
Nitrite Attenuates the In Vitro Inflammatory Response of Immune Cells to the SARS-CoV-2 S Protein without Interfering in the Antioxidant Enzyme Activation

 ,
, 
 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Subject Characterization
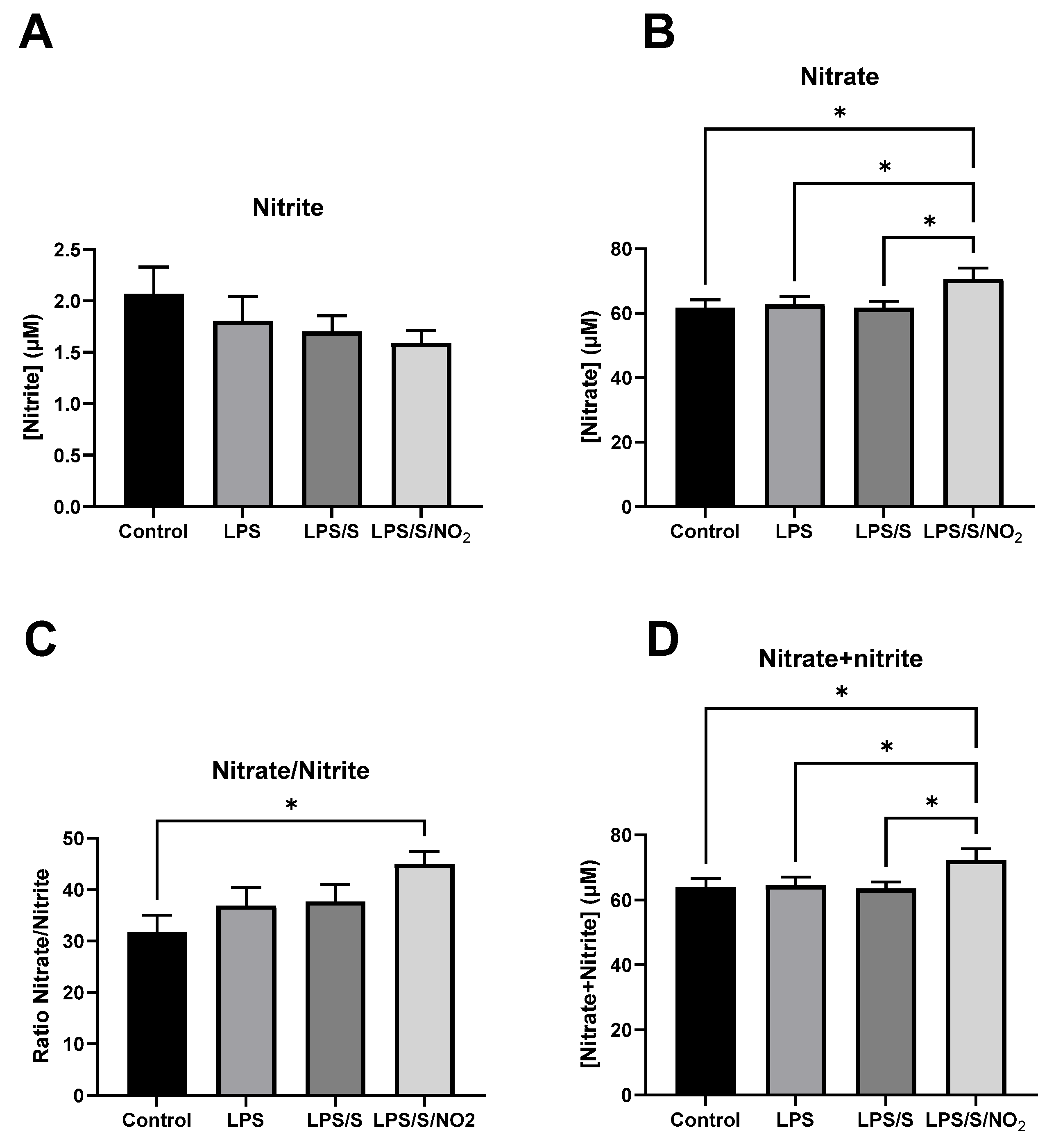
2.2. Effects of the Incubation on Nitrite and Nitrate Levels
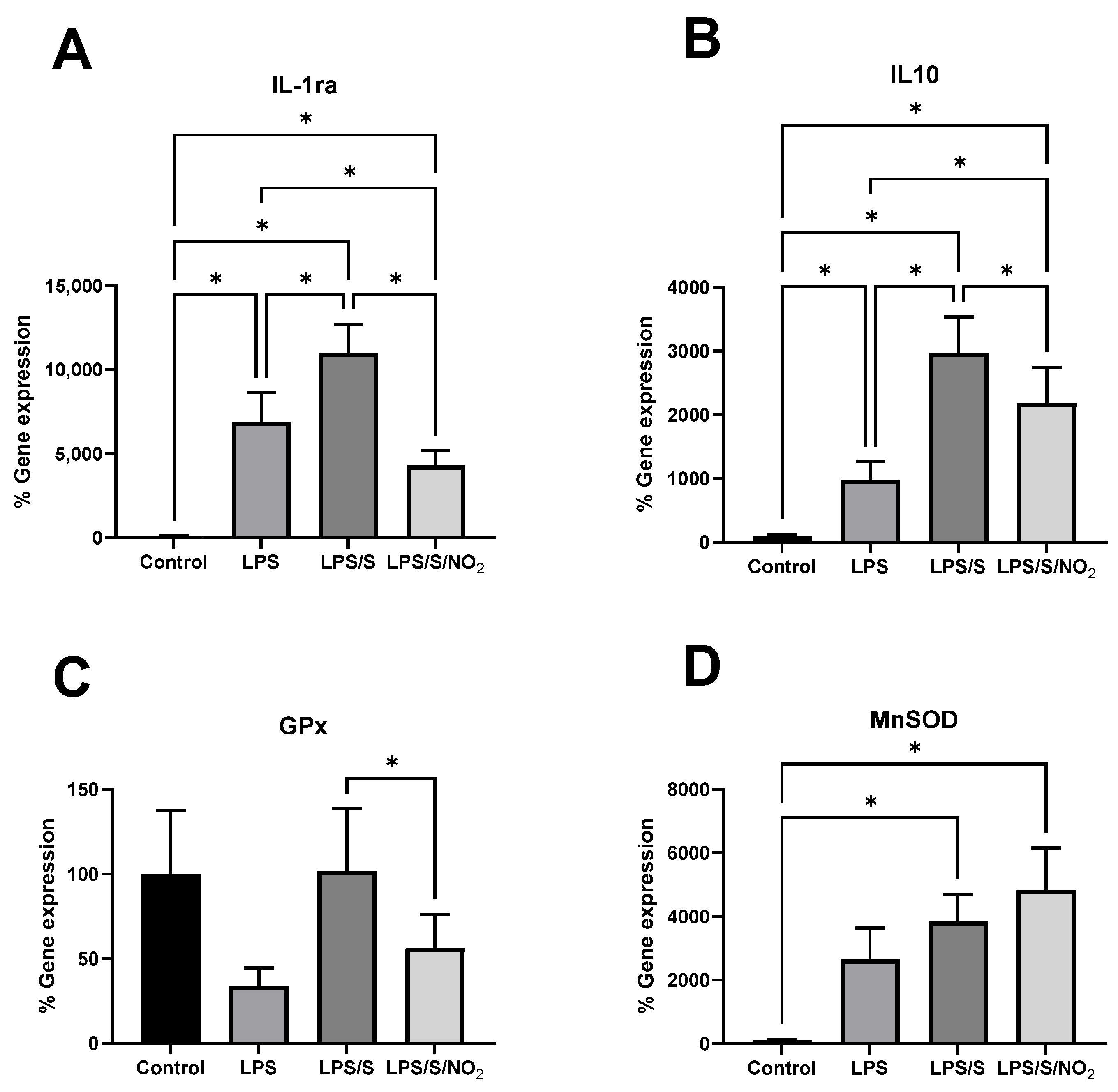
2.3. Effects of the Incubation on the Expression of Pro-inflammatory Markers
2.4. Effect of the Incubations on the Expression of Anti-Inflammatory and Antioxidant Markers
3. Discussion
4. Materials and Methods
4.1. Study Subjects and Sample Preparation
4.2. Nitrite and Nitrate Determination
4.3. RNA Isolation and Quantitative RT-PCR
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Petrilli, C.M.; Jones, S.A.; Yang, J.; Rajagopalan, H.; O’Donnell, L.; Chernyak, Y.; Tobin, K.A.; Cerfolio, R.J.; Francois, F.; Horwitz, L.I. Factors associated with hospital admission and critical illness among 5279 people with coronavirus disease 2019 in New York City: Prospective cohort study. BMJ 2020, 369, m1966. [Google Scholar] [CrossRef]
- Korakas, E.; Ikonomidis, I.; Kousathana, F.; Balampanis, K.; Kountouri, A.; Raptis, A.; Palaiodimou, L.; Kokkinos, A.; Lambadiari, V. Obesity and COVID-19: Immune and metabolic derangement as a possible link to adverse clinical outcomes. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E105–E109. [Google Scholar] [CrossRef]
- Frydrych, L.M.; Bian, G.; O’Lone, D.E.; Ward, P.A.; Delano, M.J. Obesity and type 2 diabetes mellitus drive immune dysfunction, infection development, and sepsis mortality. J. Leukoc. Biol. 2018, 104, 525–534. [Google Scholar] [CrossRef]
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef]
- Cani, P.D.; Delzenne, N.M. Interplay between obesity and associated metabolic disorders: New insights into the gut microbiota. Curr. Opin. Pharmacol. 2009, 9, 737–743. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Scherer, P.E. Preexisting and inducible endotoxemia as crucial contributors to the severity of COVID-19 outcomes. PLoS Pathog. 2021, 17, e1009306. [Google Scholar] [CrossRef]
- Petruk, G.; Puthia, M.; Petrlova, J.; Samsudin, F.; Stromdahl, A.C.; Cerps, S.; Uller, L.; Kjellstrom, S.; Bond, P.J.; Schmidtchen, A.A. SARS-CoV-2 spike protein binds to bacterial lipopolysaccharide and boosts proinflammatory activity. J. Mol. Cell Biol. 2020, 12, 916–932. [Google Scholar] [CrossRef]
- Wang, F.; Nie, J.; Wang, H.; Zhao, Q.; Xiong, Y.; Deng, L.; Song, S.; Ma, Z.; Mo, P.; Zhang, Y. Characteristics of Peripheral Lymphocyte Subset Alteration in COVID-19 Pneumonia. J. Infect. Dis. 2020, 221, 1762–1769. [Google Scholar] [CrossRef]
- Shakoory, B.; Carcillo, J.A.; Chatham, W.W.; Amdur, R.L.; Zhao, H.; Dinarello, C.A.; Cron, R.Q.; Opal, S.M. Interleukin-1 Receptor Blockade Is Associated With Reduced Mortality in Sepsis Patients With Features of Macrophage Activation Syndrome: Reanalysis of a Prior Phase III Trial. Crit. Care Med. 2016, 44, 275–281. [Google Scholar] [CrossRef]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective treatment of severe COVID-19 patients with tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar] [CrossRef]
- Li, X.; Li, T.; Wang, H. Treatment and prognosis of COVID-19: Current scenario and prospects (Review). Exp. Ther. Med. 2021, 21, 3. [Google Scholar] [CrossRef]
- Hayman, T.J.; Hsu, A.C.; Kolesnik, T.B.; Dagley, L.F.; Willemsen, J.; Tate, M.D.; Baker, P.J.; Kershaw, N.J.; Kedzierski, L.; Webb, A.I.; et al. RIPLET, and not TRIM25, is required for endogenous RIG-I-dependent antiviral responses. Immunol. Cell Biol. 2019, 97, 840–852. [Google Scholar] [CrossRef]
- Saito, T.; Gale, M., Jr. Differential recognition of double-stranded RNA by RIG-I-like receptors in antiviral immunity. J. Exp. Med. 2008, 205, 1523–1527. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Reis e Sousa, C. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef]
- Le Goffic, R.; Pothlichet, J.; Vitour, D.; Fujita, T.; Meurs, E.; Chignard, M.; Si-Tahar, M. Cutting Edge: Influenza A virus activates TLR3-dependent inflammatory and RIG-I-dependent antiviral responses in human lung epithelial cells. J. Immunol. 2007, 178, 3368–3372. [Google Scholar] [CrossRef]
- Hsu, A.C.-Y.; Wang, G.; Reid, A.T.; Veerati, P.C.; Pathinayake, P.S.; Daly, K.; Mayall, J.R.; Hansbro, P.M.; Horvat, J.C.; Wang, F.; et al. SARS-CoV-2 Spike protein promotes hyper-inflammatory response that can be ameliorated by Spike-antagonistic peptide and FDA-approved ER stress and MAP kinase inhibitors in vitro. bioRxiv 2020. [Google Scholar] [CrossRef]
- Schiffer, T.A.; Lundberg, J.O.; Weitzberg, E.; Carlstrom, M. Modulation of mitochondria and NADPH oxidase function by the nitrate-nitrite-NO pathway in metabolic disease with focus on type 2 diabetes. Biochim. Et Biophys. Acta Mol. Basis Dis. 2020, 1866, 165811. [Google Scholar] [CrossRef]
- Tian, R.; Peng, R.; Yang, Z.; Peng, Y.Y.; Lu, N. Supplementation of dietary nitrate attenuated oxidative stress and endothelial dysfunction in diabetic vasculature through inhibition of NADPH oxidase. Nitric Oxide 2020, 96, 54–63. [Google Scholar] [CrossRef]
- Peleli, M.; Ferreira, D.M.S.; Tarnawski, L.; McCann Haworth, S.; Xuechen, L.; Zhuge, Z.; Newton, P.T.; Massart, J.; Chagin, A.S.; Olofsson, P.S.; et al. Dietary nitrate attenuates high-fat diet-induced obesity via mechanisms involving higher adipocyte respiration and alterations in inflammatory status. Redox Biol. 2020, 28, 101387. [Google Scholar] [CrossRef]
- Capo, X.; Ferrer, M.D.; Olek, R.A.; Salaberry, E.; Suau, R.; Mari, B.; Llompart, I.; Tur, J.A.; Sureda, A.; Pons, A. Oral Administration of Sodium Nitrate to Metabolic Syndrome Patients Attenuates Mild Inflammatory and Oxidative Responses to Acute Exercise. Antioxidants 2020, 9, 596. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Carlstrom, M.; Larsen, F.J.; Weitzberg, E. Roles of dietary inorganic nitrate in cardiovascular health and disease. Cardiovasc. Res. 2011, 89, 525–532. [Google Scholar] [CrossRef]
- Ferrer, M.D.; Capo, X.; Reynes, C.; Quetglas, M.; Salaberry, E.; Tonolo, F.; Suau, R.; Mari, B.; Tur, J.A.; Sureda, A.; et al. Dietary Sodium Nitrate Activates Antioxidant and Mitochondrial Dynamics Genes after Moderate Intensity Acute Exercise in Metabolic Syndrome Patients. J. Clin. Med. 2021, 10, 2618. [Google Scholar] [CrossRef]
- A Healthy Lifestyle–WHO Recommendations. Available online: https://www.who.int/europe/news-room/fact-sheets/item/a-healthy-lifestyle---who-recommendations (accessed on 7 June 2023).
- Monserrat-Mesquida, M.; Quetglas-Llabres, M.; Capo, X.; Bouzas, C.; Mateos, D.; Pons, A.; Tur, J.A.; Sureda, A. Metabolic Syndrome is Associated with Oxidative Stress and Proinflammatory State. Antioxidants 2020, 9, 236. [Google Scholar] [CrossRef]
- Castro, A.M.; Macedo-de la Concha, L.E.; Pantoja-Meléndez, C.A. Low-grade inflammation and its relation to obesity and chronic degenerative diseases. Rev. Médica Del Hosp. Gen. De México 2017, 80, 101–105. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxidative Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef]
- Bansal, R.; Gubbi, S.; Muniyappa, R. Metabolic Syndrome and COVID 19: Endocrine-Immune-Vascular Interactions Shapes Clinical Course. Endocrinology 2020, 161, bqaa112. [Google Scholar] [CrossRef]
- Zhou, Y.; Chi, J.; Lv, W.; Wang, Y. Obesity and diabetes as high-risk factors for severe coronavirus disease 2019 (COVID-19). Diabetes/Metab. Res. Rev. 2021, 37, e3377. [Google Scholar] [CrossRef]
- Dejam, A.; Hunter, C.J.; Tremonti, C.; Pluta, R.M.; Hon, Y.Y.; Grimes, G.; Partovi, K.; Pelletier, M.M.; Oldfield, E.H.; Cannon, R.O., 3rd; et al. Nitrite infusion in humans and nonhuman primates: Endocrine effects, pharmacokinetics, and tolerance formation. Circulation 2007, 116, 1821–1831. [Google Scholar] [CrossRef]
- Hunault, C.C.; van Velzen, A.G.; Sips, A.J.; Schothorst, R.C.; Meulenbelt, J. Bioavailability of sodium nitrite from an aqueous solution in healthy adults. Toxicol. Lett. 2009, 190, 48–53. [Google Scholar] [CrossRef]
- Kapil, V.; Khambata, R.S.; Jones, D.A.; Rathod, K.; Primus, C.; Massimo, G.; Fukuto, J.M.; Ahluwalia, A. The Noncanonical Pathway for In Vivo Nitric Oxide Generation: The Nitrate-Nitrite-Nitric Oxide Pathway. Pharmacol. Rev. 2020, 72, 692–766. [Google Scholar] [CrossRef]
- Majumder, N.; Deepak, V.; Hadique, S.; Aesoph, D.; Velayutham, M.; Ye, Q.; Mazumder, M.H.H.; Lewis, S.E.; Kodali, V.; Roohollahi, A.; et al. Redox imbalance in COVID-19 pathophysiology. Redox Biol. 2022, 56, 102465. [Google Scholar] [CrossRef]
- Badawy, M.A.; Yasseen, B.A.; El-Messiery, R.M.; Abdel-Rahman, E.A.; Elkhodiry, A.A.; Kamel, A.G.; El-Sayed, H.; Shedra, A.M.; Hamdy, R.; Zidan, M.; et al. Neutrophil-mediated oxidative stress and albumin structural damage predict COVID-19-associated mortality. eLife 2021, 10, e69417. [Google Scholar] [CrossRef]
- Dominic, P.; Ahmad, J.; Bhandari, R.; Pardue, S.; Solorzano, J.; Jaisingh, K.; Watts, M.; Bailey, S.R.; Orr, A.W.; Kevil, C.G.; et al. Decreased availability of nitric oxide and hydrogen sulfide is a hallmark of COVID-19. Redox Biol. 2021, 43, 101982. [Google Scholar] [CrossRef]
- Alamdari, D.H.; Moghaddam, A.B.; Amini, S.; Keramati, M.R.; Zarmehri, A.M.; Alamdari, A.H.; Damsaz, M.; Banpour, H.; Yarahmadi, A.; Koliakos, G. Application of methylene blue -vitamin C -N-acetyl cysteine for treatment of critically ill COVID-19 patients, report of a phase-I clinical trial. Eur. J. Pharmacol. 2020, 885, 173494. [Google Scholar] [CrossRef]
- Lorente, L.; Gomez-Bernal, F.; Martin, M.M.; Navarro-Gonzalvez, J.A.; Argueso, M.; Perez, A.; Ramos-Gomez, L.; Sole-Violan, J.; Marcos, Y.R.J.A.; Ojeda, N.; et al. High serum nitrates levels in non-survivor COVID-19 patients. Med. Intensiv. 2022, 46, 132–139. [Google Scholar] [CrossRef]
- de Sousa, J.A.C.; Azul, F.; de Araujo, A.B.; Tome, R.C.; Silva, F.R.M.; de Vasconcelos, S.M.M.; Rios, F.J.; Leal, L. Epiisopiloturine, an Alkaloid from Pilocarpus microphyllus, Attenuates LPS-Induced Neuroinflammation by Interfering in the TLR4/NF-kappaB-MAPK Signaling Pathway in Microglial Cells. Oxidative Med. Cell. Longev. 2023, 2023, 4752502. [Google Scholar] [CrossRef]
- Kwon, J.; Arsenis, C.; Suessmilch, M.; McColl, A.; Cavanagh, J.; Morris, B.J. Differential Effects of Toll-Like Receptor Activation and Differential Mediation by MAP Kinases of Immune Responses in Microglial Cells. Cell. Mol. Neurobiol. 2022, 42, 2655–2671. [Google Scholar] [CrossRef]
- Somensi, N.; Rabelo, T.K.; Guimaraes, A.G.; Quintans-Junior, L.J.; de Souza Araujo, A.A.; Moreira, J.C.F.; Gelain, D.P. Carvacrol suppresses LPS-induced pro-inflammatory activation in RAW 264.7 macrophages through ERK1/2 and NF-kB pathway. Int. Immunopharmacol. 2019, 75, 105743. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The cytokine storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef]
- Ruan, Q.; Yang, K.; Wang, W.; Jiang, L.; Song, J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med. 2020, 46, 846–848. [Google Scholar] [CrossRef]
- Fara, A.; Mitrev, Z.; Rosalia, R.A.; Assas, B.M. Cytokine storm and COVID-19: A chronicle of pro-inflammatory cytokines. Open Biol. 2020, 10, 200160. [Google Scholar] [CrossRef]
- Capo, X.; Martorell, M.; Llompart, I.; Sureda, A.; Tur, J.A.; Pons, A. Docosahexanoic acid diet supplementation attenuates the peripheral mononuclear cell inflammatory response to exercise following LPS activation. Cytokine 2014, 69, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. CMLS 2021, 78, 1233–1261. [Google Scholar] [CrossRef]
- Sureda, A.; Martorell, M.; Bibiloni, M.D.M.; Bouzas, C.; Gallardo-Alfaro, L.; Mateos, D.; Capo, X.; Tur, J.A.; Pons, A. Effect of Free Fatty Acids on Inflammatory Gene Expression and Hydrogen Peroxide Production by Ex Vivo Blood Mononuclear Cells. Nutrients 2020, 12, 146. [Google Scholar] [CrossRef]
- Fang, H.; Wang, X.; Damarla, M.; Sun, R.; He, Q.; Li, R.; Luo, P.; Liu, J.O.; Xia, Z. Dimethyl Fumarate Protects against Lipopolysaccharide- (LPS-) Induced Sepsis through Inhibition of NF-kappaB Pathway in Mice. Mediat. Inflamm. 2023, 2023, 5133505. [Google Scholar] [CrossRef]
- Yang, T.; Peleli, M.; Zollbrecht, C.; Giulietti, A.; Terrando, N.; Lundberg, J.O.; Weitzberg, E.; Carlstrom, M. Inorganic nitrite attenuates NADPH oxidase-derived superoxide generation in activated macrophages via a nitric oxide-dependent mechanism. Free Radic. Biol. Med. 2015, 83, 159–166. [Google Scholar] [CrossRef]
- Schindler, R.; Mancilla, J.; Endres, S.; Ghorbani, R.; Clark, S.C.; Dinarello, C.A. Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumor necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood 1990, 75, 40–47. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Signaling to NF-kappaB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Sureda, A.; Batle, J.M.; Capo, X.; Martorell, M.; Cordova, A.; Tur, J.A.; Pons, A. Scuba diving induces nitric oxide synthesis and the expression of inflammatory and regulatory genes of the immune response in neutrophils. Physiol. Genom. 2014, 46, 647–654. [Google Scholar] [CrossRef]
- Deon, D.; Ahmed, S.; Tai, K.; Scaletta, N.; Herrero, C.; Lee, I.H.; Krause, A.; Ivashkiv, L.B. Cross-talk between IL-1 and IL-6 signaling pathways in rheumatoid arthritis synovial fibroblasts. J. Immunol. 2001, 167, 5395–5403. [Google Scholar] [CrossRef]
- Assar, S.; Dastbaz, M.; Amini, K.; Roghani, S.A.; Lotfi, R.; Taghadosi, M.; Kafi, H.; Abdan, Z.; Allahyari, H.; Rostampour, R.; et al. Assessing the gene expression of the adenosine 5′-monophosphate-activated protein kinase (AMPK) and its relation with the IL-6 and IL-10 plasma levels in COVID-19 patients. Mol. Biol. Rep. 2023, 50, 9925–9933. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Akerstrom, T.C.; Nielsen, A.R.; Fischer, C.P. Role of myokines in exercise and metabolism. J. Appl. Physiol. 2007, 103, 1093–1098. [Google Scholar] [CrossRef]
- Chaikijurajai, T.; Tang, W.H.W. Myeloperoxidase: A potential therapeutic target for coronary artery disease. Expert Opin. Ther. Targets 2020, 24, 695–705. [Google Scholar] [CrossRef]
- Ferrer, M.D.; Sureda, A.; Mestre, A.; Tur, J.A.; Pons, A. The double edge of reactive oxygen species as damaging and signaling molecules in HL60 cell culture. Cell. Physiol. Biochem. 2010, 25, 241–252. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age | Weight (kg) | Height (cm) | BMI (kg/cm2) | Systolic Blood Pressure (mm Hg) | Diastolic Blood Pressure (mm Hg) |
|---|---|---|---|---|---|---|---|
| 1 | Male | 67 | 95.1 | 176.5 | 30.53 | 130 | 73 |
| 2 | Female | 68 | 65.8 | 155.5 | 27.21 | 170 | 93 |
| 3 | Male | 68 | 96.2 | 166 | 34.91 | 128 | 68 |
| 4 | Male | 65 | 88.1 | 170.5 | 30.31 | 122 | 77 |
| 5 | Male | 68 | 109.7 | 174.5 | 36.03 | 170 | 88 |
| 6 | Male | 63 | 89.7 | 165.5 | 32.75 | 167 | 100 |
| Reference value | <130 | <85 |
| Measurement | Patient 1 | Patient 2 | Patient 3 | Patient 4 | Patient 5 | Patient 6 | Reference Value |
|---|---|---|---|---|---|---|---|
| Total cholesterol (mg/dL) | 170 | 124 | 132 | 151 | 110 | 253 | <200 |
| HDL (mg/dL) | 46 | 40 | 47 | 37 | 34 | 38 | ≥60 |
| LDL (mg/dL) | 95 | 63 | 71 | 65 | 60 | 181 | <100 |
| Triglycerides (mg/dL) | 147 | 106 | 73 | 244 | 133 | 172 | <149 |
| Erythrocytes (106/mm3) | 4.89 | 4.47 | 4.84 | 4.73 | 5.65 | 5.42 | 4.50–5.80 |
| Hematocrit (%) | 45.2 | 41.1 | 46.4 | 45.3 | 54.6 | 51 | 40.0–50.0 |
| Hb (g/dL) | 15 | 13.7 | 14.8 | 15.2 | 18 | 16.9 | 12.5–17.2 |
| Hba1c (%) | 5.9 | 6.3 | 6 | 6.7 | 6.9 | 5.5 | 3.8–6.2 |
| Leucocytes (103/mm3) | 7.1 | 6.75 | 7.37 | 10.5 | 5.56 | 6.74 | 4.00–11.00 |
| Neutrophils (103/mm3) | 4.12 | 3.87 | 3.71 | 6.28 | 3.04 | 3.89 | 1.8–7.5 |
| Lymphocytes (103/mm3) | 1.89 | 1.95 | 2.33 | 3.3 | 1.93 | 2.07 | 1.0–4.5 |
| Monocytes (103/mm3) | 0.84 | 0.59 | 0.72 | 0.64 | 4 | 0.55 | 2.5–13.0 |
| Eosinophils (103/mm3) | 0.23 | 0.29 | 0.53 | 0.25 | 0.12 | 0.17 | 0.5–7 |
| Basophils (103/mm3) | 0.02 | 0.05 | 0.07 | 0.07 | 0.07 | 0.06 | 0.0–2.0 |
| Platelets (103/mm3) | 231 | 231 | 190 | 190 | 196 | 214 | 150.0–400.0 |
| Variable | IL-6 | IL-1β | IL-10 | TLR4 | IL-1ra | TNFα | GPx | MnSOD |
|---|---|---|---|---|---|---|---|---|
| IL-6 | 1 | 0.537 ** | 0.724 ** | 0.246 | 0.742 ** | 0.732 ** | 0.485 ** | 0.357 * |
| IL-1β | 1 | 0.645 ** | 0.062 | 0.547 ** | 0.497 ** | 0.475 ** | 0.644 ** | |
| IL-10 | 1 | 0.520 ** | 0.797 ** | 0.658 ** | 0.444 ** | 0.558 ** | ||
| TLR4 | 1 | 0.611 ** | 0.337 * | 0.147 | 0.388 * | |||
| IL-1ra | 1 | 0.557 | 0.312 | 0.508 ** | ||||
| TNFα | 1 | 0.572 ** | 0.543 ** | |||||
| GPx | 1 | 0.523 ** | ||||||
| MnSOD | 1 |
| Gene | RV Sequence | FW Sequence | Cycle Conditions |
|---|---|---|---|
| 18S rRNA | 95 °C 10 s | ||
| 5′-GTGTAATCCGTCTCCACAGA | 5′-ATGTGAAGTCACTGTGCCAG | 60 °C 10 s | |
| 72 °C 15 s | |||
| TNFα | 95 °C 10 s | ||
| 5′-CTGGTTATCTCTCAGCTCCACGCCATT | 5′-CCCAGGCAGTCAGATCATCTTCTCGAA | 59 °C 10 s | |
| 72 °C 15 s | |||
| IL-6 | 95 °C 10 s | ||
| 5′-GTGTAATCCGTCTCCACAGA | 5′- ATGTGAAGTCACTGTGCCAG | 63 °C 10 s | |
| 72 °C 15 s | |||
| IL-1β | 95 °C 10 s | ||
| 5′-GGCAGACTCAAATTCCAGCT | 5′-GGACAGGATATGGAGCAACA | 58 °C 10 s | |
| 72 °C 15 s | |||
| GPx | 95 °C 10 s | ||
| 5′-TTCACCTCGCACTTCTCGAA | 5′-TTCCCGTGCAACCAGTTTG | 63 °C 10 s | |
| 72 °C 15 s | |||
| MnSOD | 95 °C 10 s | ||
| 5′-TGAACGTCACCGAGGAGAAG | 5′-CGTGCTCCCACACATCAATC | 60 °C 10 s | |
| 72 °C 12 s | |||
| TLR4 | 95 °C 10 s | ||
| 5′-TCAGAGGTCCATCAAACATCAC | 5′-GGTCACCTTTTCTTGATTCCA | 60 °C 10 s | |
| 72 °C 15 s | |||
| IL-10 | 95 °C 10 s | ||
| 5′-CCACGGCCTTGCTCTTGTT | 5′-AGAACCTGAAGACCCTCAGGC | 58 °C 10 s | |
| 72 °C 15 s | |||
| IL-1ra | 95 °C 10 s | ||
| 5′-CGCTCAGGTCAGTGATGTTAA | 5’-GAAGATGTGCCTGTCCTGTGT | 56 °C 10 s | |
| 72 °C 15 s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer, M.D.; Reynés, C.; Jiménez, L.; Malagraba, G.; Monserrat-Mesquida, M.; Bouzas, C.; Sureda, A.; Tur, J.A.; Pons, A. Nitrite Attenuates the In Vitro Inflammatory Response of Immune Cells to the SARS-CoV-2 S Protein without Interfering in the Antioxidant Enzyme Activation. Int. J. Mol. Sci. 2024, 25, 3001. https://doi.org/10.3390/ijms25053001
Ferrer MD, Reynés C, Jiménez L, Malagraba G, Monserrat-Mesquida M, Bouzas C, Sureda A, Tur JA, Pons A. Nitrite Attenuates the In Vitro Inflammatory Response of Immune Cells to the SARS-CoV-2 S Protein without Interfering in the Antioxidant Enzyme Activation. International Journal of Molecular Sciences. 2024; 25(5):3001. https://doi.org/10.3390/ijms25053001
Chicago/Turabian StyleFerrer, Miguel D., Clara Reynés, Laura Jiménez, Gianluca Malagraba, Margalida Monserrat-Mesquida, Cristina Bouzas, Antoni Sureda, Josep A. Tur, and Antoni Pons. 2024. "Nitrite Attenuates the In Vitro Inflammatory Response of Immune Cells to the SARS-CoV-2 S Protein without Interfering in the Antioxidant Enzyme Activation" International Journal of Molecular Sciences 25, no. 5: 3001. https://doi.org/10.3390/ijms25053001
APA StyleFerrer, M. D., Reynés, C., Jiménez, L., Malagraba, G., Monserrat-Mesquida, M., Bouzas, C., Sureda, A., Tur, J. A., & Pons, A. (2024). Nitrite Attenuates the In Vitro Inflammatory Response of Immune Cells to the SARS-CoV-2 S Protein without Interfering in the Antioxidant Enzyme Activation. International Journal of Molecular Sciences, 25(5), 3001. https://doi.org/10.3390/ijms25053001