
Lutein Production and Extraction from Microalgae: Recent Insights and Bioactive Potential

 ,
,  ,
, 
 and
and 
Abstract
1. Introduction
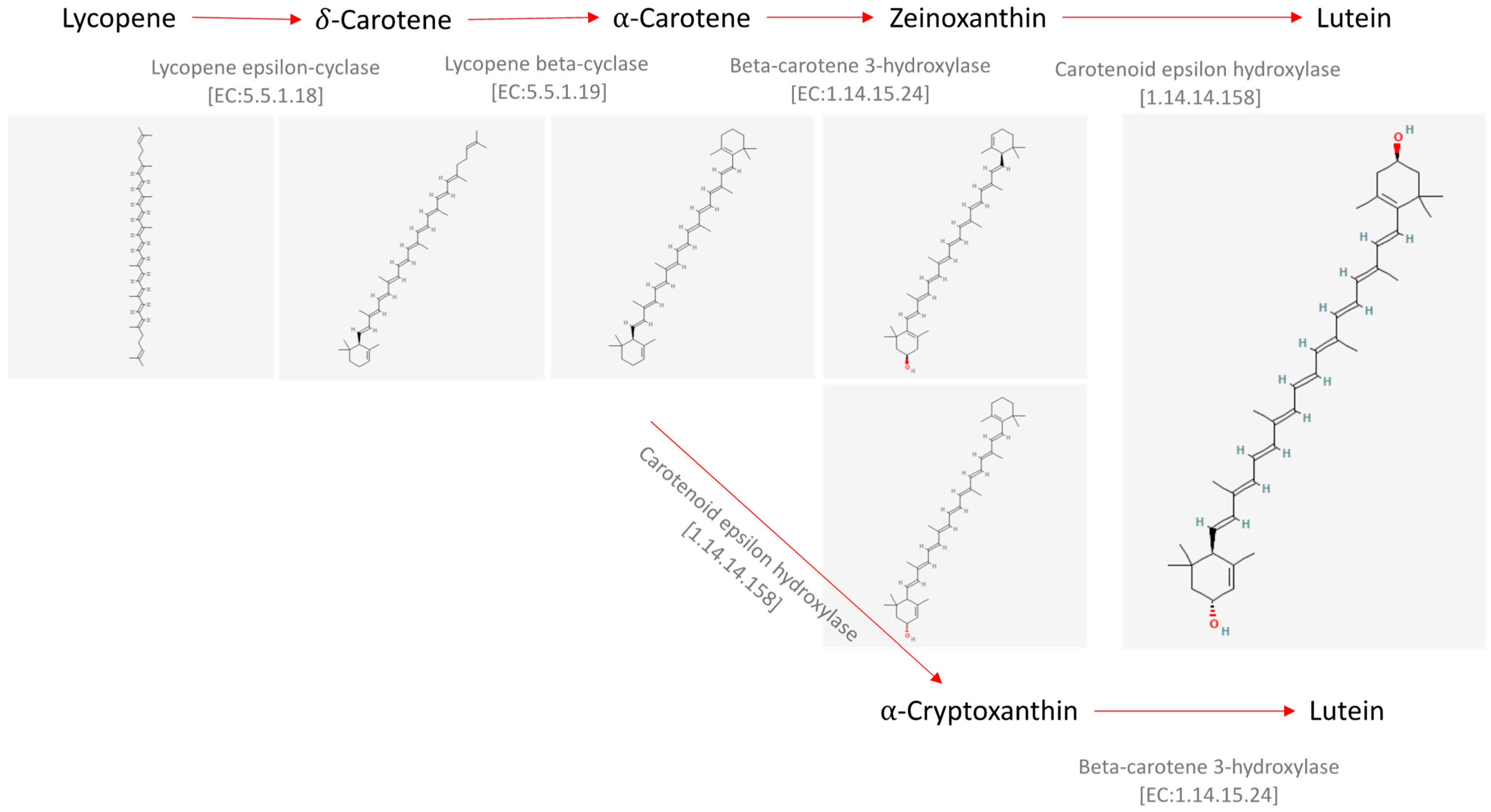
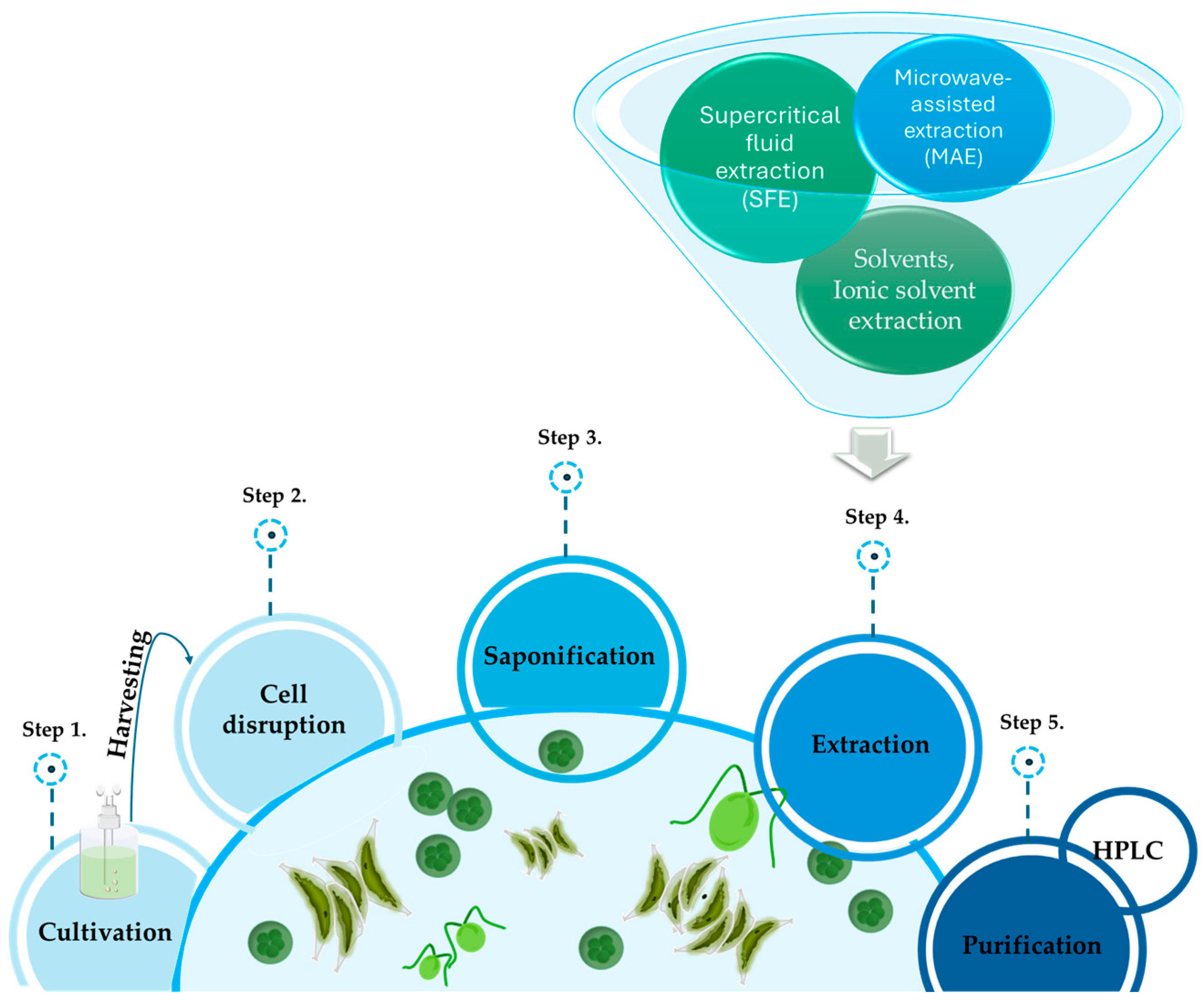
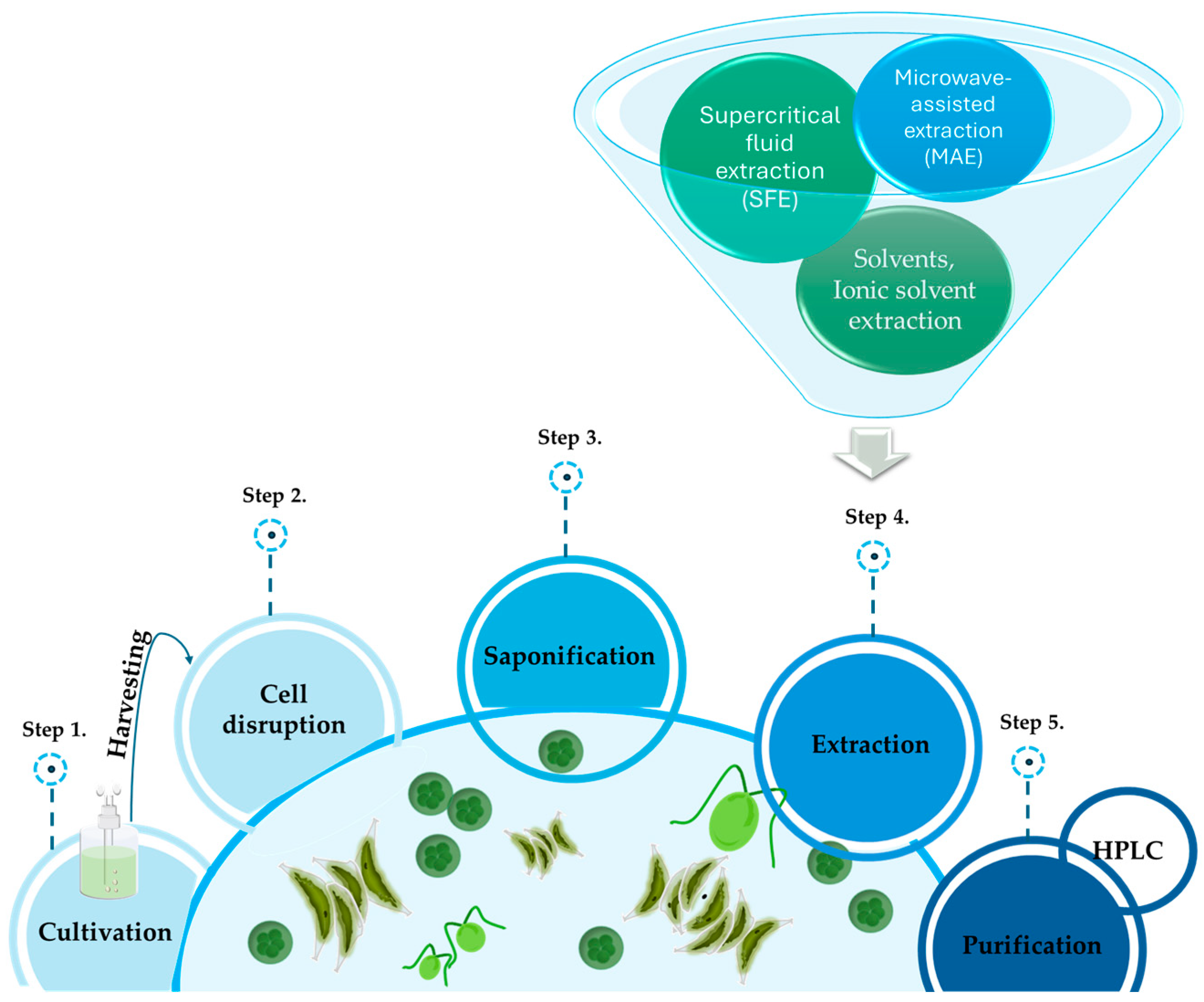
2. Extraction Procedures
2.1. Solvent Extraction
2.2. Supercritical Fluid Extraction (SFE)
2.3. Ionic Solvent Extraction
2.4. Microwave-Assisted Extraction (MAE)
2.5. Ultrasonic Extraction
2.6. Purification of Lutein
3. Lutein: A Versatile Antioxidant in Human Health
3.1. Lutein and Zeaxanthin: Guardians of Vision Health
3.2. Protection of Skin
3.3. Neurodegenerative Diseases
3.4. Lutein: A Potential Agent against Inflammation and Cancer
3.5. Diverse Protective Effects across Various Diseases

{kind=link}
{kind=link}
| Types of Cancer | Substance | Source | Model | Concentration | Mechanism of Action | Reference |
|---|---|---|---|---|---|---|
| Breast cancer | Lutein | Spinacia oleracea | In vitro (cells) MCF-7 and MDA-MB-231 cells | 1 and 5 µM for MCF-7 10 and 20 μM for MDA-MB-231 for 24 h | Inhibition of cell viability in a concentration-dependent manner Increase in cell death with higher concentrations of lutein Suppression of reactive oxygen species (ROS) levels Downregulation of Nrf-2 protein expression Decrease in expression of SOD-2, HO-2 proteins, and XBP-1 Inhibition of Akt phosphorylation and NF-κB expression Enhancement of caspase-3 activity, leading to apoptosis | [117] |
| Lutein | Strobilanthes crispus | In vivo (mouse) 4T1 breast cancer cell line | 50 mg/kg/day for 30 days | Activation of retinoid X receptor alpha (RXRa) by lutein Inhibition of breast cancer growth | [118] | |
| Lutein | - | In vitro MCF-7, MDA-MB-468 cells | 2.0 µM for 48 h | Inhibition of tumor cell growth, induction of cell cycle arrest, and cell death Increase in intracellular reactive oxygen species (ROS) levels Enhancement of the BAX/Bcl-2 ratio Activation of p53 signaling and upregulation of HSP60 | [119] | |
| Non-small-cell lung cancer (NSCLC) | Lutein | - | In vitro A549 cells | 25 and 50 μM (LC50 = 50 μM) | Inhibition of cell invasion and migration properties Suppression of PI3K/AKT signaling pathway Inhibition of intrinsic anti-apoptotic signaling molecules Bcl2 and Bcl-XL, leading to apoptosis Increase in expression of pro-apoptotic protein BAX, activating caspase 3 and inducing apoptosis | [120] |
| Lutein | - | In vitro A549 cells | 10, 30, 50 μM | Decrease in BCL2/BAX protein ratio, promoting apoptosis Reduction in gene expression of CAS3, SOX2, NANOG, SLCA11, and ABCB1 Increase in gene expression of CD44 and CD133 | [121] | |
| Colorectal Cancer | Lutein | Marigold flowers (Tagetes erecta) | In vivo | Chemoprotective effect against colon cancer Modulation of proliferative activity of K-ras, PKB, and β-catenin proteins | [122] | |
| Extract of microalgae Pressurized liquid extraction (EtOH) | Microalgae Neochloris oleoabundans | In vitro HT-29, SW480 HGUE-C-1 cells | 27, 83 or 250 μg mL−1 for 24 h | Different extracts exhibit dose-dependent anti-proliferative activities on cells | [130] | |
| Gastric Cancer | Lutein | Cayman Chemical n.10010811 | In vitro AGS, MKN-74, MKN-1, and SNU-668 cells | 5, 10, and 20 µM for 24 h | Induction of a dose-dependent decrease in cell viability Increase in ROS levels and induction of NADPH oxidase activation, promoting translocation of p47 phox subunit to the cell membrane Enhancement of NF-κB activation and induction of apoptotic indices, including Bax, caspase-3 cleavage, and DNA fragmentation Reduction in Bcl-2 levels | [123] |
| Lymphoid Leukemia | Lutein | Daucus carota L. | In vitro CCRF-CEM, Jurkat, MOLT-3 cells | 0.5 µM to100 µM for 24 and 48 h | No effect on apoptosis or cell proliferation | [124] |
| Melanoma | Lutein | - | In vitro A375 cells | 0.1, 0.3, 1, 3 and 10 µM of lutein for 24 h | No alteration in melanoma cell viability or membrane integrity by lutein | [125] |
| Prostate Cancer | Lutein | Marigold | In vitro PC-3 cells | 10 µM for 18 h | Alteration of expression of biomarker genes associated with growth and apoptosis by lutein | [126] |
| Hepatocellular carcinoma | Lutein | Marigold (Tagetes erecta L.) | In vivo | - | Reduction in levels of alanine transaminase, aspartate transaminase, and alkaline phosphatase Increase in levels of glutathione Reduction in activity of c-glutamyl transpeptidase | [127] |
| Lung cancer Cervical cancer Breast cancer Hepatocellular carcinoma Cholangiocarcinoma | Extracts of Chlorella (Gallic acid and lutein) (EtOH) | Chlorella sp. | In vitro A549, Hela, MCF7, Huh7, CCA, KKU213A cells | 400 μg mL−1 | Inhibition of growth in all types of tumor cells considered Reduction in procaspase-3, -8, and -9 levels, increase in caspase activity, and decrease in anti-apoptosis protein Bcl-2, leading to cell death Reduction in levels of phosphorylated-AKT and phosphorylated-mTOR proteins, inhibiting AKT/mTOR survival signaling | [129] |
| Breast cancer Colorectal cancer Prostate cancer Skin melanoma | Extract of Granulocystopsis (MeOH) | Granulocystopsis sp. | In vitro HTB-22, HTB-38, HTB-81 and HTB-72 cells | IC50 for 48 h: 16.70 µg/mL 17.20 µg/mL 13.74 µg/mL 17.44 µg/mL | Differential reactions of cell lines to the extract, but viability decreases in all depending on treatment duration Favorable induction of apoptosis in skin melanoma and prostate cancer cell lines Increase in caspase 3 and 7 activity in all treated tumor cells | [131] |
| Head and neck cancer (HNC): Oral and pharyngeal cancer Laryngeal cancer | Carotenoids: bcryptoxanthin, lycopene, and lutein plus zeaxanthin | Fruit and vegetables | Epidemiological studies | - | Carotenoids exhibit antioxidant, antimutagenic, and immunoregulatory actions Inverse association observed between carotenoid intake and risk of head and neck cancer (HNC) | [137] |
| Bladder cancer (BC) | Carotenoidis: lutein, αcarotene, βcarotene, lycopene, βcryptoxanthin, and zeaxanthin | Fruit and vegetables | Epidemiological studies | - | Inverse association between total carotenoid intake and risk of BC | [138] |
| Renal cell carcinoma (RCC) | Carotenoids: αcarotene, βcarotene, lutein, zeaxanthin, and lycopene | Fruit and vegetables | Epidemiological studies | - | Inverse associations between total carotenoid intake and risk of RCC | [139] |
4. Enhancement of Lutein Production in Microalgae
4.1. Metabolism
4.2. Light
4.3. Nitrogen
4.4. Temperature
4.5. Organic Carbon Source
4.6. Inorganic Carbon Source
4.7. Abiotic Stress
4.8. Growth Strategy
| Microorganism | Culture Conditions | Metabolic Mode | Enhancing Condition Applied | Lutein Content | Comment | Ref. |
|---|---|---|---|---|---|---|
| Tetraselmis sp. CTP4 | Batch cultivation in 5-L reactors with modified algal medium at 100 μmol m−2 s−1 | Photoautotrophic | Temperature | 2.15 ± 0.25 mg g−1 | Lutein content demonstrates an increase with rising temperature, reaching optimal levels at 30 °C | [165] |
| Tetraselmis sp. CTP4 | First step: batch cultivation in 5 L reactors with modified algal medium | Photoautotrophic | Combination of several strategies | 3.17 ± 0.18 mg g−1 | Effective enhancement of lutein content achieved through a two-stage cultivation process involving nitrogen depletion, controlled temperature, and light optimization | [165] |
| Chlorella protothecoides | Batch cultivation in 3.7 L fermenter with modified basal medium | Heterotrophic | Glucose addition | 4.44 mg g−1 | Optimal glucose concentrations were found to be 10 g L−1 and 40 g L−1 | [172] |
| Chlorella protothecoides | Batch cultivation in 3.7 L fermenter with modified basal medium with 40 g L−1 of glucose | Heterotrophic | Nitrogen source | 4.58 mg g−1 | Urea identified as the most effective nitrogen source for enhancing lutein content | [21] |
| Chlorella protothecoides | Batch cultivation in modified basal medium in Erlenmeyer flasks (250-mL) | Heterotrophic | Oxidative stress | 1.98 mg g−1 | Increased lutein accumulation observed in the presence of 0.01 mmol L-1 H2O2 and 0.5 mmol L−1 NaClO | [174] |
| Chlamydomonas acidophila | Batch cultivation in 1 L reactor at 25 °C | Mixotrophic | Source of carbon | 10 mg g−1 | In the presence of CO2, starch, urea, or glucose were the best carbon source | [171] |
| Desmodesmus sp. F51 | Batch cultivation in photobioreactor at 150 μmol m−2 s−1, temperature at 35 °C | Photoautotrophic | Nitrogen | 4.97 ± 0.05 mg g−1 | Optimal lutein accumulation achieved at 13.2 mM of N-nitrate | [159] |
| Desmodesmus sp. F51 | Batch cultivation in photobioreactor (PBR) at 150 μmol m−2 s−1, temperature at 35 °C; nitrate concentration, 8.8 mM | Photoautotrophic | Light intensity | 4.69 ± 0.08 mg g−1 | Increasing light intensity leads to a decrease in lutein content | [159] |
| Desmodesmus sp. F51 | Fed-batch photobioreactor at 600 μmol m−2 s−1 at 35 °C temperature, initial nitrate concentration, 8.8 mM | Photoautotrophic | Growth strategy | 5.05 ± 0.20 mg g−1 | Increased lutein content observed in fed-batch cultivation with a 2.2 mM nitrate feeding concentration | [159] |
| Chlorella protothecoides | Fed-batch growth; batch condition with 40 g L−1 of glucose; fed-batch condition limited in nitrogen | Heterotrophic | Combination of several strategies | 5.35 mg g−1 | Effective enhancement of lutein content achieved through a three-step cultivation process involving growth strategy, nitrogen depletion, and temperature control | [169] |
| S. obliquus FSP-3 | Batch growth in photobioreactor, 24 mM Ca(NO3)2 | Photoautotrophic | Nitrogen concentration | 4.95 ± 0.22 mg g−1 | Lutein content shows a positive correlation with rising nitrogen concentration | [163] |
| Chlorella minutissima MCC-27 | Batch cultivation in 2 L airlift photobioreactor with BBM | Photoautotrophic | Light supply | 8.24 ± 0.12 mg g−1 | Light supply strategy has a role on lutein accumulation | [162] |
| Desmodesmus sp. F51 | Batch cultivation in 1 L glass photobioreactor with modified Bristol’s medium at 150 μmol m−2 s−1 | Photoautotrophic | Carbon and nitrogen sources | 5.56 mg g−1 | Optimal lutein accumulation achieved at 150 mg L−1 of N-ammonium | [166] |
| Chlorella sorokiniana Mb-1 | Batch cultivation in 1 L glass photobioreactor with modified Bristol’s medium at 150 μmol m−2 s−1 with 1.83 g L−1 of nitrate | Mixotrophic | Organic carbon concentration | 4.6 mg g−1 | Increased lutein content observed at the lowest tested concentrations of acetate (4.88 g L−1) | [168] |
| Chlamydomonas sp. JSC4 | Batch cultivation in photobioreactor with modified Bold 3 N medium, sea salt, and initial nitrate-N concentrations adjusted to 2% and 1000 mg L−1 | Photoautotrophic | Light quality | 3.10 mg g−1 | Blue light increases lutein content | [157] |
| Chlamydomonas sp. JSC4 | Batch cultivation in photobioreactor with modified Bold 3 N medium/two-stage cultivation | Photoautotrophic | Combination of several strategies | 4.24 ± 0.22 mg g−1 | Efficient enhancement of lutein content achieved through two-stage cultivation involving control of light quality and temperature | [157] |
| Chlorella sorokiniana C16 | Batch cultivation on Tris acetate phosphate (TAP) media | Mixotrophic | Temperature | 17.4 mg g−1 | Lutein content exhibits a positive trend, reaching its peak with increasing temperature up to 32 °C | [170] |
| Chlorella sorokiniana FZU60 | Batch cultivation in BG11 medium at 33 °C; light intensity, 150 µmol m−2 s−1, initial NaNO3 concentration of 1 g L−1 | Mixotrophic | Organic carbon concentration | 9.65 ± 0.25 mg g−1 | Optimal lutein accumulation achieved at 1 g L−1 of sodium acetate | [167] |
| Chlorella sorokiniana FZU60 | Batch cultivation in BG11 medium; light intensity, 150 µmol m−2 s−1, 0.75 g L−1 sodium nitrate, and 1 g L−1 sodium acetate | Mixotrophic | Temperature | 9.81 ± 0.49 mg g−1 | Peak lutein content achieved at an optimal temperature of 33 °C | [156] |
| Chlorella sorokiniana Kh12 | Batch cultivation on Tris acetate phosphate (TAP) media | Mixotrophic | Temperature | 17.25 mg g−1 | Peak lutein content achieved at an optimal temperature of 30 °C | [160] |
| Scenedesmus almeriensis | Batch cultivation in vertical bubble column photobioreactor in modified Mann & Myers medium | Autotrophic | CO2 concentration | 5.71 mg g−1 | Optimal lutein accumulation observed at the highest CO2 concentration tested (3% v/v) | [173] |
| Parachlorella sp. JD-076 | Batch cultivation in tubular-type photobioreactor in BG-11 medium at 35 °C | Autotrophic | Light intensity | 11.8 mg g−1 | Lutein content shows a positive correlation with rising light intensities | [155] |
| Coccomyxa onubensis | Batch cultivation in 2L Erlenmeyer flasks 140 µmol m−2 s−1 at 26 °C | Autotrophic | Salt | 7.80 mg g−1 | Enhanced lutein content observed under UVA + PAR illumination | [175] |
| Coccomyxa onubensis | Batch cultivation in 2L Erlenmeyer flasks 140 µmol m−2 s−1 at 26 °C | Autotrophic | UVA light | 7.07 mg g−1 | Lutein content demonstrated a positive correlation with rising salinity levels | [175] |
| Chlorella sorokiniana MB-1-M12 | Batch cultivation in 5L fermenter with BG-11 medium at 27 ± 1 °C | Heterotrophic | Two-stage process | 5.88 mg g−1 | The described two-stage process integrating fed-batch and semi-batch modes induced lutein accumulation | [164] |
| Scenedesmus obliquus CWL-1 | Batch cultivation in 7 L stirred tank reactor with 4.5 g L−1 of calcium nitrate | Mixotrophic | Combination of several strategies | 2.45 mg g−1 | Photoperiod and wavelength adjustment effective in inducing lutein | [158] |
5. Pilot-Scale Systems for Lutein Production
6. Genetic Engineering Production
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buscemi, S.; Corleo, D.; Di Pace, F.; Petroni, M.; Satriano, A.; Marchesini, G. The Effect of Lutein on Eye and Extra-Eye Health. Nutrients 2018, 10, 1321. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Chemical Diversity and Defence Metabolism: How Plants Cope with Pathogens and Ozone Pollution. Int. J. Mol. Sci. 2009, 10, 3371–3399. [Google Scholar] [CrossRef] [PubMed]
- Kalariya, N.M.; Ramana, K.V.; VanKuijk, F.J.G.M. Focus on Molecules: Lutein. Exp. Eye Res. 2012, 102, 107–108. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 446925, Lycopene. 2024. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/446925#section=2D-Structure (accessed on 26 February 2024).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5281230, Delta-Carotene. 2024. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5281230#section=2D-Structure (accessed on 26 February 2024).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 6419725, Alpha-Carotene. 2024. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/6419725#section=2D-Structure (accessed on 26 February 2024).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5281234, Zeinoxanthin. 2024. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5281234#section=2D-Structure (accessed on 26 February 2024).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 23724629, Alpha-Cryptoxanthin. 2024. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/23724629#section=2D-Structure (accessed on 26 February 2024).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5281243, Lutein. 2024. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5281243#section=2D-Structure (accessed on 26 February 2024).
- Nagy, V.; Agocs, A.; Deli, J. In Vitro and In Vivo Transformations of Lutein. Mini. Rev. Org. Chem. 2009, 6, 211–219. [Google Scholar] [CrossRef]
- Saha, S.K.; Ermis, H.; Murray, P. Marine Microalgae for Potential Lutein Production. Appl. Sci. 2020, 10, 6457. [Google Scholar] [CrossRef]
- Lin, J.H.; Lee, D.J.; Chang, J.S. Lutein Production from Biomass: Marigold Flowers versus Microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Wang, Y.; Li, S.; Nagarajan, D.; Varjani, S.; Lee, D.J.; Chang, J.S. Recent Advances in Lutein Production from Microalgae. Renew. Sustain. Energy Rev. 2022, 153, 111795. [Google Scholar] [CrossRef]
- Sun, Z.; Li, T.; Zhou, Z.; Jiang, Y. Microalgae as a Source of Lutein: Chemistry, Biosynthesis, and Carotenogenesis; Springer: Berlin/Heidelberg, Germany, 2015; pp. 37–58. [Google Scholar]
- Ochoa Becerra, M.; Mojica Contreras, L.; Hsieh Lo, M.; Mateos Díaz, J.; Castillo Herrera, G. Lutein as a Functional Food Ingredient: Stability and Bioavailability. J. Funct. Foods 2020, 66, 103771. [Google Scholar] [CrossRef]
- Sulich, A.; Hamułka, J.; Nogal, D. Dietary Sources of Lutein in Adults Suffering Eye Disease (AMD/Cataracts). Rocz. Panstw. Zakl. Hig. 2015, 66, 55–60. [Google Scholar]
- Raposo, M.; de Morais, A.; de Morais, R. Carotenoids from Marine Microalgae: A Valuable Natural Source for the Prevention of Chronic Diseases. Mar. Drugs 2015, 13, 5128–5155. [Google Scholar] [CrossRef]
- Li, L.H.; Lee, J.C.-Y.; Leung, H.H.; Lam, W.C.; Fu, Z.; Lo, A.C.Y. Lutein Supplementation for Eye Diseases. Nutrients 2020, 12, 1721. [Google Scholar] [CrossRef]
- Muhammad, G.; Butler, T.O.; Chen, B.; Lv, Y.; Xiong, W.; Zhao, X.; Solovchenko, A.E.; Zhao, A.; Mofijur, M.; Xu, J.; et al. Sustainable Production of Lutein—An Underexplored Commercially Relevant Pigment from Microalgae. Biomass Convers. Biorefin. 2022, 1–22. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Rodríguez, H.; Moreno, J.; Vargas, M.Á.; Rivas, J.; Guerrero, M.G. Accumulation of Astaxanthin and Lutein in Chlorella zofingiensis (Chlorophyta). Appl. Microbiol. Biotechnol. 2004, 64, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.M.; Zhang, X.W.; Chen, F. Heterotrophic Production of Biomass and Lutein by Chlorella protothecoides on Various Nitrogen Sources. Enzyme Microb. Technol. 2000, 27, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J. Carotenoid Content of Chlorophycean Microalgae: Factors Determining Lutein Accumulation in Muriellopsis sp. (Chlorophyta). J. Biotechnol. 2000, 76, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Rodríguez, H.; Moreno, J.; Vargas, M.Á.; Rivas, J.; Guerrero, M.G. Lutein Production by Muriellopsis sp. in an Outdoor Tubular Photobioreactor. J. Biotechnol. 2001, 85, 289–295. [Google Scholar] [CrossRef]
- Sánchez, J.F.; Fernández, J.M.; Acién, F.G.; Rueda, A.; Pérez-Parra, J.; Molina, E. Influence of Culture Conditions on the Productivity and Lutein Content of the New Strain Scenedesmus almeriensis. Process Biochem. 2008, 43, 398–405. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, Y.; Yi, L.; Liu, J.; Yang, S.; Liu, B.; Chen, F.; Sun, H. Lutein Production from Microalgae: A Review. Bioresour. Technol. 2023, 376, 128875. [Google Scholar] [CrossRef]
- Available online: https://www.marketsandmarkets.com/market-reports/lutein-market-69753879.html (accessed on 26 February 2024).
- Fatima, I.; Munir, M.; Qureshi, R.; Hanif, U.; Gulzar, N.; Sheikh, A.A. Advanced Methods of Algal Pigments Extraction: A Review. Crit. Rev. Food Sci. Nutr. 2023, 1–18. [Google Scholar] [CrossRef]
- Poojary, M.M.; Barba, F.J.; Aliakbarian, B.; Donsì, F.; Pataro, G.; Dias, D.A.; Juliano, P. Innovative Alternative Technologies to Extract Carotenoids from Microalgae and Seaweeds. Mar. Drugs 2016, 14, 214. [Google Scholar] [CrossRef]
- Patel, A.K.; Vadrale, A.P.; Tseng, Y.-S.; Chen, C.-W.; Dong, C.-D.; Singhania, R.R. Bioprospecting of Marine Microalgae from Kaohsiung Seacoast for Lutein and Lipid Production. Bioresour. Technol. 2022, 351, 126928. [Google Scholar] [CrossRef] [PubMed]
- Larsen, E.; Christensen, L.P. Simple Saponification Method for the Quantitative Determination of Carotenoids in Green Vegetables. J. Agric. Food Chem. 2005, 53, 6598–6602. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-B.; Jiang, Y.; Chen, F. Isolation and Purification of Lutein from the Microalga Chlorella vulgaris by Extraction after Saponification. J. Agric. Food Chem. 2002, 50, 1070–1072. [Google Scholar] [CrossRef]
- Kumar, R.; Yu, W.; Jiang, C.; Shi, C.; Zhao, Y. Improvement of The Isolation and Purification Of Lutein From Marigold Flower (Tagetes erecta L.) and Its Antioxidant Activity. J. Food Process Eng. 2010, 33, 1065–1078. [Google Scholar] [CrossRef]
- Chan, M.-C.; Ho, S.-H.; Lee, D.-J.; Chen, C.-Y.; Huang, C.-C.; Chang, J.-S. Characterization, Extraction and Purification of Lutein Produced by an Indigenous Microalga Scenedesmus obliquus CNW-N. Biochem. Eng. J. 2013, 78, 24–31. [Google Scholar] [CrossRef]
- Mendes-Pinto, M.M.; Raposo, M.F.J.; Bowen, J.; Young, A.J.; Morais, R. Evaluation of Different Cell Disruption Processes on Encysted Cells of Haematococcus pluvialis: Effects on Astaxanthin Recovery and Implications for Bio-Availability. J. Appl. Phycol. 2001, 13, 19–24. [Google Scholar] [CrossRef]
- Jeevan Kumar, S.P.; Vijay Kumar, G.; Dash, A.; Scholz, P.; Banerjee, R. Sustainable Green Solvents and Techniques for Lipid Extraction from Microalgae: A Review. Algal Res. 2017, 21, 138–147. [Google Scholar] [CrossRef]
- Maciassanchez, M.; Mantell, C.; Rodriguez, M.; Martinezdelaossa, E.; Lubian, L.; Montero, O. Comparison of Supercritical Fluid and Ultrasound-Assisted Extraction of Carotenoids and Chlorophyll a from Dunaliella salina. Talanta 2009, 77, 948–952. [Google Scholar] [CrossRef]
- Kanda, H.; Wahyudiono; Machmudah, S.; Goto, M. Direct Extraction of Lutein from Wet Macroalgae by Liquefied Dimethyl Ether without Any Pretreatment. ACS Omega 2020, 5, 24005–24010. [Google Scholar] [CrossRef]
- Sosa-Hernández, J.; Romero-Castillo, K.; Parra-Arroyo, L.; Aguilar-Aguila-Isaías, M.; García-Reyes, I.; Ahmed, I.; Parra-Saldivar, R.; Bilal, M.; Iqbal, H. Mexican Microalgae Biodiversity and State-Of-The-Art Extraction Strategies to Meet Sustainable Circular Economy Challenges: High-Value Compounds and Their Applied Perspectives. Mar. Drugs 2019, 17, 174. [Google Scholar] [CrossRef]
- Molino, A.; Mehariya, S.; Di Sanzo, G.; Larocca, V.; Martino, M.; Leone, G.P.; Marino, T.; Chianese, S.; Balducchi, R.; Musmarra, D. Recent Developments in Supercritical Fluid Extraction of Bioactive Compounds from Microalgae: Role of Key Parameters, Technological Achievements and Challenges. J. CO2 Util. 2020, 36, 196–209. [Google Scholar] [CrossRef]
- Miguel, F.; Martín, A.; Mattea, F.; Cocero, M.J. Precipitation of Lutein and Co-Precipitation of Lutein and Poly-Lactic Acid with the Supercritical Anti-Solvent Process. Chem. Eng. Process. Process Intensif. 2008, 47, 1594–1602. [Google Scholar] [CrossRef]
- Yen, H.-W.; Chiang, W.-C.; Sun, C.-H. Supercritical Fluid Extraction of Lutein from Scenedesmus Cultured in an Autotrophical Photobioreactor. J. Taiwan Inst. Chem. Eng. 2012, 43, 53–57. [Google Scholar] [CrossRef]
- Ruen-ngam, D.; Shotipruk, A.; Pavasant, P.; Machmudah, S.; Goto, M. Selective Extraction of Lutein from Alcohol Treated Chlorella vulgaris by Supercritical CO2. Chem. Eng. Technol. 2012, 35, 255–260. [Google Scholar] [CrossRef]
- Gilbert-López, B.; Mendiola, J.A.; van den Broek, L.A.M.; Houweling-Tan, B.; Sijtsma, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Green Compressed Fluid Technologies for Downstream Processing of Scenedesmus obliquus in a Biorefinery Approach. Algal Res. 2017, 24, 111–121. [Google Scholar] [CrossRef]
- Ranjith Kumar, R.; Hanumantha Rao, P.; Arumugam, M. Lipid Extraction Methods from Microalgae: A Comprehensive Review. Front. Energy Res. 2015, 2, 61. [Google Scholar] [CrossRef]
- Pan, M.; Su, Y.; Zhu, X.; Pan, G.; Zhang, Y.; Angelidaki, I. Bioelectrochemically Assisted Sustainable Conversion of Industrial Organic Wastewater and Clean Production of Microalgal Protein. Resour. Conserv. Recycl. 2021, 168, 105441. [Google Scholar] [CrossRef]
- Khoo, K.S.; Chong, Y.M.; Chang, W.S.; Yap, J.M.; Foo, S.C.; Khoiroh, I.; Lau, P.L.; Chew, K.W.; Ooi, C.W.; Show, P.L. Permeabilization of Chlorella sorokiniana and Extraction of Lutein by Distillable CO2-Based Alkyl Carbamate Ionic Liquids. Sep. Purif. Technol. 2021, 256, 117471. [Google Scholar] [CrossRef]
- Hikmawanti, N.P.E.; Ramadon, D.; Jantan, I.; Mun’im, A. Natural Deep Eutectic Solvents (NADES): Phytochemical Extraction Performance Enhancer for Pharmaceutical and Nutraceutical Product Development. Plants 2021, 10, 2091. [Google Scholar] [CrossRef]
- Paliwal, C.; Rehmanji, M.; Shaikh, K.M.; Zafar, S.U.; Jutur, P.P. Green Extraction Processing of Lutein from Chlorella saccharophila in Water-Based Ionic Liquids as a Sustainable Innovation in Algal Biorefineries. Algal Res. 2022, 66, 102809. [Google Scholar] [CrossRef]
- Fan, C.; Liu, Y.; Shan, Y.; Cao, X. A Priori Design of New Natural Deep Eutectic Solvent for Lutein Recovery from Microalgae. Food Chem. 2022, 376, 131930. [Google Scholar] [CrossRef] [PubMed]
- Nithya, S.; Krishnan, R.R.; Rao, N.R.; Naik, K.; Praveen, N.; Vasantha, V.L. Microwave-Assisted Extraction of Phytochemicals. In Drug Discovery and Design Using Natural Products; Springer Nature Switzerland: Cham, Switzerland, 2023; pp. 209–238. [Google Scholar]
- Ekezie, F.-G.C.; Sun, D.-W.; Cheng, J.-H. Acceleration of Microwave-Assisted Extraction Processes of Food Components by Integrating Technologies and Applying Emerging Solvents: A Review of Latest Developments. Trends Food Sci. Technol. 2017, 67, 160–172. [Google Scholar] [CrossRef]
- Mary Leema, J.T.; Persia Jothy, T.; Dharani, G. Rapid Green Microwave Assisted Extraction of Lutein from Chlorella sorokiniana (NIOT-2)—Process Optimization. Food Chem. 2022, 372, 131151. [Google Scholar] [CrossRef]
- Low, K.L.; Idris, A.; Mohd Yusof, N. Novel Protocol Optimized for Microalgae Lutein Used as Food Additives. Food Chem. 2020, 307, 125631. [Google Scholar] [CrossRef]
- Gayathri, S.; Rajasree Radhika, S.R.; Suman, T.Y.; Aranganathan, L. Ultrasound-Assisted Microextraction of β, ε-Carotene-3, 3′-Diol (Lutein) from Marine Microalgae Chlorella salina: Effect of Different Extraction Parameters. Biomass Convers. Biorefinery 2018, 8, 791–797. [Google Scholar] [CrossRef]
- Available online: https://www.nobelprize.org/prizes/chemistry/1915/willstatter/biographical/ (accessed on 27 February 2024).
- Ingkasupart, P.; Manochai, B.; Song, W.T.; Hong, J.H. Antioxidant Activities and Lutein Content of 11 Marigold Cultivars (Tagetes spp.) Grown in Thailand. Food Sci. Technol. 2015, 35, 380–385. [Google Scholar] [CrossRef]
- Şahin, S.; Aybastıer, Ö.; Dawbaa, S.; Karkar, B.; Çakmak, T. Study of the Ability of Lutein and Neoxanthin as Standards and in the Extract of Chlamydomonas reinhardtii to Prevent Oxidatively Induced DNA Base Damage Using Ultrasensitive GC–MS/MS Analysis. Chromatographia 2020, 83, 919–926. [Google Scholar] [CrossRef]
- Kim, E.A.; Kang, N.; Heo, S.Y.; Oh, J.Y.; Lee, S.H.; Cha, S.H.; Kim, W.K.; Heo, S.J. Antioxidant, Antiviral, and Anti-Inflammatory Activities of Lutein-Enriched Extract of Tetraselmis Species. Mar. Drugs 2023, 21, 369. [Google Scholar] [CrossRef]
- Serpeloni, J.M.; de Cólus, I.M.S.; de Oliveira, F.S.; Aissa, A.F.; Mercadante, A.Z.; Bianchi, M.L.P.; Antunes, L.M.G. Diet Carotenoid Lutein Modulates the Expression of Genes Related to Oxygen Transporters and Decreases DNA Damage and Oxidative Stress in Mice. Food Chem. Toxicol. 2014, 70, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Qian, S.; Na, X.; Zhao, A. Association between Dietary and Supplemental Antioxidants Intake and Lung Cancer Risk: Evidence from a Cancer Screening Trial. Antioxidants 2023, 12, 338. [Google Scholar] [CrossRef]
- Jȩdrzejczak-Pospiech, K.; Błaszczyk, J. The Effect of Lutein on the Total Antioxidant Status in Human Blood. Klin. Oczna 2017, 2017, 220–223. [Google Scholar]
- Lorenzoni, F.; Giampietri, M.; Ferri, G.; Lunardi, S.; Madrigali, V.; Battini, L.; Boldrini, A.; Ghirri, P. Lutein Administration to Pregnant Women with Gestational Diabetes Mellitus Is Associated to a Decrease of Oxidative Stress in Newborns. Gynecol. Endocrinol. 2013, 29, 901–903. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Giannantonio, C.; Romagnoli, C.; Vento, G.; Gervasoni, J.; Persichilli, S.; Zuppi, C.; Cota, F. Effects of Lutein Supplementation on Biological Antioxidant Status in Preterm Infants: A Randomized Clinical Trial. J. Matern. Neonatal Med. 2013, 26, 1311–1315. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Tei, M.; Longini, M.; Santacroce, A.; Turrisi, G.; Proietti, F.; Felici, C.; Picardi, A.; Bazzini, F.; Vasarri, P.; et al. Lipid and Protein Oxidation in Newborn Infants after Lutein Administration. Oxid. Med. Cell. Longev. 2014, 2014, 781454. [Google Scholar] [CrossRef] [PubMed]
- Alves-Rodrigues, A.; Shao, A. The Science behind Lutein. Toxicol. Lett. 2004, 150, 57–83. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.J.; Kim, H. Lutein as a Modulator of Oxidative Stress-Mediated Inflammatory Diseases. Antioxidants 2021, 10, 1448. [Google Scholar] [CrossRef]
- Boehm, F.; Edge, R.; Truscott, T.G. Anti- and pro-Oxidative Mechanisms Comparing the Macular Carotenoids Zeaxanthin and Lutein with Other Dietary Carotenoids—A Singlet Oxygen, Free-Radica I in Vitro and Ex Vivo Study. Photochem. Photobiol. Sci. 2020, 19, 1001–1009. [Google Scholar] [CrossRef]
- Subagio, A.; Morita, N. No Effect of Esterification with Fatty Acid on Antioxidant Activity of Lutein. Food Res. Int. 2001, 34, 315–320. [Google Scholar] [CrossRef]
- Yang, C.; Fischer, M.; Kirby, C.; Liu, R.; Zhu, H.; Zhang, H.; Chen, Y.; Sun, Y.; Zhang, L.; Tsao, R. Bioaccessibility, Cellular Uptake and Transport of Luteins and Assessment of Their Antioxidant Activities. Food Chem. 2018, 249, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.; Beck, M.; Elliott, J.; Etheve, S.; Roberts, R.; Schalch, W. Effects of Formulation on the Bioavailability of Lutein and Zeaxanthin: A Randomized, Double-Blind, Cross-over, Comparative, Single-Dose Study in Healthy Subjects. Eur. J. Nutr. 2013, 52, 1381–1391. [Google Scholar] [CrossRef]
- Nemli, E.; Capanoglu, E.; McClements, D.J.; Tomas, M. Use of Excipient Emulsions for Improving the Bioaccessibility of Antioxidants in Tomato Sauce. Food Chem. 2023, 424, 136395. [Google Scholar] [CrossRef]
- Pugliese, A.; O’Callaghan, Y.; Tundis, R.; Galvin, K.; Menichini, F.; O’Brien, N.; Loizzo, M.R. In Vitro Investigation of the Bioaccessibility of Carotenoids from Raw, Frozen and Boiled Red Chili Peppers (Capsicum annuum). Eur. J. Nutr. 2014, 53, 501–510. [Google Scholar] [CrossRef]
- Zeb, A.; Haq, A.; Murkovic, M. Effects of Microwave Cooking on Carotenoids, Phenolic Compounds and Antioxidant Activity of Cichorium intybus L. (Chicory) Leaves. Eur. Food Res. Technol. 2019, 245, 365–374. [Google Scholar] [CrossRef]
- Mashiane, P.; Shoko, T.; Manhivi, V.; Slabbert, R.; Sultanbawa, Y.; Sivakumar, D. A Comparison of Bioactive Metabolites, Antinutrients, and Bioactivities of African Pumpkin Leaves (Momordica balsamina L.) Cooked by Different Culinary Techniques. Molecules 2022, 27, 1901. [Google Scholar] [CrossRef] [PubMed]
- Pap, R.; Pandur, E.; Jánosa, G.; Sipos, K.; Fritz, F.R.; Nagy, T.; Agócs, A.; Deli, J. Protective Effects of 3′-Epilutein and 3′-Oxolutein against Glutamate-Induced Neuronal Damage. Int. J. Mol. Sci. 2023, 24, 12008. [Google Scholar] [CrossRef]
- Cui, N.; Yang, C.; Chen, X.; Zhang, J.; Zhang, L. Anti-Oxidative and Anti-Inflammatory Effects of Different Lutein Isomers in Caco-2 Cell Model. Food Biosci. 2023, 53, 102646. [Google Scholar] [CrossRef]
- Serra, A.T.; Silva, S.D.; Pleno de Gouveia, L.; Alexandre, A.M.R.C.; Pereira, C.V.; Pereira, A.B.; Partidário, A.C.; Silva, N.E.; Bohn, T.; Gonçalves, V.S.S.; et al. A Single Dose of Marine Chlorella vulgaris Increases Plasma Concentrations of Lutein, β-Carotene and Zeaxanthin in Healthy Male Volunteers. Antioxidants 2021, 10, 1164. [Google Scholar] [CrossRef]
- Ozawa, Y.; Sasaki, M. Lutein and Oxidative Stress-Mediated Retinal Neurodegeneration in Diabetes; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9780124058859. [Google Scholar]
- Murthy, R.K.; Ravi, K.; Balaiya, S.; Brar, V.S.; Chalam, K.V. Lutein Protects Retinal Pigment Epithelium from Cytotoxic Oxidative Stress. Cutan. Ocul. Toxicol. 2014, 33, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Kamoshita, M.; Toda, E.; Osada, H.; Narimatsu, T.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Lutein Acts via Multiple Antioxidant Pathways in the Photo-Stressed Retina. Sci. Rep. 2016, 6, 30226. [Google Scholar] [CrossRef] [PubMed]
- Leung, H.H.; Galano, J.M.; Crauste, C.; Durand, T.; Lee, J.C.Y. Combination of Lutein and Zeaxanthin, and DHA Regulated Polyunsaturated Fatty Acid Oxidation in H2O2-Stressed Retinal Cells. Neurochem. Res. 2020, 45, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, W.; Zhou, X.; Long, C.; Kuang, X.; Hu, J.; Tang, Y.; Liu, L.; He, J.; Huang, Z.; et al. Protective Effect of Lutein on ARPE-19 Cells upon H2O2-Induced G2/M Arrest. Mol. Med. Rep. 2017, 16, 2069–2074. [Google Scholar] [CrossRef]
- Chae, S.; Park, S.; Park, G. Lutein Protects Human Retinal Pigment Epithelial Cells from Oxidative Stress-induced Cellular Senescence. Mol. Med. Rep. 2018, 18, 5182–5190. [Google Scholar] [CrossRef]
- Aimjongjun, S.; Sutheerawattananonda, M.; Limpeanchob, N. Silk Lutein Extract and Its Combination with Vitamin E Reduce UVB-Mediated Oxidative Damage to Retinal Pigment Epithelial Cells. J. Photochem. Photobiol. B Biol. 2013, 124, 34–41. [Google Scholar] [CrossRef]
- Fernández-Robredo, P.; Sádaba, L.M.; Salinas-Alamán, A.; Recalde, S.; Rodríguez, J.A.; García-Layana, A. Effect of Lutein and Antioxidant Supplementation on VEGF Expression, MMP-2 Activity, and Ultrastructural Alterations in Apolipoprotein E-Deficient Mouse. Oxid. Med. Cell. Longev. 2013, 2013, 213505. [Google Scholar] [CrossRef] [PubMed]
- Padmanabha, S.; Vallikannan, B. Fatty Acids Modulate the Efficacy of Lutein in Cataract Prevention: Assessment of Oxidative and Inflammatory Parameters in Rats. Biochem. Biophys. Res. Commun. 2018, 500, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Gencoglu, H.; Akdemir, F.; Orhan, C.; Tuzcu, M.; Sahin, N.; Yilmaz, I.; Juturu, V. Lutein and Zeaxanthin Isomers May Attenuate Photo-Oxidative Retinal Damage via Modulation of G Protein-Coupled Receptors and Growth Factors in Rats. Biochem. Biophys. Res. Commun. 2019, 516, 163–170. [Google Scholar] [CrossRef]
- Erhan, E.; Salcan, I.; Bayram, R.; Suleyman, B.; Dilber, M.; Yazici, G.N.; Coban, T.A.; Altuner, D.; Suleyman, H. Protective Effect of Lutein against Acrolein-Induced Ototoxicity in Rats. Biomed. Pharmacother. 2021, 137, 111281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Liu, X.B.; Chen, X.M.; Kong, Q.H.; Liu, Y.S.; So, K.F.; Chen, J.S.; Xu, Y.; Mi, X.S.; Tang, S.B. Lutein Delays Photoreceptor Degeneration in a Mouse Model of Retinitis Pigmentosa. Neural Regen. Res. 2022, 17, 1596–1603. [Google Scholar] [CrossRef]
- Roldán-Fidalgo, A.; Martín Saldaña, S.; Trinidad, A.; Olmedilla-Alonso, B.; Rodríguez-Valiente, A.; García-Berrocal, J.R.; Ramírez-Camacho, R. In Vitro and in Vivo Effects of Lutein against Cisplatin-Induced Ototoxicity. Exp. Toxicol. Pathol. 2016, 68, 197–204. [Google Scholar] [CrossRef]
- Yu, M.; Yan, W.; Beight, C. Lutein and Zeaxanthin Isomers Protect against Light-Induced Retinopathy via Decreasing Oxidative and Endoplasmic Reticulum Stress in BALB/CJ Mice. Nutrients 2018, 10, 842. [Google Scholar] [CrossRef]
- Bartlett, H.E.; Eperjesi, F. Effect of Lutein and Antioxidant Dietary Supplementation on Contrast Sensitivity in Age-Related Macular Disease: A Randomized Controlled Trial. Eur. J. Clin. Nutr. 2007, 61, 1121–1127. [Google Scholar] [CrossRef]
- Berrow, E.J.; Bartlett, H.E.; Eperjesi, F.; Gibson, J.M. The Effects of a Lutein-Based Supplement on Objective and Subjective Measures of Retinal and Visual Function in Eyes with Age-Related Maculopathy—A Randomised Controlled Trial. Br. J. Nutr. 2013, 109, 2008–2014. [Google Scholar] [CrossRef]
- Nolan, J.M.; Loskutova, E.; Howard, A.; Mulcahy, R.; Moran, R.; Stack, J.; Bolger, M.; Coen, R.F.; Dennison, J.; Akuffo, K.O.; et al. The Impact of Supplemental Macular Carotenoids in Alzheimer’s Disease: A Randomized Clinical Trial. J. Alzheimer’s Dis. 2015, 44, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.-L.; Chiu, H.-F.; Chou, H.; Liao, H.-J.; Chen, S.-T.; Wong, Y.-C.; Shen, Y.-C.; Venkatakrishnan, K.; Wang, C.-K. Influence/Impact of Lutein Complex (Marigold Flower and Wolfberry) on Visual Function with Early Age-Related Macular Degeneration Subjects: A Randomized Clinical Trial. J. Funct. Foods 2016, 24, 122–130. [Google Scholar] [CrossRef]
- Obana, A.; Gohto, Y.; Nakazawa, R.; Moriyama, T.; Gellermann, W.; Bernstein, P.S. Effect of an Antioxidant Supplement Containing High Dose Lutein and Zeaxanthin on Macular Pigment and Skin Carotenoid Levels. Sci. Rep. 2020, 10, 10262. [Google Scholar] [CrossRef]
- Ma, L.; Liu, R.; Du, J.; Liu, T.; Wu, S.; Liu, X. Lutein, Zeaxanthin and Meso-Zeaxanthin Supplementation Associated with Macular Pigment Optical Density. Nutrients 2016, 8, 426. [Google Scholar] [CrossRef] [PubMed]
- Shinojima, A.; Sawa, M.; Sekiryu, T.; Oshima, Y.; Mori, R.; Hara, C.; Sugano, Y.; Kato, A.; Asato, H.; Yuzawa, M.; et al. A Multicenter Randomized Controlled Study of Antioxidant Supplementation with Lutein for Chronic Central Serous Chorioretinopathy. Ophthalmologica 2017, 237, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.; Galasso, C.; Orefice, I.; Nuzzo, G.; Luongo, E.; Cutignano, A.; Romano, G.; Brunet, C.; Fontana, A.; Esposito, F.; et al. The Green Microalga Tetraselmis suecica Reduces Oxidative Stress and Induces Repairing Mechanisms in Human Cells. Sci. Rep. 2017, 7, 41215. [Google Scholar] [CrossRef]
- Žmitek, K.; Žmitek, J.; Rogl Butina, M.; Hristov, H.; Pogačnik, T.; Pravst, I. Dietary Lutein Supplementation Protects against Ultraviolet-Radiation-Induced Erythema: Results of a Randomized Double-Blind Placebo-Controlled Study. J. Funct. Foods 2020, 75, 104265. [Google Scholar] [CrossRef]
- Liu, T.; Liu, W.; Zhao, J.; Meng, F.; Wang, H. Lutein Protects against β-Amyloid Peptide-Induced Oxidative Stress in Cerebrovascular Endothelial Cells through Modulation of Nrf-2 and NF-κb. Cell Biol. Toxicol. 2017, 33, 57–67. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, X.; Lian, F.; Yang, J.; Xu, X. Combination of Lutein and DHA Alleviate H2O2 Induced Cytotoxicity in PC12 Cells by Regulating the MAPK Pathway. J. Nutr. Sci. Vitaminol. 2021, 67, 234–242. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, H.; Liu, G.; Kang, J.; Wang, B.; Wang, J.; Li, J.; Wang, H. Lutein Attenuated Methylglyoxal-induced Oxidative Damage and Apoptosis in PC12 Cells via the PI3K Akt Signaling Pathway. J. Food Biochem. 2022, 46, e14382. [Google Scholar] [CrossRef]
- Pap, R.; Pandur, E.; Jánosa, G.; Sipos, K.; Agócs, A.; Deli, J. Lutein Exerts Antioxidant and Anti-Inflammatory Effects and Influences Iron Utilization of BV-2 Microglia. Antioxidants 2021, 10, 363. [Google Scholar] [CrossRef]
- Pap, R.; Pandur, E.; Jánosa, G.; Sipos, K.; Nagy, T.; Agócs, A.; Deli, J. Lutein Decreases Inflammation and Oxidative Stress and Prevents Iron Accumulation and Lipid Peroxidation at Glutamate-Induced Neurotoxicity. Antioxidants 2022, 11, 2269. [Google Scholar] [CrossRef]
- Fernandes, E.J.; Poetini, M.R.; Barrientos, M.S.; Bortolotto, V.C.; Araujo, S.M.; Santos Musachio, E.A.; De Carvalho, A.S.; Leimann, F.V.; Gonçalves, O.H.; Ramborger, B.P.; et al. Exposure to Lutein-Loaded Nanoparticles Attenuates Parkinson’s Model-Induced Damage in Drosophila melanogaster: Restoration of Dopaminergic and Cholinergic System and Oxidative Stress Indicators. Chem. Biol. Interact. 2021, 340, 109431. [Google Scholar] [CrossRef] [PubMed]
- Nazari, L.; Komaki, S.; Salehi, I.; Raoufi, S.; Golipoor, Z.; Kourosh-Arami, M.; Komaki, A. Investigation of the Protective Effects of Lutein on Memory and Learning Using Behavioral Methods in a Male Rat Model of Alzheimer’s Disease. J. Funct. Foods 2022, 99, 105319. [Google Scholar] [CrossRef]
- Fatani, A.J.; Parmar, M.Y.; Abuohashish, H.M.; Ahmed, M.M.; Al-Rejaie, S.S. Protective Effect of Lutein Supplementation on Oxidative Stress and Inflammatory Progression in Cerebral Cortex of Streptozotocin-Induced Diabetes in Rats. Neurochem. J. 2016, 10, 69–76. [Google Scholar] [CrossRef]
- Qiao, Y.-Q.; Jiang, P.-F.; Gao, Y.-Z. Lutein Prevents Osteoarthritis through Nrf2 Activation and Downregulation of Inflammation. Arch. Med. Sci. 2018, 14, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Kiko, T.; Hatade, K.; Sookwong, P.; Arai, H.; Miyazawa, T. Antioxidant Effect of Lutein towards Phospholipid Hydroperoxidation in Human Erythrocytes. Br. J. Nutr. 2009, 102, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Nakagawa, K.; Takekoshi, H.; Higuchi, O.; Kato, S.; Kondo, M.; Kimura, F.; Miyazawa, T. Ingestion of Chlorella Reduced the Oxidation of Erythrocyte Membrane Lipids in Senior Japanese Subjects. J. Oleo Sci. 2013, 62, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Min, K. Serum Lycopene, Lutein and Zeaxanthin, and the Risk of Alzheimer’s Disease Mortality in Older Adults. Dement. Geriatr. Cogn. Disord. 2014, 37, 246–256. [Google Scholar] [CrossRef]
- Ademowo, O.S.; Dias, H.K.I.; Milic, I.; Devitt, A.; Moran, R.; Mulcahy, R.; Howard, A.N.; Nolan, J.M.; Griffiths, H.R. Phospholipid Oxidation and Carotenoid Supplementation in Alzheimer’s Disease Patients. Free Radic. Biol. Med. 2017, 108, 77–85. [Google Scholar] [CrossRef]
- Avila-Roman, J.; Garda-Gil, S.; Rodriguez-Luna, A.; Motilva, V.; Talero, E. Anti-Inflammatory and Anticancer Effects of Microalgal Carotenoids. Mar. Drugs 2021, 19, 531. [Google Scholar] [CrossRef]
- Lakshminarayana, R.; Sathish, U.V.; Dharmesh, S.M.; Baskaran, V. Antioxidant and Cytotoxic Effect of Oxidized Lutein in Human Cervical Carcinoma Cells (HeLa). Food Chem. Toxicol. 2010, 48, 1811–1816. [Google Scholar] [CrossRef]
- Lakshminarayana, R.; Aruna, G.; Sathisha, U.V.; Dharmesh, S.M.; Baskaran, V. Structural Elucidation of Possible Lutein Oxidation Products Mediated through Peroxyl Radical Inducer 2,2′-Azobis (2-Methylpropionamidine) Dihydrochloride: Antioxidant and Cytotoxic Influence of Oxidized Lutein in HeLa Cells. Chem. Biol. Interact. 2013, 203, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Kavalappa, Y.P.; Gopal, S.S.; Ponesakki, G. Lutein Inhibits Breast Cancer Cell Growth by Suppressing Antioxidant and Cell Survival Signals and Induces Apoptosis. J. Cell. Physiol. 2021, 236, 1798–1809. [Google Scholar] [CrossRef] [PubMed]
- Baraya, Y.S.; Yankuzo, H.M.; Wong, K.K.; Yaacob, N.S. Strobilanthes Crispus Bioactive Subfraction Inhibits Tumor Progression and Improves Hematological and Morphological Parameters in Mouse Mammary Carcinoma Model. J. Ethnopharmacol. 2021, 267, 113522. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Smith, J.R.; Swanson, H.M.; Rubin, L.P. Carotenoid Lutein Selectively Inhibits Breast Cancer Cell Growth and Potentiates the Effect of Chemotherapeutic Agents through ROS-Mediated Mechanisms. Molecules 2018, 23, 905. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.L.; Zhao, Y.N.; Shi, Z.Z.; Cong, D.; Bai, Y.S. Lutein Inhibits Cell Growth and Activates Apoptosis via the PI3K/AKT/MTOR Signaling Pathway in A549 Human Non-Small-Cell Lung Cancer Cells. J. Environ. Pathol. Toxicol. Oncol. 2018, 37, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Fujiwara, A.; Onodera, D.; Motoki, T. Lutein Regulates the Expression of Apoptosis-Related Genes and Stem Cell Markers in A549 Human Lung Cancer Cells. Nat. Prod. Commun. 2017, 12, 897–900. [Google Scholar] [CrossRef]
- Reynoso-Camacho, R.; González-Jasso, E.; Ferriz-Martínez, R.; Villalón-Corona, B.; Loarca-Pina, G.F.; Salgado, L.M.; Ramos-Gomez, M. Dietary Supplementation of Lutein Reduces Colon Carcinogenesis in DMH-Treated Rats by Modulating K-Ras, PKB, and β-Catenin Proteins. Nutr. Cancer 2011, 63, 39–45. [Google Scholar] [CrossRef]
- Eom, J.W.; Lim, J.W.; Kim, H. Lutein Induces Reactive Oxygen Species-Mediated Apoptosis in Gastric Cancer AGS Cells via NADPH Oxidase Activation. Molecules 2023, 28, 1178. [Google Scholar] [CrossRef] [PubMed]
- Zaini, R.G.; Brandt, K.; Clench, M.R.; Le Maitre, C.L. Effects of Bioactive Compounds from Carrots (Daucus carota L.), Polyacetylenes, Beta-Carotene and Lutein on Human Lymphoid Leukaemia Cells. Anticancer. Agents Med. Chem. 2012, 12, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Philips, N.; Keller, T.; Hendrix, C.; Hamilton, S.; Arena, R.; Tuason, M.; Gonzalez, S. Regulation of the Extracellular Matrix Remodeling by Lutein in Dermal Fibroblasts, Melanoma Cells, and Ultraviolet Radiation Exposed Fibroblasts. Arch. Dermatol. Res. 2007, 299, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Rafi, M.M.; Kanakasabai, S.; Gokarn, S.V.; Krueger, E.G.; Bright, J.J. Dietary Lutein Modulates Growth and Survival Genes in Prostate Cancer Cells. J. Med. Food 2015, 18, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, E.R.; Firdous, A.P.; Ramnath, V.; Kuttan, R. Effect of Carotenoid Lutein on N-Nitrosodiethylamine-Induced Hepatocellular Carcinoma and Its Mechanism of Action. Eur. J. Cancer Prev. 2013, 22, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Gansukh, E.; Mya, K.K.; Jung, M.; Keum, Y.-S.; Kim, D.H.; Saini, R.K. Lutein Derived from Marigold (Tagetes erecta) Petals Triggers ROS Generation and Activates Bax and Caspase-3 Mediated Apoptosis of Human Cervical Carcinoma (HeLa) Cells. Food Chem. Toxicol. 2019, 127, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Sawasdee, N.; Jantakee, K.; Wathikthinnakon, M.; Panwong, S.; Pekkoh, J.; Duangjan, K.; Yenchitsomanus, P.T.; Panya, A. Microalga Chlorella sp. Extract Induced Apoptotic Cell Death of Cholangiocarcinoma via AKT/MTOR Signaling Pathway. Biomed. Pharmacother. 2023, 160, 114306. [Google Scholar] [CrossRef]
- Castro-Puyana, M.; Pérez-Sánchez, A.; Valdés, A.; Ibrahim, O.H.M.; Suarez-Álvarez, S.; Ferragut, J.A.; Micol, V.; Cifuentes, A.; Ibáñez, E.; García-Cañas, V. Pressurized Liquid Extraction of Neochloris oleoabundans for the Recovery of Bioactive Carotenoids with Anti-Proliferative Activity against Human Colon Cancer Cells. Food Res. Int. 2017, 99, 1048–1055. [Google Scholar] [CrossRef]
- Tavares-Carreón, F.; De La Torre-Zavala, S.; Arocha-Garza, H.F.; Souza, V.; Galán-Wong, L.J.; Avilés-Arnaut, H. In Vitro Anticancer Activity of Methanolic Extract of Granulocystopsis sp., a Microalgae from an Oligotrophic Oasis in the Chihuahuan Desert. PeerJ 2020, 2020, e8686. [Google Scholar] [CrossRef]
- Yan, B.; Lu, M.-S.; Wang, L.; Mo, X.-F.; Luo, W.-P.; Du, Y.-F.; Zhang, C.-X. Specific Serum Carotenoids Are Inversely Associated with Breast Cancer Risk among Chinese Women: A Case–Control Study. Br. J. Nutr. 2016, 115, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.-X.; Xing, M.-Y.; Yu, L.-F.; Shen, P. Carotenoid Intake and Esophageal Cancer Risk: A Meta-Analysis. Asian Pacific J. Cancer Prev. 2013, 14, 1911–1918. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, J.; Oh, J.H.; Chang, H.J.; Sohn, D.K.; Kwon, O.; Shin, A.; Kim, J. Dietary Lutein Plus Zeaxanthin Intake and DICER1 Rs3742330 A > G Polymorphism Relative to Colorectal Cancer Risk. Sci. Rep. 2019, 9, 3406. [Google Scholar] [CrossRef]
- Jansen, R.J.; Robinson, D.P.; Stolzenberg-Solomon, R.Z.; Bamlet, W.R.; de Andrade, M.; Oberg, A.L.; Rabe, K.G.; Anderson, K.E.; Olson, J.E.; Sinha, R.; et al. Nutrients from Fruit and Vegetable Consumption Reduce the Risk of Pancreatic Cancer. J. Gastrointest. Cancer 2013, 44, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Bravi, F.; Bosetti, C.; Filomeno, M.; Levi, F.; Garavello, W.; Galimberti, S.; Negri, E.; La Vecchia, C. Foods, Nutrients and the Risk of Oral and Pharyngeal Cancer. Br. J. Cancer 2013, 109, 2904–2910. [Google Scholar] [CrossRef] [PubMed]
- Leoncini, E.; Edefonti, V.; Hashibe, M.; Parpinel, M.; Cadoni, G.; Ferraroni, M.; Serraino, D.; Matsuo, K.; Olshan, A.F.; Zevallos, J.P.; et al. Carotenoid Intake and Head and Neck Cancer: A Pooled Analysis in the International Head and Neck Cancer Epidemiology Consortium. Eur. J. Epidemiol. 2016, 31, 369–383. [Google Scholar] [CrossRef]
- Wu, S.; Liu, Y.; Michalek, J.E.; Mesa, R.A.; Parma, D.L.; Rodriguez, R.; Mansour, A.M.; Svatek, R.; Tucker, T.C.; Ramirez, A.G. Carotenoid Intake and Circulating Carotenoids Are Inversely Associated with the Risk of Bladder Cancer: A Dose-Response Meta-Analysis. Adv. Nutr. 2020, 11, 630–643. [Google Scholar] [CrossRef]
- Bock, C.H.; Ruterbusch, J.J.; Holowatyj, A.N.; Steck, S.E.; Van Dyke, A.L.; Ho, W.J.; Cote, M.L.; Hofmann, J.N.; Davis, F.; Graubard, B.I.; et al. Renal Cell Carcinoma Risk Associated with Lower Intake of Micronutrients. Cancer Med. 2018, 7, 4087–4097. [Google Scholar] [CrossRef]
- Omar, W.M.; Ahmed, A.E.; Raslan, M.; El-Nesr, K.; Ali, M.M.; De Abdelmaksoud, M.; Dahshan, D. El Effect of Lutein-Rich Extract on Human Cancer Cells. Middle East J. Cancer 2021, 12, 147–150. [Google Scholar] [CrossRef]
- Wang, S.; Wang, M.; Zhang, S.; Zhao, L. Oxidative Stress in Rats with Hyperhomo-Cysteinemia and Intervention Effect of Lutein. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 359–364. [Google Scholar]
- Li, H.; Huang, C.; Zhu, J.; Gao, K.; Fang, J.; Li, H. Lutein Suppresses Oxidative Stress and Inflammation by Nrf2 Activation in an Osteoporosis Rat Model. Med. Sci. Monit. 2018, 24, 5071–5075. [Google Scholar] [CrossRef]
- Meurer, M.C.; Mees, M.; Mariano, L.N.B.; Boeing, T.; Somensi, L.B.; Mariott, M.; da Silva, R.d.C.M.V.d.A.F.; dos Santos, A.C.; Longo, B.; Santos França, T.C.; et al. Hydroalcoholic Extract of Tagetes erecta L. Flowers, Rich in the Carotenoid Lutein, Attenuates Inflammatory Cytokine Secretion and Improves the Oxidative Stress in an Animal Model of Ulcerative Colitis. Nutr. Res. 2019, 66, 95–106. [Google Scholar] [CrossRef]
- Mammadov, R.; Suleyman, B.; Akturan, S.; Cimen, F.K.; Kurt, N.; Suleyman, Z.; Malkoc, İ. Effect of Lutein on Methotrexate-Induced Oxidative Lung Damage in Rats: A Biochemical and Histopathological Assessment. Korean J. Intern. Med. 2019, 34, 1279–1286. [Google Scholar] [CrossRef]
- Melo van Lent, D.; Leermakers, E.T.M.; Hofman, A.; Stricker, B.H.; Brusselle, G.G.; Franco, O.H.; Lahousse, L.; Kiefte-de Jong, J.C. Association between Lutein Intake and Lung Function in Adults: The Rotterdam Study. Br. J. Nutr. 2017, 117, 720–730. [Google Scholar] [CrossRef]
- Khudhai, A.R.; Al-Shawi, N.N. Possible Protective Effects of High- versus Low- Dose of Lutein in Combination with Irinotecan on Liver of Rats: Role of Oxidative Stress and Apoptosis. Indian J. Forensic Med. Toxicol. 2021, 15, 2439–2445. [Google Scholar] [CrossRef]
- Bandariyan, E.; Mogheiseh, A.; Ahmadi, A. The Effect of Lutein and Urtica dioica Extract on in Vitro Production of Embryo and Oxidative Status in Polycystic Ovary Syndrome in a Model of Mice. BMC Complement. Med. Ther. 2021, 21, 55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ding, H.; Xu, L.; Zhao, S.; Hu, S.; Ma, A.; Ma, Y. Lutein Can Alleviate Oxidative Stress, Inflammation, and Apoptosis Induced by Excessive Alcohol to Ameliorate Reproductive Damage in Male Rats. Nutrients 2022, 14, 2385. [Google Scholar] [CrossRef] [PubMed]
- Fatani, A.J.; Al-Rejaie, S.S.; Abuohashish, H.M.; Al-Assaf, A.; Parmar, M.Y.; Ahmed, M.M. Lutein Dietary Supplementation Attenuates Streptozotocin-Induced Testicular Damage and Oxidative Stress in Diabetic Rats. BMC Complement. Altern. Med. 2015, 15, 204. [Google Scholar] [CrossRef] [PubMed]
- Sharavana, G.; Joseph, G.S.; Baskaran, V. Lutein Attenuates Oxidative Stress Markers and Ameliorates Glucose Homeostasis through Polyol Pathway in Heart and Kidney of STZ-Induced Hyperglycemic Rat Model. Eur. J. Nutr. 2017, 56, 2475–2485. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Zhang, Q.; Yan, F.-F.; Wan, J.-F.; Lin, C.-S. Lutein Protects against Ischemia/Reperfusion Injury in Rat Skeletal Muscle by Modulating Oxidative Stress and Inflammation. Immunopharmacol. Immunotoxicol. 2015, 37, 329–334. [Google Scholar] [CrossRef]
- Tan, D.; Yu, X.; Chen, M.; Chen, J.; Xu, J. Lutein Protects against Severe Traumatic Brain Injury through Anti-Inflammation and Antioxidative Effects via ICAM-1/Nrf-2. Mol. Med. Rep. 2017, 16, 4235–4240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, L.; Jia, L.; Liu, J. Association between Carotenoid Intake and Metabolic Dysfunction-Associated Fatty Liver Disease among US Adults: A Cross-Sectional Study. Medicine 2023, 102, e36658. [Google Scholar] [CrossRef]
- Yen, H.W.; Sun, C.H.; Ma, T.W. The Comparison of Lutein Production by Scenesdesmus sp. in the Autotrophic and the Mixotrophic Cultivation. Appl. Biochem. Biotechnol. 2011, 164, 353–361. [Google Scholar] [CrossRef]
- Heo, J.; Shin, D.S.; Cho, K.; Cho, D.H.; Lee, Y.J.; Kim, H.S. Indigenous Microalga Parachlorella sp. JD-076 as a Potential Source for Lutein Production: Optimization of Lutein Productivity via Regulation of Light Intensity and Carbon Source. Algal Res. 2018, 33, 1–7. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, Z.; Ho, S.H.; Ruan, C.; Li, J.; Xie, Y.; Shi, X.; Liu, L.; Chen, J. Two-Stage Bioprocess for Hyper-Production of Lutein from Microalga Chlorella sorokiniana FZU60: Effects of Temperature, Light Intensity, and Operation Strategies. Algal Res. 2020, 52, 102119. [Google Scholar] [CrossRef]
- Zhao, X.; Ma, R.; Liu, X.; Ho, S.H.; Xie, Y.; Chen, J. Strategies Related to Light Quality and Temperature to Improve Lutein Production of Marine Microalga Chlamydomonas sp. Bioprocess Biosyst. Eng. 2019, 42, 435–443. [Google Scholar] [CrossRef]
- Chen, W.C.; Hsu, Y.C.; Chang, J.S.; Ho, S.H.; Wang, L.F.; Wei, Y.H. Enhancing Production of Lutein by a Mixotrophic Cultivation System Using Microalga Scenedesmus obliquus CWL-1. Bioresour. Technol. 2019, 291, 121891. [Google Scholar] [CrossRef]
- Xie, Y.; Ho, S.H.; Chen, C.N.N.; Chen, C.Y.; Ng, I.S.; Jing, K.J.; Chang, J.S.; Lu, Y. Phototrophic Cultivation of a Thermo-Tolerant Desmodesmus sp. for Lutein Production: Effects of Nitrate Concentration, Light Intensity and Fed-Batch Operation. Bioresour. Technol. 2013, 144, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Vadrale, A.P.; Dong, C.-D.; Haldar, D.; Wu, C.-H.; Chen, C.-W.; Singhania, R.R.; Patel, A.K. Bioprocess Development to Enhance Biomass and Lutein Production from Chlorella sorokiniana Kh12. Bioresour. Technol. 2023, 370, 128583. [Google Scholar] [CrossRef]
- Ma, R.; Zhao, X.; Xie, Y.; Ho, S.-H.; Chen, J. Enhancing Lutein Productivity of Chlamydomonas sp. via High-Intensity Light Exposure with Corresponding Carotenogenic Genes Expression Profiles. Bioresour. Technol. 2019, 275, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Dineshkumar, R.; Subramanian, G.; Dash, S.K.; Sen, R. Development of an Optimal Light-Feeding Strategy Coupled with Semi-Continuous Reactor Operation for Simultaneous Improvement of Microalgal Photosynthetic Efficiency, Lutein Production and CO2 Sequestration. Biochem. Eng. J. 2016, 113, 47–56. [Google Scholar] [CrossRef]
- Ho, S.H.; Xie, Y.; Chan, M.C.; Liu, C.C.; Chen, C.Y.; Lee, D.J.; Huang, C.C.; Chang, J.S. Effects of Nitrogen Source Availability and Bioreactor Operating Strategies on Lutein Production with Scenedesmus obliquus FSP-3. Bioresour. Technol. 2015, 184, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lu, I.C.; Nagarajan, D.; Chang, C.H.; Ng, I.S.; Lee, D.J.; Chang, J.S. A Highly Efficient Two-Stage Cultivation Strategy for Lutein Production Using Heterotrophic Culture of Chlorella sorokiniana MB-1-M12. Bioresour. Technol. 2018, 253, 141–147. [Google Scholar] [CrossRef]
- Schüler, L.M.; Santos, T.; Pereira, H.; Duarte, P.; Katkam, N.G.; Florindo, C.; Schulze, P.S.C.; Barreira, L.; Varela, J.C.S. Improved Production of Lutein and β-Carotene by Thermal and Light Intensity Upshifts in the Marine Microalga Tetraselmis sp. CTP4. Algal Res. 2020, 45, 101732. [Google Scholar] [CrossRef]
- Xie, Y.; Zhao, X.; Chen, J.; Yang, X.; Ho, S.H.; Wang, B.; Chang, J.S.; Shen, Y. Enhancing Cell Growth and Lutein Productivity of Desmodesmus sp. F51 by Optimal Utilization of Inorganic Carbon Sources and Ammonium Salt. Bioresour. Technol. 2017, 244, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, J.; Ma, R.; Ho, S.H.; Shi, X.; Liu, L.; Chen, J. Bioprocess Operation Strategies with Mixotrophy/Photoinduction to Enhance Lutein Production of Microalga Chlorella sorokiniana FZU60. Bioresour. Technol. 2019, 290, 121798. [Google Scholar] [CrossRef]
- Chen, C.Y.; Ho, S.H.; Liu, C.C.; Chang, J.S. Enhancing Lutein Production with Chlorella sorokiniana Mb-1 by Optimizing Acetate and Nitrate Concentrations under Mixotrophic Growth. J. Taiwan Inst. Chem. Eng. 2017, 79, 88–96. [Google Scholar] [CrossRef]
- Shi, X.M.; Jiang, Y.; Chen, F. High-Yield Production of Lutein by the Green Microalga Chlorella protothecoides in Heterotrophic Fed-Batch Culture. Biotechnol. Prog. 2002, 18, 723–727. [Google Scholar] [CrossRef]
- Patel, A.K.; Vadrale, A.P.; Singhania, R.-R.; Chen, C.-W.; Chang, J.S.; Dong, C.-D. Enhanced Mixotrophic Production of Lutein and Lipid from Potential Microalgae Isolate Chlorella sorokiniana C16. Bioresour. Technol. 2023, 386, 129477. [Google Scholar] [CrossRef]
- Cuaresma, M.; Casal, C.; Forján, E.; Vílchez, C. Productivity and Selective Accumulation of Carotenoids of the Novel Extremophile Microalga Chlamydomonas acidophila Grown with Different Carbon Sources in Batch Systems. J. Ind. Microbiol. Biotechnol. 2011, 38, 167–177. [Google Scholar] [CrossRef]
- Shi, X.M.; Liu, H.J.; Zhang, X.W.; Chen, F. Production of Biomass and Lutein by Chlorella Protothecoides at Various Glucose Concentrations in Heterotrophic Cultures. Process Biochem. 1999, 34, 341–347. [Google Scholar] [CrossRef]
- Molino, A.; Mehariya, S.; Iovine, A.; Casella, P.; Marino, T.; Karatza, D.; Chianese, S.; Musmarra, D. Enhancing Biomass and Lutein Production from Scenedesmus almeriensis: Effect of Carbon Dioxide Concentration and Culture Medium Reuse. Front. Plant Sci. 2020, 11, 415. [Google Scholar] [CrossRef]
- Wei, D.; Chen, F.; Chen, G.; Zhang, X.W.; Liu, L.J.; Zhang, H. Enhanced Production of Lutein in Heterotrophic Chlorella protothecoides by Oxidative Stress. Sci. China Ser. C Life Sci. 2008, 51, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, E.; Ruiz-Domínguez, M.C.; Cuaresma, M.; Vaquero, I.; Ramos-Merchante, A.; Vega, J.M.; Vílchez, C.; Garbayo, I. Production of Lutein, and Polyunsaturated Fatty Acids by the Acidophilic Eukaryotic Microalga Coccomyxa onubensis under Abiotic Stress by Salt or Ultraviolet Light. J. Biosci. Bioeng. 2018, 125, 669–675. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, Z.; Tang, Z.; Ho, S.H.; Shi, X.; Liu, L.; Xie, Y.; Chen, J. Enhancement of Co-Production of Lutein and Protein in Chlorella sorokiniana FZU60 Using Different Bioprocess Operation Strategies. Bioresour. Bioprocess. 2021, 8, 82. [Google Scholar] [CrossRef]
- Moulton, T.P.; Burford, M.A. The Mass Culture of Dunaliella viridis (Volvocales, Chlorophyta) for Oxygenated Carotenoids: Laboratory and Pilot Plant Studies. Hydrobiologia 1990, 204–205, 401–408. [Google Scholar] [CrossRef]
- Jeon, J.Y.; Kwon, J.S.; Kang, S.T.; Kim, B.R.; Jung, Y.; Han, J.G.; Park, J.H.; Hwang, J.K. Optimization of Culture Media for Large-Scale Lutein Production by Heterotrophic Chlorella vulgaris. Biotechnol. Prog. 2014, 30, 736–743. [Google Scholar] [CrossRef]
- McClure, D.D.; Nightingale, J.K.; Luiz, A.; Black, S.; Zhu, J.; Kavanagh, J.M. Pilot-Scale Production of Lutein Using Chlorella vulgaris. Algal Res. 2019, 44, 101707. [Google Scholar] [CrossRef]
- Xie, Y.; Li, J.; Ho, S.H.; Ma, R.; Shi, X.; Liu, L.; Chen, J. Pilot-Scale Cultivation of Chlorella sorokiniana FZU60 with a Mixotrophy/Photoautotrophy Two-Stage Strategy for Efficient Lutein Production. Bioresour. Technol. 2020, 314, 123767. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, J.; Montero, Z.; Cuaresma, M.; Ruiz-Domínguez, M.-C.; Mogedas, B.; Nores, I.G.; González del Valle, M.; Vílchez, C. Outdoor Large-Scale Cultivation of the Acidophilic Microalga Coccomyxa onubensis in a Vertical Close Photobioreactor for Lutein Production. Processes 2020, 8, 324. [Google Scholar] [CrossRef]
- Cavieres, L.; Bazaes, J.; Marticorena, P.; Riveros, K.; Medina, P.; Sepúlveda, C.; Riquelme, C. Pilot-Scale Phycoremediation Using Muriellopsis sp. For Wastewater Reclamation in the Atacama Desert: Microalgae Biomass Production and Pigment Recovery. Water Sci. Technol. 2021, 83, 331–343. [Google Scholar] [CrossRef]
- Van, T.; Do, C.; Dinh, C.T.; Dang, M.T.; Dang Tran, T.; Giang Le, T. A Novel Flat-Panel Photobioreactor for Simultaneous Production of Lutein and Carbon Sequestration by Chlorella sorokiniana TH01. Bioresour. Technol. 2022, 345, 126552. [Google Scholar] [CrossRef]
- Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Microalgal Lutein Biosynthesis: Recent Trends and Challenges to Enhance the Lutein Content in Microalgal Cell Factories. Front. Mar. Sci. 2022, 9, 1015419. [Google Scholar] [CrossRef]
- Huang, W.; Lin, Y.; He, M.; Gong, Y.; Huang, J. Induced High-Yield Production of Zeaxanthin, Lutein, and β-Carotene by a Mutant of Chlorella zofingiensis. J. Agric. Food Chem. 2018, 66, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Cordero, B.F.; Obraztsova, I.; Couso, I.; Leon, R.; Vargas, M.A.; Rodriguez, H. Enhancement of Lutein Production in Chlorella sorokiniana (Chorophyta) by Improvement of Culture Conditions and Random Mutagenesis. Mar. Drugs 2011, 9, 1607–1624. [Google Scholar] [CrossRef] [PubMed]
- Cordero, B.F.; Couso, I.; León, R.; Rodríguez, H.; Vargas, M.Á. Enhancement of Carotenoids Biosynthesis in Chlamydomonas reinhardtii by Nuclear Transformation Using a Phytoene Synthase Gene Isolated from Chlorella zofingiensis. Appl. Microbiol. Biotechnol. 2011, 91, 341–351. [Google Scholar] [CrossRef]
- Liu, J.; Gerken, H.; Huang, J.; Chen, F. Engineering of an Endogenous Phytoene Desaturase Gene as a Dominant Selectable Marker for Chlamydomonas reinhardtii Transformation and Enhanced Biosynthesis of Carotenoids. Process Biochem. 2013, 48, 788–795. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Gerken, H.; Huang, J.; Jiang, Y.; Chen, F. Genetic Engineering of the Green Alga Chlorella zofingiensis: A Modified Norflurazon-Resistant Phytoene Desaturase Gene as a Dominant Selectable Marker. Appl. Microbiol. Biotechnol. 2014, 98, 5069–5079. [Google Scholar] [CrossRef] [PubMed]
- Rathod, J.P.; Vira, C.; Lali, A.M.; Prakash, G. Metabolic Engineering of Chlamydomonas reinhardtii for Enhanced β-Carotene and Lutein Production. Appl. Biochem. Biotechnol. 2020, 190, 1457–1469. [Google Scholar] [CrossRef]
- Takemura, M.; Kubo, A.; Watanabe, A.; Sakuno, H.; Minobe, Y.; Sahara, T.; Murata, M.; Araki, M.; Harada, H.; Terada, Y.; et al. Pathway Engineering for High-Yield Production of Lutein in Escherichia coli. Synth. Biol. 2021, 6, ysab012. [Google Scholar] [CrossRef]
- Bian, Q.; Zhou, P.; Yao, Z.; Li, M.; Yu, H.; Ye, L. Heterologous Biosynthesis of Lutein in S. cerevisiae Enabled by Temporospatial Pathway Control. Metab. Eng. 2021, 67, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the Health Potential of Microalgae as Sustainable Sources of Bioactive Compounds. Int. J. Mol. Sci. 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montuori, E.; Lima, S.; Marchese, A.; Scargiali, F.; Lauritano, C. Lutein Production and Extraction from Microalgae: Recent Insights and Bioactive Potential. Int. J. Mol. Sci. 2024, 25, 2892. https://doi.org/10.3390/ijms25052892
Montuori E, Lima S, Marchese A, Scargiali F, Lauritano C. Lutein Production and Extraction from Microalgae: Recent Insights and Bioactive Potential. International Journal of Molecular Sciences. 2024; 25(5):2892. https://doi.org/10.3390/ijms25052892
Chicago/Turabian StyleMontuori, Eleonora, Serena Lima, Arima Marchese, Francesca Scargiali, and Chiara Lauritano. 2024. "Lutein Production and Extraction from Microalgae: Recent Insights and Bioactive Potential" International Journal of Molecular Sciences 25, no. 5: 2892. https://doi.org/10.3390/ijms25052892
APA StyleMontuori, E., Lima, S., Marchese, A., Scargiali, F., & Lauritano, C. (2024). Lutein Production and Extraction from Microalgae: Recent Insights and Bioactive Potential. International Journal of Molecular Sciences, 25(5), 2892. https://doi.org/10.3390/ijms25052892