
The Role of Polyphenols in Modulating PON1 Activity Regarding Endothelial Dysfunction and Atherosclerosis

 ,
,  ,
,  ,
,  , ,
, , 
Abstract
1. Introduction
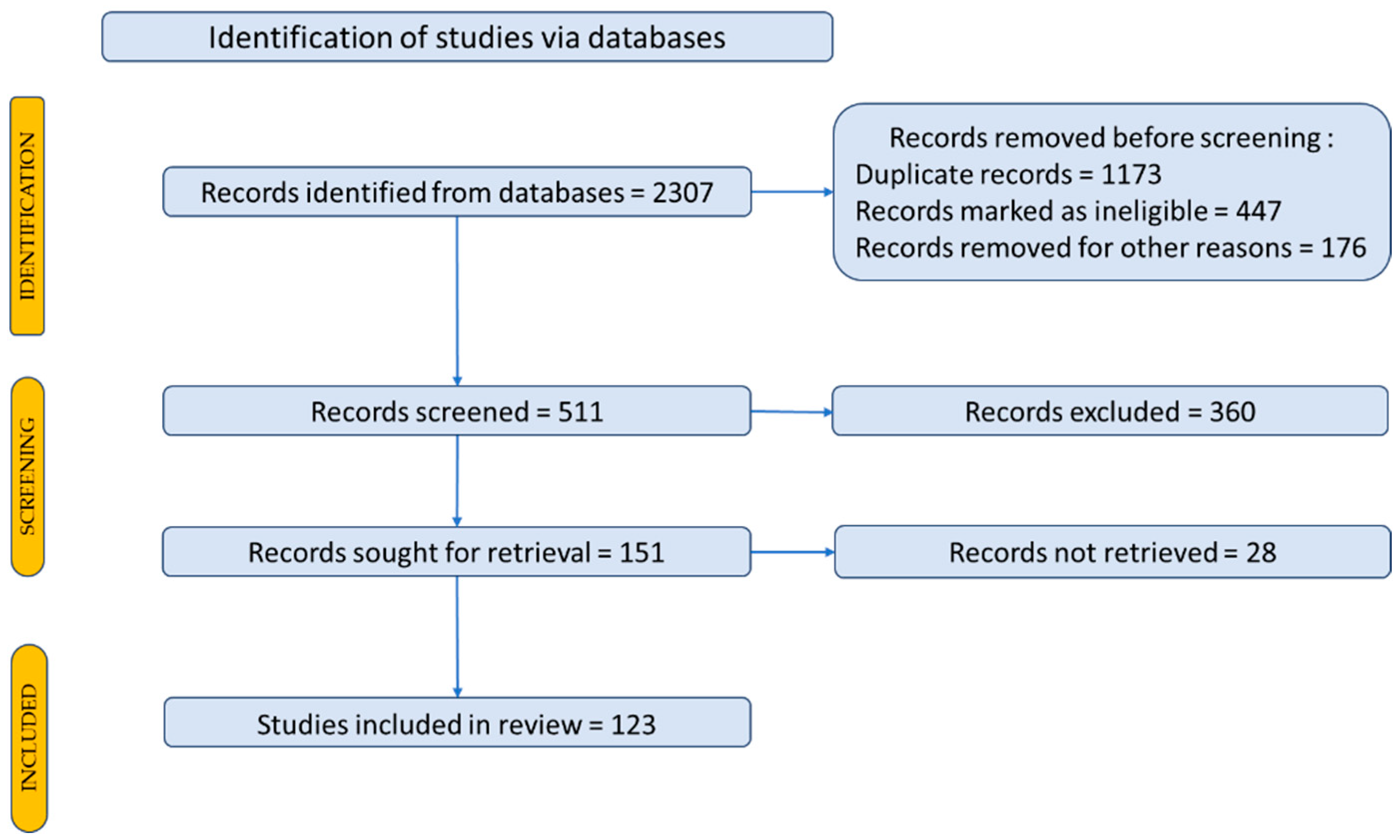
2. Materials and Methods
3. Paraoxonase 1 and Its Implications in Various Diseases
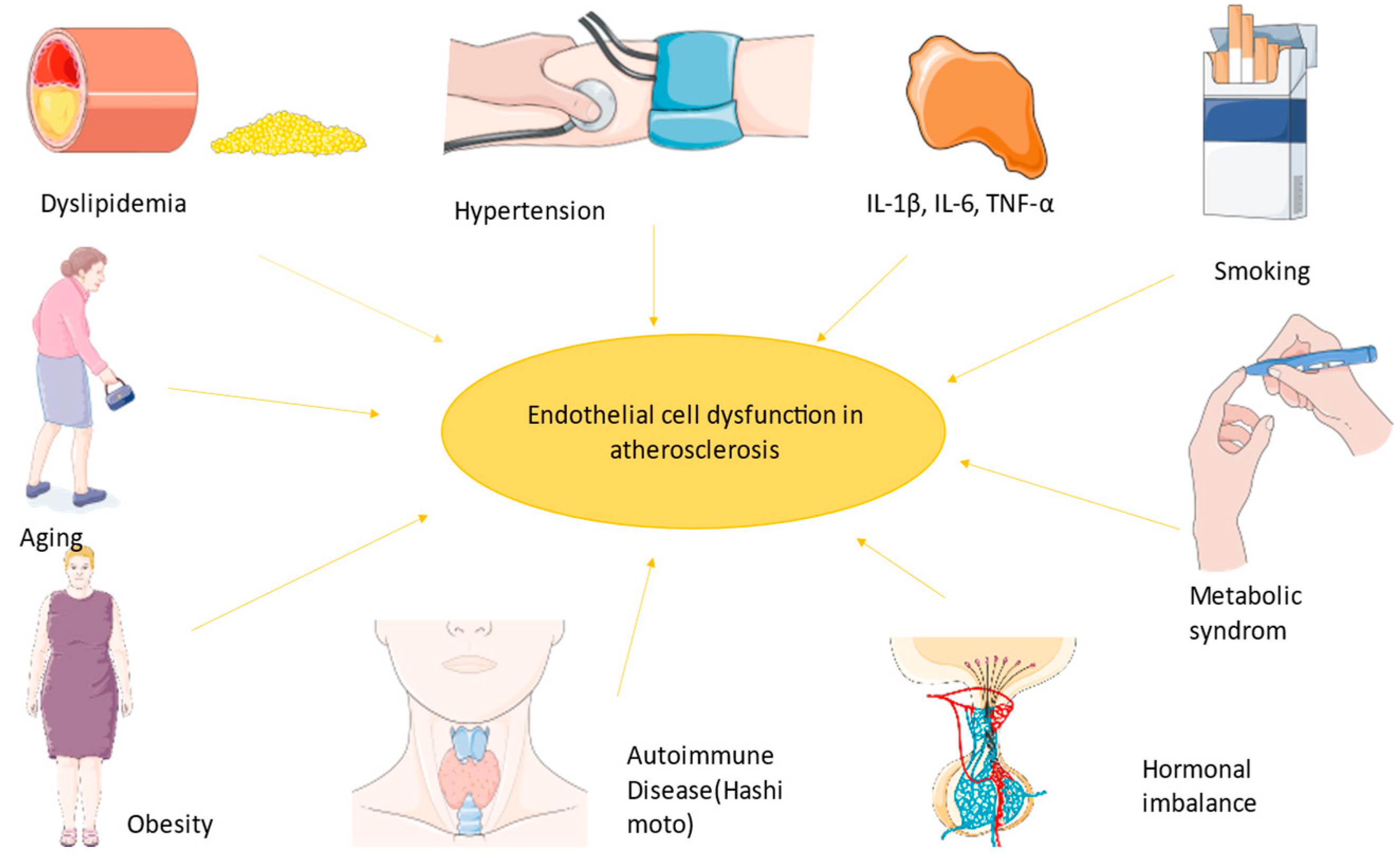
3.1. Endothelial Dysfunction
3.2. PON1 Function
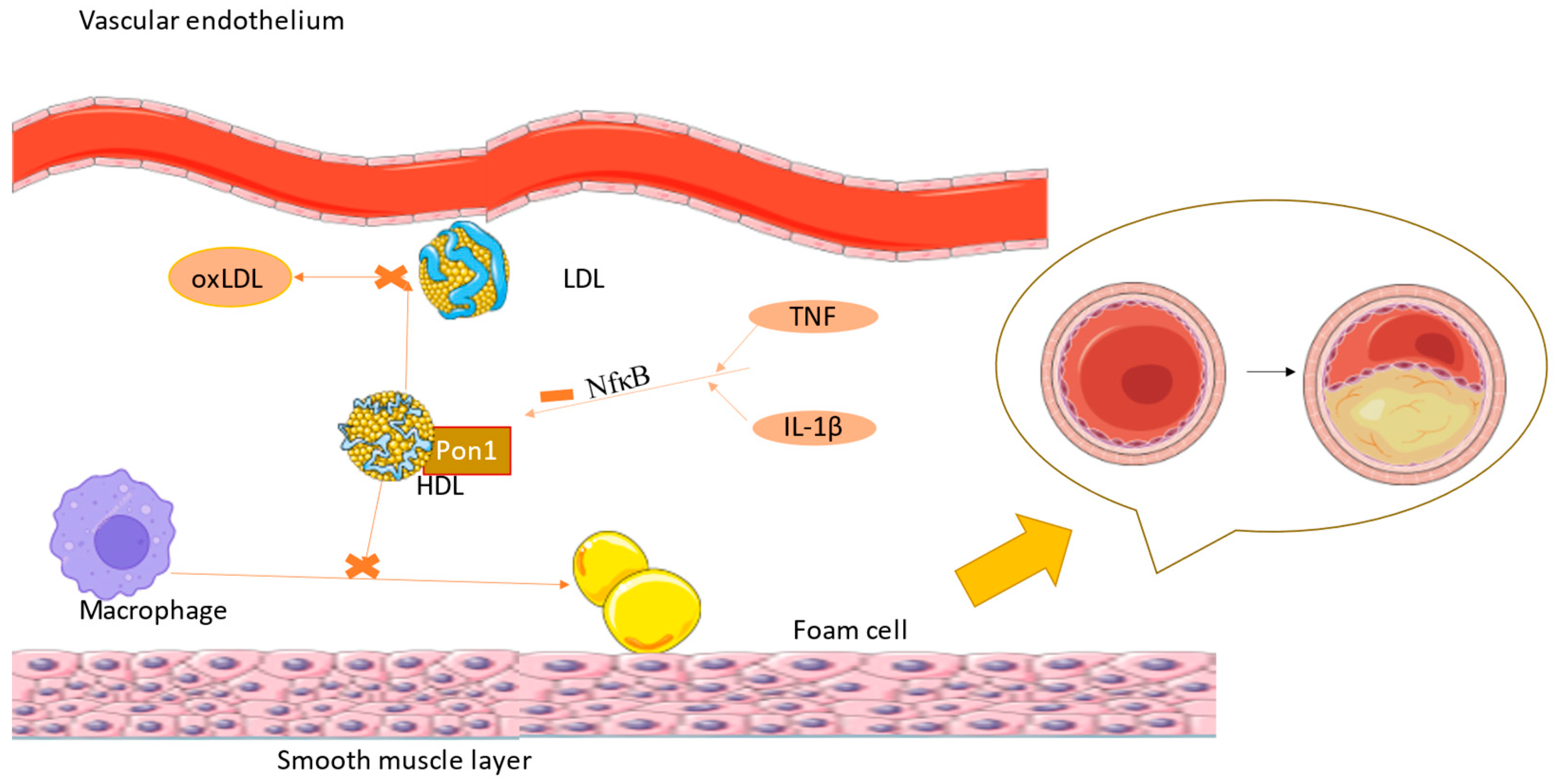
3.3. PON1 Implication in Atherogenesis and Cardiovascular Diseases
3.4. PON1 and Diabetes
3.5. PON1 and Stroke
3.6. PON1 and Other Diseases
3.7. PON1 and Therapy
3.8. PON1 and Lifestyle
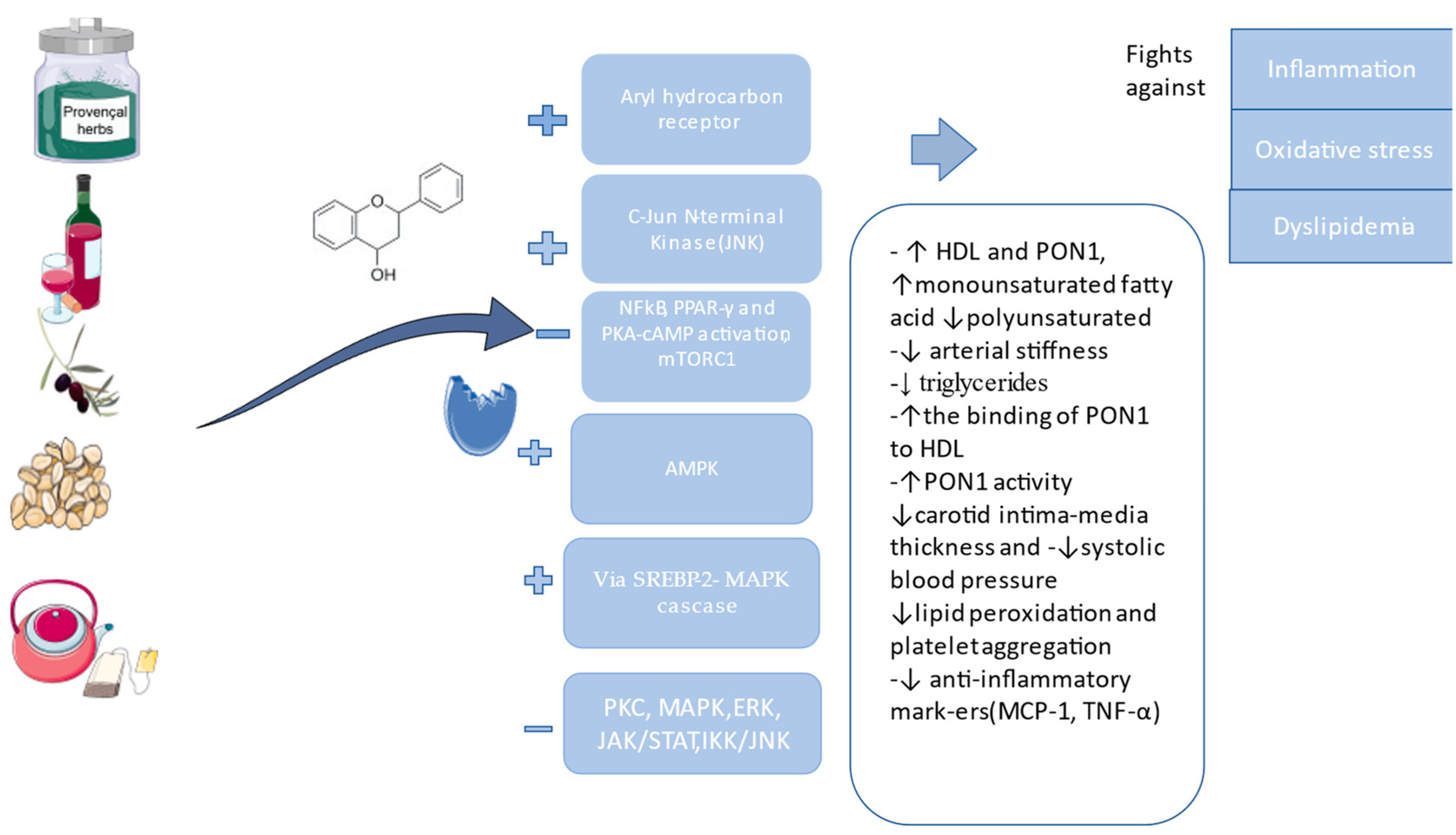
4. Plant Extracts and Phytochemical Compounds with a Positive Effect on PON1
5. Conclusions
Funding
Conflicts of Interest
Abbreviations
| ADP | adenosine diphosphate |
| AhR | aryl hydrocarbon receptor |
| AKT | serine/threonine protein kinase, also called protein kinase B. |
| AP-1 | activator protein-1 |
| ApoA-1 | apolipoprotein(a) |
| ARNT | aryl hydrocarbon receptor nuclear translocator |
| CYP1A1 | cytochrome P450 Family 1 Subfamily A Member 1 |
| CAT | chloramphenicol acetyltransferase |
| COPD | chronic obstructive pulmonary disease |
| CRP | C reactive protein |
| CVD | cardiovascular disease |
| ERK | extracellular signal-regulated protein kinase |
| FMD | flow-mediated vasodilatation |
| HDL | high-density lipoprotein |
| HT | hydroxytyrosol |
| ICAM1 | intercellular adhesion molecules |
| IL-1 | interleukin-1 |
| IL-6 | interleukin-6 |
| JAK | Janus kinase |
| JAK/STAT 3 | Janus kinase/signal transducers and activators of transcription |
| JNK | c-Jun N-terminal kinases |
| LDL | low-density lipoprotein |
| L-NAME | L-NG-Nitro arginine methyl ester |
| MAPKs | mitogen-activated protein kinases |
| MCP-1 | monocyte chemoattractant protein-1 |
| MPO | pro-oxidant myeloperoxidase |
| NAFLD | nonalcoholic fatty liver disease |
| NF-kB | nuclear factor kappa B |
| NO | nitric oxide |
| Nrf2 | nuclear factor erythroid-related factor 2 |
| PCOS | polycystic ovary syndrome |
| PKC | protein kinase C |
| PON1 | paraoxonase 1 |
| PON2 | paraoxonase 2 |
| PON3 | paraoxonase 3 |
| PPAR | γ-peroxisome-proliferator-activated receptor gamma |
| PUFA | polyunsaturated fatty acid |
| RAAS | renin–angiotensin–aldosterone system |
| ROS | reactive oxygen species |
| SR-BI | scavenger receptor class B type |
| SOD | superoxide dismutase |
| TAC | total non-enzymatic antioxidant capacity |
| TNF | tumor necrosis factor |
| VCAM1 | endothelial–leukocyte adhesion molecule |
| VLDL | very low-density lipoprotein |
References
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The Vascular Endothelium and Human Diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Gimbrone, M.A.; García-Cardeña, G. Endothelial Cell Dysfunction and the Pathobiology of Atherosclerosis. Circ. Res. 2016, 118, 620–636. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Shen, T.; Lou, H. Dietary Polyphenols and Their Biological Significance. IJMS 2007, 8, 950–988. [Google Scholar] [CrossRef]
- Sierra-Campos, E.; Valdez-Solana, M.; Avitia-Domínguez, C.; Campos-Almazán, M.; Flores-Molina, I.; García-Arenas, G.; Téllez-Valencia, A. Effects of Moringa Oleifera Leaf Extract on Diabetes-Induced Alterations in Paraoxonase 1 and Catalase in Rats Analyzed through Progress Kinetic and Blind Docking. Antioxidants 2020, 9, 840. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Moher, D. Updating Guidance for Reporting Systematic Reviews: Development of the PRISMA 2020 Statement. J. Clin. Epidemiol. 2021, 134, 103–112. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Xu, S.; Ilyas, I.; Little, P.J.; Li, H.; Kamato, D.; Zheng, X.; Luo, S.; Li, Z.; Liu, P.; Han, J.; et al. Endothelial Dysfunction in Atherosclerotic Cardiovascular Diseases and Beyond: From Mechanism to Pharmacotherapies. Pharmacol. Rev. 2021, 73, 924–967. [Google Scholar] [CrossRef]
- Cyr, A.R.; Huckaby, L.V.; Shiva, S.S.; Zuckerbraun, B.S. Nitric Oxide and Endothelial Dysfunction. Crit. Care Clin. 2020, 36, 307–321. [Google Scholar] [CrossRef]
- Meneses, M.J.; Silvestre, R.; Sousa-Lima, I.; Macedo, M.P. Macedo Paraoxonase-1 as a Regulator of Glucose and Lipid Homeostasis: Impact on the Onset and Progression of Metabolic Disorders. IJMS 2019, 20, 4049. [Google Scholar] [CrossRef]
- Schiffrin, E.L.; Park, J.B.; Pu, Q. Effect of Crossing over Hypertensive Patients from a Beta-Blocker to an Angiotensin Receptor Antagonist on Resistance Artery Structure and on Endothelial Function. J. Hypertens. 2002, 20, 71–78. [Google Scholar] [CrossRef]
- Husain, K. Inflammation, Oxidative Stress and Renin Angiotensin System in Atherosclerosis. WJBC 2015, 6, 209. [Google Scholar] [CrossRef] [PubMed]
- Landmesser, U.; Harrison, D.G. Oxidant Stress as a Marker for Cardiovascular Events: Ox Marks the Spot. Circulation 2001, 104, 2638–2640. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Galley, H.F.; Webster, N.R. Physiology of the Endothelium. Br. J. Anaesth. 2004, 93, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Best, P.J.M.; Lerman, A. Endothelin in Cardiovascular Disease: From Atherosclerosis to Heart Failure. J. Cardiovasc. Pharmacol. 2000, 35, S61–S63. [Google Scholar] [CrossRef]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative Stress and Reactive Oxygen Species in Endothelial Dysfunction Associated with Cardiovascular and Metabolic Diseases. Vasc. Pharmacol. 2018, 100, 1–19. [Google Scholar] [CrossRef]
- Tabas, I.; García-Cardeña, G.; Owens, G.K. Recent Insights into the Cellular Biology of Atherosclerosis. J. Cell Biol. 2015, 209, 13–22. [Google Scholar] [CrossRef]
- Sandoo, A.; Veldhuijzen Van Zanten, J.J.C.S.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The Endothelium and Its Role in Regulating Vascular Tone. TOCMJ 2010, 4, 302–312. [Google Scholar] [CrossRef]
- Balta, S. Endothelial Dysfunction and Inflammatory Markers of Vascular Disease. CVP 2020, 19, 243–249. [Google Scholar] [CrossRef]
- Arab, Z.N.; Khayatan, D.; Razavi, S.M.; Zare, K.; Kheradkhah, E.; Momtaz, S.; Ferretti, G.; Bacchetti, T.; Sathyapalan, T.; Emami, S.A.; et al. Phytochemicals as Modulators of Paraoxonase-1 in Health and Diseases. Antioxidants 2022, 11, 1273. [Google Scholar] [CrossRef] [PubMed]
- Taler-Verčič, A.; Goličnik, M.; Bavec, A. The Structure and Function of Paraoxonase-1 and Its Comparison to Paraoxonase-2 and -3. Molecules 2020, 25, 5980. [Google Scholar] [CrossRef] [PubMed]
- Mahrooz, A.; Mackness, M.; Bagheri, A.; Ghaffari-Cherati, M.; Masoumi, P. The Epigenetic Regulation of Paraoxonase 1 (PON1) as an Important Enzyme in HDL Function: The Missing Link between Environmental and Genetic Regulation. Clin. Biochem. 2019, 73, 1–10. [Google Scholar] [CrossRef]
- Khalil, A.; Fulop, T.; Berrougui, H. Role of Paraoxonase1 in the Regulation of High-Density Lipoprotein Functionality and in Cardiovascular Protection. Antioxid. Redox Signal. 2021, 34, 191–200. [Google Scholar] [CrossRef]
- Draganov, D.I.; La Du, B.N. Pharmacogenetics of Paraoxonases: A Brief Review. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2004, 369, 78–88. [Google Scholar] [CrossRef]
- Priyanka, K.; Singh, S.; Gill, K. Paraoxonase 3: Structure and Its Role in Pathophysiology of Coronary Artery Disease. Biomolecules 2019, 9, 817. [Google Scholar] [CrossRef]
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What Are Their Functions? Chem. Biol. Interact. 2016, 259, 51–62. [Google Scholar] [CrossRef]
- Mohamed, R.H.; Mohamed, R.H.; Karam, R.A.; Abd El-Aziz, T.A. The Relationship between Paraoxonase1-192 Polymorphism and Activity with Coronary Artery Disease. Clin. Biochem. 2010, 43, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Soflaei, S.S.; Baktashian, M.; Moghaddam, K.H.; Saberi-Karimian, M.; Kosari, N.; Hashemi, S.M.; Mouhebati, M.; Amini, M.; Dehghani, M.; Esmaily, H.; et al. Association of Paraoxonase-1 Genotype and Phenotype with Angiogram Positive Coronary Artery Disease. Arq. Bras. Cardiol. 2022, 119, 593–601. [Google Scholar] [CrossRef]
- Ashiq, S.; Ashiq, K. The Role of Paraoxonase 1 (PON1) Gene Polymorphisms in Coronary Artery Disease: A Systematic Review and Meta-Analysis. Biochem. Genet. 2021, 59, 919–939. [Google Scholar] [CrossRef] [PubMed]
- Deakin, S.P.; Bioletto, S.; Bochaton-Piallat, M.-L.; James, R.W. HDL-Associated Paraoxonase-1 Can Redistribute to Cell Membranes and Influence Sensitivity to Oxidative Stress. Free Radic. Biol. Med. 2011, 50, 102–109. [Google Scholar] [CrossRef]
- Gamliel-Lazarovich, A.; Abassi, Z.; Khatib, S.; Tavori, H.; Vaya, J.; Aviram, M.; Keidar, S. Paraoxonase1 Deficiency in Mice Is Associated with Hypotension and Increased Levels of 5,6-Epoxyeicosatrienoic Acid. Atherosclerosis 2012, 222, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Gilad, D.; Atiya, S.; Mozes-Autmazgin, Z.; Ben-Shushan, R.S.; Ben-David, R.; Amram, E.; Tamir, S.; Chuyun, D.; Szuchman-Sapir, A. Paraoxonase 1 in Endothelial Cells Impairs Vasodilation Induced by Arachidonic Acid Lactone Metabolite. Biochim. Et Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2019, 1864, 386–393. [Google Scholar] [CrossRef]
- Raz, B.-D.; Dimitry, C.; Andrea, S.-S. The Uptake Mechanism and Intracellular Fate of Paraoxonase-1 in Endothelial Cells. Free Radic. Biol. Med. 2020, 153, 26–33. [Google Scholar] [CrossRef]
- Camps, J.; Iftimie, S.; García-Heredia, A.; Castro, A.; Joven, J. Paraoxonases and Infectious Diseases. Clin. Biochem. 2017, 50, 804–811. [Google Scholar] [CrossRef]
- Durrington, P.N.; Bashir, B.; Soran, H. Paraoxonase 1 and Atherosclerosis. Front. Cardiovasc. Med. 2023, 10, 1065967. [Google Scholar] [CrossRef]
- Park, S.; Mathis, K.W.; Lee, I.K. The Physiological Roles of Apolipoprotein J/Clusterin in Metabolic and Cardiovascular Diseases. Rev. Endocr. Metab. Disord. 2014, 15, 45–53. [Google Scholar] [CrossRef]
- Kowalska, K.; Socha, E.; Milnerowicz, H. Review: The Role of Paraoxonase in Cardiovascular Diseases. Ann. Clin. Lab. Sci. 2015, 45, 226–233. [Google Scholar]
- Jeelani, H.; Tabassum, N.; Afroze, D.; Rashid, F. Association of Paraoxonase1 Enzyme and Its Genetic Single Nucleotide Polymorphisms with Cardio-Metabolic and Neurodegenerative Diseases. Gene Rep. 2020, 20, 100775. [Google Scholar] [CrossRef]
- Variji, A.; Shokri, Y.; Fallahpour, S.; Zargari, M.; Bagheri, B.; Abediankenari, S.; Alizadeh, A.; Mahrooz, A. The Combined Utility of Myeloperoxidase (MPO) and Paraoxonase 1 (PON1) as Two Important HDL-Associated Enzymes in Coronary Artery Disease: Which Has a Stronger Predictive Role? Atherosclerosis 2019, 280, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Bacchetti, T.; Ferretti, G.; Carbone, F.; Ministrini, S.; Montecucco, F.; Jamialahmadi, T.; Sahebkar, A. Dysfunctional High-Density Lipoprotein: The Role of Myeloperoxidase and Paraoxonase-1. CMC 2021, 28, 2842–2850. [Google Scholar] [CrossRef] [PubMed]
- Yunoki, K.; Naruko, T.; Inaba, M.; Inoue, T.; Nakagawa, M.; Sugioka, K.; Ohsawa, M.; Iwasa, Y.; Komatsu, R.; Itoh, A.; et al. Gender-Specific Correlation between Plasma Myeloperoxidase Levels and Serum High-Density Lipoprotein-Associated Paraoxonase-1 Levels in Patients with Stable and Unstable Coronary Artery Disease. Atherosclerosis 2013, 231, 308–314. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Hsueh, C.-M.; Chen, C.-Y.; Chen, T.-H.; Hsu, S.-L. Interleukin-6 Upregulates Paraoxonase 1 Gene Expression via an AKT/NF-κB-Dependent Pathway. Biochem. Biophys. Res. Commun. 2013, 437, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Kunutsor, S.K.; Bakker, S.J.L.; James, R.W.; Dullaart, R.P.F. Serum Paraoxonase-1 Activity and Risk of Incident Cardiovascular Disease: The PREVEND Study and Meta-Analysis of Prospective Population Studies. Atherosclerosis 2016, 245, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Zuin, M.; Trentini, A.; Marsillach, J.; D’Amuri, A.; Bosi, C.; Roncon, L.; Passaro, A.; Zuliani, G.; Mackness, M.; Cervellati, C. Paraoxonase-1 (PON-1) Arylesterase Activity Levels in Patients with Coronary Artery Disease: A Meta-Analysis. Dis. Markers 2022, 2022, 4264314. [Google Scholar] [CrossRef] [PubMed]
- Martini, D.; Del Bo’, C.; Porrini, M.; Ciappellano, S.; Riso, P. Role of Polyphenols and Polyphenol-Rich Foods in the Modulation of PON1 Activity and Expression. J. Nutr. Biochem. 2017, 48, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kunachowicz, D.; Ściskalska, M.; Kepinska, M. Modulatory Effect of Lifestyle-Related, Environmental and Genetic Factors on Paraoxonase-1 Activity: A Review. Int. J. Environ. Res. Public Health 2023, 20, 2813. [Google Scholar] [CrossRef]
- Mackness, M.; Mackness, B. Human Paraoxonase-1 (PON1): Gene Structure and Expression, Promiscuous Activities and Multiple Physiological Roles. Gene 2015, 567, 12–21. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, Y.; Fang, Y.; Liu, L.; Wu, S.; Fu, D.; Wang, X. Association between PON1 Activity and Coronary Heart Disease Risk: A Meta-Analysis Based on 43 Studies. Mol. Genet. Metab. 2012, 105, 141–148. [Google Scholar] [CrossRef]
- Bacchetti, T.; Ferretti, G.; Sahebkar, A. The Role of Paraoxonase in Cancer. Semin. Cancer Biol. 2019, 56, 72–86. [Google Scholar] [CrossRef]
- Précourt, L.-P.; Amre, D.; Denis, M.-C.; Lavoie, J.-C.; Delvin, E.; Seidman, E.; Levy, E. The Three-Gene Paraoxonase Family: Physiologic Roles, Actions and Regulation. Atherosclerosis 2011, 214, 20–36. [Google Scholar] [CrossRef]
- Bełtowski, J.; Wójcicka, G.; Mydlarczyk, M.; Jamroz, A. The Effect of Peroxisome Proliferator-Activated Receptors Alpha (PPARalpha) Agonist, Fenofibrate, on Lipid Peroxidation, Total Antioxidant Capacity, and Plasma Paraoxonase 1 (PON 1) Activity. J. Physiol. Pharmacol. 2002, 53, 463–475. [Google Scholar]
- Yesilbursa, D.; Serdar, A.; Saltan, Y.; Serdar, Z.; Heper, Y.; Guclu, S.; Cordan, J. The Effect of Fenofibrate on Serum Paraoxonase Activity and Inflammatory Markers in Patients with Combined Hyperlipidemia. Kardiol. Pol. 2005, 62, 526–530. [Google Scholar] [PubMed]
- Mackness, B.; Hine, D.; McElduff, P.; Mackness, M. High C-Reactive Protein and Low Paraoxonase1 in Diabetes as Risk Factors for Coronary Heart Disease. Atherosclerosis 2006, 186, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Van Lenten, B.J.; Wagner, A.C.; Navab, M.; Fogelman, A.M. Oxidized Phospholipids Induce Changes in Hepatic Paraoxonase and ApoJ but Not Monocyte Chemoattractant Protein-1 via Interleukin-6. J. Biol. Chem. 2001, 276, 1923–1929. [Google Scholar] [CrossRef] [PubMed]
- Istratoaie, S.; Boroş, B.; Vesa, Ş.C.; Maria Pop, R.; Cismaru, G.; Pop, D.; Vasile Milaciu, M.; Ciumărnean, L.; Văcăraş, V.; Dana Buzoianu, A. Paraoxonase 1 and Atrial Fibrillation: Is There a Relationship? Medicine 2022, 101, e31553. [Google Scholar] [CrossRef]
- Lioudaki, S.; Verikokos, C.; Kouraklis, G.; Kontopodis, N.; Markakis, G.; Ioannou, C.; Daskalopoulou, A.; Perrea, D.; Klonaris, C. Paraoxonase-1 and Symptomatic Status in Carotid Artery Disease. Ann. Vasc. Surg. 2020, 64, 355–360. [Google Scholar] [CrossRef]
- Leocádio, P.C.L.; Goulart, A.C.; Santos, I.S.; Lotufo, P.A.; Bensenor, I.M.; Alvarez-Leite, J.I. Lower Paraoxonase 1 Paraoxonase Activity Is Associated with a Worse Prognosis in Patients with Non-ST-Segment Elevation Myocardial Infarction in Long-Term Follow-Up. Coron. Artery Dis. 2022, 33, 515–522. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Melnichenko, A.A.; Orekhov, A.N.; Bobryshev, Y.V. Paraoxonase and Atherosclerosis-Related Cardiovascular Diseases. Biochimie 2017, 132, 19–27. [Google Scholar] [CrossRef]
- Pei, J.-F.; Yan, Y.-F.; Tang, X.; Zhang, Y.; Cui, S.-S.; Zhang, Z.-Q.; Chen, H.-Z.; Liu, D.-P. Human Paraoxonase Gene Cluster Overexpression Alleviates Angiotensin II-Induced Cardiac Hypertrophy in Mice. Sci. China Life Sci. 2016, 59, 1115–1122. [Google Scholar] [CrossRef][Green Version]
- Santhakumar, A.B.; Bulmer, A.C.; Singh, I. A Review of the Mechanisms and Effectiveness of Dietary Polyphenols in Reducing Oxidative Stress and Thrombotic Risk. J. Hum. Nutr. Diet. 2014, 27, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Adiga, U.; Banawalikar, N.; Menambath, D.T. Association of Paraoxonase 1 Activity and Insulin Resistance Models in Type 2 Diabetes Mellitus: Cross-Sectional Study. J. Chin. Med. Assoc. 2022, 85, 77. [Google Scholar] [CrossRef] [PubMed]
- Shokri, Y.; Variji, A.; Nosrati, M.; Khonakdar-Tarsi, A.; Kianmehr, A.; Kashi, Z.; Bahar, A.; Bagheri, A.; Mahrooz, A. Importance of Paraoxonase 1 (PON1) as an Antioxidant and Antiatherogenic Enzyme in the Cardiovascular Complications of Type 2 Diabetes: Genotypic and Phenotypic Evaluation. Diabetes Res. Clin. Pract. 2020, 161, 108067. [Google Scholar] [CrossRef]
- Vaisar, T.; Kanter, J.E.; Wimberger, J.; Irwin, A.D.; Gauthier, J.; Wolfson, E.; Bahnam, V.; Wu, I.-H.; Shah, H.; Keenan, H.A.; et al. High Concentration of Medium-Sized HDL Particles and Enrichment in HDL Paraoxonase 1 Associate With Protection From Vascular Complications in People With Long-Standing Type 1 Diabetes. Diabetes Care 2020, 43, 178–186. [Google Scholar] [CrossRef]
- Nessler, K.; Grzybczak, R.; Nessler, M.; Zalewski, J.; Gajos, G.; Windak, A. Associations between Myeloperoxidase and Paraoxonase-1 and Type 2 Diabetes in Patients with Ischemic Heart Disease. BMC Cardiovasc. Disord. 2022, 22, 521. [Google Scholar] [CrossRef]
- Koren-Gluzer, M.; Aviram, M.; Hayek, T. Paraoxonase1 (PON1) Reduces Insulin Resistance in Mice Fed a High-Fat Diet, and Promotes GLUT4 Overexpression in Myocytes, via the IRS-1/Akt Pathway. Atherosclerosis 2013, 229, 71–78. [Google Scholar] [CrossRef]
- Kota, S.K.; Meher, L.K.; Kota, S.K.; Jammula, S.; Krishna, S.V.S.; Modi, K.D. Implications of Serum Paraoxonase Activity in Obesity, Diabetes Mellitus, and Dyslipidemia. Indian J. Endocrinol. Metab. 2013, 17, 402–412. [Google Scholar] [CrossRef]
- Dada, A.O.; Ikpegbu, U.A.; Okunowo, L.O.; Ajibare, A.O.; Adekiitan, M.E.; Shasore, H.O. Plasma Paraoxonase-1 Activity Levels in Patients with Type 2 Diabetes Mellitus in Lagos State University Teaching Hospital, Lagos, Southwest Nigeria: A Cross-Sectional Study. Pan Afr. Med. J. 2023, 45, 40. [Google Scholar] [CrossRef]
- Murphy, S.J.; Werring, D.J. Stroke: Causes and Clinical Features. Medicine 2020, 48, 561–566. [Google Scholar] [CrossRef]
- Pan, Y.; He, B.; Sun, H.; Xu, T.; Pan, B.; Wang, S.; Mei, Y. Susceptibility of PON1/PON2 Genetic Variations to Ischemic Stroke Risk in a Chinese Han Population. PGPM 2020, 13, 563–570. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, K.; Wang, Q.; Ma, Y.; Liu, X. The Antioxidant Enzyme PON1: A Potential Prognostic Predictor of Acute Ischemic Stroke. Oxidative Med. Cell. Longev. 2021, 2021, e6677111. [Google Scholar] [CrossRef]
- Goswami, B.; Tayal, D.; Gupta, N.; Mallika, V. Paraoxonase: A Multifaceted Biomolecule. Clin. Chim. Acta 2009, 410, 1–12. [Google Scholar] [CrossRef]
- Zeng, Q.; Zeng, J. A Meta-Analysis on Relationship between Paraoxonase 1 Polymorphisms and Atherosclerotic Cardiovascular Diseases. Life Sci. 2019, 232, 116646. [Google Scholar] [CrossRef]
- Tajbakhsh, A.; Rezaee, M.; Rivandi, M.; Forouzanfar, F.; Afzaljavan, F.; Pasdar, A. Paraoxonase 1 (PON1) and Stroke; the Dilemma of Genetic Variation. Clin. Biochem. 2017, 50, 1298–1305. [Google Scholar] [CrossRef]
- Sun, J.; Wang, L.; Yang, Q.; Zhou, T.; Ding, X.; Yang, K.; Zhou, Z. The Association of Paraoxonase-1 Polymorphism with Carotid Artery Stenosis among Elderly Chinese Population. Oxid. Med. Cell. Longev. 2020, 2020, 3084120. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, T.; Liu, J. Role of Paraoxonase 1 Activity and Oxidative/Antioxidative Stress Markers in Patients with Acute Cerebral Infarction. Clin. Lab. 2018, 64, 180201. [Google Scholar] [CrossRef]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, Inflammation, and Metabolic Disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef]
- Kotur-Stevuljević, J.; Vekić, J.; Stefanović, A.; Zeljković, A.; Ninić, A.; Ivanišević, J.; Miljković, M.; Sopić, M.; Munjas, J.; Mihajlović, M.; et al. Paraoxonase 1 and Atherosclerosis-related Diseases. BioFactors 2020, 46, 193–205. [Google Scholar] [CrossRef]
- Bassu, S.; Mangoni, A.A.; Argiolas, D.; Carru, C.; Pirina, P.; Fois, A.G.; Zinellu, A. A Systematic Review and Meta-Analysis of Paraoxonase-1 Activity in Asthma. Clin. Exp. Med. 2023, 23, 1067–1074. [Google Scholar] [CrossRef]
- Bassu, S.; Mangoni, A.A.; Satta, R.; Argiolas, D.; Carru, C.; Zinellu, A. Paraoxonase and Arylesterase Activity of Serum PON-1 Enzyme in Psoriatic Patients: A Systematic Review and Meta-Analysis. Clin. Exp. Med. 2023, 23, 301–311. [Google Scholar] [CrossRef]
- Jeelani, H.; Ganie, M.A.; Amin, S.; Fatima, Q.; Kawa, I.A.; Manzoor, S.; Parvez, T.; Ahmad, D.N.; Rashid, F. Effect of Paraoxonase1 (PON1) Gene Polymorphisms on PON1 Activity, HDL, LDL and MDA Levels in Women with Polycystic Ovary Syndrome (PCOS): A Case-Control Study. Meta Gene 2019, 20, 100552. [Google Scholar] [CrossRef]
- Kotani, K.; Watanabe, J.; Miura, K.; Gugliucci, A. Paraoxonase 1 and Non-Alcoholic Fatty Liver Disease: A Meta-Analysis. Molecules 2021, 26, 2323. [Google Scholar] [CrossRef]
- Deakin, S.; Leviev, I.; Guernier, S.; James, R.W. Simvastatin Modulates Expression of the PON1 Gene and Increases Serum Paraoxonase: A Role for Sterol Regulatory Element–Binding Protein-2. ATVB 2003, 23, 2083–2089. [Google Scholar] [CrossRef]
- Kumar, A. Effect of Simvastatin on Paraoxonase 1 (PON1) Activity and Oxidative Stress. Asian Pac. J. Trop. Med. 2010, 3, 310–314. [Google Scholar] [CrossRef]
- De Souza, J.A.; Menin, A.; Lima, L.O.; Smiderle, L.; Hutz, M.H.; Van Der Sand, C.R.; Van Der Sand, L.C.; Ferreira, M.E.W.; Pires, R.C.; Almeida, S.; et al. PON1 Polymorphisms Are Predictors of Ability to Attain HDL-C Goals in Statin-Treated Patients. Clin. Biochem. 2015, 48, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Akram, N.; Mustafa, G.; Hanif, A.A.; Tawwab, S.; Hussain, S.; Kaul, H.; Mohsin, S. Cytochrome 2C19 and Paraoxonase-1 Polymorphisms and Clopidogrel Resistance in Ischemic Heart Disease Patients. Per. Med. 2019, 16, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Halim, H.; Pinkaew, D.; Chunhacha, P.; Sinthujaroen, P.; Thiagarajan, P.; Fujise, K. Ticagrelor Induces Paraoxonase-1 (PON1) and Better Protects Hypercholesterolemic Mice against Atherosclerosis Compared to Clopidogrel. PLoS ONE 2019, 14, e0218934. [Google Scholar] [CrossRef]
- Blatter-Garin, M.C.; Kalix, B.; De Pree, S.; James, R.W. Aspirin Use Is Associated with Higher Serum Concentrations of the Anti-Oxidant Enzyme, Paraoxonase-1. Diabetologia 2003, 46, 594–595. [Google Scholar] [CrossRef]
- Turfaner, N.; Uzun, H.; Balci, H.; Ercan, M.A.; Karter, Y.H.; Caner, M.; Sipahioglu, F.; Genc, H. Ezetimibe Therapy and Its Influence on Oxidative Stress and Fibrinolytic Activity. South. Med. J. 2010, 103, 428–433. [Google Scholar] [CrossRef]
- Macan, M.; Vrkić, N.; Vrdoljak, A.L.; Radić, B.; Bradamante, V. Effects of High Sucrose Diet, Gemfibrozil, and Their Combination on Plasma Paraoxonase 1 Activity and Lipid Levels in Rats. Acta Biochim. Pol. 2010, 57, 321–326. [Google Scholar] [CrossRef]
- Mouhamed, D.H.; Ezzaher, A.; Araoud, M.; Neffati, F.; Douki, W.; Fadhel Najjar, M. Etude de l’activité de La Paraoxonase 1 (PON1) et Du Profil Lipidique Dans Une Population de Fumeurs Tunisiens. Ann. De Biol. Clin. 2010, 68, 143–147. [Google Scholar] [CrossRef]
- Otocka-Kmiecik, A. Effect of Carotenoids on Paraoxonase-1 Activity and Gene Expression. Nutrients 2022, 14, 2842. [Google Scholar] [CrossRef]
- Rao, M.N.; Marmillot, P.; Gong, M.; Palmer, D.A.; Seeff, L.B.; Strader, D.B.; Lakshman, M.R. Light, but Not Heavy Alcohol Drinking, Stimulates Paraoxonase by Upregulating Liver mRNA in Rats and Humans. Metabolism 2003, 52, 1287–1294. [Google Scholar] [CrossRef]
- Seres, I.; Bajnok, L.; Harangi, M.; Sztanek, F.; Koncsos, P.; Paragh, G. Alteration of PON1 Activity in Adult and Childhood Obesity and Its Relation to Adipokine Levels. In Paraoxonases in Inflammation, Infection, and Toxicology; Reddy, S.T., Ed.; Advances in Experimental Medicine and Biology; Humana Press: Totowa, NJ, USA, 2010; Volume 660, pp. 129–142. ISBN 978-1-60761-349-7. [Google Scholar]
- Cakmak, A.; Zeyrek, D.; Atas, A.; Erel, O. Paraoxonase Activity in Athletic Adolescents. Pediatr. Exerc. Sci. 2010, 22, 93–104. [Google Scholar] [CrossRef]
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR Signaling Pathways and Regulatory Functions. Biochim. Open 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Gouédard, C.; Barouki, R.; Morel, Y. Induction of the Paraoxonase-1 Gene Expression by Resveratrol. ATVB 2004, 24, 2378–2383. [Google Scholar] [CrossRef]
- Zhu, K.; Meng, Q.; Zhang, Z.; Yi, T.; He, Y.; Zheng, J.; Lei, W. Aryl Hydrocarbon Receptor Pathway: Role, Regulation and Intervention in Atherosclerosis Therapy (Review). Mol. Med. Rep. 2019, 20, 4763–4773. [Google Scholar] [CrossRef]
- Luna-Castillo, K.P.; Lin, S.; Muñoz-Valle, J.F.; Vizmanos, B.; López-Quintero, A.; Márquez-Sandoval, F. Functional Food and Bioactive Compounds on the Modulation of the Functionality of HDL-C: A Narrative Review. Nutrients 2021, 13, 1165. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.; Puri, B.K.; Bortolasci, C.C.; Carvalho, A.; Berk, M.; Walder, K.; Moreira, E.G.; Maes, M. The Role of High-Density Lipoprotein Cholesterol, Apolipoprotein A and Paraoxonase-1 in the Pathophysiology of Neuroprogressive Disorders. Neurosci. Biobehav. Rev. 2021, 125, 244–263. [Google Scholar] [CrossRef] [PubMed]
- Chu, A.J. Quarter-Century Explorations of Bioactive Polyphenols: Diverse Health Benefits. FBL 2022, 27, 134. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, N.; Aksoy, M.; Bagci, C.; Gergerlioglu, H.S.; Celik, H.; Herken, E.; Yaman, A.; Tarakcioglu, M.; Soydinc, S.; Sari, I.; et al. Pistachio Intake Increases High Density Lipoprotein Levels and Inhibits Low-Density Lipoprotein Oxidation in Rats. Tohoku J. Exp. Med. 2007, 212, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Barreca, D.; Gervasi, T.; Roussell, M.A.; Klein, B.; Feeney, M.J.; Carughi, A. Pistachio Nuts (Pistacia vera L.): Production, Nutrients, Bioactives and Novel Health Effects. Plants 2022, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Seeberger, J.; Alberico, T.; Wang, C.; Wheeler, C.T.; Schauss, A.G.; Zou, S. Açai Palm Fruit (Euterpe oleracea Mart.) Pulp Improves Survival of Flies on a High Fat Diet. Exp. Gerontol. 2010, 45, 243–251. [Google Scholar] [CrossRef]
- Pérez Méndez, O.; García Hernández, L. [High-density lipoproteins (HDL) size and composition are modified in the rat by a diet supplemented with “Hass” avocado (Persea americana Miller)]. Arch. Cardiol. Mex. 2007, 77, 17–24. [Google Scholar] [PubMed]
- Alidadi, M.; Jamialahmadi, T.; Cicero, A.F.G.; Bianconi, V.; Pirro, M.; Banach, M.; Sahebkar, A. The Potential Role of Plant-Derived Natural Products in Improving Arterial Stiffness: A Review of Dietary Intervention Studies. Trends Food Sci. Technol. 2020, 99, 426–440. [Google Scholar] [CrossRef]
- Lou-Bonafonte, J.; Gabás-Rivera, C.; Navarro, M.; Osada, J. The Search for Dietary Supplements to Elevate or Activate Circulating Paraoxonases. IJMS 2017, 18, 416. [Google Scholar] [CrossRef] [PubMed]
- Tirla, A.; Timar, A.V.; Becze, A.; Memete, A.R.; Vicas, S.I.; Popoviciu, M.S.; Cavalu, S. Designing New Sport Supplements Based on Aronia melanocarpa and Bee Pollen to Enhance Antioxidant Capacity and Nutritional Value. Molecules 2023, 28, 6944. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, C.B.; Mellbye, F.B.; Hermansen, K.; Jeppesen, P.B.; Gregersen, S. Effects of Aronia melanocarpa on Cardiometabolic Diseases: A Systematic Review of Quasi-Design Studies and Randomized Controlled Trials. Rev. Diabet. Stud. 2022, 18, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, H.; Hegazi, N.; El-Shamy, S.; Farag, M.A. Pomegranate Juice as a Functional Food: A Comprehensive Review of Its Polyphenols, Therapeutic Merits, and Recent Patents. Food Funct. 2020, 11, 5768–5781. [Google Scholar] [CrossRef] [PubMed]
- Ziółkiewicz, A.; Kasprzak-Drozd, K.; Rusinek, R.; Markut-Miotła, E.; Oniszczuk, A. The Influence of Polyphenols on Atherosclerosis Development. Int. J. Mol. Sci. 2023, 24, 7146. [Google Scholar] [CrossRef] [PubMed]
- Abudahab, S.; Price, E.T.; Dozmorov, M.G.; Deshpande, L.S.; McClay, J.L. The Aryl Hydrocarbon Receptor, Epigenetics and the Aging Process. J. Nutr. Health Aging 2023, 27, 291–300. [Google Scholar] [CrossRef]
- Garjani, A.; Fathiazad, F.; Zakheri, A.; Akbari, N.A.; Azarmie, Y.; Fakhrjoo, A.; Andalib, S.; Maleki-Dizaji, N. The Effect of Total Extract of Securigera securidaca L. Seeds on Serum Lipid Profiles, Antioxidant Status, and Vascular Function in Hypercholesterolemic Rats. J. Ethnopharmacol. 2009, 126, 525–532. [Google Scholar] [CrossRef]
- Alizadeh-Fanalou, S.; Nazarizadeh, A.; Babaei, M.; Khosravi, M.; Farahmandian, N.; Bahreini, E. Effects of Securigera Securidaca (L.) Degen & Dorfl Seed Extract Combined with Glibenclamide on Paraoxonase1 Activity, Lipid Profile and Peroxidation, and Cardiovascular Risk Indices in Diabetic Rats. Bioimpacts 2019, 10, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, G.; Wang, Y.; Zhang, Y. Moringa oleifera Leaf Flavonoids Protect Bovine Mammary Epithelial Cells from Hydrogen Peroxide-induced Oxidative Stress in Vitro. Reprod. Dom. Anim. 2020, 55, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Aekthammarat, D.; Pannangpetch, P.; Tangsucharit, P. Moringa oleifera Leaf Extract Induces Vasorelaxation via Endothelium-Dependent Hyperpolarization and Calcium Channel Blockade in Mesenteric Arterial Beds Isolated from L-NAME Hypertensive Rats. Clin. Exp. Hypertens. 2020, 42, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.-Y.; Choi, M.-S. Chapter 44—Polyphenols and Nutrigenetic/Nutrigenomic Associations With Obesity-Related Metabolic Diseases. In Principles of Nutrigenetics and Nutrigenomics; Caterina, R.D., Martinez, J.A., Kohlmeier, M., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 327–334. ISBN 978-0-12-804572-5. [Google Scholar]
- Li, H.; Zhang, Q. Research Progress of Flavonoids Regulating Endothelial Function. Pharmaceuticals 2023, 16, 1201. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhao, J.; Xin, Q.; Yuan, R.; Miao, Y.; Yang, M.; Mo, H.; Chen, K.; Cong, W. Protective Effects of Oleic Acid and Polyphenols in Extra Virgin Olive Oil on Cardiovascular Diseases. Food Sci. Hum. Wellness 2024, 13, 529–540. [Google Scholar] [CrossRef]
- Prakash Mishra, A.; Sharifi-Rad, M.; Shariati, M.A.; Mabkhot, Y.N.; Al-Showiman, S.S.; Rauf, A.; Salehi, B.; Župunski, M.; Sharifi-Rad, M.; Gusain, P.; et al. Bioactive Compounds and Health Benefits of Edible Rumex Species-A Review. Cell. Mol. Biol. 2018, 64, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Bello, O.M.; Fasinu, P.S.; Bello, O.E.; Ogbesejana, A.B.; Adetunji, C.O.; Dada, A.O.; Ibitoye, O.S.; Aloko, S.; Oguntoye, O.S. Wild Vegetable Rumex acetosa Linn.: Its Ethnobotany, Pharmacology and Phytochemistry—A Review. S. Afr. J. Bot. 2019, 125, 149–160. [Google Scholar] [CrossRef]
- Li, J.-J.; Li, Y.-X.; Li, N.; Zhu, H.-T.; Wang, D.; Zhang, Y.-J. The Genus Rumex (Polygonaceae): An Ethnobotanical, Phytochemical and Pharmacological Review. Nat. Prod. Bioprospect. 2022, 12, 21. [Google Scholar] [CrossRef]
- Jeong, D.; Irfan, M.; Lee, D.-H.; Hong, S.-B.; Oh, J.-W.; Rhee, M.H. Rumex acetosa Modulates Platelet Function and Inhibits Thrombus Formation in Rats. BMC Complement. Med. Ther. 2020, 20, 98. [Google Scholar] [CrossRef]
- Qamar, H.M.-U.-D.; Qayyum, R.; Salma, U.; Khan, S.; Khan, T.; Shah, A.J. Vascular Mechanisms Underlying the Hypotensive Effect of Rumex Acetosa. Pharm. Biol. 2018, 56, 225–234. [Google Scholar] [CrossRef]
- Kucekova, Z.; Mlcek, J.; Humpolicek, P.; Rop, O.; Valasek, P.; Saha, P. Phenolic Compounds from Allium schoenoprasum, Tragopogon pratensis and Rumex acetosa and Their Antiproliferative Effects. Molecules 2011, 16, 9207–9217. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.-J.; Choi, J.-H.; Koo, B.-S.; Ryu, S.-Y.; Han, Y.-H.; Lee, S.-I.; Lee, D.-U. Antimutagenicity and Cytotoxicity of the Constituents from the Aerial Parts of Rumex acetosa. Biol. Pharm. Bull. 2005, 28, 2158–2161. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Rizvi, S.I. Onion Extract (Allium cepa L.), Quercetin and Catechin up-Regulate Paraoxonase 1 Activity with Concomitant Protection against Low-Density Lipoprotein Oxidation in Male Wistar Rats Subjected to Oxidative Stress: Up-Regulation of PON1 Activity and Protection of LDL Oxidation by Onion Extract and Flavonoids. J. Sci. Food Agric. 2014, 94, 2752–2757. [Google Scholar] [CrossRef] [PubMed]
- Kaya, E.D.; Erğun, B.; Demir, Y.; Alım, Z.; Beydemir, Ş. The In Vitro Impacts of Some Plant Extracts on Carbonic Anhydrase I, II and Paraoxonase-1. Hacet. J. Biol. Chem. 2019, 47, 51–59. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1. Preservation of HDL by fighting against its oxidation [23] -higher cholesterol efflux [23] |
| 2. Preventing LDL oxidation [23] -less oxidized lipids, which are responsible for inflammation [49] -lower LDL levels [23] |
| 3. Protects against insulin resistance [9] |
| 4. Ameliorates effects of oxidized LDL [23] -↓ inflammatory and cytotoxic oxidized phospholipids [23] -↓ LDL uptake by macrophages [23] -↓ monocyte transmigration induced by oxidized LDL [23] |
| 5. Atheroprotective [21] -decreases lipid peroxides in atherosclerotic lesions [23] -reduces macrophage oxidative stress and the ability of macrophages to oxidize LDL [21] -contributes to the metabolism of homocysteine thiolactones [21] -prevents the oxidative inactivation of lecithin cholesterol acyltransferase [21] -reduces monocyte–macrophage inflammatory response [21] -reduces foam cell formation [72] -restores normal endothelial function [72] |
| 6. Antiapoptotic [76] |
| 7. Vasodilative [76] |
| Plant Extracts | Compounds | Mechanism of Action | Refs. |
|---|---|---|---|
| Pistachia vera L. | lutein, β-carotene, and γ-tocopherol in addition to containing selenium, flavonoids, and proanthocyanidins | -increases HDL and PON1, enhancing antioxidative defense by ↑ monounsaturated fatty acid and ↓polyunsaturated and saturated fatty acid uptake -reduces arterial stiffness -degrades NF-kB, TNF-α, and IL-1β | [100,101,104] |
| Euterpe oleracea Mart. | polyphenolic flavonoid–anthocyanins (cyanidin 3-rutinoside, cyanidin 3-glucoside, cyanidin 3-sambubioside, peonidin 3-rutinoside, peonidin 3-glucoside) and flavonoids (chrysoeriol, luteolin orientin, homoorientin, quercetin, isovitexin, vitexin, dihydrokaempferol) | -long-lasting vasodilation dependent on activation of the nitric oxide–cGMP pathway using endothelium-derived hyperpolarizing factor (EDHF) | [102] |
| Persea americana Mill. | -↓triglycerides -↑HDL-cholesterol and activity of PON1 | [49] | |
| Aronia melanocarpa Michx.-Elliott | anthocyanins (cyanidin–glucoside, cyanidin–arabinoside), phenolic acids, proanthocyanidins, and flavonols (quercetin–rutinoside (rutin quercetin)) -tannins, anthocyanins (punicalagin, punicalain, gallic acid, and urolithins A and B) | -↑binding of PON1 to HDL -↑PON1 activity -↓carotid intima media thickness -↓systolic blood pressure -↓lipid peroxidation and platelet aggregation -modulates NF-kB pathway | [105,106,107] |
| Securigera securidaca L. | phenolic acids, cardenolides, flavonoids, pentacyclic triterpenoid-type saponins, steroidal | -↓oxidative stress -↑antioxidant defense system -regulates the balance between antioxidant/oxidant -antihyperlipidemic -↑PON1 -↓markers of lipid peroxidation (MDA) -no effect on hs-CRP, TNF-α | [19,111] |
| Moringa oleifera Lam. | phenolic acids, flavonoids, tannins, triterpenes, glucosinolates | -protects against oxidative stress in the liver cells -vasorelaxation inhibits Ca channels -reduces diabetes onset -↑PON1, CAT, and HDL, -↓triglyceride and LDL | [99] |
| Rumex acetosa L. | Phenols (cis-resveratrol, trans-resveratrol, vanillic acid, sinapic acid, catechin rutin, hyproside, quercetin, avicularin, orientin, and iso-orientin) anthraquinones, tannins, alkaloids, stilbenes, lignans, naphthalenes, diterpene terpenes | -↓LDL cholesterol, -↓total cholesterol, -↓liver enzyme levels -↑HDL levels but improved glucose tolerance -↑storage of glycogen in the liver -antimutagenic activity -inhibit thrombus formation (inhibits the phosphorylation of ERK1/2 and JNK, lowered ATP release in collagen-stimulated platelets) | [119,120,121] |
| Allium cepa L. | quercetin and catechin | -protection of LDL oxidation and lipid peroxidation | [125] |
| Olea europaea L. | mono- and polyunsaturated fatty acid | -↑PON1 activity (PON and arylesterase) | [49] |
| Resveratrol(3,5,4′-trihydroxystilbene) | -modulates NF-kB pathway -↓LDL, ↑HDL and PON1 activity -↓anti-inflammatory markers (MCP-1, TNF-α) | [19] | |
| Luteolin(3′,4′,5,7-tetrahydroxyflavone) | -suppresses TNF-α, MCP-1, VCAM-1, IKBα/NF-kB pathway -inhibits the JAK/STAT3 pathway, IL-1β | [115,116] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirca, T.B.; Mureșan, M.E.; Pallag, A.; Marian, E.; Jurca, T.; Vicaș, L.G.; Tunduc, I.P.; Manole, F.; Ștefan, L. The Role of Polyphenols in Modulating PON1 Activity Regarding Endothelial Dysfunction and Atherosclerosis. Int. J. Mol. Sci. 2024, 25, 2962. https://doi.org/10.3390/ijms25052962
Sirca TB, Mureșan ME, Pallag A, Marian E, Jurca T, Vicaș LG, Tunduc IP, Manole F, Ștefan L. The Role of Polyphenols in Modulating PON1 Activity Regarding Endothelial Dysfunction and Atherosclerosis. International Journal of Molecular Sciences. 2024; 25(5):2962. https://doi.org/10.3390/ijms25052962
Chicago/Turabian StyleSirca, Teodora Bianca, Mariana Eugenia Mureșan, Annamaria Pallag, Eleonora Marian, Tunde Jurca, Laura Grațiela Vicaș, Ioana Paula Tunduc, Felicia Manole, and Liana Ștefan. 2024. "The Role of Polyphenols in Modulating PON1 Activity Regarding Endothelial Dysfunction and Atherosclerosis" International Journal of Molecular Sciences 25, no. 5: 2962. https://doi.org/10.3390/ijms25052962
APA StyleSirca, T. B., Mureșan, M. E., Pallag, A., Marian, E., Jurca, T., Vicaș, L. G., Tunduc, I. P., Manole, F., & Ștefan, L. (2024). The Role of Polyphenols in Modulating PON1 Activity Regarding Endothelial Dysfunction and Atherosclerosis. International Journal of Molecular Sciences, 25(5), 2962. https://doi.org/10.3390/ijms25052962