
The Resistance of Soybean Variety Heinong 84 to Apple Latent Spherical Virus Is Controlled by Two Genetic Loci


 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Screening of Susceptible Soybean Varieties in Northeast China
2.2. Genetic Analysis of the Resistance of Heinong 84 to ALSV
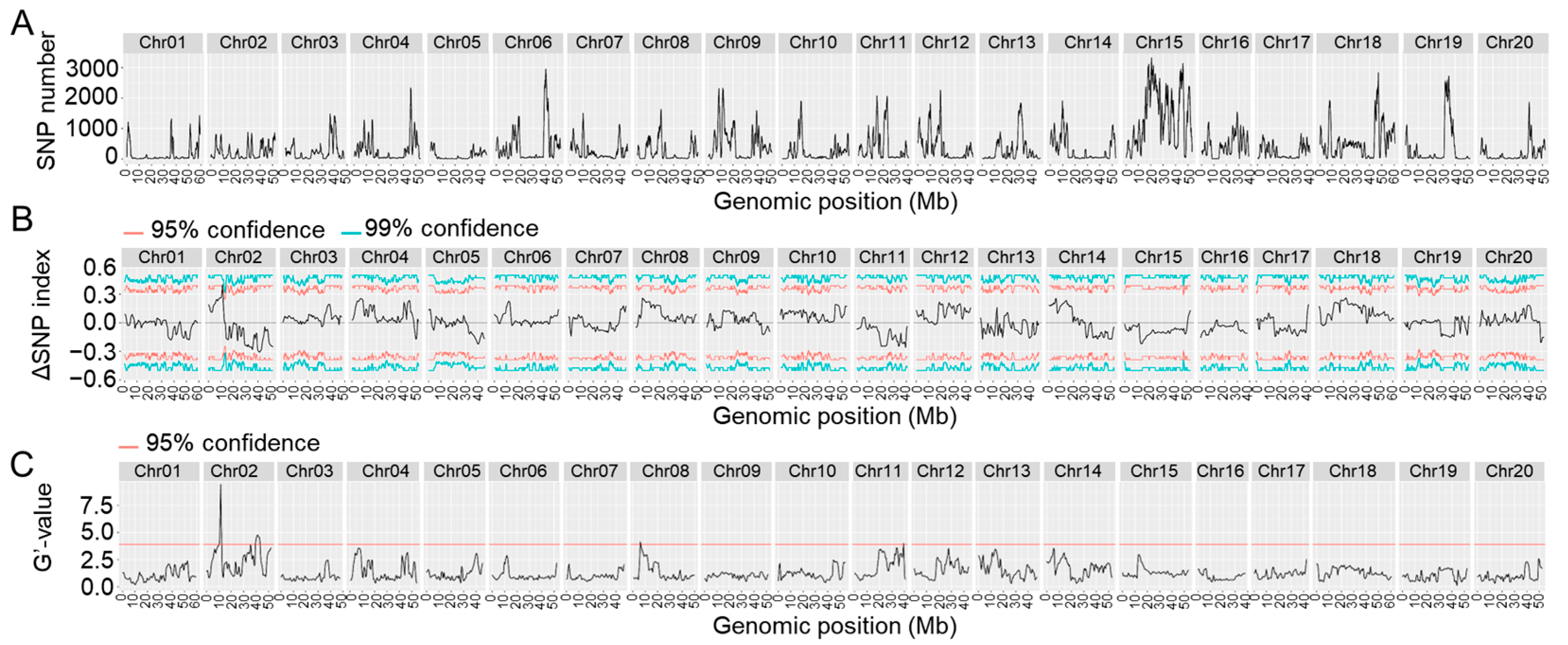
2.3. Location of the Resistance Loci in Heinong 84 by HGS–BSA
2.4. Dissection the Genetic Basis of RALSV-L1 and RALSV-L2 Using CAPS Markers
2.5. Prediction of Candidate Genes in the Resistance Loci RALSV-L1 and RALSV-L1
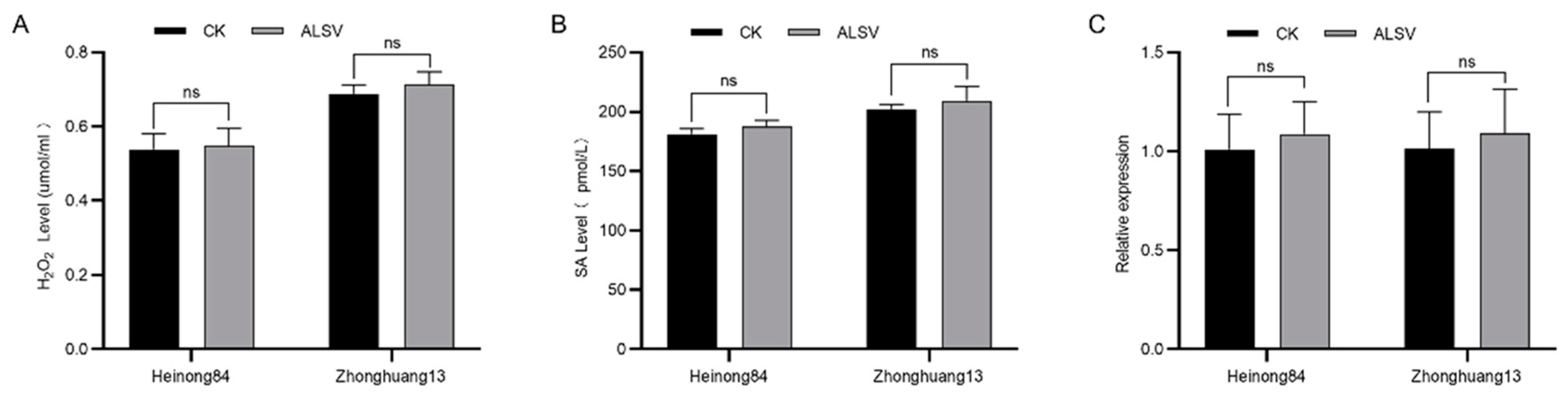
2.6. The Resistance Is Not Associated with Innate Immunity
2.7. There Are Resistance Genes Other Than RALSV-L1 or RALSV-L2 in Soybeans
3. Discussion
4. Materials and Methods
4.1. Soybean Varieties and Growth Conditions
4.2. ALSV Inoculation
4.3. RNA Extraction and RT–qPCR
4.4. DNA Extraction, DNA Pool Preparation, and HTS
4.5. Bulk Segregation Analysis (BSA)
4.6. Design and Validation of CAPS Primers
4.7. SA and H2O2 Quantification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops that feed the World 2. Soybean—Worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- SoyStat. Available online: http://www.soystats.com/ (accessed on 12 June 2023).
- Huang, C.; Qian, Y.; Li, Z.; Zhou, X. Virus-induced gene silencing and its applications in plant functional genomics. Sci. China Life Sci. 2012, 55, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, C.; Whitham, S.A.; Hill, J.H. Development and Use of an Efficient DNA-Based Viral Gene Silencing Vector for Soybean. Mol. Plant-Microbe Interact. 2009, 22, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ghabrial, S.A. Development of Bean pod mottle virus-based vectors for stable protein expression and sequence-specific virus-induced gene silencing in soybean. Virology 2006, 344, 401–411. [Google Scholar] [CrossRef]
- Lim, S.; Nam, M.; Kim, K.H.; Lee, S.-H.; Moon, J.-K.; Lim, H.-S.; Choung, M.-G.; Kim, S.-M.; Moon, J.S. Development of a new vector using Soybean yellow common mosaic virus for gene function study or heterologous protein expression in soybeans. J. Virol. Methods 2016, 228, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Lim, S.; Kang, Y.J.; Yoon, M.Y.; Nam, M.; Jun, T.H.; Seo, M.-J.; Baek, S.-B.; Lee, J.-H.; Moon, J.-K.; et al. Optimization of a virus-induced gene silencing system with Soybean yellow common mosaic virus for gene function studies in soybeans. Plant Pathol. J. 2016, 32, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Yoshikawa, N. Virus-induced gene silencing in soybean seeds and the emergence stage of soybean plants with Apple latent spherical virus vectors. Plant Mol. Biol. 2009, 71, 15–24. [Google Scholar] [CrossRef]
- Zhang, C.; Bradshaw, J.D.; Whitham, S.A.; Hill, J.H. The development of an efficient multipurpose bean pod mottle virus viral vector set for foreign gene expression and RNA silencing. Plant Physiol. 2010, 153, 52–65. [Google Scholar] [CrossRef]
- Le Gall, O.; Sanfaçon, H.; Ikegami, M.; Iwanami, T.; Jones, T.; Karasev, A.; Lehto, K.; Wellink, J.; Wetzel, T.; Yoshikawa, N. Cheravirus and Sadwavirus: Two unassigned genera of plant positive-sense single-stranded RNA viruses formerly considered atypical members of the genus Nepovirus (family Comoviridae). Arch. Virol. 2007, 152, 1767–1774. [Google Scholar] [CrossRef]
- Li, C.; Yoshikawa, N.; Takahashi, T.; Ito, T.; Yoshida, K.; Koganezawa, H. Nucleotide sequence and genome organization of Apple latent spherical virus: A new virus classified into the family Comoviridae. J. Gen. Virol. 2000, 81, 541–547. [Google Scholar] [CrossRef]
- Koganezawa, H. An isometric viruslike particle isolated from russet ring-diseased apple. Ann. Phytopath. Soc. Jpn. 1985, 51, 363. [Google Scholar]
- Li, C.; Yamagishi, N.; Kasajima, I.; Yoshikawa, N. Virus-induced gene silencing and virus-induced flowering in strawberry (Fragaria× ananassa) using apple latent spherical virus vectors. Hortic. Res. 2019, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, A.; Yamagata, K.; Sugai, T.; Takahashi, Y.; Sugawara, E.; Tamura, A.; Yaegashi, H.; Yamagishi, N.; Takahashi, T.; Isogai, M. Apple latent spherical virus vectors for reliable and effective virus-induced gene silencing among a broad range of plants including tobacco, tomato, Arabidopsis thaliana, cucurbits, and legumes. Virology 2009, 386, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N.; Kon, T.; Yamagishi, N.; Takahashi, T.; Natsuaki, T.; Yoshikawa, N. Apple latent spherical virus vector as vaccine for the prevention and treatment of mosaic diseases in pea, broad bean, and eustoma plants by bean yellow mosaic virus. Viruses 2014, 6, 4242–4257. [Google Scholar] [CrossRef] [PubMed]
- Izuishi, Y.; Isaka, N.; Li, H.; Nakanishi, K.; Kageyama, J.; Ishikawa, K.; Shimada, T.; Masuta, C.; Yoshikawa, N.; Kusano, H.; et al. Apple latent spherical virus (ALSV)-induced gene silencing in a medicinal plant, Lithospermum erythrorhizon. Sci. Rep. 2020, 10, 13555. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Kikuchi, T.; Kasajima, I.; Li, C.; Yamagishi, N.; Yamashita, H.; Yoshikawa, N. Virus-induced flowering by Apple latent spherical virus vector: Effective use to accelerate breeding of grapevine. Viruses 2020, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Sasaki, S.; Yamagata, K.; Komori, S.; Nagase, M.; Wada, M.; Yamamoto, T.; Yoshikawa, N. Promotion of flowering and reduction of a generation time in apple seedlings by ectopical expression of the Arabidopsis thaliana FT gene using the Apple latent spherical virus vector. Plant Mol. Biol. 2011, 75, 193–204. [Google Scholar] [CrossRef]
- Nakamura, K.; Yamagishi, N.; Isogai, M.; Komori, S.; Ito, T.; Yoshikawa, N. Seed and pollen transmission of Apple latent spherical virus in apple. J. Gen. Plant Pathol. 2011, 77, 48–53. [Google Scholar] [CrossRef]
- Don, Y.; Wei, Q.; Hong, H.; Huang, Y.; Zhao, Y.; Feng, M.; Dou, D.; Xu, Y.; Tao, X. Establishment of ALSV-induced gene silencing in Chinese soybean cultivars. Sci. Agric. Sin. 2022, 55, 1710–1722. [Google Scholar]
- Gedling, C.R.; Ali, E.M.; Gunadi, A.; Finer, J.J.; Xie, K.; Liu, Y.; Yoshikawa, N.; Qu, F.; Dorrance, A.E. Improved apple latent spherical virus-induced gene silencing in multiple soybean genotypes through direct inoculation of agro-infiltrated Nicotiana benthamiana extract. Plant Methods 2018, 14, 19. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Deng, W.; Liu, J.; Fang, Y.; Liu, Y.; Ma, T.; Zhang, Y.; Xue, Y.; Tang, X.; et al. Fine mapping the soybean mosaic virus resistance gene in soybean cultivar Heinong 84 and development of CAPS markers for rapid identification. Viruses 2022, 14, 2533. [Google Scholar] [CrossRef]
- Thiel, T.; Kota, R.; Grosse, I.; Stein, N.; Graner, A. SNP2CAPS: A SNP and INDEL analysis tool for CAPS marker development. Nucleic Acids Res. 2004, 32, e5. [Google Scholar] [CrossRef]
- Yeam, I.; Kang, B.C.; Lindeman, W.; Frantz, J.D.; Faber, N.; Jahn, M.M. Allele-specific CAPS markers based on point mutations in resistance alleles at the pvr1 locus encoding eIF4E in Capsicum. Theor. Appl. Genet. 2005, 112, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Valli, A.; García, J.A.; Zhou, X.; Cheng, X. The tug-of-war between plants and viruses: Great progress and many remaining questions. Viruses 2019, 11, 203. [Google Scholar] [CrossRef]
- Chisholm, S.T.; Mahajan, S.K.; Whitham, S.A.; Yamamoto, M.L.; Carrington, J.C. Cloning of the Arabidopsis RTM1 gene, which controls restriction of long-distance movement of tobacco etch virus. Proc. Natl. Acad. Sci. USA 2000, 97, 489–494. [Google Scholar] [CrossRef]
- Yamaji, Y.; Maejima, K.; Komatsu, K.; Shiraishi, T.; Okano, Y.; Himeno, M.; Sugawara, K.; Neriya, Y.; Minato, N.; Miura, C.; et al. Lectin-mediated resistance impairs plant virus infection at the cellular level. Plant Cell 2012, 24, 778–793. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Kezuka, Y.; Kobayashi, C.; Kato, M.; Inoue, T.; Nonaka, T.; Ishikawa, M.; Matsumura, H.; Katoh, E. Structural basis for the recognition–evasion arms race between Tomato mosaic virus and the resistance gene Tm-1. Proc. Natl. Acad. Sci. USA 2014, 111, E3486–E3495. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Masuda, K.; Naito, S.; Meshi, T.; Ishikawa, M. An inhibitor of viral RNA replication is encoded by a plant resistance gene. Proc. Natl. Acad. Sci. USA 2007, 104, 13833–13838. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, H.; Gong, Y.; Tao, Y.; Jiang, L.; Zuo, W.; Yang, Q.; Ye, J.; Lai, J.; Wu, J.; et al. An atypical thioredoxin imparts early resistance to Sugarcane mosaic virus in Maize. Mol. Plant 2017, 10, 483–497. [Google Scholar] [CrossRef]
- Hashimoto, M.; Neriya, Y.; Yamaji, Y.; Namba, S. Recessive Resistance to Plant Viruses: Potential Resistance Genes Beyond Translation Initiation Factors. Front. Microbiol. 2016, 7, 1695. [Google Scholar] [CrossRef]
- Robaglia, C.; Caranta, C. Translation initiation factors: A weak link in plant RNA virus infection. Trends Plant Sci. 2006, 11, 40–45. [Google Scholar] [CrossRef]
- Bruckner, F.P.; da Silva Xavier, A.; de Souza Cascardo, R.; Otoni, W.C.; Zerbini, F.M.; Alfenas-Zerbini, P. Translationally controlled tumor protein (TCTP) from tomato and Nicotiana benthamiana is necessary for successful infection by A Potyvirus. Mol. Plant Pathol. 2017, 18, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Neriya, Y.; Keima, T.; Iwabuchi, N.; Koinuma, H.; Hagiwara-Komoda, Y.; Ishikawa, K.; Himeno, M.; Maejima, K.; Yamaji, Y.; et al. EXA1, a GYF domain protein, is responsible for loss-of-susceptibility to plantago asiatica mosaic virus in Arabidopsis thaliana. Plant J. 2016, 88, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Mansfeld, B.N.; Grumet, R. QTLseqr: An R package for bulk segregant analysis with next-generation sequencing. Plant Genome 2018, 11, 180006. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R. BEDTools: The Swiss-army tool for genome feature analysis. Curr. Protoc. Bioinform. 2014, 47, 11–12. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parent and Offspring | Resistant | Susceptible | Total | Theoretical Separation Ratio | χ2 | p |
|---|---|---|---|---|---|---|
| Heinong 84 | 6 | 0 | 6 | |||
| Zhonghuang 13 | 0 | 6 | 6 | |||
| F2 | 60 | 40 | 3:1 | 4.4672 | 0.03445 | |
| 9:7 | 0.185 | 0.6673 |
| Chromosome | Start Position | End Position | Length | Number of SNPs | p Value |
|---|---|---|---|---|---|
| 2 | 39,213,950 | 43,829,476 | 4,615,526 | 1710 | 0.00004 |
| 11 | 19,104,714 | 25,514,536 | 6,409,822 | 5666 | 0.00040 |
| Genotype | Resistant | Susceptible | Total | p Value |
|---|---|---|---|---|
| A1A1 | 12 | 3 | 15 | |
| A1a1 | 13 | 9 | 22 | |
| a1a1 | 5 | 18 | 23 | |
| A1A1 + A1a1 + a1a1 | 30 | 30 | 60 | 0.001115 |
| B2B2 | 11 | 19 | 30 | |
| B2b2 | 0 | 1 | 1 | |
| b2b2 | 19 | 10 | 29 | |
| B2B2 + B2b2 + b2b2 | 30 | 30 | 60 | 0.03789 |
| A1A1b2b2 | 10 | 1 | 11 | |
| A1a1b2b2 | 8 | 3 | 11 | |
| A1A1B2B2 | 2 | 2 | 4 | |
| A1a1B2B2 | 5 | 6 | 11 | |
| a1a1B2B2 | 4 | 11 | 15 | |
| a1a1b2b2 | 1 | 6 | 7 | |
| a1a1B2b2 | 0 | 1 | 1 | |
| All | 30 | 30 | 60 | 0.003263 |
| Primer Names | Sequences (5′-3′) | Usage |
|---|---|---|
| GmPR1-F | TGCTAATCAACGCAAAGG | RT–qPCR |
| GmPR1-R | CATCCAAGACGCACCGAG | RT–qPCR |
| Cons6-F | AAGTTAGGAGCCCAAGACAT | RT–qPCR |
| Cons6-R | AGCGAGTTCATTGAAGCAGA | RT–qPCR |
| C2-SNP3999-F | GGACATACTTGCAAGTTTAGGG | CAPS analysis |
| C2-SNP3999-R | GAGGTTGGCCTATAAACCTC | CAPS analysis |
| C2-SNP4209-F | GCAGGGCATAAACTTTCAATC | CAPS analysis |
| C2-SNP4209-R | CAGTTAAGTGATTGCATTGGC | CAPS analysis |
| C2-SNP4232-F | CCACATCACTAACCTAAAGGC | CAPS analysis |
| C2-SNP4232-R | TTCACTTATGCCCAAGGGC | CAPS analysis |
| C11-SNP2115-F | GATCGAGGTAGTGGTAGACATC | CAPS analysis |
| C11-SNP2115-R | CACTTGTGCAGCAGAATCGTG | CAPS analysis |
| C11-SNP2130-F | GCAGCGGTTCAAAACCGTCC | CAPS analysis |
| C11-SNP2130-R | TGATCCTGAAGGTTGAGGATGC | CAPS analysis |
| C11-SNP2298-F | ACTCTGAAGCGTATCCATGACC | CAPS analysis |
| C11-SNP2298-R | TGTTGACGGTTTATTTAGATGAC | CAPS analysis |
| ALSV2-F | GCTCGTCACCTGTTCAGCTC | RT–PCR |
| ALSV2-R | CTAGGTGTAACCAGCTTTGAGC | RT–PCR |
| MP944-F | CTGATGGTGTCCTCAAAGAGG | RT–PCR |
| VP195-R | GGTAAATTCTGGAGTAGAAG | RT–PCR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, T.; Zhang, Y.; Li, Y.; Zhao, Y.; Attiogbe, K.B.; Fan, X.; Fan, W.; Sun, J.; Luo, Y.; Yu, X.; et al. The Resistance of Soybean Variety Heinong 84 to Apple Latent Spherical Virus Is Controlled by Two Genetic Loci. Int. J. Mol. Sci. 2024, 25, 2034. https://doi.org/10.3390/ijms25042034
Ma T, Zhang Y, Li Y, Zhao Y, Attiogbe KB, Fan X, Fan W, Sun J, Luo Y, Yu X, et al. The Resistance of Soybean Variety Heinong 84 to Apple Latent Spherical Virus Is Controlled by Two Genetic Loci. International Journal of Molecular Sciences. 2024; 25(4):2034. https://doi.org/10.3390/ijms25042034
Chicago/Turabian StyleMa, Tingshuai, Ying Zhang, Yong Li, Yu Zhao, Kekely Bruno Attiogbe, Xinyue Fan, Wenqian Fan, Jiaxing Sun, Yalou Luo, Xinwei Yu, and et al. 2024. "The Resistance of Soybean Variety Heinong 84 to Apple Latent Spherical Virus Is Controlled by Two Genetic Loci" International Journal of Molecular Sciences 25, no. 4: 2034. https://doi.org/10.3390/ijms25042034
APA StyleMa, T., Zhang, Y., Li, Y., Zhao, Y., Attiogbe, K. B., Fan, X., Fan, W., Sun, J., Luo, Y., Yu, X., Ji, W., Cheng, X., & Wu, X. (2024). The Resistance of Soybean Variety Heinong 84 to Apple Latent Spherical Virus Is Controlled by Two Genetic Loci. International Journal of Molecular Sciences, 25(4), 2034. https://doi.org/10.3390/ijms25042034