef]
- Zhao, S.; Mehta, A.S.; Zhao, M. Biomedical applications of electrical stimulation. Cell Mol. Life Sci. 2020, 77, 2681–2699. [Google Scholar] [CrossRef]
- Balint, R.; Cassidy, N.J.; Cartmell, S.H. Electrical Stimulation: A Novel Tool for Tissue Engineering. Tissue Eng. Part B Rev. 2012, 19, 48–57. [Google Scholar] [CrossRef]
- Kloth, L.C. Electrical stimulation for wound healing: A review of evidence from in vitro studies, animal experiments, and clinical trials. Int. J. Low. Extrem. Wounds 2005, 4, 23–44. [Google Scholar] [CrossRef]
- Kapeller, B.; Mueller, J.; Losert, U.; Podesser, B.K.; Macfelda, K. Microcurrent stimulation promotes reverse remodelling in cardiomyocytes. ESC Heart Fail. 2016, 3, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Macfelda, K.; Kapeller, B.; Holly, A.; Podesser, B.K.; Losert, U.; Brandes, K.; Goettel, P.; Mueller, J. Bioelectrical signals improve cardiac function and modify gene expression of extracellular matrix components. ESC Heart Fail. 2017, 4, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Schmitto, J.D.; Napp, L.C.; Mariani, S.; Hanke, J.S.; Li, T.; Vogel-Claussen, J.; Brandes, K.; Göttel, P.; Müller, J.; Dogan, G.; et al. First-in-man Implantation of a Cardiac Microcurrent Device for Chronic Systolic Heart Failure. ASAIO J. 2022, 68, e121–e123. [Google Scholar] [CrossRef]
- Kosevic, D.; Wiedemann, D.; Vukovic, P.; Ristic, V.; Riebandt, J.; Radak, U.; Brandes, K.; Goettel, P.; Duengen, H.D.; Tahirovic, E.; et al. Cardio-microcurrent device for chronic heart failure: First-in-human clinical study. ESC Heart Fail. 2021, 8, 962–970. [Google Scholar] [CrossRef]
- Haberland, A.; Wallukat, G.; Dahmen, C.; Kage, A.; Schimke, I. Aptamer neutralization of beta1-adrenoceptor autoantibodies isolated from patients with cardiomyopathies. Circ. Res. 2011, 109, 986–992. [Google Scholar] [CrossRef]
- Ehler, E.; Moore-Morris, T.; Lange, S. Isolation and culture of neonatal mouse cardiomyocytes. J. Vis. Exp. 2013, 79, 50154. [Google Scholar] [CrossRef]
- Cho, N.; Razipour, S.E.; McCain, M.L. Featured Article: TGF-β1 dominates extracellular matrix rigidity for inducing differentiation of human cardiac fibroblasts to myofibroblasts. Exp. Biol. Med. 2018, 243, 601–612. [Google Scholar] [CrossRef]
- Juan, G.; Zoog, S.J.; Ferbas, J. Leveraging image cytometry for the development of clinically feasible biomarkers: Evaluation of activated caspase-3 in fine needle aspirate biopsies. Methods Cell Biol. 2011, 102, 309–320. [Google Scholar] [CrossRef]
- Bravo, M.P.; Soares, G.P.; Daniele de Oliveira, P.; Szezerbaty, S.K.; Frederico, R.C.P.; Maia, L.P. Microcurrent stimulates cell proliferation and modulates cytokine release in fibroblast cells. J. Wound Care 2021, 30, IIIi–IIIix. [Google Scholar] [CrossRef]
- Li, Y.; Wei, L.; Lan, L.; Gao, Y.; Zhang, Q.; Dawit, H.; Mao, J.; Guo, L.; Shen, L.; Wang, L. Conductive biomaterials for cardiac repair: A review. Acta Biomater. 2021, 139, 157–178. [Google Scholar] [CrossRef]
- Liu, M.; López de Juan Abad, B.; Cheng, K. Cardiac fibrosis: Myofibroblast-mediated pathological regulation and drug delivery strategies. Adv. Drug Deliv. Rev. 2021, 173, 504–519. [Google Scholar] [CrossRef]
- Thomas, T.P.; Grisanti, L.A. The Dynamic Interplay Between Cardiac Inflammation and Fibrosis. Front. Physiol. 2020, 11, 529075. [Google Scholar] [CrossRef]
- Feng, J.; Armillei, M.K.; Yu, A.S.; Liang, B.T.; Runnels, L.W.; Yue, L. Ca2+ Signaling in Cardiac Fibroblasts and Fibrosis-Associated Heart Diseases. J. Cardiovasc. Dev. Dis. 2019, 6, 34. [Google Scholar] [CrossRef]
- Grotendorst, G.R.; Rahmanie, H.; Duncan, M.R. Combinatorial signaling pathways determine fibroblast proliferation and myofibroblast differentiation. FASEB J. 2004, 18, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.-J.; Dangerfield, A.L.; Rattan, S.G.; Bathe, K.L.; Cunnington, R.H.; Raizman, J.E.; Bedosky, K.M.; Freed, D.H.; Kardami, E.; Dixon, I.M.C. Cardiac fibroblast to myofibroblast differentiation in vivo and in vitro: Expression of focal adhesion components in neonatal and adult rat ventricular myofibroblasts. Dev. Dyn. 2010, 239, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.A.; Porter, K.E. Function and fate of myofibroblasts after myocardial infarction. Fibrogenesis Tissue Repair 2013, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Tarbit, E.; Singh, I.; Peart, J.N.; Rose’Meyer, R.B. Biomarkers for the identification of cardiac fibroblast and myofibroblast cells. Heart Fail. Rev. 2019, 24, 1–15. [Google Scholar] [CrossRef]
- Creemers, E.E.; Cleutjens, J.P.; Smits, J.F.; Daemen, M.J. Matrix metalloproteinase inhibition after myocardial infarction: A new approach to prevent heart failure? Circ. Res. 2001, 89, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Szóstek-Mioduchowska, A.Z.; Lukasik, K.; Skarzynski, D.J.; Okuda, K. Effect of transforming growth factor-β1 on α-smooth muscle actin and collagen expression in equine endometrial fibroblasts. Theriogenology 2019, 124, 9–17. [Google Scholar] [CrossRef]
- Davis, J.; Molkentin, J.D. Myofibroblasts: Trust your heart and let fate decide. J. Mol. Cell Cardiol. 2014, 70, 9–18. [Google Scholar] [CrossRef]
- Edwards, D.R.; Murphy, G.; Reynolds, J.J.; Whitham, S.E.; Docherty, A.J.; Angel, P.; Heath, J.K. Transforming growth factor beta modulates the expression of collagenase and metalloproteinase inhibitor. EMBO J. 1987, 6, 1899–1904. [Google Scholar] [CrossRef]
- Desmoulière, A.; Redard, M.; Darby, I.; Gabbiani, G. Apoptosis mediates the decrease in cellularity during the transition between granulation tissue and scar. Am. J. Pathol. 1995, 146, 56–66. [Google Scholar] [PubMed]
- Uemura, M.; Sugimoto, M.; Yoshikawa, Y.; Hiramatsu, T.; Inoue, T. Monophasic Pulsed Current Stimulation of Duty Cycle 10% Promotes Differentiation of Human Dermal Fibroblasts into Myofibroblasts. Phys. Ther. Res. 2021, 24, 145–152. [Google Scholar] [CrossRef]
- Silva, D.F.D.; de Oliveira Fujii, L.; Chiarotto, G.B.; de Oliveira, C.A.; de Andrade, T.A.M.; de Oliveira, A.L.R.; Esquisatto, M.A.M.; Mendonça, F.A.S.; Dos Santos, G.M.T.; de Aro, A.A. Influence of microcurrent on the modulation of remodelling genes in a wound healing assay. Mol. Biol. Rep. 2021, 48, 1233–1241. [Google Scholar] [CrossRef]
- Rouabhia, M.; Park, H.; Meng, S.; Derbali, H.; Zhang, Z. Electrical stimulation promotes wound healing by enhancing dermal fibroblast activity and promoting myofibroblast transdifferentiation. PLoS ONE 2013, 8, e71660. [Google Scholar] [CrossRef]
- Rouabhia, M.; Park, H.J.; Abedin-Do, A.; Douville, Y.; Méthot, M.; Zhang, Z. Electrical stimulation promotes the proliferation of human keratinocytes, increases the production of keratin 5 and 14, and increases the phosphorylation of ERK1/2 and p38 MAP kinases. J. Tissue Eng. Regen. Med. 2020, 14, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Wolcott, L.E.; Wheeler, P.C.; Hardwicke, H.M.; Rowley, B.A. Accelerated healing of skin ulcer by electrotherapy: Preliminary clinical results. S. Med. J. 1969, 62, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Carley, P.J.; Wainapel, S.F. Electrotherapy for acceleration of wound healing: Low intensity direct current. Arch. Phys. Med. Rehabil. 1985, 66, 443–446. [Google Scholar]
- Hinz, B. Formation and function of the myofibroblast during tissue repair. J. Investig. Dermatol. 2007, 127, 526–537. [Google Scholar] [CrossRef]
- Feaster, T.K.; Ewoldt, J.K.; Avila, A.; Casciola, M.; Narkar, A.; Chen, C.S.; Blinova, K. Nonclinical evaluation of chronic cardiac contractility modulation on 3D human engineered cardiac tissues. J. Cardiovasc. Electrophysiol. 2024, 1–11. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunogen, Conjugate | Host | Dilution Factor | Product Details |
|---|---|---|---|
| Primary antibodies | |||
| α-SMA (Anti-α smooth muscle) | Mouse | 1:200 | A2547, Sigma Aldrich |
| Col1a1 (Anti-Collagen Type I) | Rabbit | 1:200 | 234167, Sigma Aldrich |
| Eda-Fn (Anti-Fibronectin (EDA)) | Mouse | 1:200 | AG-20B-6001PF-C100 AdipoGen |
| Secondary antibodies | |||
| Donkey anti-Mouse IgG, Alexa Fluor 488 | Donkey | 1:1000 | A-21202, Invitrogen |
| Donkey anti-Rabbit IgG, Alexa Fluor 488 | Donkey | 1:1000 | A-21206, Invitrogen |
| Gene | Forward Primer 5′ → 3′ | Reverse Primer 5′ → 3′ |
|---|---|---|
| αSma | ACCTTCAATGTCCCTGCCATGTA | ACGAAGGAATAGCCACGCTCA |
| ßAct | GGACCTGACAGACTACCTCA | GTTGCCAATAGTGATGACCT |
| Col1a1 | CCCTAATGGTGAGACGTGGA | CTTGGGTCCCTCGACTCCTA |
| Col1a2 | CAGCTCCACTCTCACCTG | CAAGCCGGGAGAAAGGG |
| Ctgf | TGACCTGGAGGAAAACATTAAGA | AGCCCTGTATGTCTTCACACTG |
| Eda-fn | GGCAAGTTTCCAGGTACAGG | GCAAGGCAACCACACTGACT |
| Gapdh | AGACAGCCGCATCTTCTTGT | CTTGCCGTGGGTAGAGTCAT |
| Igf | ATCCGGCGAGGCAATAACAT | ACGGATGTGGTCGTTTTCCA |
| Il-6 | CCGGAGAGGAGACTTCACAG | ACAGTGCATCATCGCTGTTC |
| Lox | GGCACCGACCTGGATATGGCACC | CGGTGAAATGGTGCAGCCTGAGG |
| Lox1 | CGTCGTTACTCGGCATAGCCT | CCATGCTGTGGTAATGTTGGTG |
| Mmp2 | ATCTGGTGTCTCCCTTACGG | GTGCAGTGATGTCCGACAAC |
| Mmp9 | CTTCGAGGGCCACTCCTACT | CAGTGACGTCGGCTCGAGT |
| Nppa | CAGGGCCTCAAGGCACTTTT | GGTGGTCTAGCAGGTTCTTGAAA |
| Nppb | CAGCTCTCAAAGGACCAAGG | CGGTCTATCTTCTGCCCAAA |
| Opn | GAAGCCAGCCAAGGTAAGC | CACTGCCAGTCTCATGGTTG |
| Postn | ACACGGCATGGTTATTCCTTC | GAAGTCTTGGATGGAGGTGC |
| Pdgfr-α | AGGCTTGGGGCTCACTTTTT | CTCGGCCCTGTGAGGAGA |
| Sm22 | AAGCAGATGGAACAGGTGGC | TGCCCAAAGCCATTACAGTCC |
| SMemb | GTGAAGCCTCTCCTCCAAGTG | TCCGTTCCATCTCCTCAAGTTC |
| Tgf-ß | AACATGCACACCCCCAAGAT | CAGGGCCTCAAGGCACTTTT |
| Timp1 | ACGCTAGAGCAGATACCACG | AGCGTCGAATCCTTTGAGCA |
| Thy1 | TCCTGCTTTCAGTCTTGCAG | TCATGCTGGATGGGCAAGTT |
| Vegf | GCCCTGAGTCAAGAGGACAG | CAGGCTCCTGATTCTTCCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bachamanda Somesh, D.; Jürchott, K.; Giesel, T.; Töllner, T.; Prehn, A.; Richters, J.-P.; Kosevic, D.; Eduardo Rame, J.; Göttel, P.; Müller, J. Microcurrent-Mediated Modulation of Myofibroblasts for Cardiac Repair and Regeneration. Int. J. Mol. Sci. 2024, 25, 3268. https://doi.org/10.3390/ijms25063268
Bachamanda Somesh D, Jürchott K, Giesel T, Töllner T, Prehn A, Richters J-P, Kosevic D, Eduardo Rame J, Göttel P, Müller J. Microcurrent-Mediated Modulation of Myofibroblasts for Cardiac Repair and Regeneration. International Journal of Molecular Sciences. 2024; 25(6):3268. https://doi.org/10.3390/ijms25063268
Chicago/Turabian StyleBachamanda Somesh, Dipthi, Karsten Jürchott, Thomas Giesel, Thomas Töllner, Alexander Prehn, Jan-Peter Richters, Dragana Kosevic, Jesus Eduardo Rame, Peter Göttel, and Johannes Müller. 2024. "Microcurrent-Mediated Modulation of Myofibroblasts for Cardiac Repair and Regeneration" International Journal of Molecular Sciences 25, no. 6: 3268. https://doi.org/10.3390/ijms25063268
APA StyleBachamanda Somesh, D., Jürchott, K., Giesel, T., Töllner, T., Prehn, A., Richters, J.-P., Kosevic, D., Eduardo Rame, J., Göttel, P., & Müller, J. (2024). Microcurrent-Mediated Modulation of Myofibroblasts for Cardiac Repair and Regeneration. International Journal of Molecular Sciences, 25(6), 3268. https://doi.org/10.3390/ijms25063268