
Yeast-Derived Nucleotides Enhance Fibroblast Migration and Proliferation and Provide Clinical Benefits in Atopic Dermatitis

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
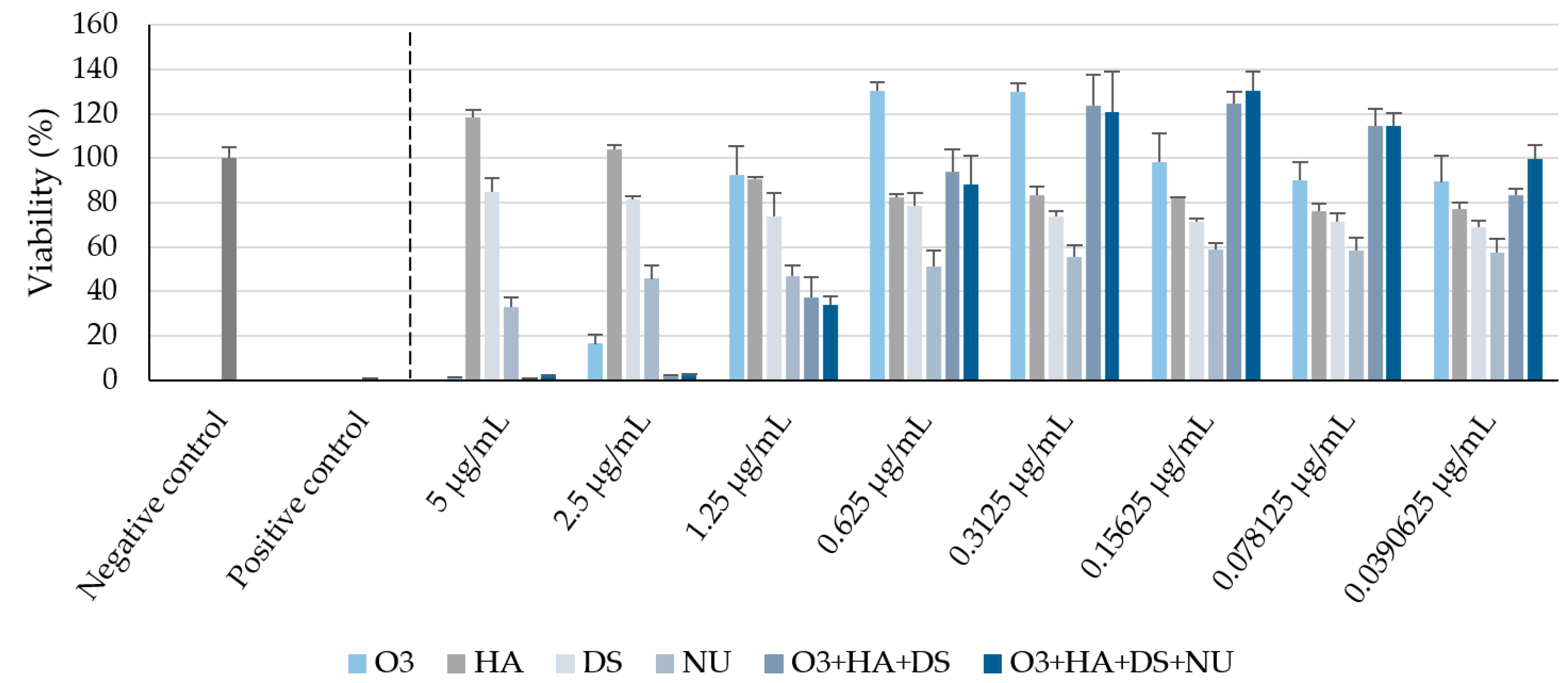
2.1. Cell Viability
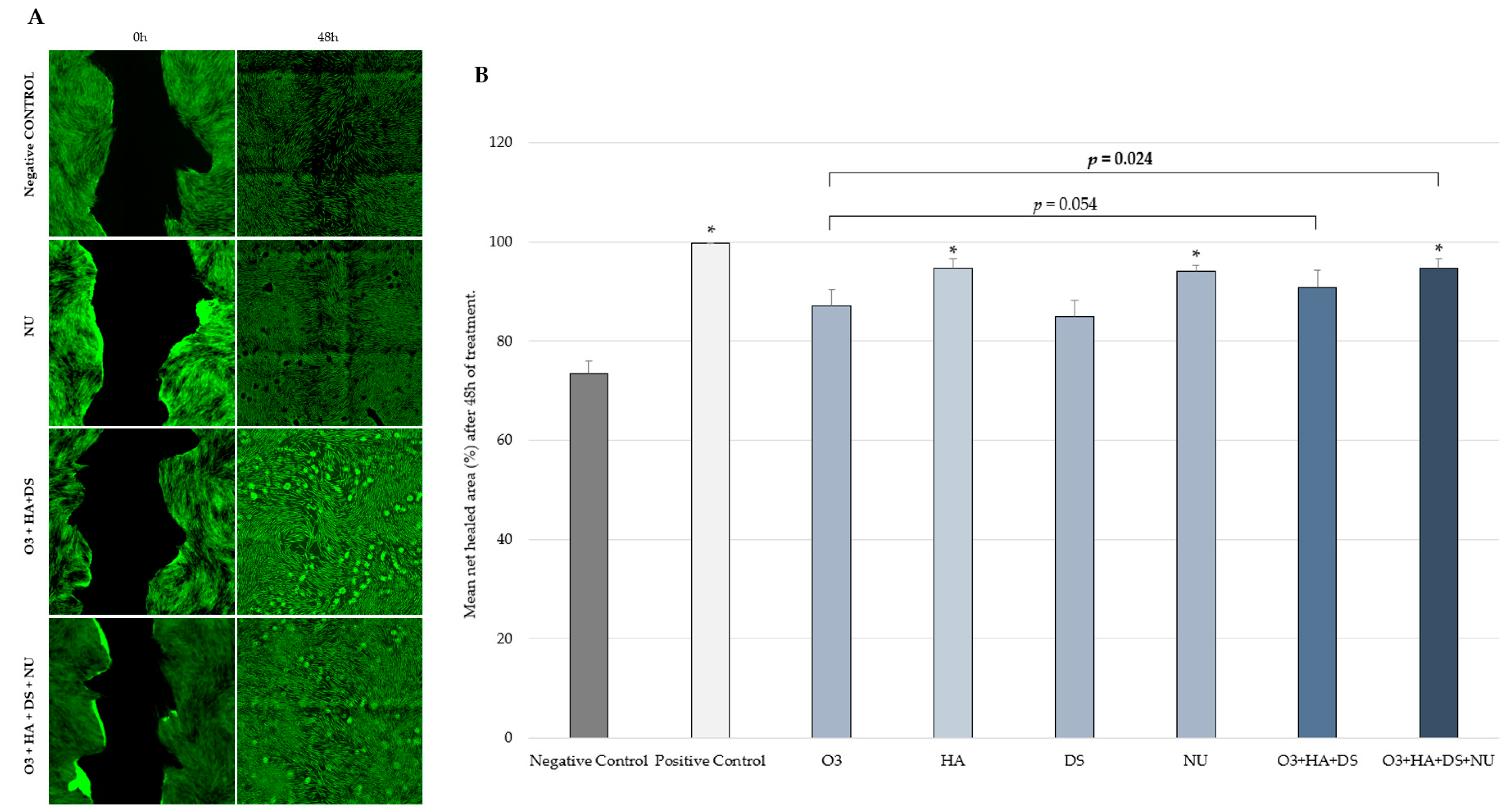
2.2. Fibroblast Migration
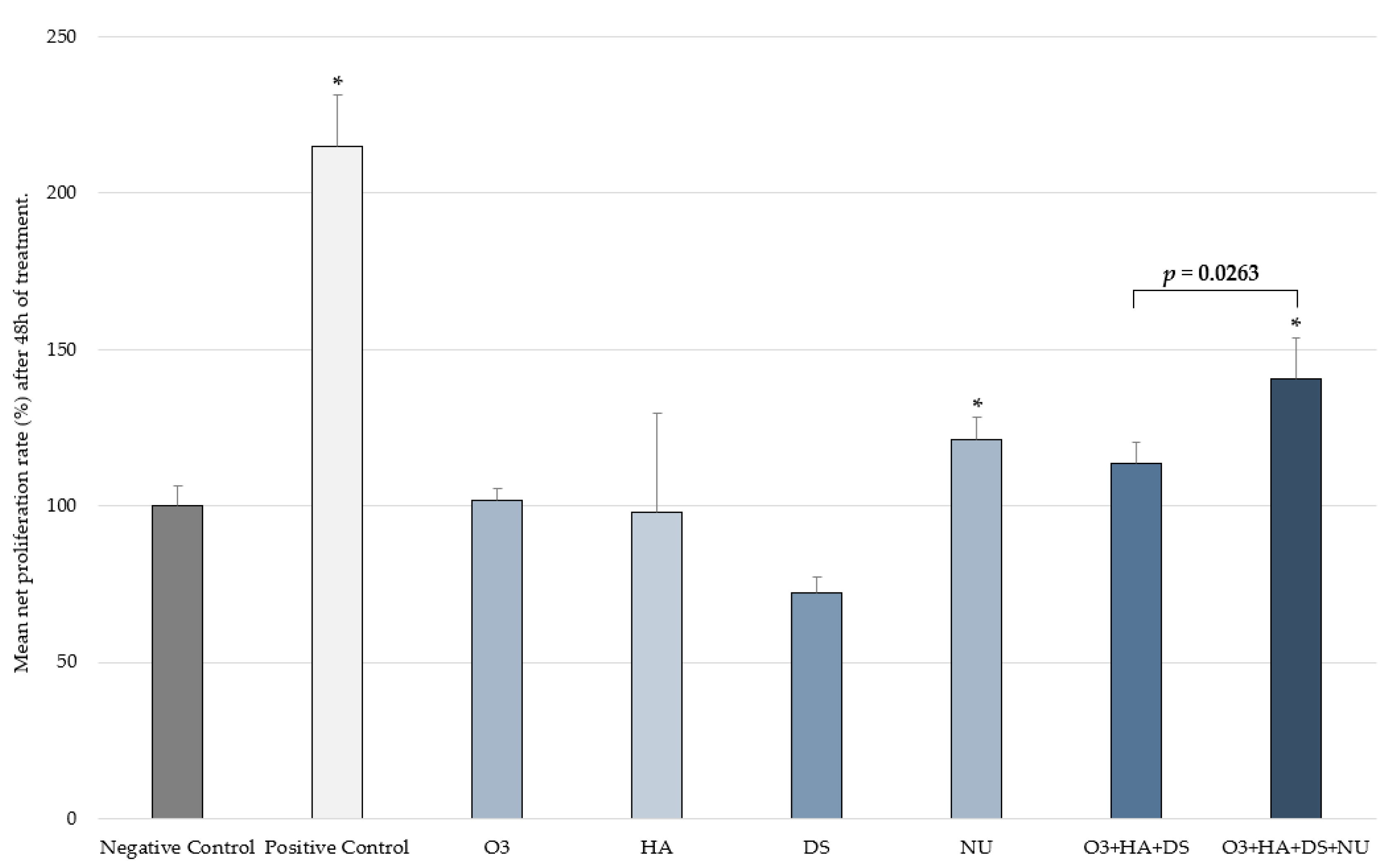
2.3. Fibroblast Proliferation
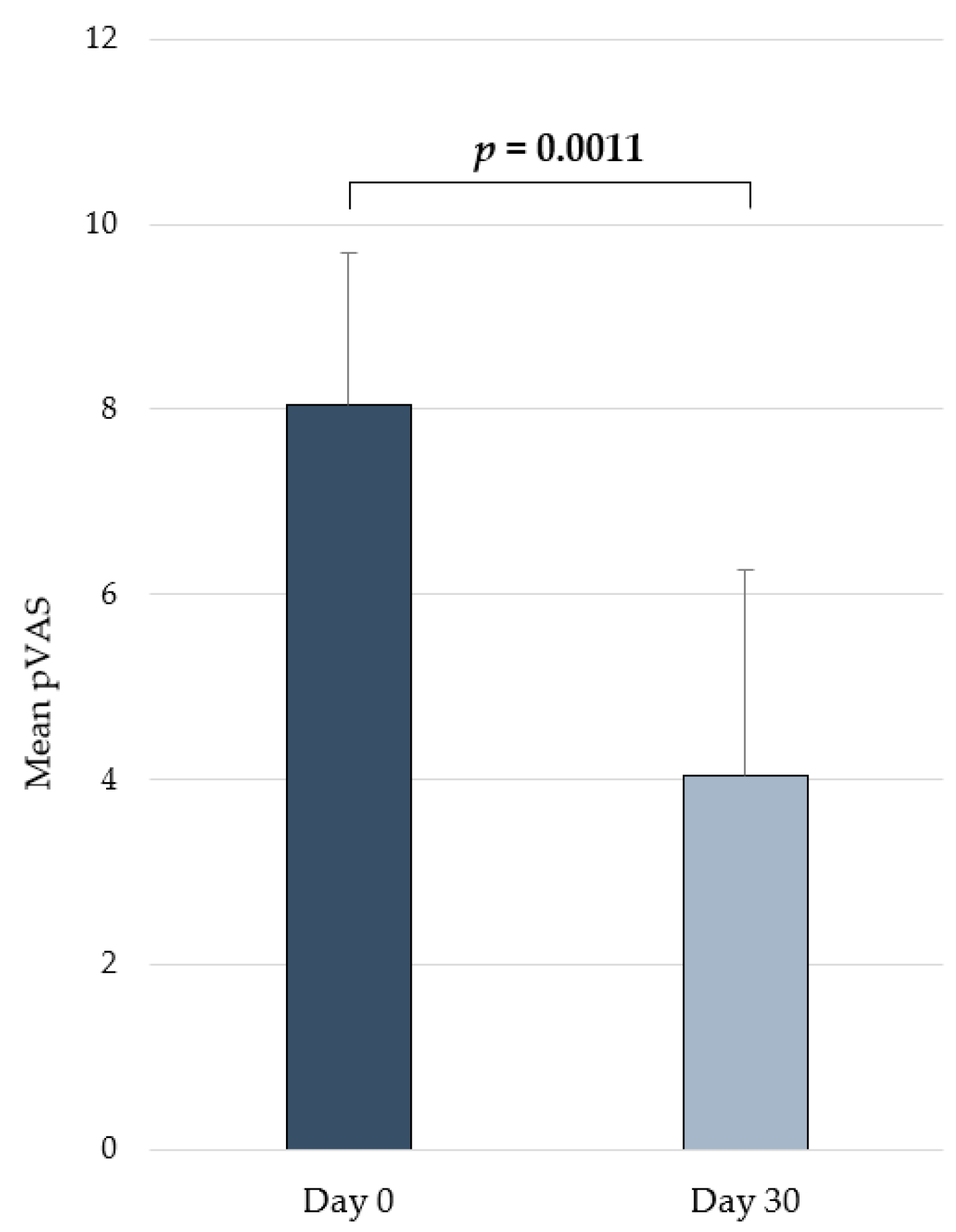
2.4. Clinical Efficacy in Atopic Dogs
3. Discussion
4. Materials and Methods
4.1. In Vitro Studies
4.2. In Vivo Testing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fontana, L.; Martínez-Augustin, O.; Gil, Á. Role of Dietary Nucleotides in Immunity. Funct. Food Rev. 2010, 2, 91–100. [Google Scholar]
- Gil, A. Modulation of the Immune Response Mediated by Dietary Nucleotides. Eur. J. Clin. Nutr. 2002, 56, S1–S4. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.R.; Greenberg, N.A. The Role of Nucleotides in the Immune and Gastrointestinal Systems. Nutr. Clin. Pract. 2012, 27, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Song, G.; Liu, X.; Xu, M.; Li, Y. Nucleotides as Optimal Candidates for Essential Nutrients in Living Organisms: A Review. J. Funct. Foods 2021, 82, 104498. [Google Scholar] [CrossRef]
- Segarra, S. Nutritional Modulation of the Immune Response Mediated by Nucleotides in Canine Leishmaniosis. Microorganisms 2021, 9, 2601. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.D.; Fanslow, W.C.; Higley, H.; Pizzini, R.; Rudolph, F.B.; van Buren, C.T. Expression of Immune Cell Surface Markers in Vivo and Immune Competence in Mice by Dietary Nucleotides. Transplant. Proc. 1989, 21, 121–124. [Google Scholar]
- Van Buren, C.T.; Rudolph, F. Dietary Nucleotides: A Conditional Requirement. Nutrition 1997, 13, 470–472. [Google Scholar] [CrossRef]
- Tao, Z.; Yuan, H.; Liu, M.; Liu, Q.; Zhang, S.; Liu, H.; Jiang, Y.; Huang, D.; Wang, T. Yeast Extract: Characteristics, Production, Applications and Future Perspectives. J. Microbiol. Biotechnol. 2023, 33, 151–166. [Google Scholar] [CrossRef]
- Maturana, M.; Castillejos, L.; Martin-Orue, S.M.; Minel, A.; Chetty, O.; Felix, A.P.; Adib, A. Potential Benefits of Yeast Saccharomyces and Their Derivatives in Dogs and Cats: A Review. Front. Vet. Sci. 2023, 10, 1279506. [Google Scholar] [CrossRef]
- Marsella, R. Advances in Our Understanding of Canine Atopic Dermatitis. Vet. Dermatol. 2021, 32, 547-e151. [Google Scholar] [CrossRef]
- Marsella, R. Atopic Dermatitis in Domestic Animals: What Our Current Understanding Is and How This Applies to Clinical Practice. Vet. Sci. 2021, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- Marsella, R.; De Benedetto, A. Atopic Dermatitis in Animals and People: An Update and Comparative Review. Vet. Sci. 2017, 4, 37. [Google Scholar] [CrossRef] [PubMed]
- Marsella, R.; Segarra, S.; Ahrens, K.; Alonso, C.; Ferrer, L. Topical Treatment with Sphingolipids and Glycosaminoglycans for Canine Atopic Dermatitis. BMC Vet. Res. 2020, 16, 92. [Google Scholar] [CrossRef] [PubMed]
- Egawa, G.; Kabashima, K. Barrier Dysfunction in the Skin Allergy. Allergol. Int. 2018, 67, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Combarros, D.; Goudounèche, D.; Cadiergues, M.C.; Simon, M. The Upper Epidermis of Atopic Dogs Is Altered at the Functional and Structural Levels. Vet. Dermatol. 2021, 32, 620-e165. [Google Scholar] [CrossRef]
- Ständer, S. Atopic Dermatitis. N. Engl. J. Med. 2021, 384, 1136–11434. [Google Scholar] [CrossRef]
- Mesjasz, A.; Zawadzka, M.; Chałubiński, M.; Trzeciak, M. Is Atopic Dermatitis Only a Skin Disease? Int. J. Mol. Sci. 2023, 24, 837. [Google Scholar] [CrossRef]
- Santoro, D.; Saridomichelakis, M.; Eisenschenk, M.; Tamamoto-Mochizuki, C.; Hensel, P.; Pucheu-Haston, C. Update on the Skin Barrier, Cutaneous Microbiome and Host Defence Peptides in Canine Atopic Dermatitis. Vet. Dermatol. 2023, 35, 5–14. [Google Scholar] [CrossRef]
- Fernandes, B.; Alves, S.; Schmidt, V.; Bizarro, A.F.; Pinto, M.; Pereira, H.; Marto, J.; Lourenço, A.M. Primary Prevention of Canine Atopic Dermatitis: Breaking the Cycle—A Narrative Review. Vet. Sci. 2023, 10, 659. [Google Scholar] [CrossRef]
- Miller, J.; Simpson, A.; Bloom, P.; Diesel, A.; Friedeck, A.; Paterson, T.; Wisecup, M.; Yu, C.M. 2023 AAHA Management of Allergic Skin Diseases in Dogs and Cats Guidelines. J. Am. Anim. Hosp. Assoc. 2023, 59, 255–284. [Google Scholar] [CrossRef] [PubMed]
- Olivry, T.; DeBoer, D.J.; Favrot, C.; Jackson, H.A.; Mueller, R.S.; Nuttall, T.; Prélaud, P. Treatment of Canine Atopic Dermatitis: 2015 Updated Guidelines from the International Committee on Allergic Diseases of Animals (ICADA). BMC Vet. Res. 2015, 11, 210. [Google Scholar] [CrossRef] [PubMed]
- Popa, I.; Pin, D.; Remoué, N.; Remoué, N.; Osta, B.; Callejon, S.; Videmont, E.; Gatto, H.; Portoukalian, J.; Haftek, M. Analysis of Epidermal Lipids in Normal and Atopic Dogs, before and after Administration of an Oral Omega-6/Omega-3 Fatty Acid Feed Supplement. A Pilot Study. Vet. Res. Commun. 2011, 35, 501–509. [Google Scholar] [CrossRef]
- Mueller, R.S.; Fieseler, K.V.; Fettman, M.J.; Zabel, S.; Rosychuk, R.A.W.; Ogilvie, G.K.; Greenwalt, T.L. Effect of Omega-3 Fatty Acids on Canine Atopic Dermatitis. J. Small Anim. Pract. 2004, 45, 293–297. [Google Scholar] [CrossRef]
- Olivry, T.; Marsella, R.; Hillier, A. The ACVD Task Force on Canine Atopic Dermatitis (XXIII): Are Essential Fatty Acids Effective? Vet. Immunol. Immunopathol. 2001, 81, 347–362. [Google Scholar] [CrossRef]
- Paterson, S. Supplementary Therapy in Canine Atopic Dermatitis. Companion Anim. 2019, 24, 400–407. [Google Scholar] [CrossRef]
- Grimble, R.F. Nutritional Modulation of Immune Function. Proc. Nutr. Soc. 2001, 60, 389–397. [Google Scholar] [CrossRef]
- De Abreu Nascimento, M.; da Matta Alvarez Pimenta, N.; Aiceles de Medeiros Pinto Polastri, V.; Cardoso Chamon, R.; Sarto Figueiredo, M. Immunonutrients and Intestinal Microbiota: A Gap in the Literature. Crit. Rev. Food Sci. Nutr. 2023, ahead of print. [Google Scholar] [CrossRef]
- Smith, M.M.; Melrose, J. Proteoglycans in Normal and Healing Skin. Adv. Wound Care 2013, 4, 152–173. [Google Scholar] [CrossRef] [PubMed]
- Callejas-quijada, G.; Campos-Lozada, G.; Aguirre, G. Hyaluronic Acid—Extraction Methods, Sources and Applications. Polymers 2023, 15, 3473. [Google Scholar] [CrossRef]
- Torrent, A.; Ruhí, R.; Martínez, C.; Castells, G.; de Castellarnau-Castellà, C. Anti-Inflammatory Activity and Absorption of a Natural Rooster Comb Extract (Hyal-Joint®). Osteoarthr. Cartil. 2010, 18, S246–S247. [Google Scholar] [CrossRef]
- Torrent, A.; Montell, E.; Verges, J.; Ruhi, R.; Dalmau, P.; Zurbano, M.J.; Romero, J. A New Natural Extract with Anti-Aging and Regenerative Properties for Skin. FASEB J. 2015, 29, 740–743. [Google Scholar] [CrossRef]
- Galvez-Martin, P.; Martinez-Puig, D.; Romero-Rueda, J. Comparative in Vitro Efficacy of a Hyaluronic Acid (HA) Matrix Ingredient Containing HA, Dermatan Sulphate and Collagen (Dermial®) versus Pure HA from Extraction or Fermentation Origin Introduction. FASEB J. 2022, 36, 5–7. [Google Scholar] [CrossRef]
- Galvez-martin, P.; Soto-fernandez, C.; Romero-rueda, J.; Cabañas, J.; Torrent, A.; Castells, G.; Martinez-puig, D. A Novel Hyaluronic Acid Matrix Ingredient with Regenerative, Anti-Aging and Antioxidant Capacity. Int. J. Mol. Sci. 2023, 24, 4774. [Google Scholar] [CrossRef]
- Gatmaitan, J.G.; Lee, J.H. Challenges and Future Trends in Atopic Dermatitis. Int. J. Mol. Sci. 2023, 24, 1380. [Google Scholar] [CrossRef]
- Berroth, A.; Kühnl, J.; Kurschat, N.; Schwarz, A.; Stäb, F.; Schwarz, T.; Wenck, H.; Fölster-Holst, R.; Neufang, G. Role of Fibroblasts in the Pathogenesis of Atopic Dermatitis. J. Allergy Clin. Immunol. 2013, 131, 1547–1554. [Google Scholar] [CrossRef]
- Marsella, R.; Olivry, T.; Carlotti, D.-N. Current Evidence of Skin Barrier Dysfunction in Human and Canine Atopic Dermatitis. Vet. Dermatol. 2011, 22, 239–248. [Google Scholar] [CrossRef]
- Nagafuchi, S.; Hachimura, S.; Totsuka, M.; Takahashi, T.; Goto, M.; Yajima, T.; Kuwata, T.; Habu, S.; Kaminogawa, S. Dietary Nucleotides Can Up-Regulate Antigen-Specific Th1 Immune Responses and Suppress Antigen-Specific IgE Responses in Mice. Int. Arch. Allergy Immunol. 2000, 122, 33–41. [Google Scholar] [CrossRef]
- Davies, J. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef]
- Caneschi, A.; Bardhi, A.; Barbarossa, A.; Zaghini, A. The Use of Antibiotics and Antimicrobial Resistance in Veterinary Medicine, a Complex Phenomenon: A Narrative Review. Antibiotics 2023, 12, 487. [Google Scholar] [CrossRef]
- Tao, R.; Li, R.; Wang, R. Dysbiosis of Skin Mycobiome in Atopic Dermatitis. Mycoses 2022, 65, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.; Künstner, A.; Wohlers, I.; Olbrich, M.; Lenfers, T.; Osumi, T.; Shimazaki, Y.; Nishifuji, K.; Ibrahim, S.M.; Watson, A.; et al. A Comprehensive Analysis of Gut and Skin Microbiota in Canine Atopic Dermatitis in Shiba Inu Dogs. Microbiome 2022, 11, 232. [Google Scholar] [CrossRef]
- Marques, V.S.; Pimenta, A.; Bicalho, V. Evaluation of Quality of Life in Dogs with Atopic Dermatitis and Their Owners after Lokivetmab Therapy. Res. Soc. Dev. 2021, 2021, e392101119775. [Google Scholar] [CrossRef]
- Souto, E.B.; Dias-Ferreira, J.; Oliveira, J.; Sanchez-Lopez, E.; Lopez-Machado, A.; Espina, M.; Garcia, M.L.; Souto, S.B.; Martins-Gomes, C.; Silva, A.M. Trends in Atopic Dermatitis—From Standard Pharmacotherapy to Novel Drug Delivery Systems. Int. J. Mol. Sci. 2019, 20, 5659. [Google Scholar] [CrossRef]
- Segarra, S.; Sanmiguel, D.; Zuriaga, E.; Leclerc, S.; Cabañas, J.; Seigneuric, E.; Miquel, A.; Vázquez, A.; Ferrer, L. Sphingomyelin-Rich Lipid Extract Collar for Canine Atopic Dermatitis. Vet. Sci. 2023, 10, 389. [Google Scholar] [CrossRef]
- Nuttall, T.J.; Marsella, R.; Rosenbaum, M.R.; Gonzales, A.J.; Fadok, V.A. Update on Pathogenesis, Diagnosis, and Treatment of Atopic Dermatitis in Dogs. J. Am. Vet. Med. Assoc. 2019, 254, 1291–1300. [Google Scholar] [CrossRef]
- Gedon, N.K.Y.; Mueller, R.S. Atopic Dermatitis in Cats and Dogs: A Difficult Disease for Animals and Owners. Clin. Transl. Allergy 2018, 8, 41. [Google Scholar] [CrossRef]
- Segarra, S.; Naiken, T.; Garnier, J.; Hamon, V.; Coussay, N.; Bernard, F.X. Enhanced In Vitro Expression of Filaggrin and Antimicrobial Peptides Following Application of Glycosaminoglycans and a Sphingomyelin-Rich Lipid Extract. Vet. Sci. 2022, 9, 323. [Google Scholar] [CrossRef]
- Bruet, V.; Mosca, M.; Briand, A.; Bourdeau, P.; Pin, D.; Cochet-faivre, N.; Cadiergues, M. Clinical Guidelines for the Use of Antipruritic Drugs in the Control of the Most Frequent Pruritic Skin Diseases in Dogs. Vet. Sci. 2022, 9, 149. [Google Scholar] [CrossRef]
- Outerbridge, C.A.; Jordan, T.J.M. Current Knowledge on Canine Atopic Dermatitis. Adv. Small Anim. Care 2021, 2, 101–115. [Google Scholar] [CrossRef]
- Segarra, S.; Miró, G.; Montoya, A.; Pardo-Marín, L.; Boqué, N.; Ferrer, L.; Cerón, J. Randomized, Allopurinol-Controlled Trial of the Effects of Dietary Nucleotides and Active Hexose Correlated Compound in the Treatment of Canine Leishmaniosis. Vet. Parasitol. 2017, 239, 50–56. [Google Scholar] [CrossRef]
- Segarra, S.; Miró, G.; Montoya, A.; Pardo-Marín, L.; Teichenné, J.; Ferrer, L.; Cerón, J.J. Prevention of Disease Progression in Leishmania Infantum-Infected Dogs with Dietary Nucleotides and Active Hexose Correlated Compound. Parasites Vectors 2018, 11, 103. [Google Scholar] [CrossRef]
- Barroso, C.D.N.; Imamura, L.M.; Cordeiro, C.C.; Ingberman, M.; Caron, L.F. Nucleotide Supplementation as a Novel Adjunctive Therapy for Canine Atopic Dermatitis. Vet. Res. Commun. 2023, 1–11. [Google Scholar] [CrossRef]
- Santoro, D.; Pucheu-Haston, C.M.; Prost, C.; Mueller, R.S.; Jackson, H. Clinical Signs and Diagnosis of Feline Atopic Syndrome: Detailed Guidelines for a Correct Diagnosis. Vet. Dermatol. 2021, 32, 26-e6. [Google Scholar] [CrossRef]
- Halliwell, R.; Pucheu-Haston, C.M.; Olivry, T.; Prost, C.; Jackson, H.; Banovic, F.; Nuttall, T.; Santoro, D.; Bizikova, P.; Mueller, R.S. Feline Allergic Diseases: Introduction and Proposed Nomenclature. Vet. Dermatol. 2021, 32, 8-e2. [Google Scholar] [CrossRef]
- Favrot, C.; Steffan, J.; Seewald, W.; Picco, F. A Prospective Study on the Clinical Features of Chronic Canine Atopic Dermatitis and Its Diagnosis. Vet. Dermatol. 2010, 21, 23–31. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segarra, S.; Bošnjak, I.; Mioč, I.; Čurčija, B.; Csik, V.-A.; Krešić, S.; Romero-Rueda, J.; Rodríguez, A.; Martínez-Puig, D. Yeast-Derived Nucleotides Enhance Fibroblast Migration and Proliferation and Provide Clinical Benefits in Atopic Dermatitis. Int. J. Mol. Sci. 2024, 25, 2890. https://doi.org/10.3390/ijms25052890
Segarra S, Bošnjak I, Mioč I, Čurčija B, Csik V-A, Krešić S, Romero-Rueda J, Rodríguez A, Martínez-Puig D. Yeast-Derived Nucleotides Enhance Fibroblast Migration and Proliferation and Provide Clinical Benefits in Atopic Dermatitis. International Journal of Molecular Sciences. 2024; 25(5):2890. https://doi.org/10.3390/ijms25052890
Chicago/Turabian StyleSegarra, Sergi, Ivica Bošnjak, Igor Mioč, Bojana Čurčija, Vlatka-Antonija Csik, Srećko Krešić, Jessica Romero-Rueda, Anna Rodríguez, and Daniel Martínez-Puig. 2024. "Yeast-Derived Nucleotides Enhance Fibroblast Migration and Proliferation and Provide Clinical Benefits in Atopic Dermatitis" International Journal of Molecular Sciences 25, no. 5: 2890. https://doi.org/10.3390/ijms25052890
APA StyleSegarra, S., Bošnjak, I., Mioč, I., Čurčija, B., Csik, V.-A., Krešić, S., Romero-Rueda, J., Rodríguez, A., & Martínez-Puig, D. (2024). Yeast-Derived Nucleotides Enhance Fibroblast Migration and Proliferation and Provide Clinical Benefits in Atopic Dermatitis. International Journal of Molecular Sciences, 25(5), 2890. https://doi.org/10.3390/ijms25052890