
Mitochondrial Ribosomal Protein MRPS15 Is a Component of Cytosolic Ribosomes and Regulates Translation in Stressed Cardiomyocytes

 , , , , ,
, , , , ,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results
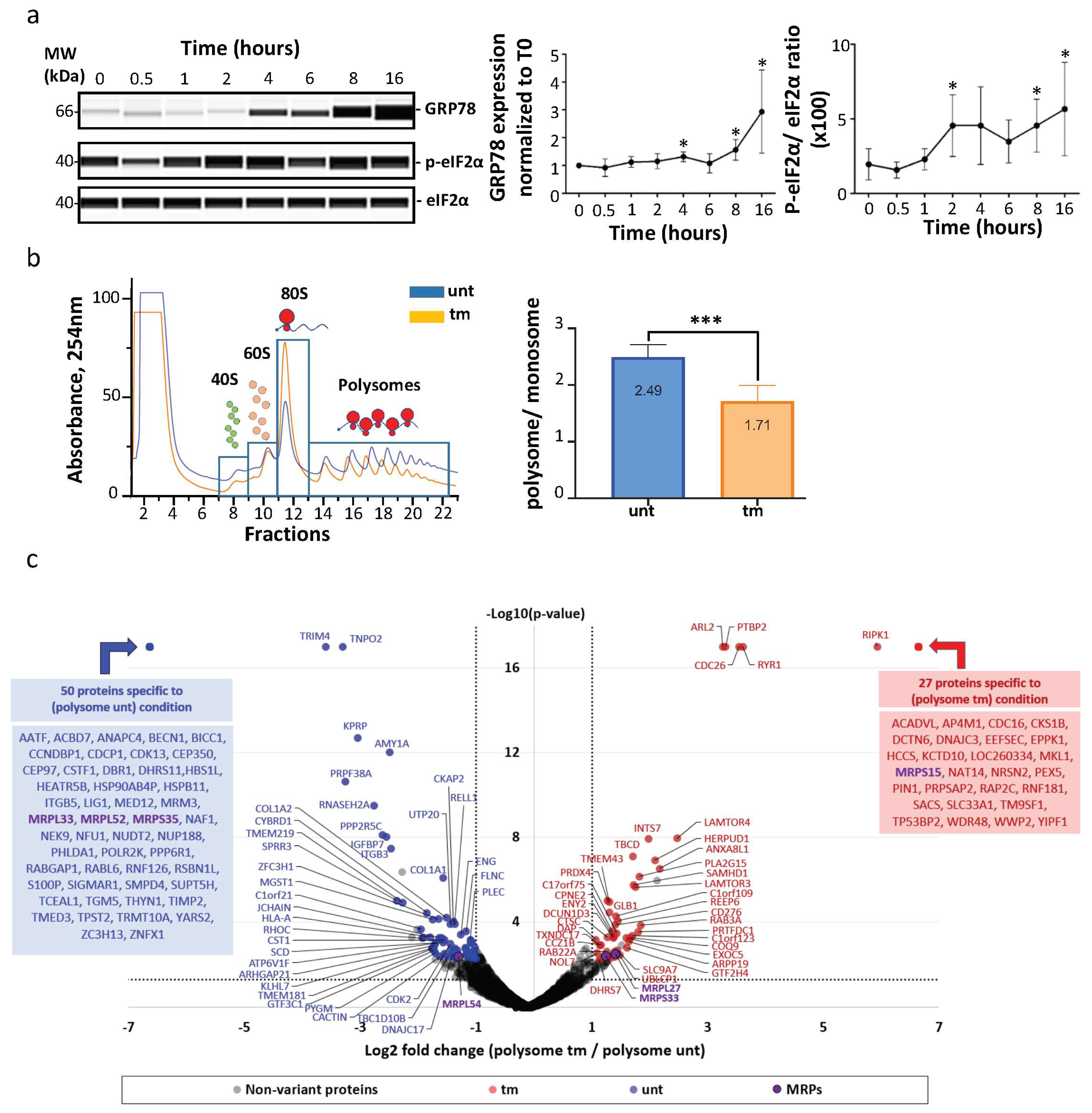
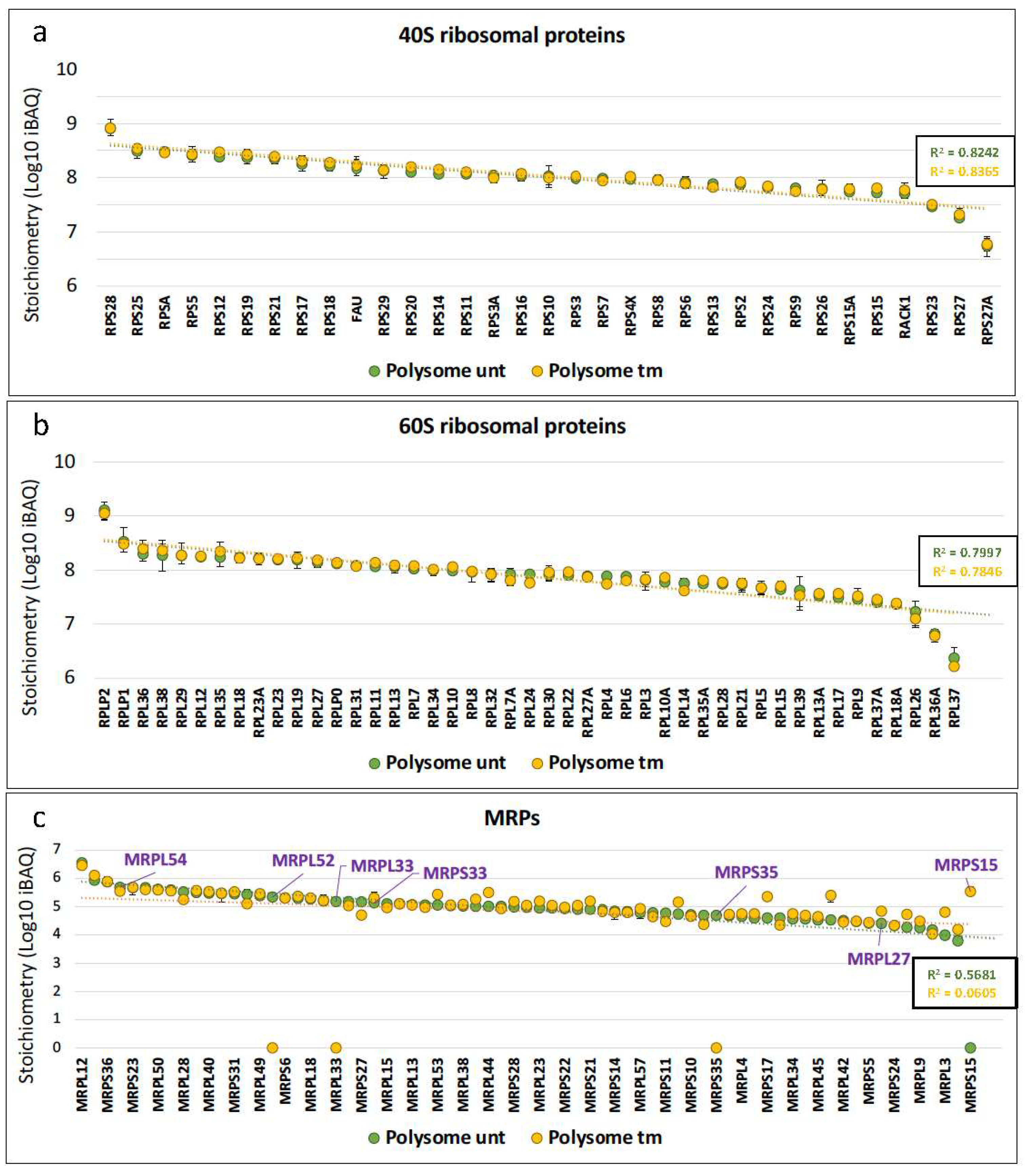
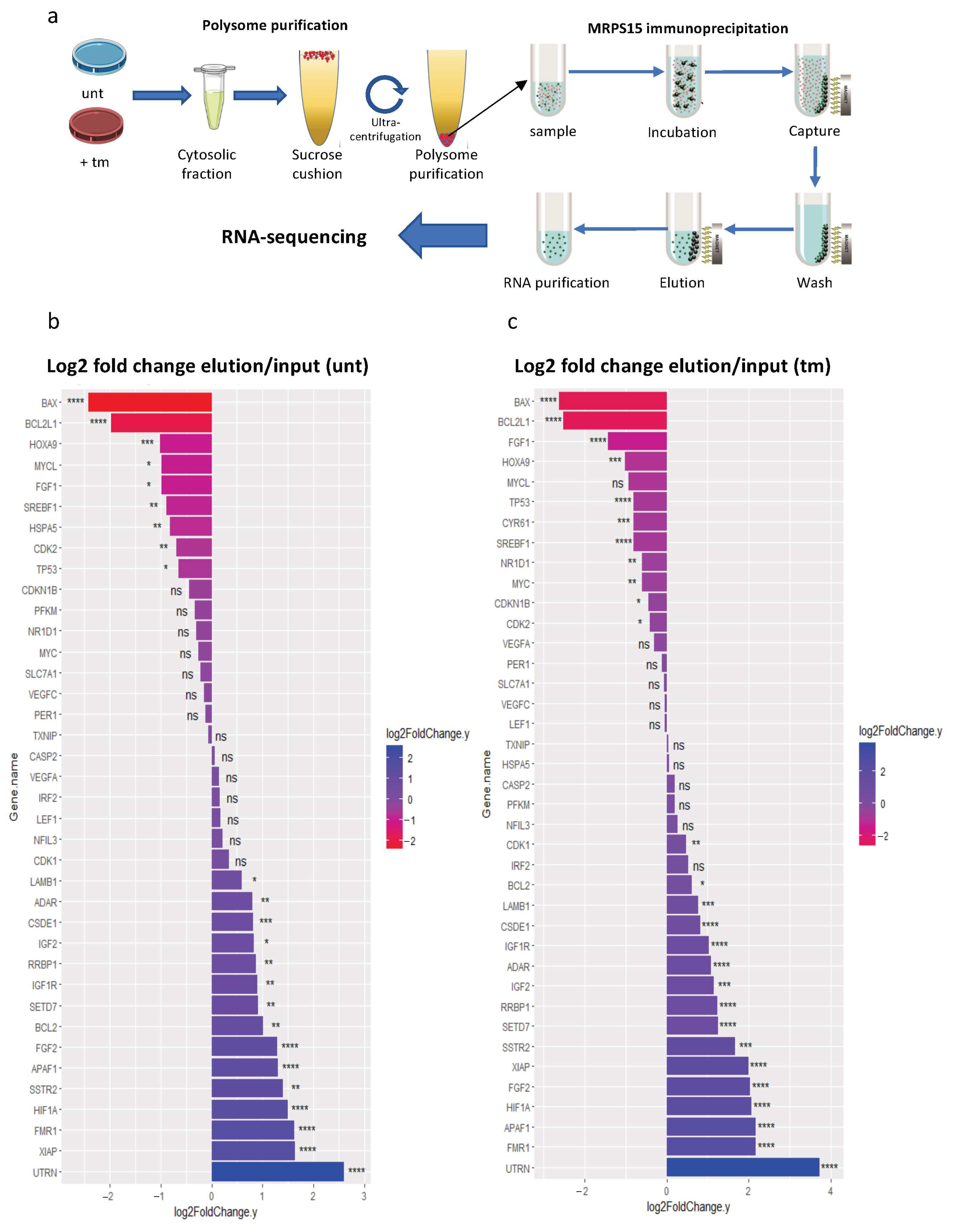
2.1. Mitochondrial Ribosomal Proteins Are Differently Associated with Polysomes in Cardiomyocytes in Response to Stress
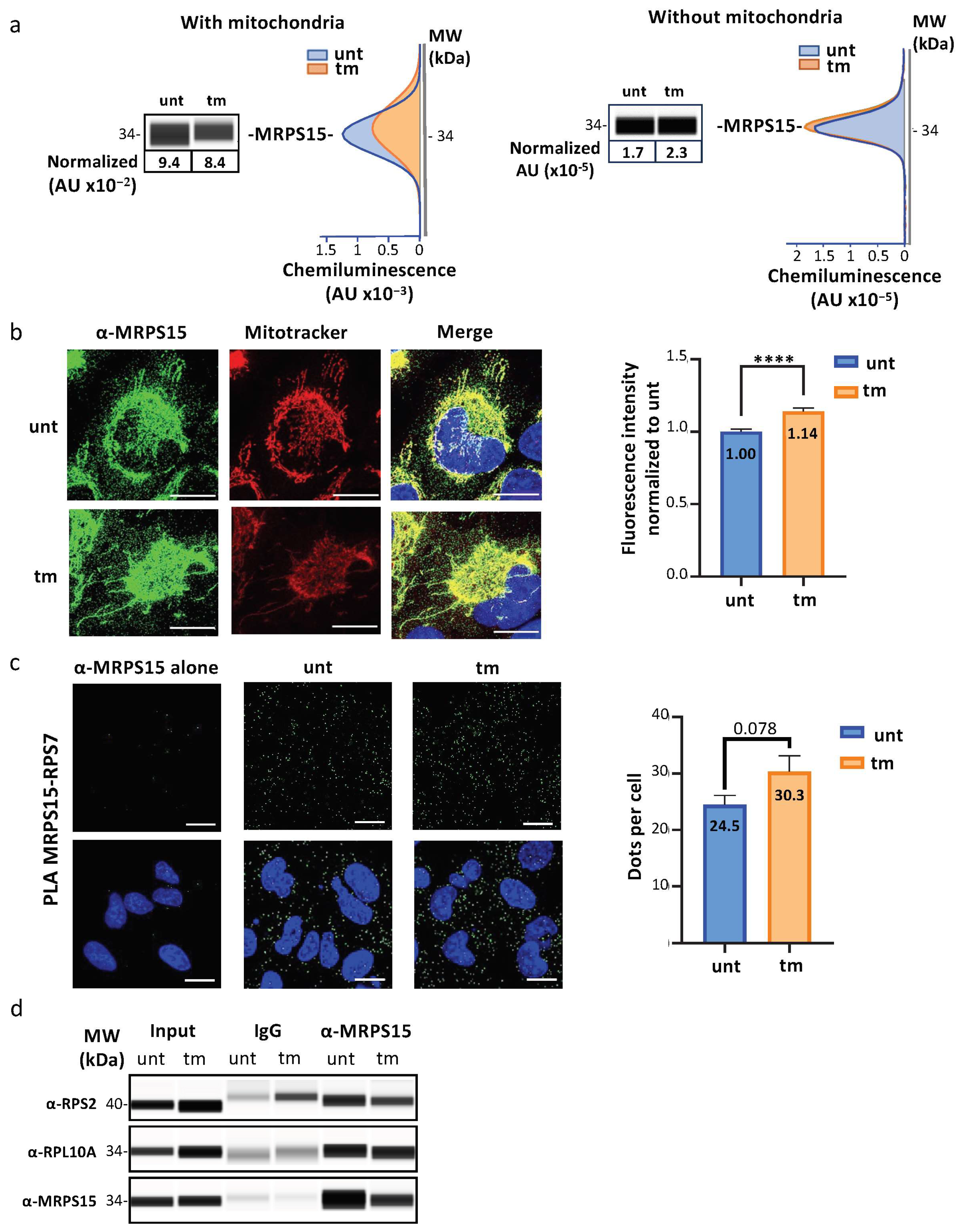
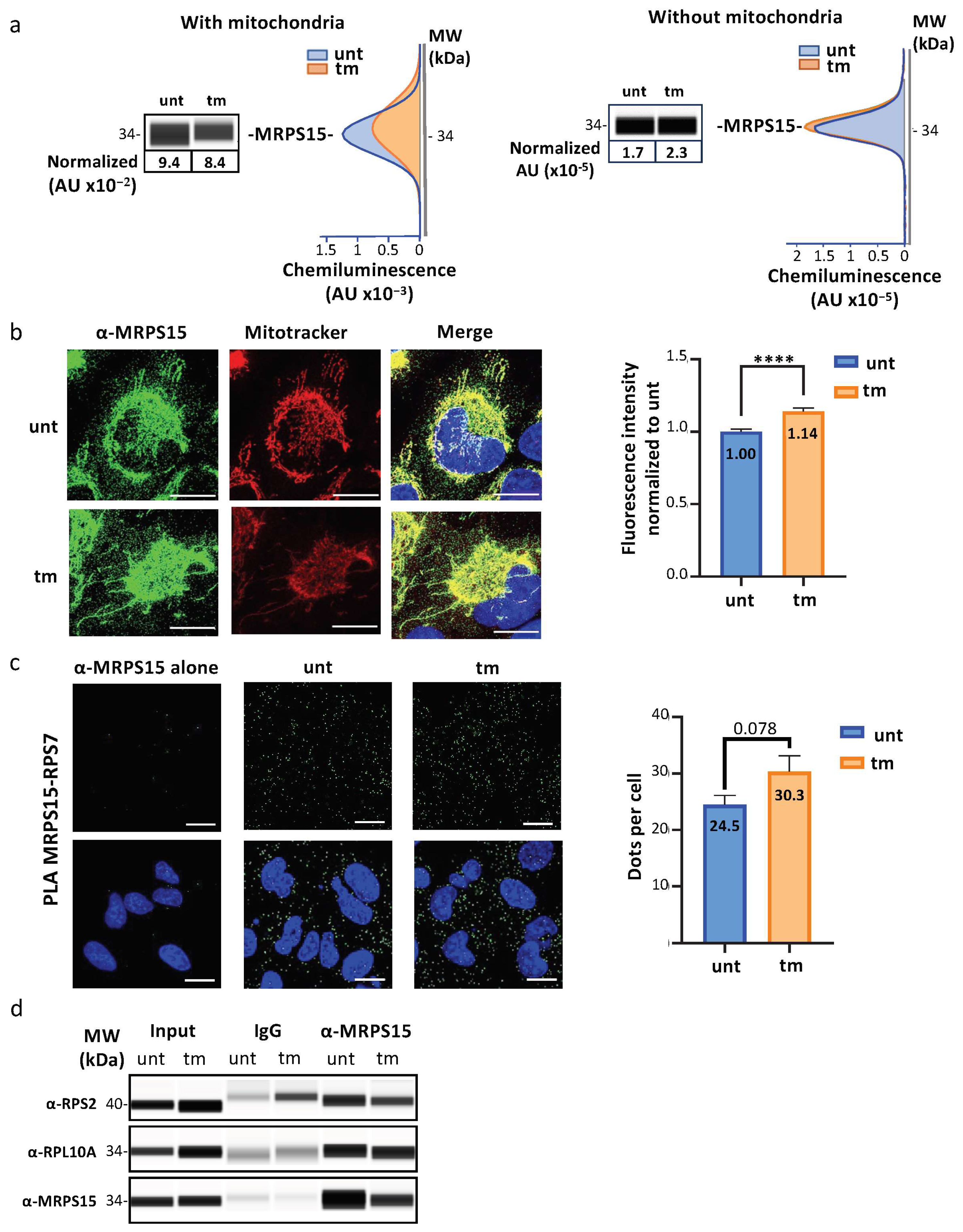
2.2. MRPS15 Is Partly Located in the Cytosol and Interacts with Ribosomal Proteins
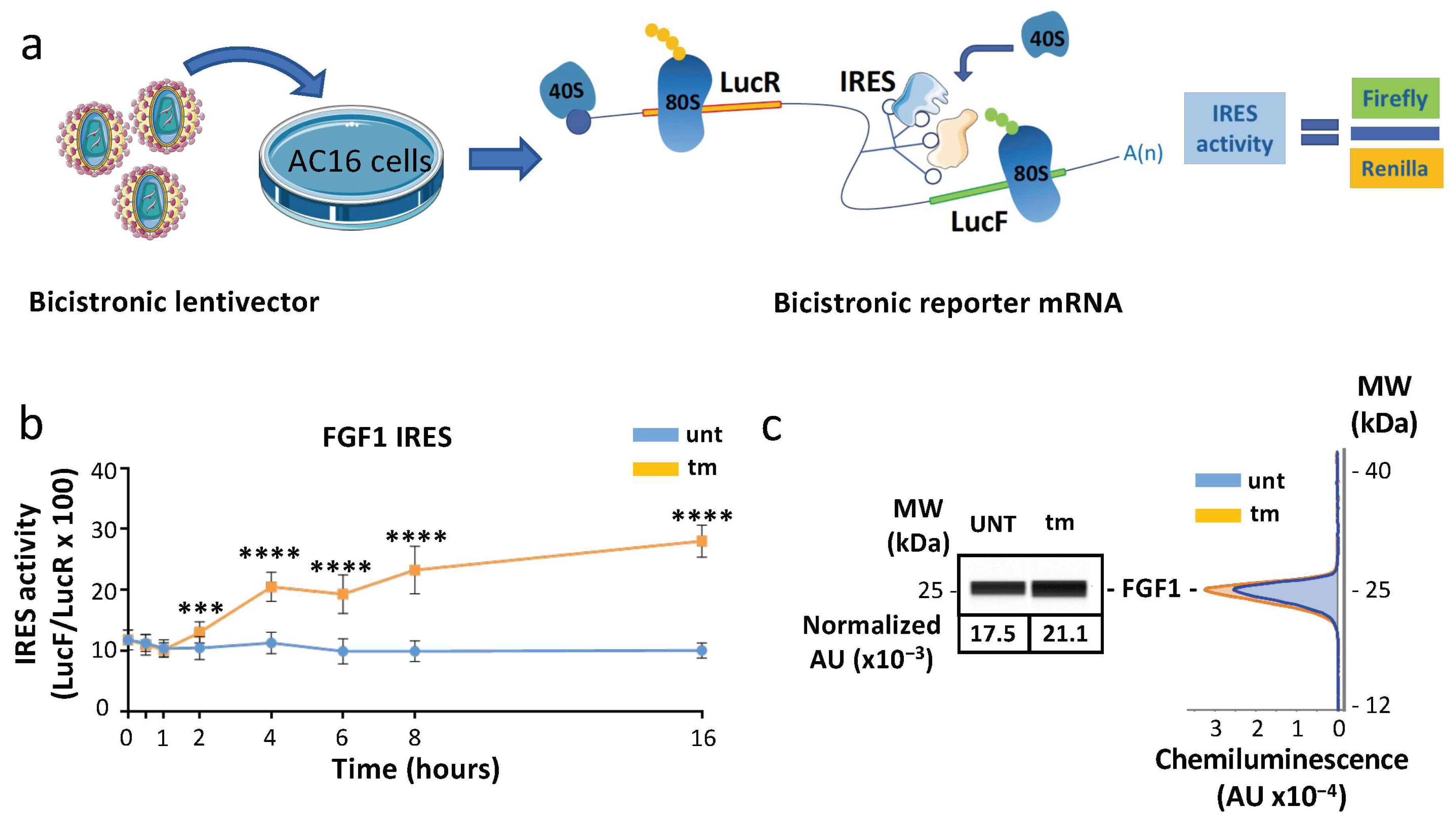
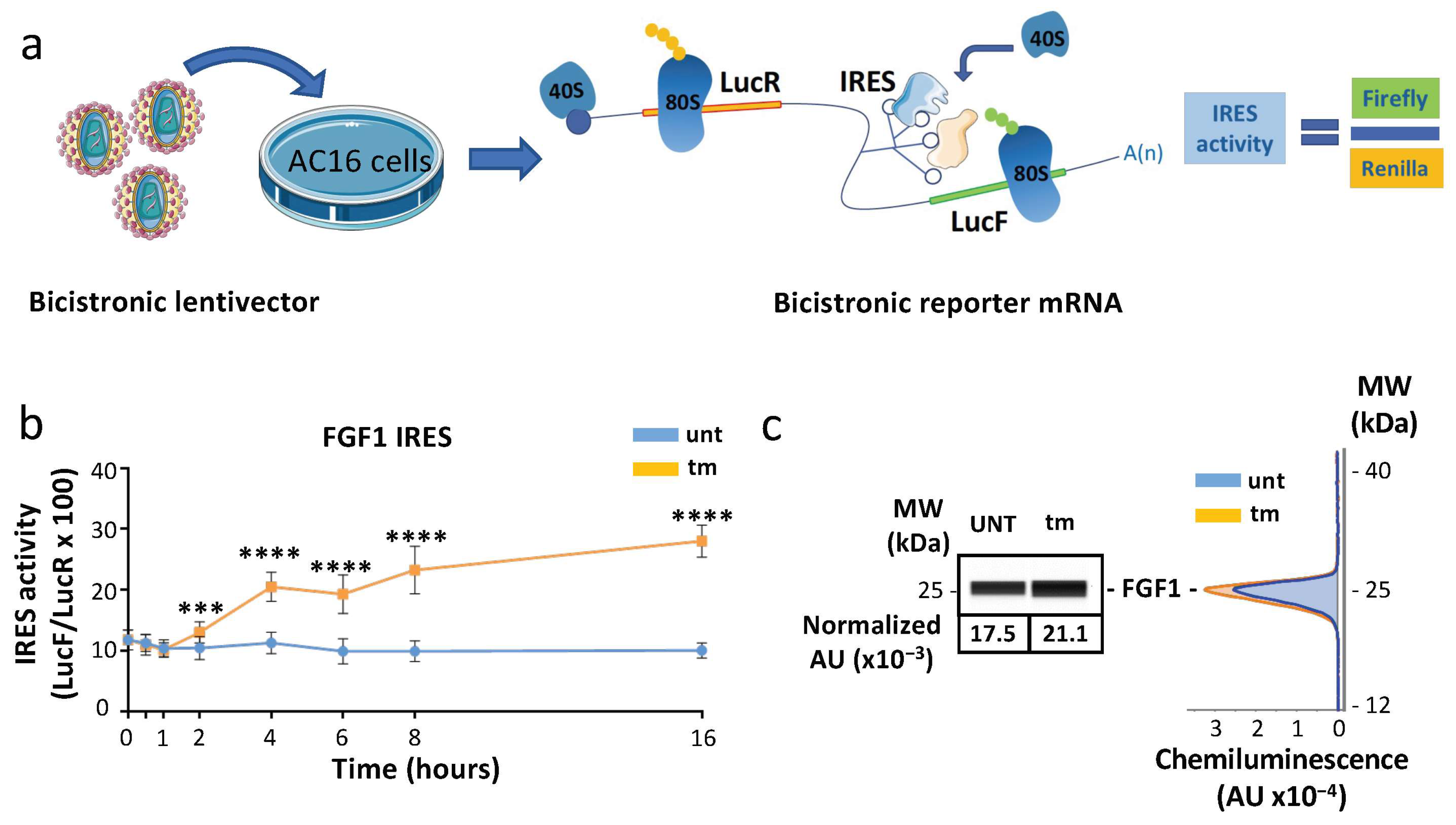
2.3. IRES-Dependent Translation Is Activated by ER Stress in Cardiomyocytes
2.4. MRPS15 Knock-Down Inhibits IRES Activation during ER Stress
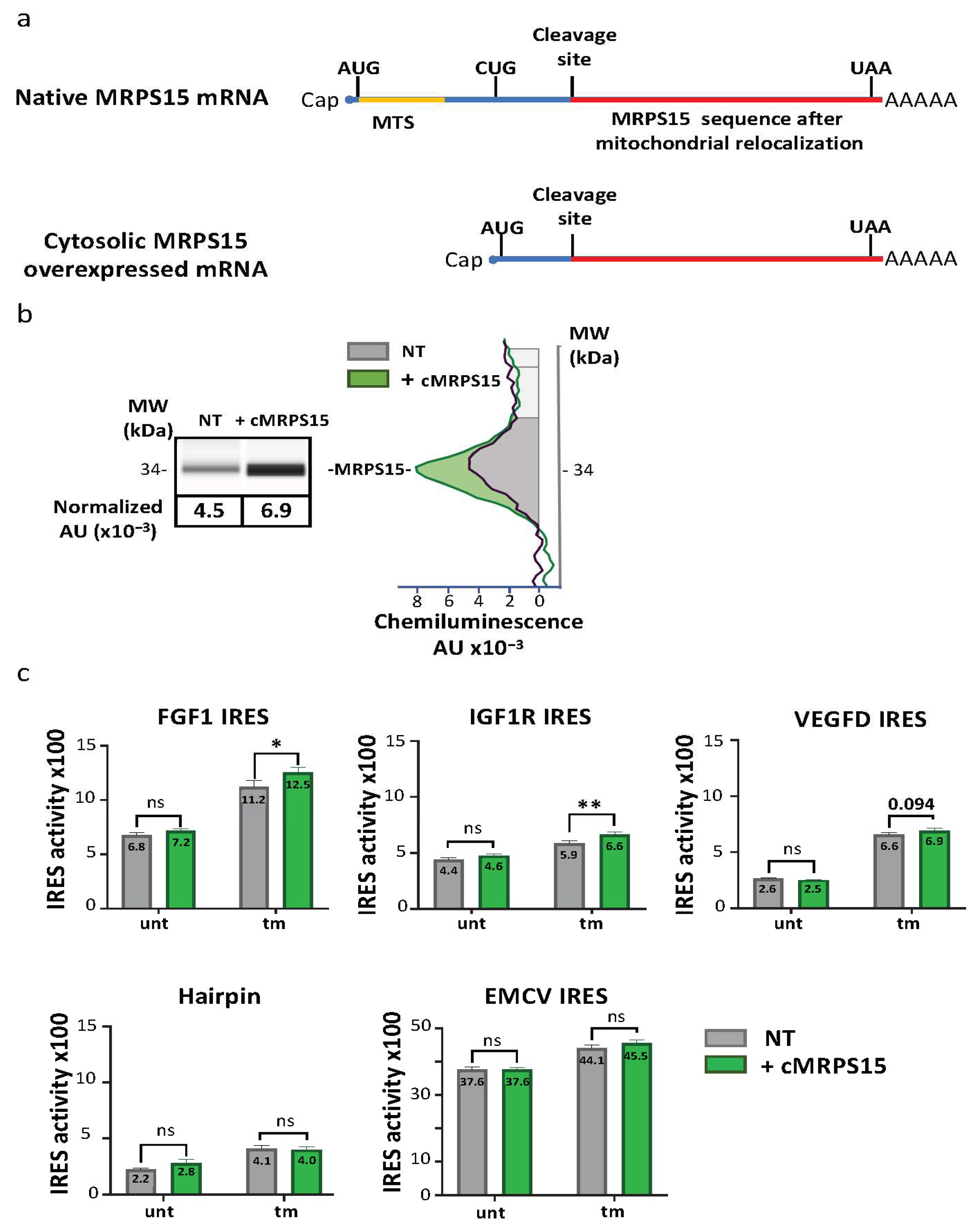
2.5. Cytosolic MRPS15 Overexpression Promotes IRES Activation during ER Stress
2.6. Ribosomes Containing MRPS15 Are More Associated with IRES-Containing mRNAs
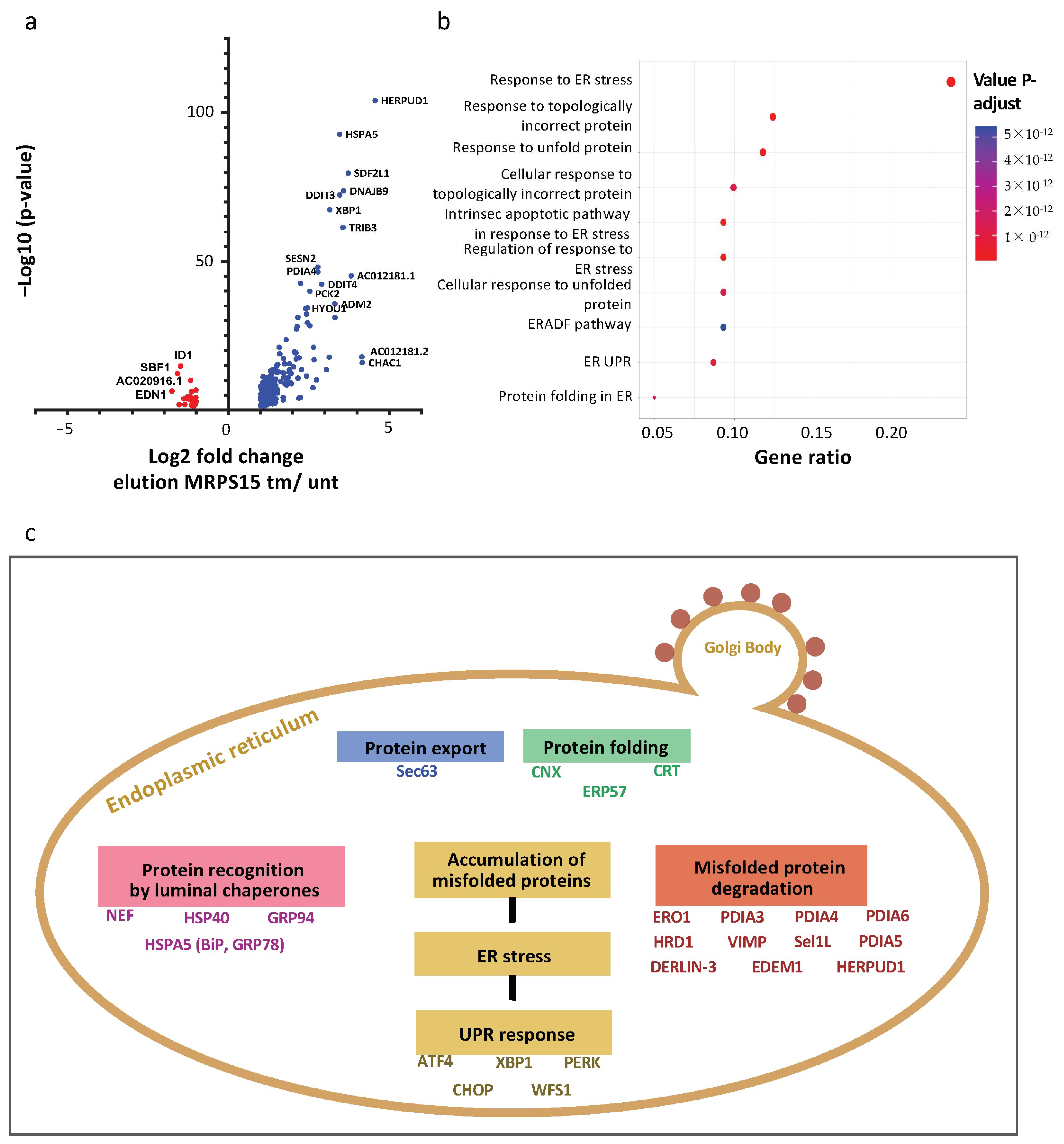
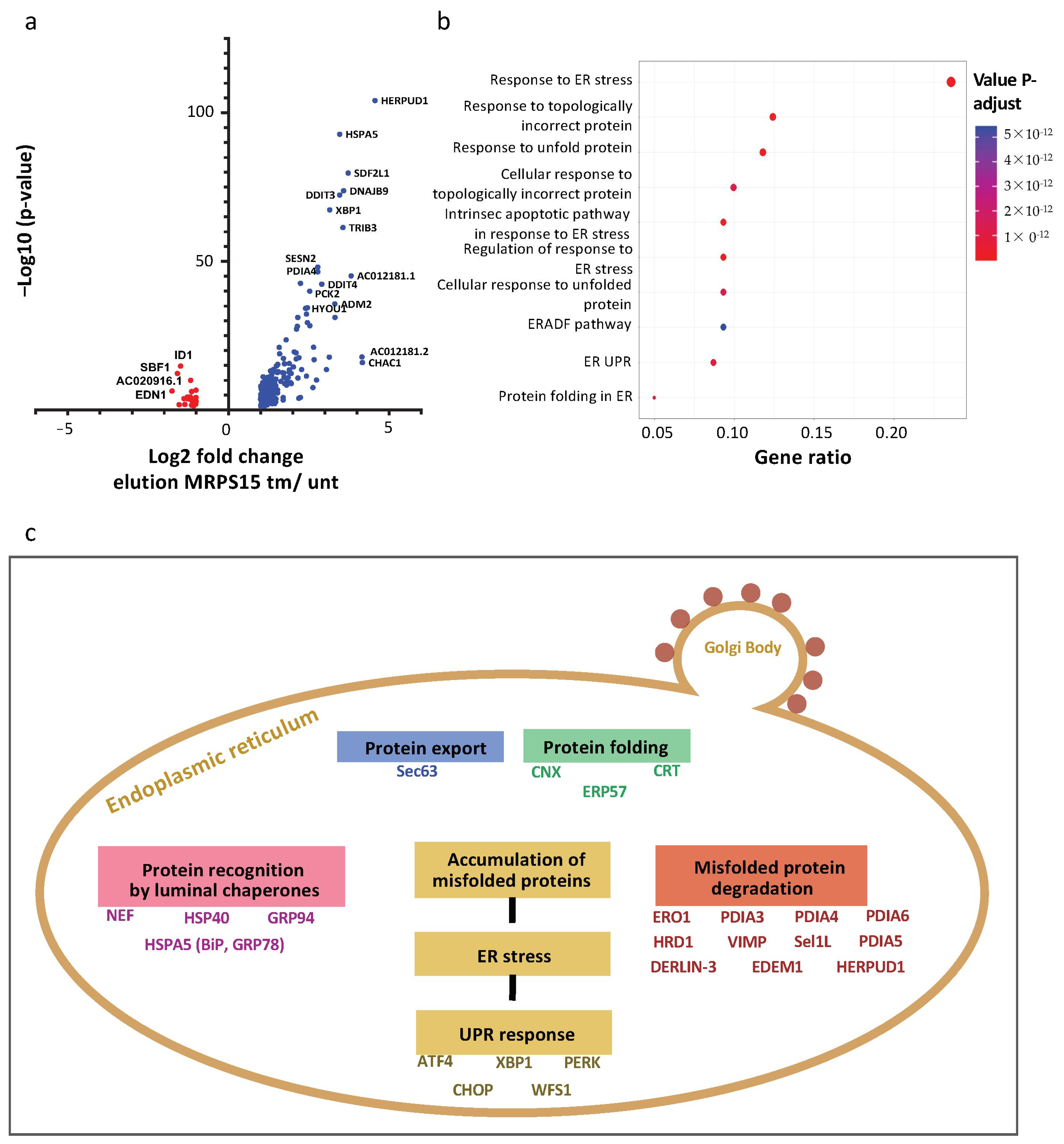
2.7. Ribosomes Containing MRPS15 Are Specialized in Translation of UPR mRNAs
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Bacterial Strains
4.3. Cell Transduction
4.4. Cell Transfection
4.5. Lentivector Construction
4.6. Lentivector Production
4.7. Reporter Activity Assay
4.8. Capillary Western
4.9. RNA Purification and cDNA Synthesis
4.10. QPCR
4.11. Polysomal RNA Preparation from Sucrose Gradients
4.12. Protein Purification from Sucrose Gradient
4.13. Polysome Purification from Sucrose Cushions
4.14. Preparation of Cell Extracts with or without Mitochondria
4.15. In-Gel Trypsin Digestion and Mass Spectrometry Analysis
4.16. MS-Based Protein Identification and Label-Free Quantification
4.17. Immunoprecipitation
4.18. Immunofluorescence
4.19. Proximal Ligation Assay
4.20. Statistical Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resource | Source | Identifier |
|---|---|---|
| Antibodies | ||
| Mouse anti-eIF2α | Cell Signaling | #2103 |
| Rabbit anti-Phospho-eIF2α (Ser51) | Cell Signaling | #9721 |
| Rabbit anti-GRP78 | Novus Biological | NB-56411 |
| Rabbit anti-MRPS15 | Abcam | Ab137070 |
| Rabbit anti-FGF1 | Abcam | Ab207321 |
| Anti-IGF1R | Abcam | Ab182408 |
| Rabbit anti-RPS2 | Bethyl | A303-794A |
| Rabbit anti-RPS25 | Abcam | Ab254671 |
| Rabbit anti-RPL10A | Abcam | Ab240179 |
| Mouse anti-RPS7 | Santa Cruz | Sc-377317 |
| Mouse anti-RPS13 | Santa Cruz | Sc-398690 |
| Mouse anti-HSP60 | Santa Cruz | Sc-13115 |
| Rabbit IgG serum | Sigma | I5006-1MG |
| Rabbit detection module | Protein Simple | DM-001 |
| Mouse detection module | Protein Simple | DM-002 |
| Secondary streptavidine-HRP | Protein Simple | 043-459-2 |
| Secondary donkey anti-Rabbit Alexa 488 | Jackson | 711-545-152 |
| Secondary donkey anti-mouse Alexa 594 | Jackson | 715-585-150 |
| Secondary donkey anti-mouse Alexa 647 | Interchim | FP-SC4110 |
| Bacteria and Virus Strains | ||
| Escherichia Coli Top10 | InVitrogen | C404003 |
| Escherichia Coli Strataclone | Agilent technologies | 200185 |
| Cell lines | ||
| AC-16 cardiomyocytes | ATCC (Manassas, VA, USA) | CRL-3568TM |
| HEK293 FT | Invitrogen | R700-07 |
| Chemicals, Peptides and Recombinant Proteins | ||
| TRI-Reagent | MRC Inc (Houston, TX, USA) | TR118 |
| Isopropanol | Sigma-Aldrich | 33539 |
| Ethanol | Sigma-Aldrich | 32221 |
| Cycloheximide | Merck (Lowe, NJ, USA) | 239764-100MG |
| NP40 (IGEPAL 630) | Sigma-Aldrich | I8896 |
| Magnesium chloride | Serva (Heidelberg, Germany) | 39772 |
| Potassium chloride | Prolabo (Fontenay-sous-Bois, France) | 26764.298 |
| Sodium chloride | Honeywell (Charlott, NC, USA) | 31434 |
| Sucrose | Sigma | S7903 |
| Proteinase inhibitor cocktail | Sigma-Aldrich | P2714 |
| RNAse inhibitor | AppliedBiosystem | N8080119 |
| Acetic acid 100% | Prolabo | 20 104.298 |
| Vanadyl ribonucleoside complexe | Sigma | R3380 |
| PhoSTOP | Roche (Basel, Switzerland) | 04 906 837 001 |
| Bovine serum albumin standard | Euromedex (Souffelweyersheim, France) | 04-100-812-E |
| Sodium dodecyl sulfate 20 % | Biosolve (Dieuze, France) | 0019812323BS |
| RIPA | BioBasic (New York, NY, USA) | RB4476 |
| MitoTracker®Red CMXRos | Cell Signaling | #9082 |
| Critical Commercial Assays | ||
| High capacity cDNA Reverse transcription kit | Thermofisher | 4368814 |
| ONEGreen FAST qPCR Premix | Ozyme | OZYA008 |
| EZ-10 Spin Column Plasmid DNA Miniprep Kit | BioBasic | BS413 |
| 13.2 mL, Open-Top Thinwall Ultra-Clear Tube, 14 × 89 mm | Beckman | 344059 |
| StrataClone Blunt PCR Cloning Kit | Agilent | 240207 |
| Duolink In situ Red Starter kit Mouse/Rabbit | Sigma | DUO92101 |
| Dual-Luciferase® Reporter Assay system | Promega | E1980 |
| Jess or Wes Separation Module | ProteinSimple | SM-SW004 |
| Fluorescent 5x Master Mix 1 | ProteinSimple | PS-FL01-8 |
| Experimental Models: Cell Lines and Medium | ||
| 293FT | Invitrogen | R700-07 |
| AC16 Human cardiomyocyte cell line | Sigma | SCC109 |
| Dulbecco′s Modified Eagle′s Medium/Nutrient Mixture F-12 Ham | Sigma | D6434 |
| Fetal bovine serum | Sigma | ES-009-B |
| L-Glutamine | Sigma | TMS-002-C |
| Penicillin-streptomycin | Sigma | TMS-AB2-C |
| Opti-MEM, reduced serum, no-phenol red | Gibco | 11058021 |
| Oligonucleotides | ||
| HPRT | F: 5′-TGCTTTCCTTGGTCAGGCAGT-3′ | R: 5′-CTTCGTGGGGTCCTTTTCACC-3′ |
| FLuc | F: 5′-GTGTTGTTCCATTCCATCA-3′ | R: 5′-TTGGCGAAGAAGGAGAATA-3′ |
| Rluc | F: 5′-GCCTGATATTGAAGAAGATATTG-3′ | R: 5′-CCTTTCTCTTTGAATGGTTC-3′ |
| FGF1 CDS | F: 5′-GCT GAA GGG GAA ATC ACC AC-3′ | R: 5′-CCC GTT GCT ACA GTA GAG GAG-3′ |
| FGF1A | F: 5′-CCT CCT TTT CTG GGA GGA CA-3′ | R: 5′-C AGC TTC TGC AAT GTC CAC-3′ |
| FGF2 | F: 5′-TGGTATGTGGCACTGAAACGA- 3′ | R: 5′-GCCCAGGTCCTGTTTTGGAT-3′ |
| VEGFA | F: 5′-TGCTGTCTTGGGTGCATTGGA-3′ | R: 5′-CCACTTCGTGATGATTCTGCC-3′ |
| VEGFC | F: 5′-AAAGAAGTTCCACCACCAAAC-3′ | R: 5′-AGGGACACAACGACACACTTC-3′ |
| 18S | F: 5′-CAACTAAGAACGGCCATGCA-3′ | R: 5′-AGCCTGCGGCTTAATTTGAC-3′ |
| MRPS15 primer 1 | F: 5′-CAAGATCCGCAGTTATGAAGAACAC-3′ | R: 5′-TCCTCTGGTCAATGCTCATTAGC-3′ |
| MRPS15 primer 2 | F: 5′-CGTGACCAAGAAGGCTCTGTG-3′ | R: 5′-GCTGCAGCCTTTAAGGCTCT-3′ |
| Recombinant DNA | ||
| pTRIP-CRHL+ | Sequence available on Dryad, (2) | https://doi.org/10.5061/dryad.nvx0k6dq7, accessed on 20 February 2024 |
| pTRIP-CRF1AL+ | Sequence available on Dryad, (17; 26) | https://doi.org/10.5061/dryad.nvx0k6dq7, accessed on 20 February 2024 |
| pTRIP-CRIGL+ | ||
| pTRIP-CRhVDL+ | ||
| pTRIP-CREL+ | Sequence available on Dryad, (13) | https://doi.org/10.5061/dryad.nvx0k6dq7, accessed on 20 February 2024 |
| pCMV-dR8.91 | Addgene (Watertown, MA, USA) | |
| pCMV-VSV-G | Addgene | |
| pTRIP MRPS15 | ||
| ON-TARGETplus Human MRPS15 (64960) siRNA–SMARTpool | Dharmacon (Cambridge, UK) | L-013609-02-0020 |
| ON-TARGETplus Non-targeting Pool | Dharmacon | D-001810-10-20 |
| Software and Algorithms | ||
| Prism 7 | Graphpad (Boston, MA, USA) | https://www.graphpad.com/scientific-software/prism/, accessed on 20 February 2024 |
| Microsoft 365 (excel, word, powerpoint) version 16.82 | Microsoft office (Redmond, WA, USA) | |
| FIJI 1.530 | FIJI (Madison, WI, USA) | https://fiji.sc/, accessed on 20 February 2024 |
| ImageJ version 1.53 | ImageJ (NIH, Stapleton, NY, USA) | https://imagej.nih.gov/ij/download.html, accessed on 20 February 2024 |
| Zen black/Blue edition version 2.3 SP1 FP3 | Zeiss (Rueil Malmaison, France) | https://www.zeiss.fr/microscopie/produits/microscope-software/zen-lite.html, accessed on 20 February 2024 |
| QuantStudio | Applied Biosystems | https://www.thermofisher.com/fr/fr/home/global/forms/life-science/quantstudio-3-5-software.html, accessed on 20 February 2024 |
| Microwin 2000 | Berthold | https://fr.freedownloadmanager.org/Windows-PC/MikroWin-2000.html, accessed on 20 February 2024 |
| Other | ||
| LSM780 Zeiss confocal microscope | Zeiss | - |
| Jess capillary western | Protein Simple | - |
| SW 41 Ti Swinging-Bucket Rotor | Beckman | 331362 |
| Optima XL-100K Ultracentrifugeuse | Beckman | - |
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Röst, H.L.; Teruel, M.N.; Barna, M. Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-Wide. Mol. Cell 2017, 67, 71–83.e7. [Google Scholar] [CrossRef]
- Genuth, N.R.; Barna, M. The Discovery of Ribosome Heterogeneity and Its Implications for Gene Regulation and Organismal Life. Mol. Cell 2018, 71, 364–374. [Google Scholar] [CrossRef]
- Kochavi, A.; Lovecchio, D.; Faller, W.J.; Agami, R. Proteome Diversification by mRNA Translation in Cancer. Mol. Cell 2023, 83, 469–480. [Google Scholar] [CrossRef]
- Cinque, S.; Verheyden, Y.; Katopodi, V.; Knezevic, Z.; Demesmaeker, E.; Adnane, S.; Hanache, S.; Vendramin, R.; Stinkens, F.; Vervloesem, F.; et al. The Generation of Cancer-Specific Ribosomes by the lncRNA LISR Suppresses Melanoma Anti-Tumour Immunity; SSRN: London, UK, 2023. [Google Scholar]
- Boussaid, I.; Fontenay, M. Translation Defects in Ribosomopathies. Curr. Opin. Hematol. 2022, 29, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Natchiar, S.K.; Myasnikov, A.G.; Kratzat, H.; Hazemann, I.; Klaholz, B.P. Visualization of Chemical Modifications in the Human 80S Ribosome Structure. Nature 2017, 551, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Poole, A.R.; Vicino, I.; Adachi, H.; Yu, Y.-T.; Hebert, M.D. Regulatory RNPs: A Novel Class of Ribonucleoproteins That Potentially Contribute to Ribosome Heterogeneity. Biol. Open 2017, 6, 1342–1354. [Google Scholar] [CrossRef] [PubMed]
- Marcel, V.; Kielbassa, J.; Marchand, V.; Natchiar, K.S.; Paraqindes, H.; Nguyen Van Long, F.; Ayadi, L.; Bourguignon-Igel, V.; Lo Monaco, P.; Monchiet, D.; et al. Ribosomal RNA 2′O-Methylation as a Novel Layer of Inter-Tumour Heterogeneity in Breast Cancer. NAR Cancer 2020, 2, zcaa036. [Google Scholar] [CrossRef]
- Marcel, V.; Ghayad, S.E.; Belin, S.; Therizols, G.; Morel, A.-P.; Solano-Gonzàlez, E.; Vendrell, J.A.; Hacot, S.; Mertani, H.C.; Albaret, M.A.; et al. P53 Acts as a Safeguard of Translational Control by Regulating Fibrillarin and rRNA Methylation in Cancer. Cancer Cell 2013, 24, 318–330. [Google Scholar] [CrossRef]
- Jansson, M.D.; Häfner, S.J.; Altinel, K.; Tehler, D.; Krogh, N.; Jakobsen, E.; Andersen, J.V.; Andersen, K.L.; Schoof, E.M.; Ménard, P.; et al. Regulation of Translation by Site-Specific Ribosomal RNA Methylation. Nat. Struct. Mol. Biol. 2021, 28, 889–899. [Google Scholar] [CrossRef]
- Penzo, M.; Rocchi, L.; Brugiere, S.; Carnicelli, D.; Onofrillo, C.; Couté, Y.; Brigotti, M.; Montanaro, L. Human Ribosomes from Cells with Reduced Dyskerin Levels Are Intrinsically Altered in Translation. FASEB J. 2015, 29, 3472–3482. [Google Scholar] [CrossRef]
- Yoon, A.; Peng, G.; Brandenburg, Y.; Zollo, O.; Xu, W.; Rego, E.; Ruggero, D. Impaired Control of IRES-Mediated Translation in X-Linked Dyskeratosis Congenita. Science 2006, 312, 902–906. [Google Scholar] [CrossRef]
- Singleton, R.S.; Liu-Yi, P.; Formenti, F.; Ge, W.; Sekirnik, R.; Fischer, R.; Adam, J.; Pollard, P.J.; Wolf, A.; Thalhammer, A.; et al. OGFOD1 Catalyzes Prolyl Hydroxylation of RPS23 and Is Involved in Translation Control and Stress Granule Formation. Proc. Natl. Acad. Sci. USA 2014, 111, 4031–4036. [Google Scholar] [CrossRef] [PubMed]
- Landry, D.M.; Hertz, M.I.; Thompson, S.R. RPS25 Is Essential for Translation Initiation by the Dicistroviridae and Hepatitis C Viral IRESs. Genes Dev. 2009, 23, 2753–2764. [Google Scholar] [CrossRef]
- Godet, A.-C.; David, F.; Hantelys, F.; Tatin, F.; Lacazette, E.; Garmy-Susini, B.; Prats, A.-C. IRES Trans-Acting Factors, Key Actors of the Stress Response. Int. J. Mol. Sci. 2019, 20, 924. [Google Scholar] [CrossRef] [PubMed]
- Holcik, M.; Sonenberg, N. Translational Control in Stress and Apoptosis. Nat. Rev. Mol. Cell Biol. 2005, 6, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Walters, B.; Axhemi, A.; Jankowsky, E.; Thompson, S.R. Binding of a Viral IRES to the 40S Subunit Occurs in Two Successive Steps Mediated by eS25. Nucleic Acids Res. 2020, 48, 8063–8073. [Google Scholar] [CrossRef]
- Renaud-Gabardos, E.; Tatin, F.; Hantelys, F.; Lebas, B.; Calise, D.; Kunduzova, O.; Masri, B.; Pujol, F.; Sicard, P.; Valet, P.; et al. Therapeutic Benefit and Gene Network Regulation by Combined Gene Transfer of Apelin, FGF2, and SERCA2a into Ischemic Heart. Mol. Ther. 2018, 26, 902–916. [Google Scholar] [CrossRef]
- Giraud, S.; Greco, A.; Brink, M.; Diaz, J.-J.; Delafontaine, P. Translation Initiation of the Insulin-like Growth Factor I Receptor mRNA Is Mediated by an Internal Ribosome Entry Site. J. Biol. Chem. 2001, 276, 5668–5675. [Google Scholar] [CrossRef]
- Knezevic, I.; Patel, A.; Sundaresan, N.R.; Gupta, M.P.; Solaro, R.J.; Nagalingam, R.S.; Gupta, M. A Novel Cardiomyocyte-Enriched MicroRNA, miR-378, Targets Insulin-like Growth Factor 1 Receptor. J. Biol. Chem. 2012, 287, 12913–12926. [Google Scholar] [CrossRef]
- Bogenhagen, D.F.; Ostermeyer-Fay, A.G.; Haley, J.D.; Garcia-Diaz, M. Kinetics and Mechanism of Mammalian Mitochondrial Ribosome Assembly. Cell Rep. 2018, 22, 1935–1944. [Google Scholar] [CrossRef]
- Greber, B.J.; Ban, N. Structure and Function of the Mitochondrial Ribosome. Annu. Rev. Biochem. 2016, 85, 103–132. [Google Scholar] [CrossRef]
- Macejak, D.G.; Sarnow, P. Internal Initiation of Translation Mediated by the 5′ Leader of a Cellular mRNA. Nature 1991, 353, 90–94. [Google Scholar] [CrossRef]
- Drogat, B.; Auguste, P.; Nguyen, D.T.; Bouchecareilh, M.; Pineau, R.; Nalbantoglu, J.; Kaufman, R.J.; Chevet, E.; Bikfalvi, A.; Moenner, M. IRE1 Signaling Is Essential for Ischemia-Induced Vascular Endothelial Growth Factor-A Expression and Contributes to Angiogenesis and Tumor Growth In Vivo. Cancer Res. 2007, 67, 6700–6707. [Google Scholar] [CrossRef]
- Guo, W.; Wang, M.; Yang, Z.; Liu, D.; Ma, B.; Zhao, Y.; Chen, Y.; Hu, Y. Recent Advances in Small Molecule and Peptide Inhibitors of Glucose-Regulated Protein 78 for Cancer Therapy. Eur. J. Med. Chem. 2023, 261, 115792. [Google Scholar] [CrossRef]
- Xu, C. Endoplasmic Reticulum Stress: Cell Life and Death Decisions. J. Clin. Investig. 2005, 115, 2656–2664. [Google Scholar] [CrossRef]
- Zhu, H.; Zhou, H. Novel Insight into the Role of Endoplasmic Reticulum Stress in the Pathogenesis of Myocardial Ischemia-Reperfusion Injury. Oxidative Med. Cell. Longev. 2021, 2021, 5529810. [Google Scholar] [CrossRef]
- Mainali, N.; Li, X.; Wang, X.; Balasubramaniam, M.; Ganne, A.; Kore, R.; Shmookler Reis, R.J.; Mehta, J.L.; Ayyadevara, S. Myocardial Infarction Elevates Endoplasmic Reticulum Stress and Protein Aggregation in Heart as Well as Brain. Mol. Cell Biochem. 2023, 1–13. [Google Scholar] [CrossRef]
- Philippe, C.; Dubrac, A.; Quelen, C.; Desquesnes, A.; Van Den Berghe, L.; Ségura, C.; Filleron, T.; Pyronnet, S.; Prats, H.; Brousset, P.; et al. PERK Mediates the IRES-Dependent Translational Activation of mRNAs Encoding Angiogenic Growth Factors after Ischemic Stress. Sci. Signal. 2016, 9, ra44. [Google Scholar] [CrossRef]
- Fernandez, J.; Bode, B.; Koromilas, A.; Diehl, J.A.; Krukovets, I.; Snider, M.D.; Hatzoglou, M. Translation Mediated by the Internal Ribosome Entry Site of the Cat-1 mRNA Is Regulated by Glucose Availability in a PERK Kinase-Dependent Manner. J. Biol. Chem. 2002, 277, 11780–11787. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.; Nesti, C.; Palenzuela, L.; Walker, W.; Hernandez, E.; Protas, L.; Hirano, M.; Isaac, N. Novel Cell Lines Derived from Adult Human Ventricular Cardiomyocytes. J. Mol. Cell. Cardiol. 2005, 39, 133–147. [Google Scholar] [CrossRef]
- Hantelys, F.; Godet, A.-C.; David, F.; Tatin, F.; Renaud-Gabardos, E.; Pujol, F.; Diallo, L.H.; Ader, I.; Ligat, L.; Henras, A.K.; et al. Vasohibin1, a New Mouse Cardiomyocyte IRES Trans-Acting Factor That Regulates Translation in Early Hypoxia. eLife 2019, 8, e50094. [Google Scholar] [CrossRef]
- Matsuki, Y.; Matsuo, Y.; Nakano, Y.; Iwasaki, S.; Yoko, H.; Udagawa, T.; Li, S.; Saeki, Y.; Yoshihisa, T.; Tanaka, K.; et al. Ribosomal Protein S7 Ubiquitination during ER Stress in Yeast Is Associated with Selective mRNA Translation and Stress Outcome. Sci. Rep. 2020, 10, 19669. [Google Scholar] [CrossRef]
- Ikeuchi, K.; Ivic, N.; Buschauer, R.; Cheng, J.; Fröhlich, T.; Matsuo, Y.; Berninghausen, O.; Inada, T.; Becker, T.; Beckmann, R. Molecular Basis for Recognition and Deubiquitination of 40S Ribosomes by Otu2. Nat. Commun. 2023, 14, 2730. [Google Scholar] [CrossRef]
- Godet, A.-C.; Roussel, E.; David, F.; Hantelys, F.; Morfoisse, F.; Alves, J.; Pujol, F.; Ader, I.; Bertrand, E.; Burlet-Schiltz, O.; et al. Long Non-Coding RNA Neat1 and Paraspeckle Components Are Translational Regulators in Hypoxia. eLife 2022, 11, e69162. [Google Scholar] [CrossRef]
- Martineau, Y.; Le Bec, C.; Monbrun, L.; Allo, V.; Chiu, I.-M.; Danos, O.; Moine, H.; Prats, H.; Prats, A.-C. Internal Ribosome Entry Site Structural Motifs Conserved among Mammalian Fibroblast Growth Factor 1 Alternatively Spliced mRNAs. Mol. Cell. Biol. 2004, 24, 7622–7635. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, X.; Coots, R.A.; Conn, C.S.; Liu, B.; Qian, S.-B. Translational Control of the Cytosolic Stress Response by Mitochondrial Ribosomal Protein L18. Nat. Struct. Mol. Biol. 2015, 22, 404–410. [Google Scholar] [CrossRef]
- Casad, M.E.; Abraham, D.; Kim, I.-M.; Frangakis, S.; Dong, B.; Lin, N.; Wolf, M.J.; Rockman, H.A. Cardiomyopathy Is Associated with Ribosomal Protein Gene Haplo-Insufficiency in Drosophila Melanogaster. Genetics 2011, 189, 861–870. [Google Scholar] [CrossRef]
- Cavdar Koc, E.; Burkhart, W.; Blackburn, K.; Moseley, A.; Spremulli, L.L. The Small Subunit of the Mammalian Mitochondrial Ribosome. J. Biol. Chem. 2001, 276, 19363–19374. [Google Scholar] [CrossRef]
- Wilson, D.N.; Doudna Cate, J.H. The Structure and Function of the Eukaryotic Ribosome. Cold Spring Harb. Perspect. Biol. 2012, 4, a011536. [Google Scholar] [CrossRef]
- Melnikov, S.; Ben-Shem, A.; Garreau De Loubresse, N.; Jenner, L.; Yusupova, G.; Yusupov, M. One Core, Two Shells: Bacterial and Eukaryotic Ribosomes. Nat. Struct. Mol. Biol. 2012, 19, 560–567. [Google Scholar] [CrossRef]
- Malygin, A.; Parakhnevitch, N.; Karpova, G. Human Ribosomal Protein S13: Cloning, Expression, Refolding, and Structural Stability. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2005, 1747, 93–97. [Google Scholar] [CrossRef]
- Jiao, J.; Kavdia, K.; Pagala, V.; Palmer, L.; Finkelstein, D.; Fan, Y.; Peng, J.; Demontis, F. An Age-Downregulated Ribosomal RpS28 Protein Variant Regulates the Muscle Proteome. G3 Genes|Genomes|Genet. 2021, 11, jkab165. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.C.; Nie, Y.; Ren, S.; De Costa, N.T.T.S.; Pandey, R.K.; Hong, J.; Smith, D.B.; Symons, J.A.; Beigelman, L.; Blatt, L.M. Mechanism of Action of Hepatitis B Virus S Antigen Transport-Inhibiting Oligonucleotide Polymer, STOPS, Molecules. Mol. Ther.-Nucleic Acids 2022, 27, 335–348. [Google Scholar] [CrossRef]
- Dong, H.-J.; Wang, J.; Zhang, X.-Z.; Li, C.-C.; Liu, J.-F.; Wang, X.-J. Proteomic Screening Identifies RPLp2 as a Specific Regulator for the Translation of Coronavirus. Int. J. Biol. Macromol. 2023, 230, 123191. [Google Scholar] [CrossRef]
- Luo, J.; Zhao, H.; Chen, L.; Liu, M. Multifaceted Functions of RPS27a: An Unconventional Ribosomal Protein. J. Cell. Physiol. 2023, 238, 485–497. [Google Scholar] [CrossRef]
- Nosrati, N.; Kapoor, N.R.; Kumar, V. DNA Damage Stress Induces the Expression of Ribosomal Protein S27a Gene in a P53-Dependent Manner. Gene 2015, 559, 44–51. [Google Scholar] [CrossRef]
- Riepe, C.; Zelin, E.; Frankino, P.A.; Meacham, Z.A.; Fernandez, S.G.; Ingolia, N.T.; Corn, J.E. Double Stranded DNA Breaks and Genome Editing Trigger Loss of Ribosomal Protein RPS27A. FEBS J. 2022, 289, 3101–3114. [Google Scholar] [CrossRef]
- Chen, T.-W.; Chang, K.-P.; Cheng, C.-C.; Chen, C.-Y.; Hong, S.-W.; Sie, Z.-L.; Cheng, H.-W.; Yen, W.-C.; Huang, Y.; Liu, S.-C.; et al. Characterization of Recurrent Relevant Genes Reveals a Novel Role of RPL36A in Radioresistant Oral Squamous Cell Carcinoma. Cancers 2021, 13, 5623. [Google Scholar] [CrossRef]
- Chen, Q.; Li, L.; Samidurai, A.; Thompson, J.; Hu, Y.; Willard, B.; Lesnefsky, E.J. Acute Endoplasmic Reticulum Stress-induced Mitochondria Respiratory Chain Damage: The Role of Activated Calpains. FASEB J. 2024, 38, e23404. [Google Scholar] [CrossRef]
- Ravi; Kumar, A.; Bhattacharyya, S.; Singh, J. Thiol Reductive Stress Activates the Hypoxia Response Pathway. EMBO J. 2023, 42, e114093. [Google Scholar] [CrossRef]
- Reinhard, J.; Mattes, C.; Väth, K.; Radanović, T.; Surma, M.A.; Klose, C.; Ernst, R. A Quantitative Analysis of Cellular Lipid Compositions During Acute Proteotoxic ER Stress Reveals Specificity in the Production of Asymmetric Lipids. Front. Cell Dev. Biol. 2020, 8, 756. [Google Scholar] [CrossRef]
- Kumar, R.A.; Thome, T.; Sharaf, O.M.; Ryan, T.E.; Arnaoutakis, G.J.; Jeng, E.I.; Ferreira, L.F. Reversible Thiol Oxidation Increases Mitochondrial Electron Transport Complex Enzyme Activity but Not Respiration in Cardiomyocytes from Patients with End-Stage Heart Failure. Cells 2022, 11, 2292. [Google Scholar] [CrossRef] [PubMed]
- Dever, T.E.; Ivanov, I.P.; Hinnebusch, A.G. Translational Regulation by uORFs and Start Codon Selection Stringency. Genes Dev. 2023, 37, 474–489. [Google Scholar] [CrossRef]
- Jaud, M.; Philippe, C.; Di Bella, D.; Tang, W.; Pyronnet, S.; Laurell, H.; Mazzolini, L.; Rouault-Pierre, K.; Touriol, C. Translational Regulations in Response to Endoplasmic Reticulum Stress in Cancers. Cells 2020, 9, 540. [Google Scholar] [CrossRef]
- Starck, S.R.; Tsai, J.C.; Chen, K.; Shodiya, M.; Wang, L.; Yahiro, K.; Martins-Green, M.; Shastri, N.; Walter, P. Translation from the 5′ Untranslated Region Shapes the Integrated Stress Response. Science 2016, 351, aad3867. [Google Scholar] [CrossRef]
- Palam, L.R.; Baird, T.D.; Wek, R.C. Phosphorylation of eIF2 Facilitates Ribosomal Bypass of an Inhibitory Upstream ORF to Enhance CHOP Translation. J. Biol. Chem. 2011, 286, 10939–10949. [Google Scholar] [CrossRef]
- Sirven, A.; Ravet, E.; Charneau, P.; Zennou, V.; Coulombel, L.; Guétard, D.; Pflumio, F.; Dubart-Kupperschmitt, A. Enhanced Transgene Expression in Cord Blood CD34+-Derived Hematopoietic Cells, Including Developing T Cells and NOD/SCID Mouse Repopulating Cells, Following Transduction with Modified TRIP Lentiviral Vectors. Mol. Ther. 2001, 3, 438–448. [Google Scholar] [CrossRef]
- Meng, Z.; Jackson, N.L.; Choi, H.; King, P.H.; Emanuel, P.D.; Blume, S.W. Alterations in RNA-binding Activities of IRES-regulatory Proteins as a Mechanism for Physiological Variability and Pathological Dysregulation of IGF-IR Translational Control in Human Breast Tumor Cells. J. Cell. Physiol. 2008, 217, 172–183. [Google Scholar] [CrossRef]
- HaileMariam, M.; Eguez, R.V.; Singh, H.; Bekele, S.; Ameni, G.; Pieper, R.; Yu, Y. S-Trap, an ultrafast sample-preparation approach for shotgun proteomics. J. Proteome Res. 2018, 17, 2917–2924. [Google Scholar] [CrossRef] [PubMed]
- Schwanhäusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global Quantification of Mammalian Gene Expression Control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Smits, A.H.; Jansen, P.W.T.C.; Poser, I.; Hyman, A.A.; Vermeulen, M. Stoichiometry of Chromatin-Associated Protein Complexes Revealed by Label-Free Quantitative Mass Spectrometry-Based Proteomics. Nucleic Acids Res. 2013, 41, e28. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
David, F.; Roussel, E.; Froment, C.; Draia-Nicolau, T.; Pujol, F.; Burlet-Schiltz, O.; Henras, A.K.; Lacazette, E.; Morfoisse, F.; Tatin, F.; et al. Mitochondrial Ribosomal Protein MRPS15 Is a Component of Cytosolic Ribosomes and Regulates Translation in Stressed Cardiomyocytes. Int. J. Mol. Sci. 2024, 25, 3250. https://doi.org/10.3390/ijms25063250
David F, Roussel E, Froment C, Draia-Nicolau T, Pujol F, Burlet-Schiltz O, Henras AK, Lacazette E, Morfoisse F, Tatin F, et al. Mitochondrial Ribosomal Protein MRPS15 Is a Component of Cytosolic Ribosomes and Regulates Translation in Stressed Cardiomyocytes. International Journal of Molecular Sciences. 2024; 25(6):3250. https://doi.org/10.3390/ijms25063250
Chicago/Turabian StyleDavid, Florian, Emilie Roussel, Carine Froment, Tangra Draia-Nicolau, Françoise Pujol, Odile Burlet-Schiltz, Anthony K. Henras, Eric Lacazette, Florent Morfoisse, Florence Tatin, and et al. 2024. "Mitochondrial Ribosomal Protein MRPS15 Is a Component of Cytosolic Ribosomes and Regulates Translation in Stressed Cardiomyocytes" International Journal of Molecular Sciences 25, no. 6: 3250. https://doi.org/10.3390/ijms25063250
APA StyleDavid, F., Roussel, E., Froment, C., Draia-Nicolau, T., Pujol, F., Burlet-Schiltz, O., Henras, A. K., Lacazette, E., Morfoisse, F., Tatin, F., Diaz, J.-J., Catez, F., Garmy-Susini, B., & Prats, A.-C. (2024). Mitochondrial Ribosomal Protein MRPS15 Is a Component of Cytosolic Ribosomes and Regulates Translation in Stressed Cardiomyocytes. International Journal of Molecular Sciences, 25(6), 3250. https://doi.org/10.3390/ijms25063250