
A Pharmacological Investigation of the TMEM16A Currents in Murine Skeletal Myogenic Precursor Cells

 , ,
, , 
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
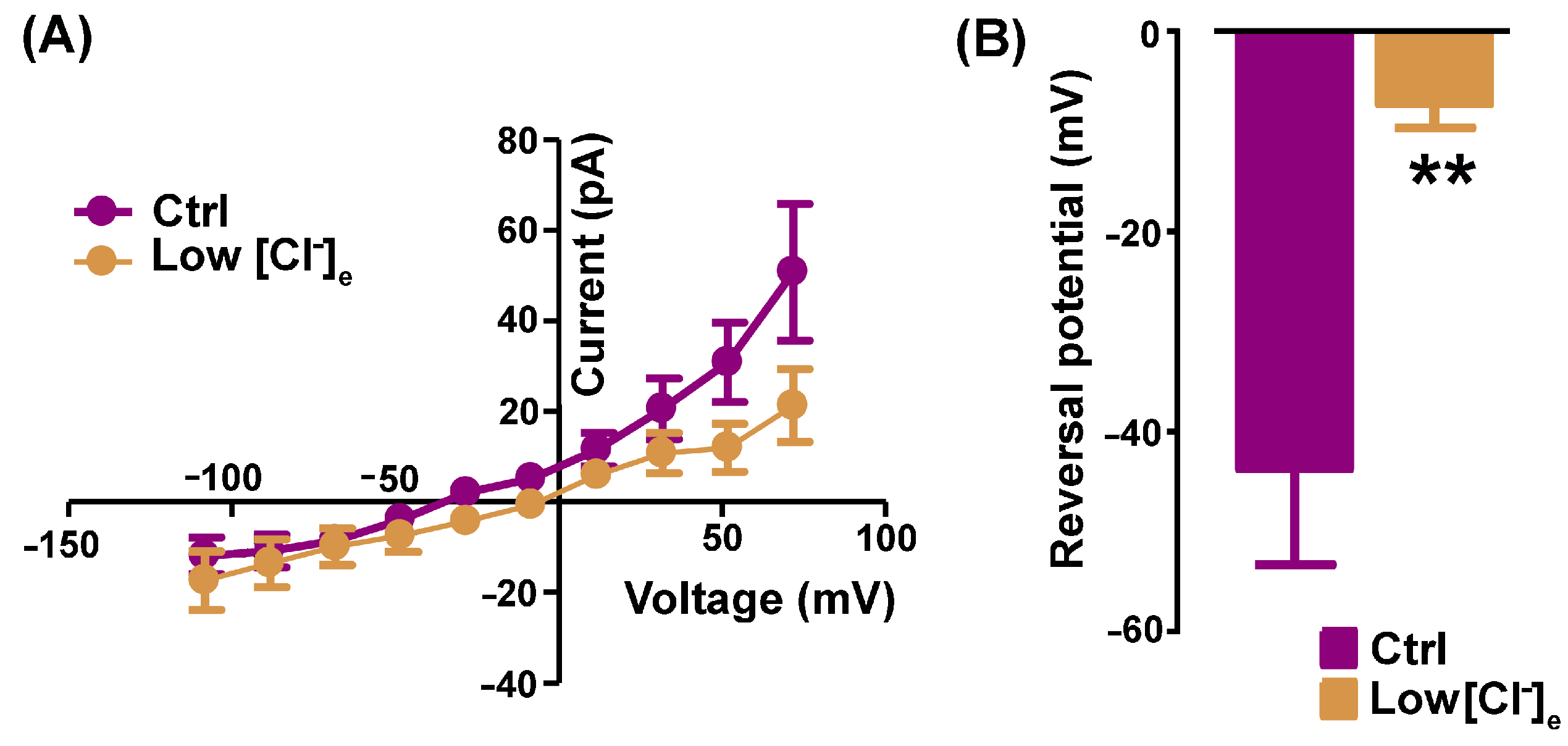
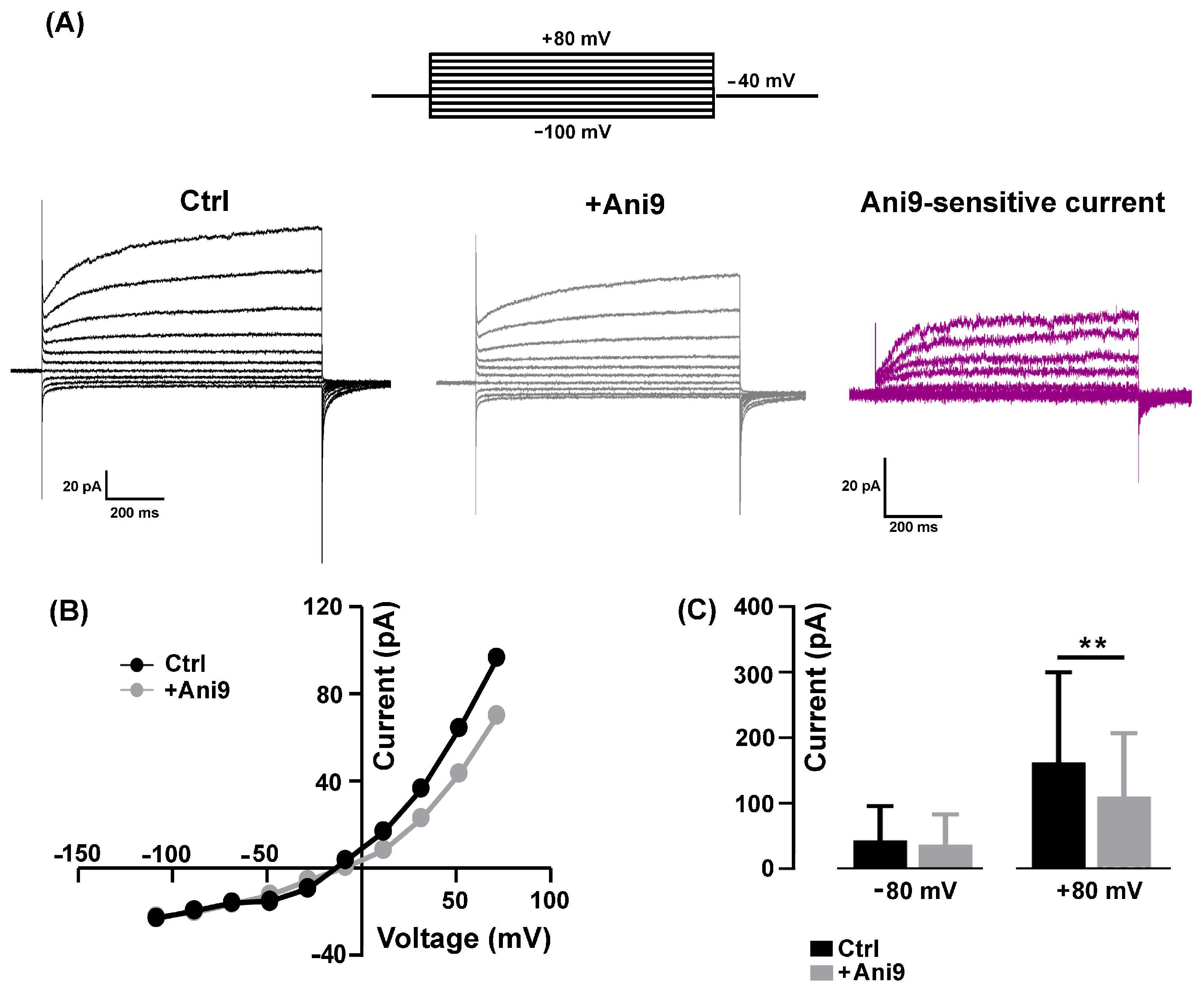
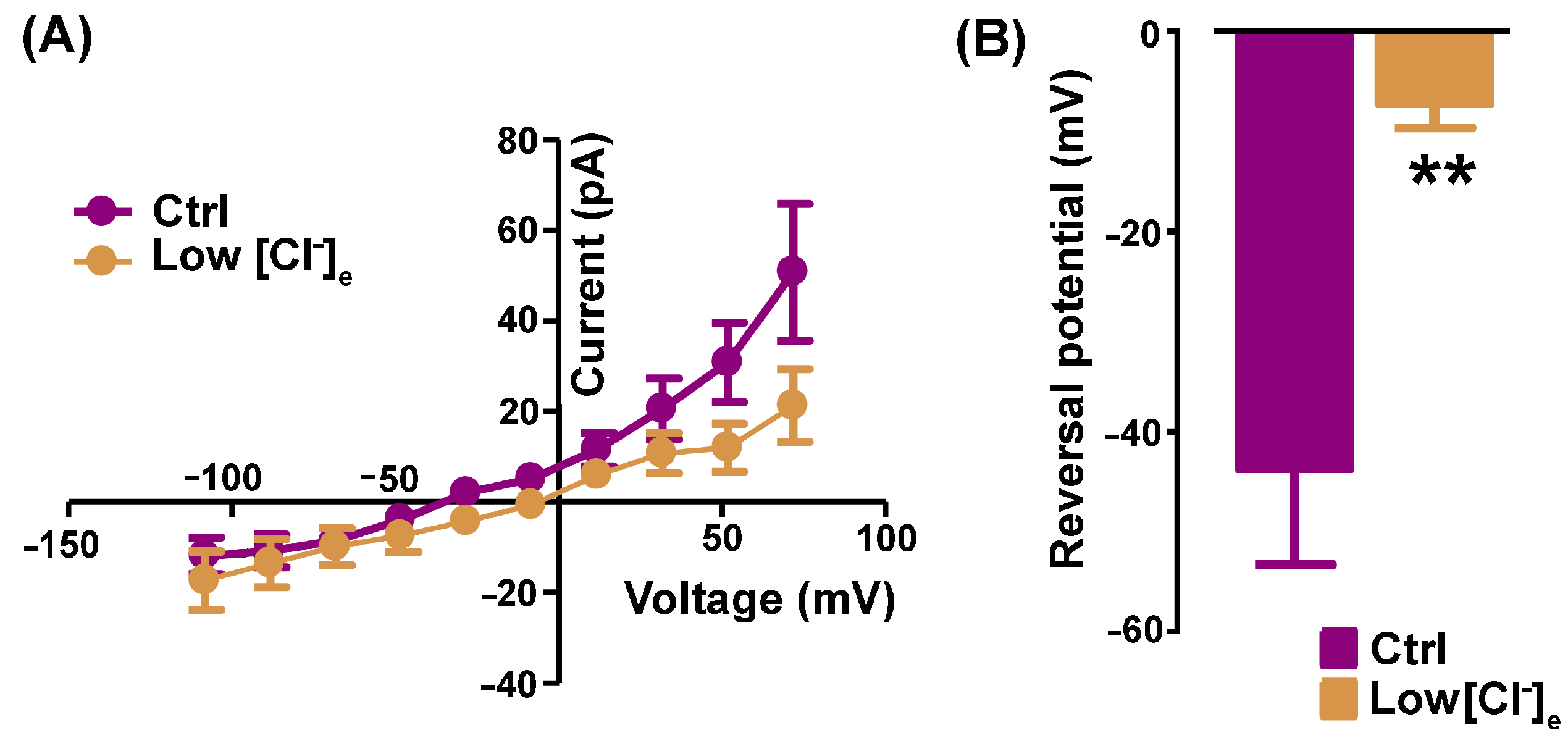
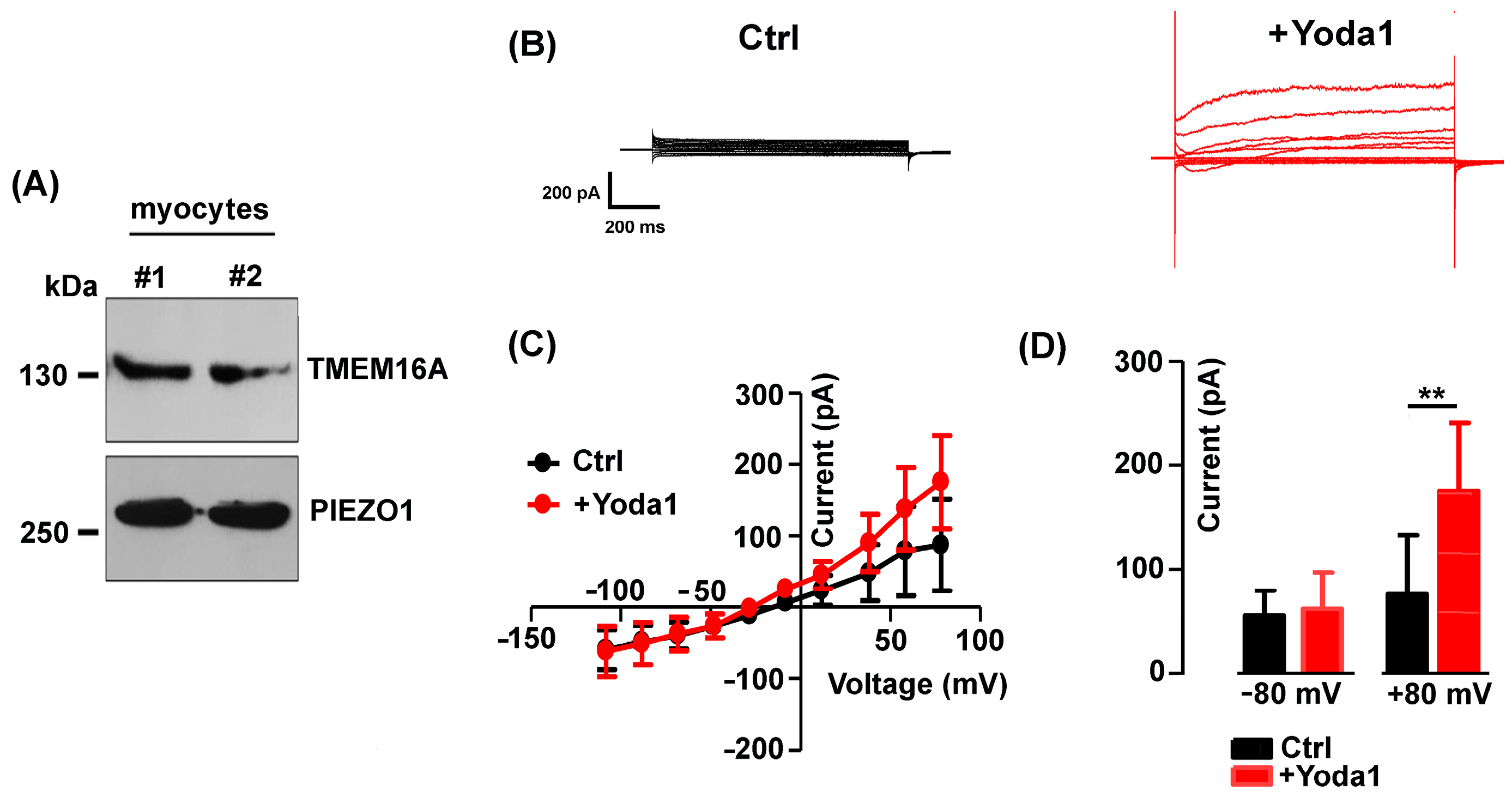
2.1. Pharmacological Identification of TMEM16A Currents in Mouse Skeletal Myocytes
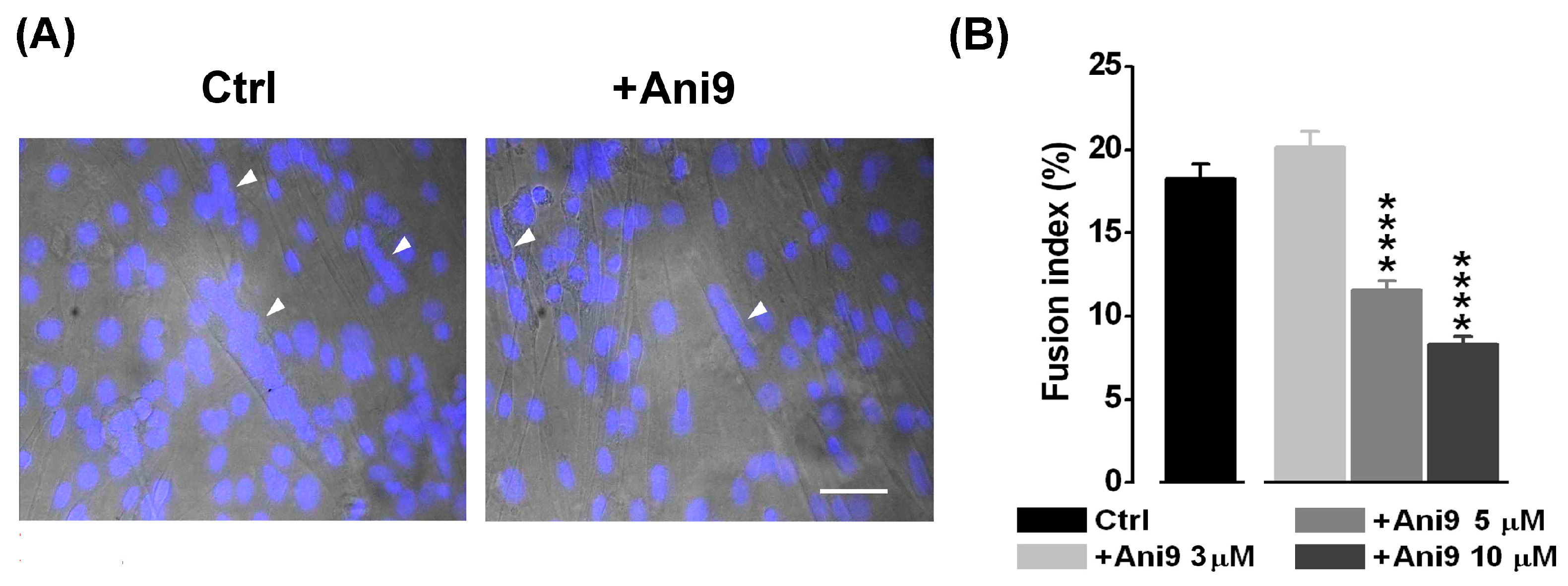
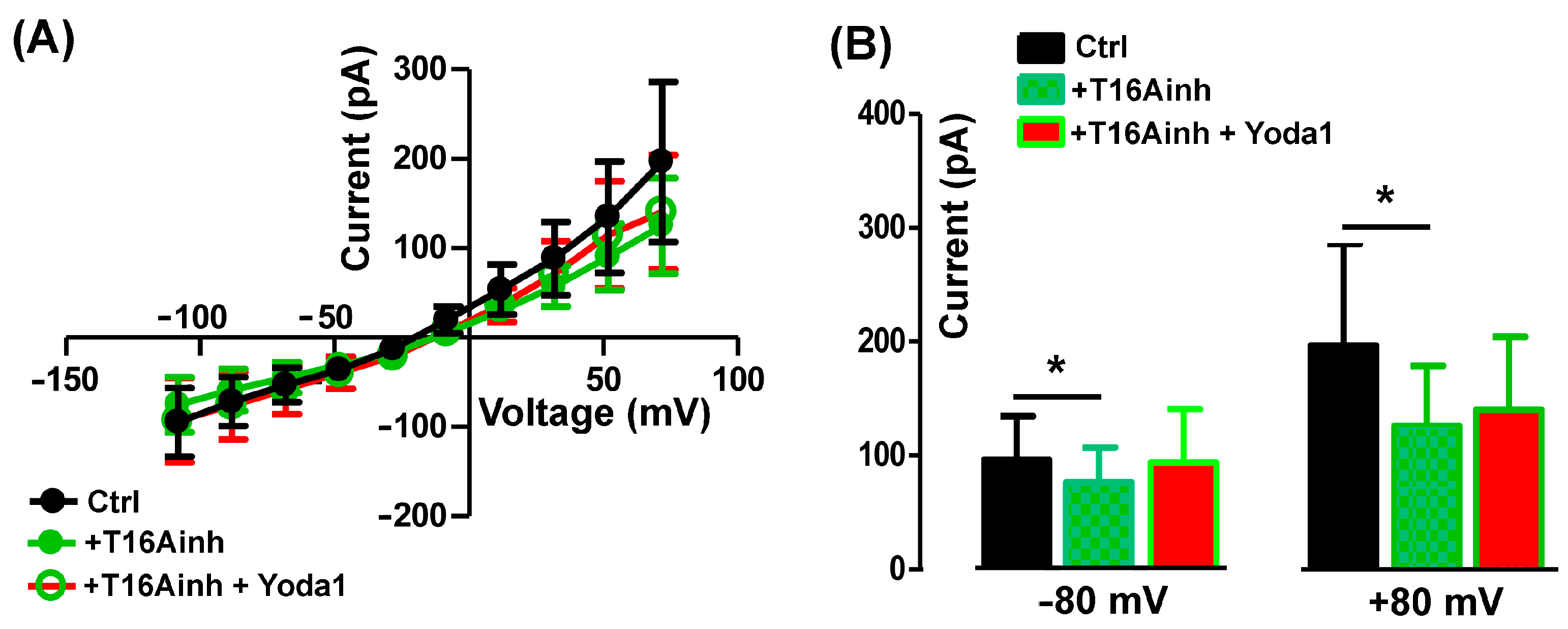
2.2. The Blockage of TMEM16A Currents Reduces Myoblast Fusion
2.3. The TMEM16A Currents Are Modulated by the Piezo1 Agonist Yoda1
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Electrophysiological Recordings
4.3. Western Blotting
4.4. Fusion Index
4.5. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef]
- Fennelly, C.; Soker, S. Bioelectric Properties of Myogenic Progenitor Cells. Bioelectricity 2019, 1, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Miledi, R. A calcium-dependent transient outward current in Xenopus laevis oocytes. Proc. R. Soc. London Ser. B Biol. Sci. 1982, 215, 491–497. [Google Scholar] [CrossRef]
- Barish, M.E. A transient calcium-dependent chloride current in the immature Xenopus oocyte. J. Physiol. 1983, 342, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.A.; Cozens, A.L.; Schulman, H.; Gruenert, D.C.; Stryer, L.; Gardner, P. Activation of chloride channels in normal and cystic fibrosis airway epithelial cells by multifunctional calcium/calmodulin-dependent protein kinase. Nature 1991, 349, 793–796. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Shah, S.; Liu, Y.; Zhang, H.; Lees, M.; Fu, Z.; Lippiat, J.D.; Beech, D.J.; Sivaprasadarao, A.; Baldwin, S.A.; et al. Activation of the Cl− channel ANO1 by localized calcium signals in nociceptive sensory neurons requires coupling with the IP3 receptor. Sci. Signal. 2013, 6, ra73. [Google Scholar] [CrossRef]
- Large, W.A.; Wang, Q. Characteristics and physiological role of the Ca(2+)-activated Cl− conductance in smooth muscle. Am. J. Physiol. 1996, 271, C435–C454. [Google Scholar] [CrossRef] [PubMed]
- Frings, S.; Reuter, D.; Kleene, S.J. Neuronal Ca2+-activated Cl− channels–homing in on an elusive channel species. Prog. Neurobiol. 2000, 60, 247–289. [Google Scholar] [CrossRef]
- Dauner, S.; Lissmann, J.; Jeridi, S.; Frings, S.; Mohrlen, F. Expression patterns of anoctamin 1 and anoctamin 2 chloride channels in the mammalian nose. Cell Tissue Res. 2012, 347, 327–341. [Google Scholar] [CrossRef]
- Maurya, D.K.; Menini, A. Developmental expression of the calcium-activated chloride channels TMEM16A and TMEM16B in the mouse olfactory epithelium. Dev. Neurobiol. 2014, 74, 657–675. [Google Scholar] [CrossRef]
- Ferrera, L.; Zegarra-Moran, O.; Galietta, L.J. Ca2+-activated Cl− channels. Compr. Physiol. 2011, 1, 2155–2174. [Google Scholar] [CrossRef]
- Steinmeyer, K.; Ortland, C.; Jentsch, T.J. Primary structure and functional expression of a developmentally regulated skeletal muscle chloride channel. Nature 1991, 354, 301–304. [Google Scholar] [CrossRef]
- Bretag, A.H. Muscle chloride channels. Physiol. Rev. 1987, 67, 618–724. [Google Scholar] [CrossRef]
- Bardouille, C.; Vullhorst, D.; Jockusch, H. Expression of chloride channel 1 mRNA in cultured myogenic cells: A marker of myotube maturation. FEBS Lett. 1996, 396, 177–180. [Google Scholar] [CrossRef]
- Pedemonte, N.; Galietta, L.J. Structure and function of TMEM16 proteins (anoctamins). Physiol. Rev. 2014, 94, 419–459. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.C.; Cheng, T.; Jan, Y.N.; Jan, L.Y. Expression cloning of TMEM16A as a calcium-activated chloride channel subunit. Cell 2008, 134, 1019–1029. [Google Scholar] [CrossRef]
- Caputo, A.; Caci, E.; Ferrera, L.; Pedemonte, N.; Barsanti, C.; Sondo, E.; Pfeffer, U.; Ravazzolo, R.; Zegarra-Moran, O.; Galietta, L.J. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science 2008, 322, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.D.; Cho, H.; Koo, J.Y.; Tak, M.H.; Cho, Y.; Shim, W.S.; Park, S.P.; Lee, J.; Lee, B.; Kim, B.M.; et al. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 2008, 455, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Bradley, E.; Fedigan, S.; Webb, T.; Hollywood, M.A.; Thornbury, K.D.; McHale, N.G.; Sergeant, G.P. Pharmacological characterization of TMEM16A currents. Channels 2014, 8, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Hume, R.I.; Thomas, S.A. A calcium- and voltage-dependent chloride current in developing chick skeletal muscle. J. Physiol. 1989, 417, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Dayal, A.; Ng, S.F.J.; Grabner, M. Ca2+-activated Cl− channel TMEM16A/ANO1 identified in zebrafish skeletal muscle is crucial for action potential acceleration. Nat. Commun. 2019, 10, 115. [Google Scholar] [CrossRef]
- Yuan, W.; Cui, C.C.; Li, J.; Xu, Y.H.; Fan, C.E.; Chen, Y.C.; Fan, H.W.; Hu, B.X.; Shi, M.Y.; Sun, Z.Y.; et al. Intracellular TMEM16A is necessary for myogenesis of skeletal muscle. iScience 2022, 25, 105446. [Google Scholar] [CrossRef]
- Bosutti, A.; Giniatullin, A.; Odnoshivkina, Y.; Giudice, L.; Malm, T.; Sciancalepore, M.; Giniatullin, R.; D’Andrea, P.; Lorenzon, P.; Bernareggi, A. “Time window” effect of Yoda1-evoked Piezo1 channel activity during mouse skeletal muscle differentiation. Acta Physiol. 2021, 233, e13702. [Google Scholar] [CrossRef]
- Bernareggi, A.; Bosutti, A.; Massaria, G.; Giniatullin, R.; Malm, T.; Sciancalepore, M.; Lorenzon, P. The State of the Art of Piezo1 Channels in Skeletal Muscle Regeneration. Int. J. Mol. Sci. 2022, 23, 6616. [Google Scholar] [CrossRef]
- Sciancalepore, M.; Massaria, G.; Tramer, F.; Zacchi, P.; Lorenzon, P.; Bernareggi, A. A preliminary study on the role of Piezo1 channels in myokine release from cultured mouse myotubes. Biochem. Biophys. Res. Commun. 2022, 623, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Cahalan, S.M.; Lukacs, V.; Ranade, S.S.; Chien, S.; Bandell, M.; Patapoutian, A. Piezo1 links mechanical forces to red blood cell volume. eLife 2015, 4, e07370. [Google Scholar] [CrossRef] [PubMed]
- Gnanasambandam, R.; Bae, C.; Gottlieb, P.A.; Sachs, F. Ionic Selectivity and Permeation Properties of Human PIEZO1 Channels. PLoS ONE 2015, 10, e0125503. [Google Scholar] [CrossRef] [PubMed]
- Vladislav Chubinskiy-Nadezhdin, V.; Semenova, S.; Vasileva, V.; Shatrova, A.; Pugovkina, N.; Negulyaev, Y. Store-Operated Ca2+ Entry Contributes to Piezo1-Induced Ca2+ Increase in Human Endometrial Stem Cells. Int. J. Mol. Sci. 2022, 23, 3763. [Google Scholar] [CrossRef] [PubMed]
- Syeda, R.; Xu, J.; Dubin, A.E.; Coste, B.; Mathur, J.; Huynh, T.; Matzen, J.; Lao, J.; Tully, D.C.; Engels, I.H.; et al. Chemical activation of the mechanotransduction channel Piezo1. eLife 2015, 4, e07369. [Google Scholar] [CrossRef] [PubMed]
- Centeio, R.; Cabrita, I.; Benedetto, R.; Talbi, K.; Ousingsawat, J.; Schreiber, R.; Sullivan, J.K.; Kunzelmann, K. Pharmacological Inhibition and Activation of the Ca2+ Activated Cl− Channel TMEM16A. Int. J. Mol. Sci. 2020, 21, 2557. [Google Scholar] [CrossRef] [PubMed]
- Valdivieso, Á.G.; Santa-Coloma, T.A. The chloride anion as a signalling effector. Biol. Rev. Camb. Philos. Soc. 2019, 94, 1839–1856. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Lee, H.K.; Park, J.; Jeon, D.K.; Jo, S.; Jo, M.; Namkung, W. Ani9, A Novel Potent Small-Molecule ANO1 Inhibitor with Negligible Effect on ANO2. PLoS ONE 2016, 11, e0155771. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Ryu, S.; Sim, K.; Song, C.; Shin, I.; Kim, S.S.; Lee, Y.-S.; Park, J.-Y.; Sim, T. Anti-glioma effects of 2-aminothiophene-3-carboxamide derivatives, ANO1 channel blockers. Eur. J. Med. Chem. 2020, 208, 112688. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, K.L.; Phelps, W.A.; Tembo, M.; Lee, M.T.; Carlson, A.E. The TMEM16A channel mediates the fast polyspermy block in Xenopus laevis. J. Gen. Physiol. 2018, 150, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, Y.; Liang, H. Interactive association of drugs binding to human serum albumin. Int. J. Mol. Sci. 2014, 15, 3580–3595. [Google Scholar] [CrossRef] [PubMed]
- Haroon, M.; Klein-Nulend, J.; Bakker, A.D.; Jin, J.; Seddiqi, H.; Offringa, C.; de Wit, G.M.J.; Le Grand, F.; Giordani, L.; Liu, K.J.; et al. Myofiber stretch induces tensile and shear deformation of muscle stem cells in their native niche. Biophys. J. 2021, 120, 2665–2678. [Google Scholar] [CrossRef]
- Seale, P.; Rudnicki, M.A. A new look at the origin, function, and “stem-cell” status of muscle satellite cells. Dev. Biol. 2000, 218, 115–124. [Google Scholar] [CrossRef]
- Hawke, T.J.; Garry, D.J. Myogenic satellite cells: Physiology to molecular biology. J. Appl. Physiol. 2001, 91, 534–551. [Google Scholar] [CrossRef]
- Botello-Smith, W.M.; Jiang, W.; Zhang, H.; Ozkan, A.D.; Lin, Y.C.; Pham, C.N.; Lacroix, J.J.; Luo, Y. A mechanism for the activation of the mechanosensitive Piezo1 channel by the small molecule Yoda1. Nat. Commun. 2019, 10, 4503. [Google Scholar] [CrossRef]
- Wijerathne, T.D.; Alper, A.D.; Lacroix, J.J. Yoda1’s energetic footprint on Piezo1 channels and its modulation by voltage and temperature. Proc. Natl. Acad. Sci. USA 2022, 119, e2202269119. [Google Scholar] [CrossRef]
- Bernheim, L.; Liu, J.H.; Hamann, M.; Haenggeli, C.A.; Fischer-Lougheed, J.; Bader, C.R. Contribution of a non-inactivating potassium current to the resting membrane potential of fusion-competent human myoblasts. J. Physiol. 1996, 493, 129–141. [Google Scholar] [CrossRef]
- Bijlenga, P.; Occhiodoro, T.; Liu, J.H.; Bader, C.R.; Bernheim, L.; Fischer-Lougheed, J. An ether -à-go-go K+ current, Ih-eag, contributes to the hyperpolarization of human fusion-competent myoblasts. J. Physiol. 1998, 512, 317–323. [Google Scholar] [CrossRef]
- Liu, J.H.; Bijlenga, P.; Fischer-Lougheed, J.; Occhiodoro, T.; Kaelin, A.; Bader, C.R.; Bernheim, L. Role of an inward rectifier K+ current and of hyperpolarization in human myoblast fusion. J. Physiol. 1998, 510, 467–476. [Google Scholar] [CrossRef]
- Chen, L.; König, B.; Stauber, T. LRRC8 channel activation and reduction in cytosolic chloride concentration during early differentiation of C2C12 myoblasts. Biochem. Biophys. Res. Commun. 2020, 532, 482–488. [Google Scholar] [CrossRef]
- Dulhunty, A.F. The dependence of membrane potential on extracellular chloride concentration in mammalian skeletal muscle fibres. J. Physiol. 1978, 276, 67–82. [Google Scholar] [CrossRef]
- Ritchie, A.K.; Fambrough, D.M. Electrophysiological properties of the membrane and acetylcholine receptor in developingrat and chick myotubes. J. Gen. Physiol. 1975, 66, 327–355. [Google Scholar] [CrossRef]
- Spector, I.; Prives, J.M. Development of electrophysiological and biochemical membrane properties during differentiation of embryonic skeletal muscle in culture. Proc. Natl. Acad. Sci. USA 1977, 74, 5166–5170. [Google Scholar] [CrossRef]
- Stauber, T.; Jentsch, T.J. Chloride in vesicular trafficking and function. Annu. Rev. Physiol. 2013, 75, 453–477. [Google Scholar] [CrossRef]
- He, M.; Ye, W.; Wang, W.J.; Sison, E.S.; Jan, Y.N.; Jan, L.Y. Cytoplasmic Cl− couples membrane remodeling to epithelial morphogenesis. Proc. Natl. Acad. Sci. USA 2017, 114, E11161–E11169. [Google Scholar] [CrossRef]
- Perez-Cornejo, P.; Gokhale, A.; Duran, C.; Cui, Y.; Xiao, Q.; Hartzell, H.C.; Faundez, V. Anoctamin 1 (Tmem16A) Ca2+-activated chloride channel stoichiometrically interacts with an ezrin-radixin-moesin network. Proc. Natl. Acad. Sci. USA 2012, 109, 10376–10381. [Google Scholar] [CrossRef]
- Irintchev, A.; Langer, M.; Zweyer, M.; Theisen, R.; Wernig, A. Functional improvement of damaged adult mouse muscle by implantation of primary myoblasts. J. Physiol. 1997, 500, 775–785. [Google Scholar] [CrossRef]
- Barry, P.H. JPCalc, a software package for calculating liquid junction potential corrections in patch-clamp, intracellular, epithelial and bilayer measurements and for correcting junction potential measurements. J. Neurosci. Methods 1994, 51, 107–116. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciancalepore, M.; Ragnini, A.; Zacchi, P.; Borelli, V.; D’Andrea, P.; Lorenzon, P.; Bernareggi, A. A Pharmacological Investigation of the TMEM16A Currents in Murine Skeletal Myogenic Precursor Cells. Int. J. Mol. Sci. 2024, 25, 2225. https://doi.org/10.3390/ijms25042225
Sciancalepore M, Ragnini A, Zacchi P, Borelli V, D’Andrea P, Lorenzon P, Bernareggi A. A Pharmacological Investigation of the TMEM16A Currents in Murine Skeletal Myogenic Precursor Cells. International Journal of Molecular Sciences. 2024; 25(4):2225. https://doi.org/10.3390/ijms25042225
Chicago/Turabian StyleSciancalepore, Marina, Asja Ragnini, Paola Zacchi, Violetta Borelli, Paola D’Andrea, Paola Lorenzon, and Annalisa Bernareggi. 2024. "A Pharmacological Investigation of the TMEM16A Currents in Murine Skeletal Myogenic Precursor Cells" International Journal of Molecular Sciences 25, no. 4: 2225. https://doi.org/10.3390/ijms25042225
APA StyleSciancalepore, M., Ragnini, A., Zacchi, P., Borelli, V., D’Andrea, P., Lorenzon, P., & Bernareggi, A. (2024). A Pharmacological Investigation of the TMEM16A Currents in Murine Skeletal Myogenic Precursor Cells. International Journal of Molecular Sciences, 25(4), 2225. https://doi.org/10.3390/ijms25042225