
Transcriptome Analysis Reveals the Mechanism by Which Exogenous Melatonin Treatment Delays Leaf Senescence of Postharvest Chinese Kale (Brassica oleracea var. alboglabra)

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
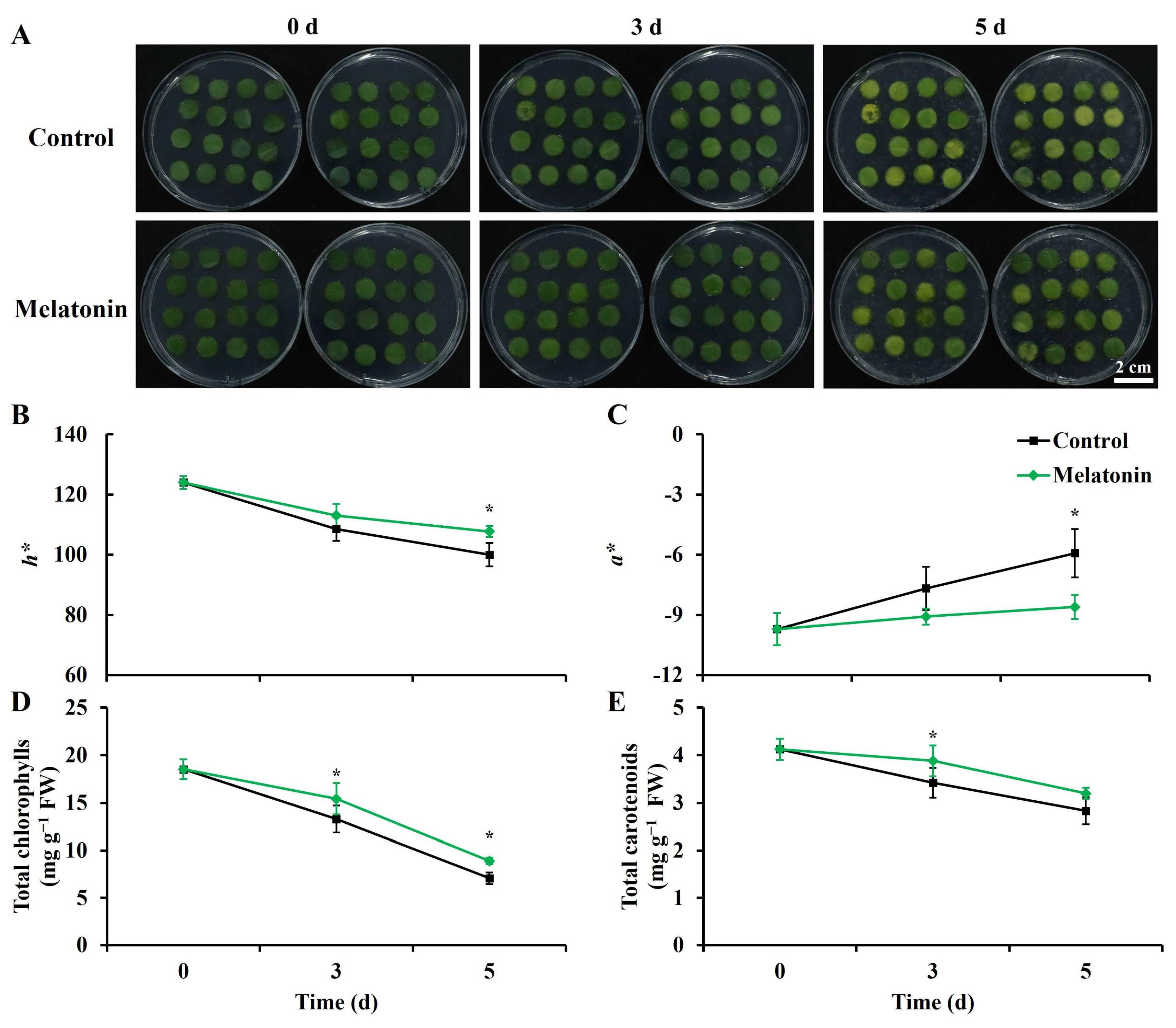
2.1. Melatonin Reduced Leaf Senescence in Postharvest Chinese Kale
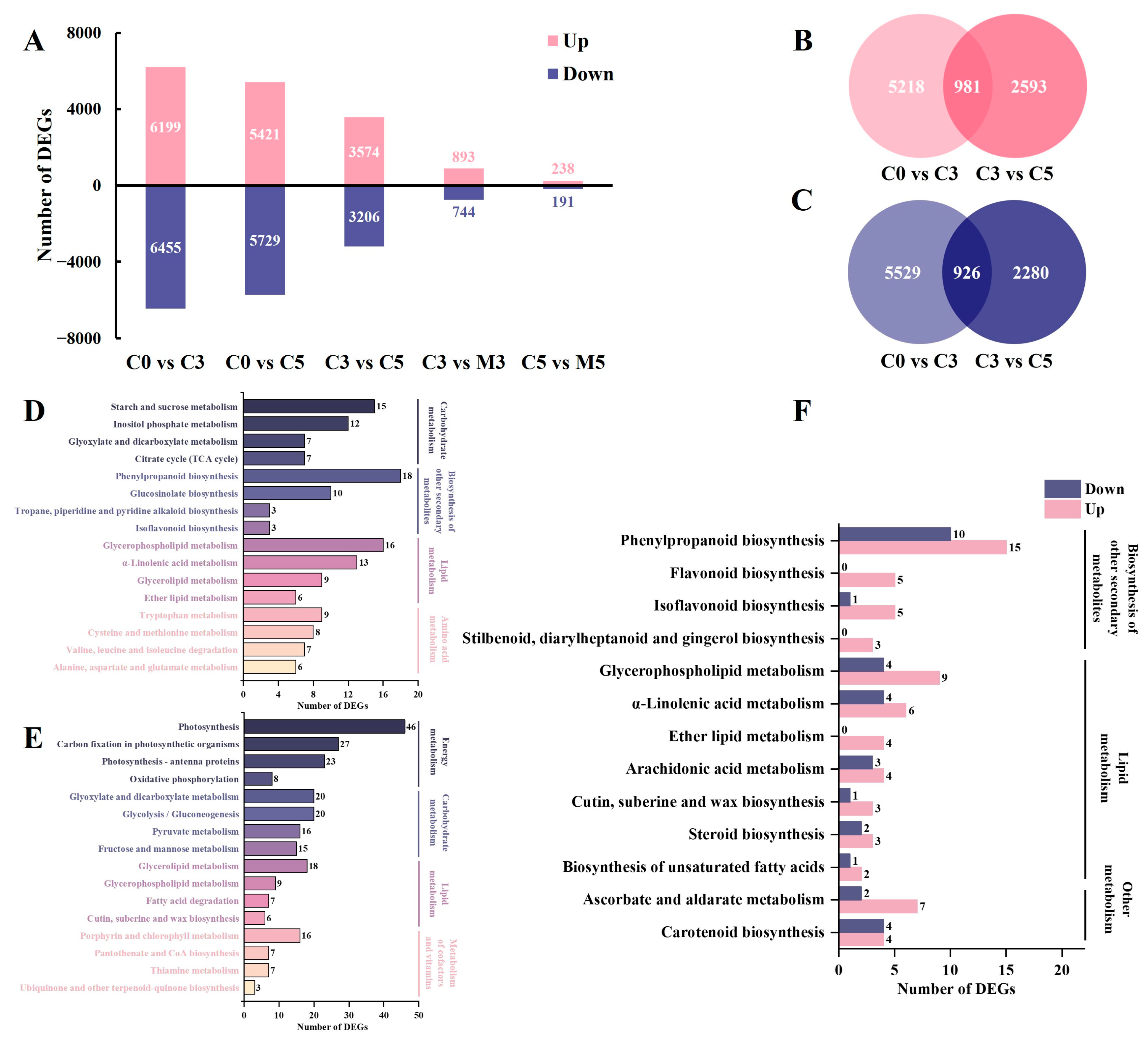
2.2. Differential Gene Expression
2.3. Verification of RNA-Seq Data by qRT-PCR
2.4. Global mRNA Response to Senescence and Melatonin Treatment
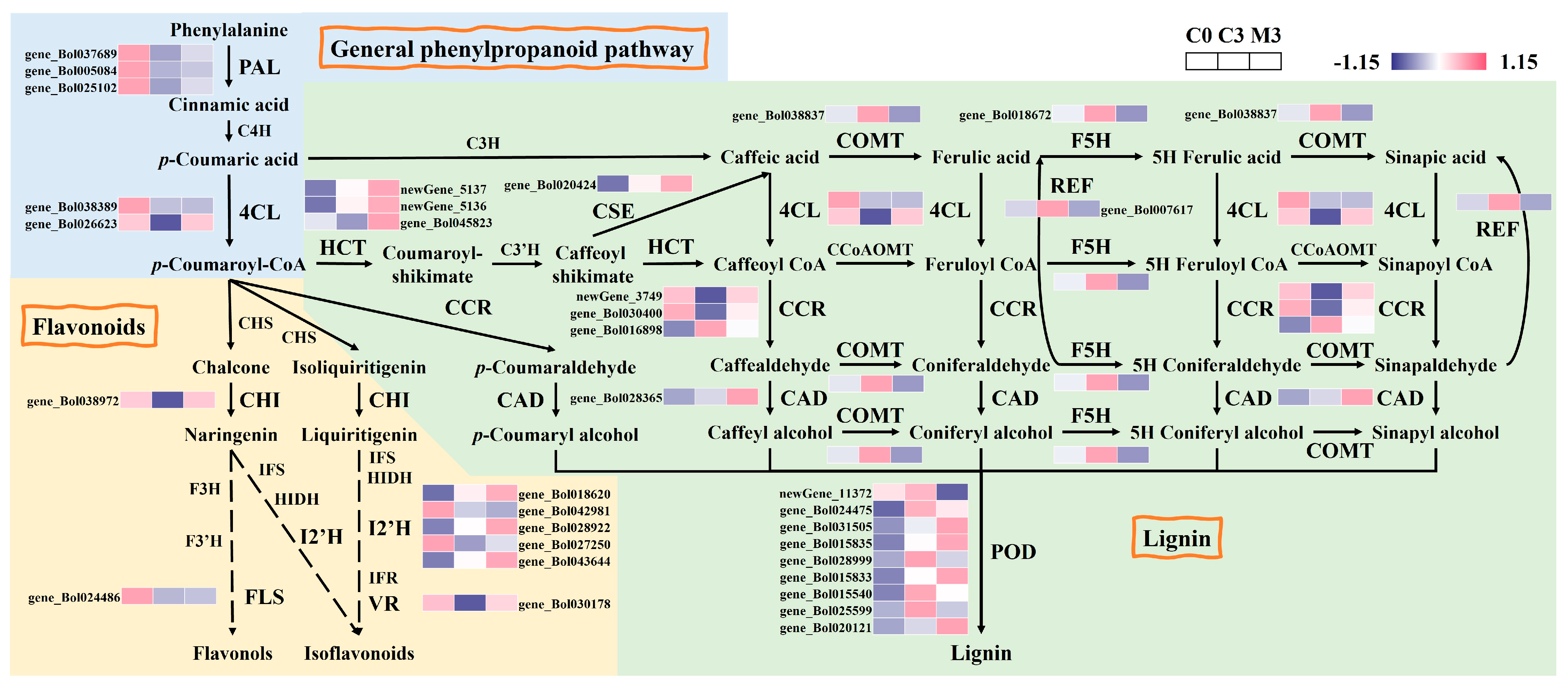
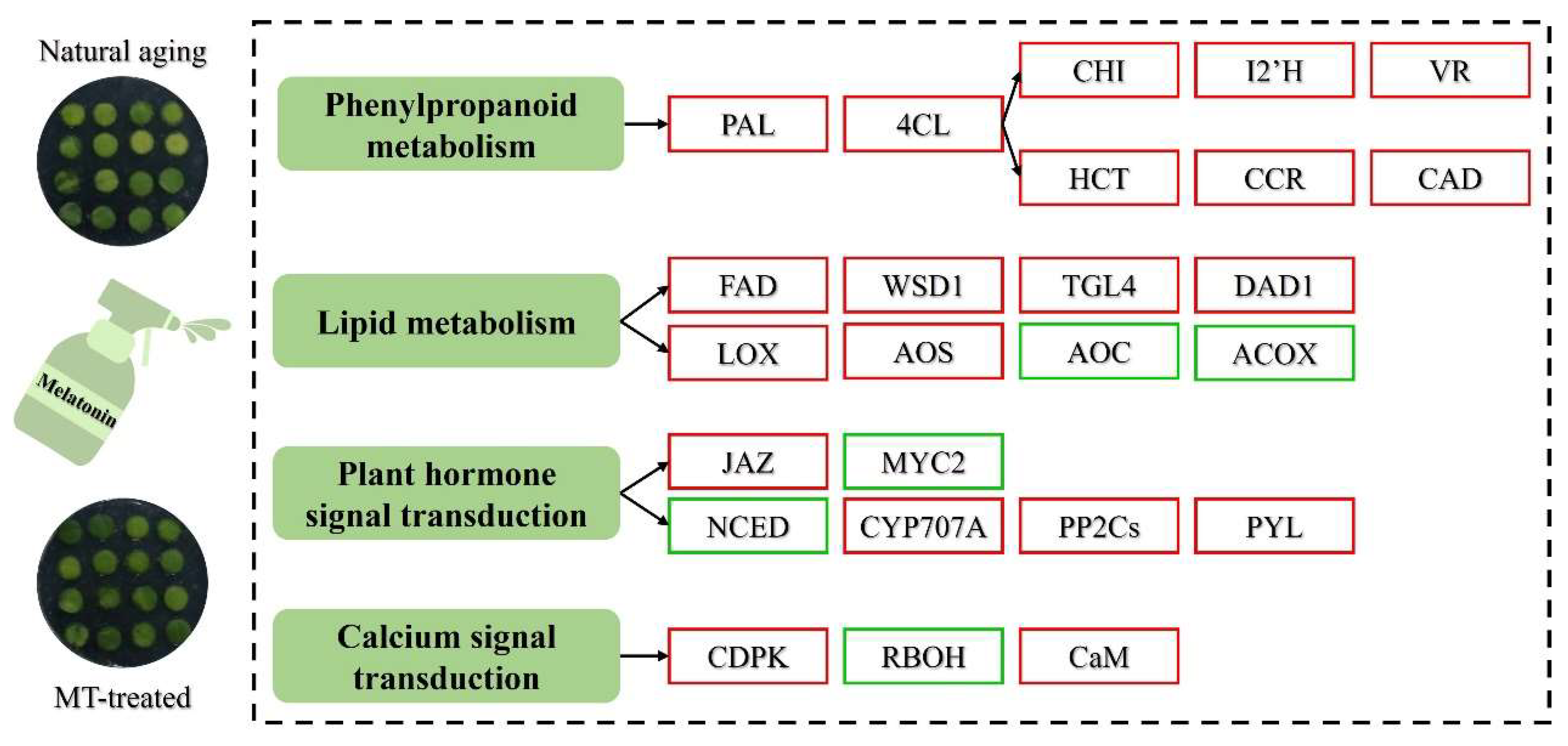
2.5. Melatonin Treatment Activated the Phenylpropanoid Biosynthesis in Postharvest Chinese Kale
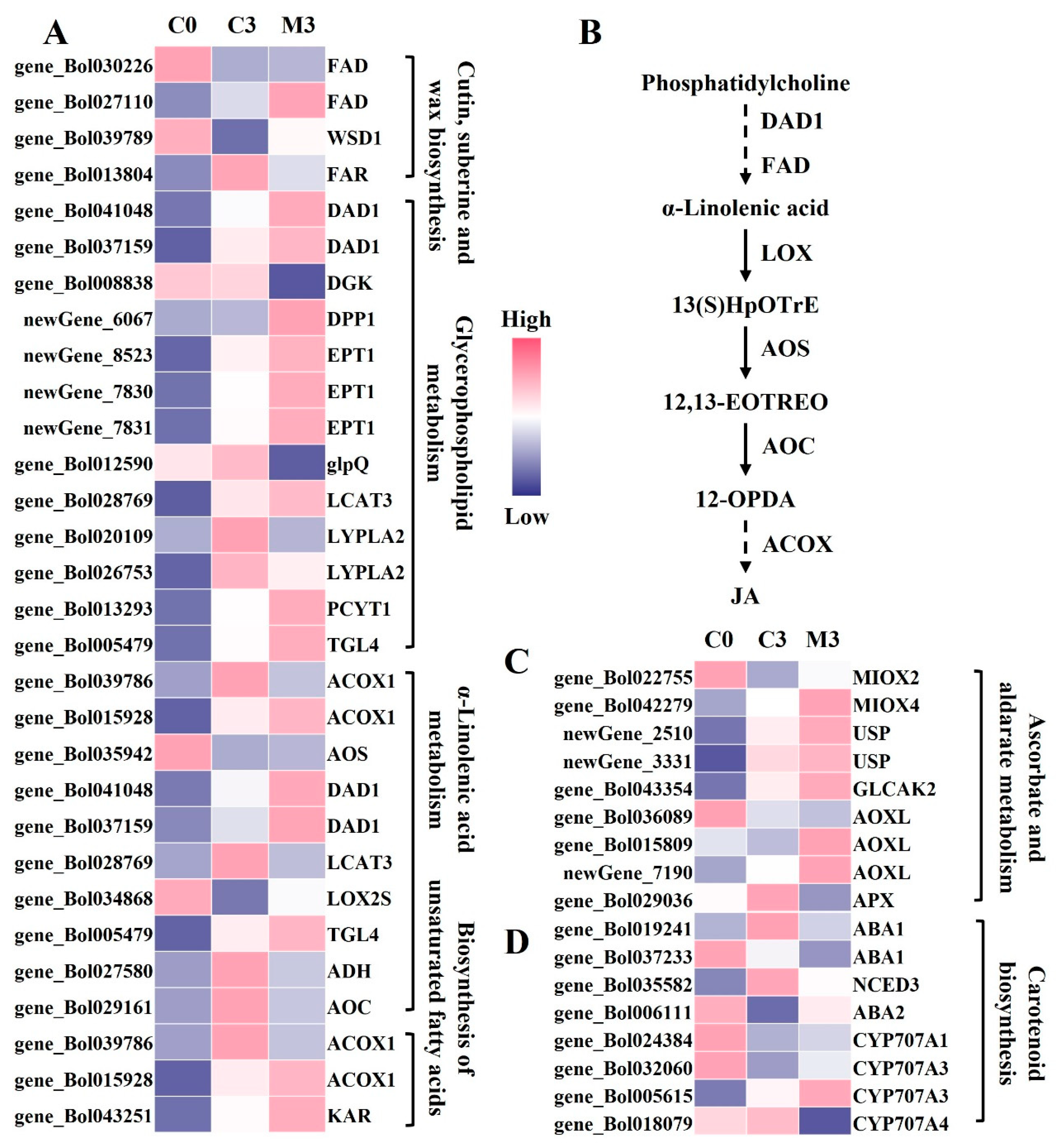
2.6. Melatonin Treatment Induced the Lipid Metabolism in Postharvest Chinese Kale
2.7. Melatonin Treatment Influenced Other Metabolic Pathways in Postharvest Chinese Kale
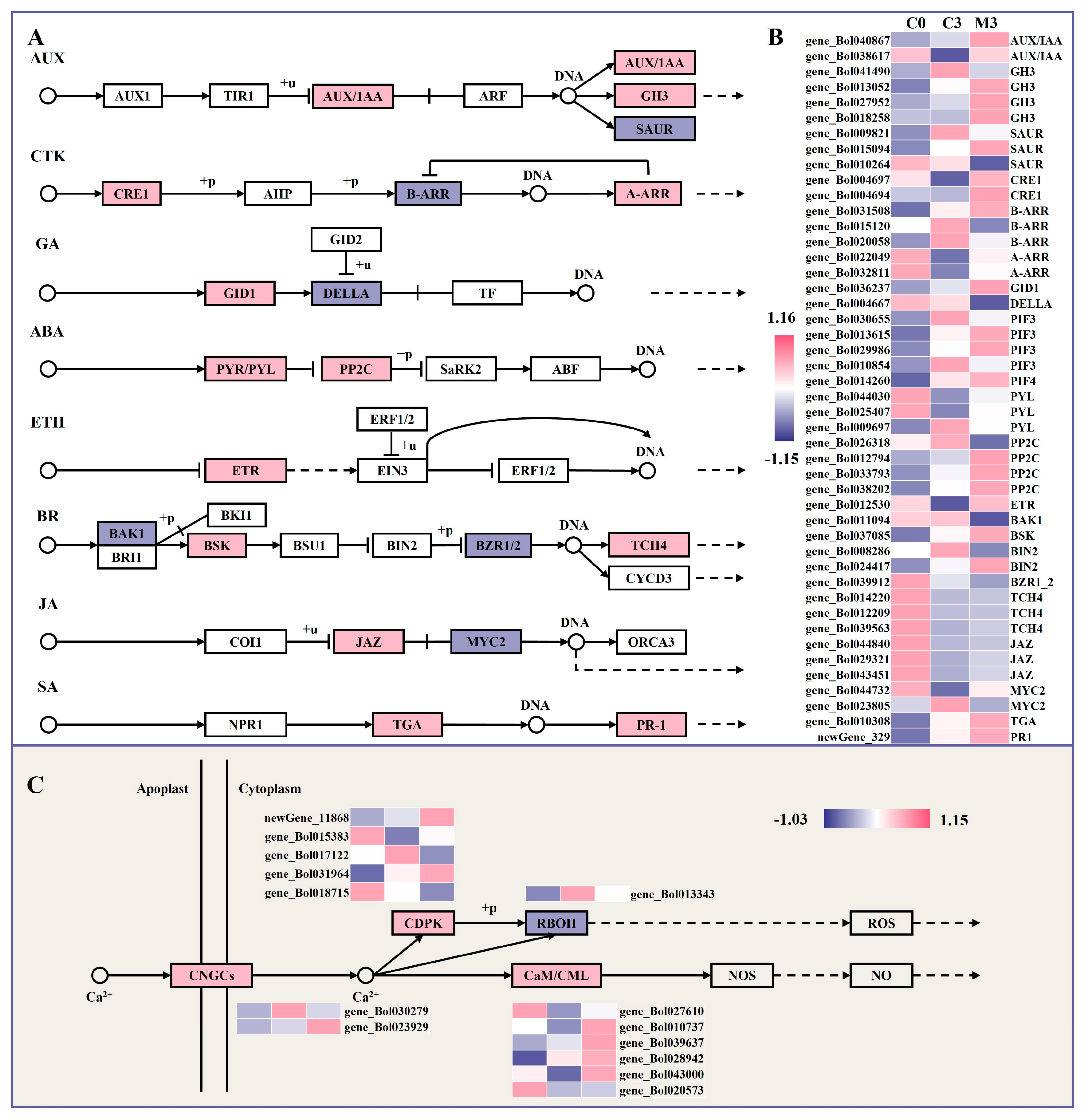
2.8. Melatonin Treatment Induced Plant Hormone Signal Transduction in Postharvest Chinese Kale
2.9. Melatonin Treatment Enhanced Calcium Signal Transduction in Postharvest Chinese Kale
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Melatonin Treatments
4.2. Surface Color Measurement
4.3. Chlorophyll and Carotenoid Content
4.4. Transcriptome Sequencing
4.5. qRT-PCR Validation
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, B.; Jiang, M.; Zheng, H.; Jian, Y.; Huang, W.L.; Yuan, Q.; Zheng, A.H.; Chen, Q.; Zhang, Y.T.; Lin, Y.X.; et al. Color-related chlorophyll and carotenoid concentrations of Chinese kale can be altered through CRISPR/Cas9 targeted editing of the carotenoid isomerase gene BoaCRTISO. Hortic. Res. 2020, 7, 161. [Google Scholar] [CrossRef]
- Wang, J.S.; Yu, H.F.; Zhao, Z.Q.; Sheng, X.G.; Shen, Y.S.; Gu, H.H. Natural variation of glucosinolates and their breakdown products in broccoli (Brassica oleracea var. italica) seeds. J. Agric. Food Chem. 2019, 67, 12528–12537. [Google Scholar] [CrossRef]
- Xiao, Z.L.; Leater, G.E.; Luo, Y.G.; Wang, Q. Assessment of vitamin and carotenoid concentrations of emerging food products: Edible microgreens. J. Agric. Food Chem. 2012, 60, 7644–7651. [Google Scholar] [CrossRef]
- Wang, L.; Wu, S.; Huang, H.; Chen, F.; Ye, M.; Yin, J.; Luo, Z.; Qi, Y.; Chen, M.; Chen, Y. High oxygen atmospheric packaging treatment regulates the postharvest changes of Chinese kale (Brassica oleracea var. alboglabra) during storage. J. Food Sci. 2021, 86, 3884–3895. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Yan, H.; Liu, N.; Wei, J.; Wang, Q. Effect of 1-MCP treatment on postharvest quality characters, antioxidants and glucosinolates of Chinese kale. Food Chem. 2012, 131, 519–526. [Google Scholar] [CrossRef]
- Noichinda, S.; Bodhipadma, K.; Mahamontri, C.; Narongruk, T.; Ketsa, S. Light during storage prevents loss of ascorbic acid, and increases glucose and fructose levels in Chinese kale (Brassica oleracea var. alboglabra). Postharvest Biol. Technol. 2007, 44, 312–315. [Google Scholar] [CrossRef]
- Tada, Y.; Kawano, R.; Komatsubara, S.; Nishimura, H.; Katsuhara, M.; Ozaki, S.; Terashima, S.; Yano, K.; Endo, C.; Sato, M.; et al. Functional screening of salt tolerance genes from a halophyte Sporobolus virginicus and transcriptomic and metabolomic analysis of salt tolerant plants expressing glycine-rich RNA-binding protein. Plant Sci. 2019, 278, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A new plant hormone and/or a plant master regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.L.; Zhang, Y.; Cheng, Y.X.; Tian, Y.L.; Luo, J.J.; Hu, Z.L.; Chen, G.P. The role of melatonin in tomato stress response, growth and development. Plant Cell Rep. 2022, 41, 1631–1650. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.L.; Fan, Z.Q.; Kuang, J.F.; Lu, W.J.; Reiter, R.J.; Lakshmanan, P.; Su, X.G.; Chen, J.Y.; Shan, W. Melatonin delays leaf senescence of Chinese flowering cabbage by suppressing ABFs-mediated abscisic acid biosynthesis and chlorophyll degradation. J. Pineal Res. 2019, 67, e12570. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Protective effect of melatonin against chlorophyll degradation during the senescence of barley leaves. J. Pineal Res. 2009, 46, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.S.; Li, Y.T.; Zhu, J.Z.; Li, P.X. Integrative analysis of transcriptome reveals the possible mechanism of delayed leaf senescence in pak choi (Brassica rapa subsp. chinensis) following melatonin treatment. Food Qual. Saf. 2022, 7, fyac064. [Google Scholar] [CrossRef]
- Liu, X.; An, R.; Li, G.; Luo, S.; Hu, H.; Li, P. Melatonin delays leaf senescence in pak choi (Brassica rapa subsp. chinensis) by regulating biosynthesis of the second messenger cGMP. Hortic. Plant J. 2023, 10, 145–155. [Google Scholar]
- Wei, L.Y.; Liu, C.H.; Zheng, H.H.; Zheng, L. Melatonin treatment affects the glucoraphanin-sulforaphane system in postharvest fresh-cut broccoli (Brassica oleracea L.). Food Chem. 2020, 307, 125562. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Cao, S.F.; Xie, K.Q.; Chi, Z.Y.; Wang, J.; Wang, H.F.; Wei, Y.Y.; Shao, X.F.; Zhang, C.D.; Xu, F.; et al. Melatonin delays yellowing of broccoli during storage by regulating chlorophyll catabolism and maintaining chloroplast ultrastructure. Postharvest Biol. Technol. 2021, 172, 111378. [Google Scholar] [CrossRef]
- Miao, H.Y.; Zeng, W.; Zhao, M.; Wang, J.S.; Wang, Q.M. Effect of melatonin treatment on visual quality and health-promoting properties of broccoli florets under room temperature. Food Chem. 2020, 319, 126498. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, M.; Li, H.; Liu, Y.; Fan, S.; Zhang, N.; Guo, Y.D. Strategies and prospects for melatonin to alleviate abiotic stress in horticultural plants. Hortic. Plant J. 2023. [Google Scholar] [CrossRef]
- Wang, Z.; Pu, H.; Shan, S.; Zhang, P.; Li, J.; Song, H.; Xu, X. Melatonin enhanced chilling tolerance and alleviated peel browning of banana fruit under low temperature storage. Postharvest Biol. Technol. 2021, 179, 111571. [Google Scholar] [CrossRef]
- Li, C.T.; Suo, J.W.; Xuan, L.L.; Ding, M.Z.; Zhang, H.; Song, L.L.; Ying, Y.Q. Bamboo shoot-lignification delay by melatonin during low temperature storage. Postharvest Biol. Technol. 2019, 156, 110933. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, X.P.; Yang, Q.Z.; Zhao, Q.F. Exogenous melatonin delays postharvest fruit senescence and maintains the quality of sweet cherries. Food Chem. 2019, 301, 125311. [Google Scholar] [CrossRef]
- Jannatizadeh, A. Exogenous melatonin applying confers chilling tolerance in pomegranate fruit during cold storage. Sci. Hortic. 2019, 246, 544–549. [Google Scholar] [CrossRef]
- Onik, J.C.; Wai, S.C.; Li, A.; Lin, Q.; Sun, Q.Q.; Wang, Z.D.; Duan, Y.Q. Melatonin treatment reduces ethylene production and maintains fruit quality in apple during postharvest storage. Food Chem. 2021, 337, 127753. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, G.; Huang, Z.; Duan, W.; Tan, H.; Li, Y.; Hou, X. Temperature expression patterns of genes and their coexpression with LncRNAs revealed by RNA-Seq in non-heading Chinese cabbage. BMC Genom. 2016, 17, 297. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiang, H.; Gao, M.; He, R.; Liu, X.; Su, W.; Liu, H. Far-red-light-induced morphology changes, phytohormone, and transcriptome reprogramming of Chinese kale (Brassica alboglabra Bailey). Int. J. Mol. Sci. 2023, 24, 5563. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, J.; Mao, S.; Xu, H.; Wu, Q.; Liang, M.; Yuan, Y.; Liu, M.; Huang, K. Comparative transcriptome analyses of genes involved in sulforaphane metabolism at different treatment in Chinese kale using full-length transcriptome sequencing. BMC Genom. 2019, 20, 377. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant-environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.T.; Liao, Y.X.; Kan, J.Q.; Han, L.; Zheng, Y.H. Response of direct or priming defense against Botrytis cinerea to methyl jasmonate treatment at different concentrations in grape berries. Int. J. Food Microbiol. 2015, 194, 32–39. [Google Scholar] [CrossRef]
- van Tunen, A.J.; Mur, L.A.; Recourt, K.; Gerats, A.G.; Mol, J.N. Regulation and manipulation of flavonoid gene expression in anthers of petunia: The molecular basis of the Po mutation. Plant Cell 1991, 3, 39–48. [Google Scholar]
- Mottiar, Y.; Vanholme, R.; Boerjan, W.; Ralph, J.; Mansfield, S.D. Designer lignins: Harnessing the plasticity of lignification. Curr. Opin. Biotechnol. 2016, 37, 190–200. [Google Scholar] [CrossRef]
- Yan, H.L.; Chen, J.J.; Liu, J. The involvement of energy metabolism and lipid peroxidation in lignin accumulation of postharvest pumelos. Membranes 2020, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.N.; Li, B.J.; Su, G.Q.; Zhang, M.X.; Grierson, D.; Chen, K.S. Transcriptional regulation of fleshy fruit texture. J. Integr. Plant Biol. 2022, 64, 1649–1672. [Google Scholar] [CrossRef]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, L.; Wang, Z.; Zhang, R.; Liu, P.; Liu, M.; Liu, Z.; Zhao, Z.; Wang, L.; Chen, X.; et al. The regulation of cell wall lignification and lignin biosynthesis during pigmentation of winter jujube. Hortic. Res. 2021, 8, 238. [Google Scholar] [CrossRef]
- Li, S.E.; Cheng, Y.; Yan, R.; Liu, Y.; Huan, C.; Zheng, X.L. Preharvest spray with melatonin improves postharvest disease resistance in cherry tomato fruit. Postharvest Biol. Technol. 2022, 193, 112055. [Google Scholar] [CrossRef]
- Li, S.E.; Xu, Y.H.; Bi, Y.; Zheng, B.; Shen, S.L.; Jiang, T.J.; Zheng, X.L. Melatonin treatment inhibits gray mold and induces disease resistance in cherry tomato fruit during postharvest. Postharvest Biol. Technol. 2019, 157, 110962. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Wang, T.; Liu, G.S.; Hu, M.J.; Yun, Z.; Duan, X.W.; Cai, K.; Jiang, G.X. Inhibition of downy blight and enhancement of resistance in litchi fruit by postharvest application of melatonin. Food Chem. 2021, 347, 129009. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.C.; Han, P.; Bai, S.Q.; Ma, N.; Fang, D.H.; Yang, W.J.; Hu, Q.H.; Pei, F. Transcriptome analysis reveals the mechanism of caffeic acid-grafted-chitosan/polylactic acid film packaging to delay quality deterioration in postharvest Agaricus bisporus. Sci. Hortic. 2023, 309, 111647. [Google Scholar] [CrossRef]
- Bohinc, T.; Markovič, D.; Trdan, S. Leaf epicuticular wax as a factor of antixenotic resistance of cabbage to cabbage flea beetles and cabbage stink bugs attack. Acta Agric. Scand. 2014, 64, 493–500. [Google Scholar] [CrossRef]
- Segado, P.; Domínguez, E.; Heredia, A. Ultrastructure of the epidermal cell wall and cuticle of tomato fruit (Solanum lycopersicum L.) during development. Plant Physiol. 2016, 170, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wu, X.; Lam, P.; Bird, D.; Zheng, H.; Samuels, L.; Jetter, R.; Kunst, L. Identification of the wax ester synthase/acyl-coenzyme A: Diacylglycerol acyltransferase WSD1 required for stem wax ester biosynthesis in Arabidopsis. Plant Physiol. 2008, 148, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Bi, Y.; Zhang, X.; Han, Z.; Zong, Y.; Li, Y.; Sionov, E.; Prusky, D. Benzothiadiazole treatment inhibits membrane lipid metabolism and straight-chain volatile compound release in Penicillium expansum-inoculated apple fruit. Postharvest Biol. Technol. 2021, 181, 111671. [Google Scholar] [CrossRef]
- Min, T.; Niu, L.F.; Feng, X.Y.; Yi, Y.; Wang, L.M.; Zhao, Y.; Wang, H.X. The effects of different temperatures on the storage characteristics of lotus (Nelumbo nucifera G.) root. Food Chem. 2021, 348, 129109. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Chen, Y.; Shi, Z.J.; Jin, Y.F.; Sun, H.S.; Xie, F.C.; Zhang, L. Biosynthesis and signal transduction of ABA, JA, and BRs in response to drought stress of Kentucky bluegrass. Int. J. Mol. Sci. 2019, 20, 1289. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Yang, M.; Wu, X.; Wang, Y.; Zhang, R. Physiological and transcriptomic analyses of the effects of exogenous melatonin on drought tolerance in maize (Zea mays L.). Plant Physiol. Biochem. 2021, 168, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Schaller, A.; Stintzi, A. Enzymes in jasmonate biosynthesis-structure, function, regulation. Phytochemistry 2009, 70, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.C.; Fu, Q.S.; Zheng, J.; Zhang, A.A.; Wang, H.S. Transcriptomic and metabolomic analyses reveal that melatonin promotes melon root development under copper stress by inhibiting jasmonic acid biosynthesis. Hortic. Res. 2020, 7, 79. [Google Scholar] [CrossRef]
- Li, C.; Tan, D.X.; Liang, D.; Chang, C.; Jia, D.; Ma, F. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 2015, 66, 669–680. [Google Scholar] [CrossRef]
- Wang, D.; Chen, Q.Y.; Chen, W.W.; Guo, Q.G.; Xia, Y.; Wang, S.M.; Jing, D.L.; Liang, G.L. Physiological and transcription analyses reveal the regulatory mechanism of melatonin in inducing drought resistance in loquat (Eriobotrya japonica Lindl.) seedlings. Environ. Exp. Bot. 2021, 181, 104291. [Google Scholar] [CrossRef]
- Duszyn, M.; Swiezawska, B.; Szmidt-Jaworska, A.; Jaworski, K. Cyclic nucleotide gated channels (CNGCs) in plant signalling-current knowledge and perspectives. J. Plant Physiol. 2019, 241, 153035. [Google Scholar] [CrossRef]
- Stefano, M.D.; Vandelle, E.; Polverari, A.; Ferrarini, A.; Delledonne, M. Nitric oxide-mediated signaling functions during the plant hypersensitive response. In Nitric Oxide in Plant Growth, Development and Stress Physiology; Springer: Berlin/Heidelberg, Germany, 2007; pp. 207–222. [Google Scholar]
- Sun, B.; Di, H.M.; Zhang, J.Q.; Xia, P.X.; Huang, W.L.; Jian, Y.; Zhang, C.L.; Zhang, F. Effect of light on sensory quality, health-promoting phytochemicals and antioxidant capacity in post-harvest baby mustard. Food Chem. 2021, 339, 128057. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L.P. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di, H.; Zhang, C.; Zhou, A.; Huang, H.; Tang, Y.; Li, H.; Huang, Z.; Zhang, F.; Sun, B. Transcriptome Analysis Reveals the Mechanism by Which Exogenous Melatonin Treatment Delays Leaf Senescence of Postharvest Chinese Kale (Brassica oleracea var. alboglabra). Int. J. Mol. Sci. 2024, 25, 2250. https://doi.org/10.3390/ijms25042250
Di H, Zhang C, Zhou A, Huang H, Tang Y, Li H, Huang Z, Zhang F, Sun B. Transcriptome Analysis Reveals the Mechanism by Which Exogenous Melatonin Treatment Delays Leaf Senescence of Postharvest Chinese Kale (Brassica oleracea var. alboglabra). International Journal of Molecular Sciences. 2024; 25(4):2250. https://doi.org/10.3390/ijms25042250
Chicago/Turabian StyleDi, Hongmei, Chenlu Zhang, Aolian Zhou, Huanhuan Huang, Yi Tang, Huanxiu Li, Zhi Huang, Fen Zhang, and Bo Sun. 2024. "Transcriptome Analysis Reveals the Mechanism by Which Exogenous Melatonin Treatment Delays Leaf Senescence of Postharvest Chinese Kale (Brassica oleracea var. alboglabra)" International Journal of Molecular Sciences 25, no. 4: 2250. https://doi.org/10.3390/ijms25042250
APA StyleDi, H., Zhang, C., Zhou, A., Huang, H., Tang, Y., Li, H., Huang, Z., Zhang, F., & Sun, B. (2024). Transcriptome Analysis Reveals the Mechanism by Which Exogenous Melatonin Treatment Delays Leaf Senescence of Postharvest Chinese Kale (Brassica oleracea var. alboglabra). International Journal of Molecular Sciences, 25(4), 2250. https://doi.org/10.3390/ijms25042250