

Untargeted Metabolomic Analyses and Antilipidemic Effects of Citrus Physiological Premature Fruit Drop

Abstract

1. Introduction
2. Results
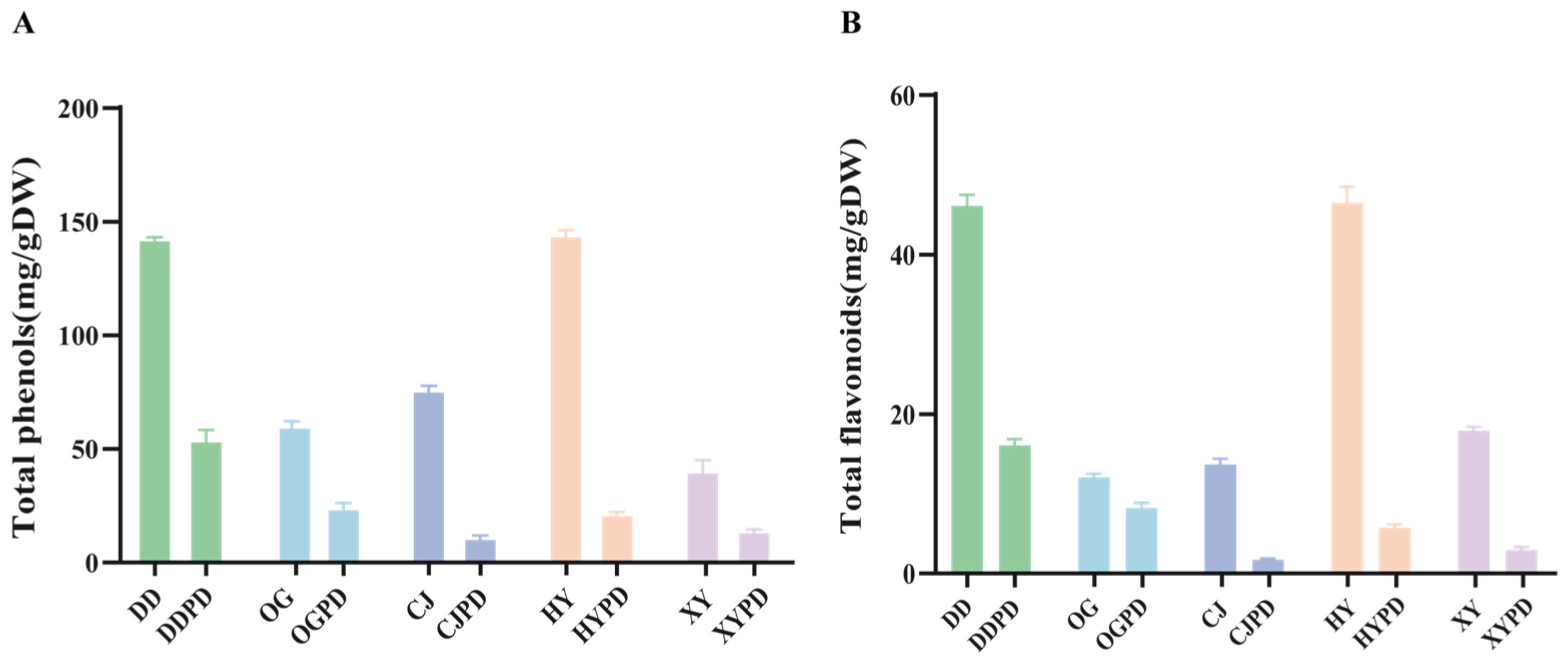
2.1. Differences in Citrus Fruits Chemical Composition
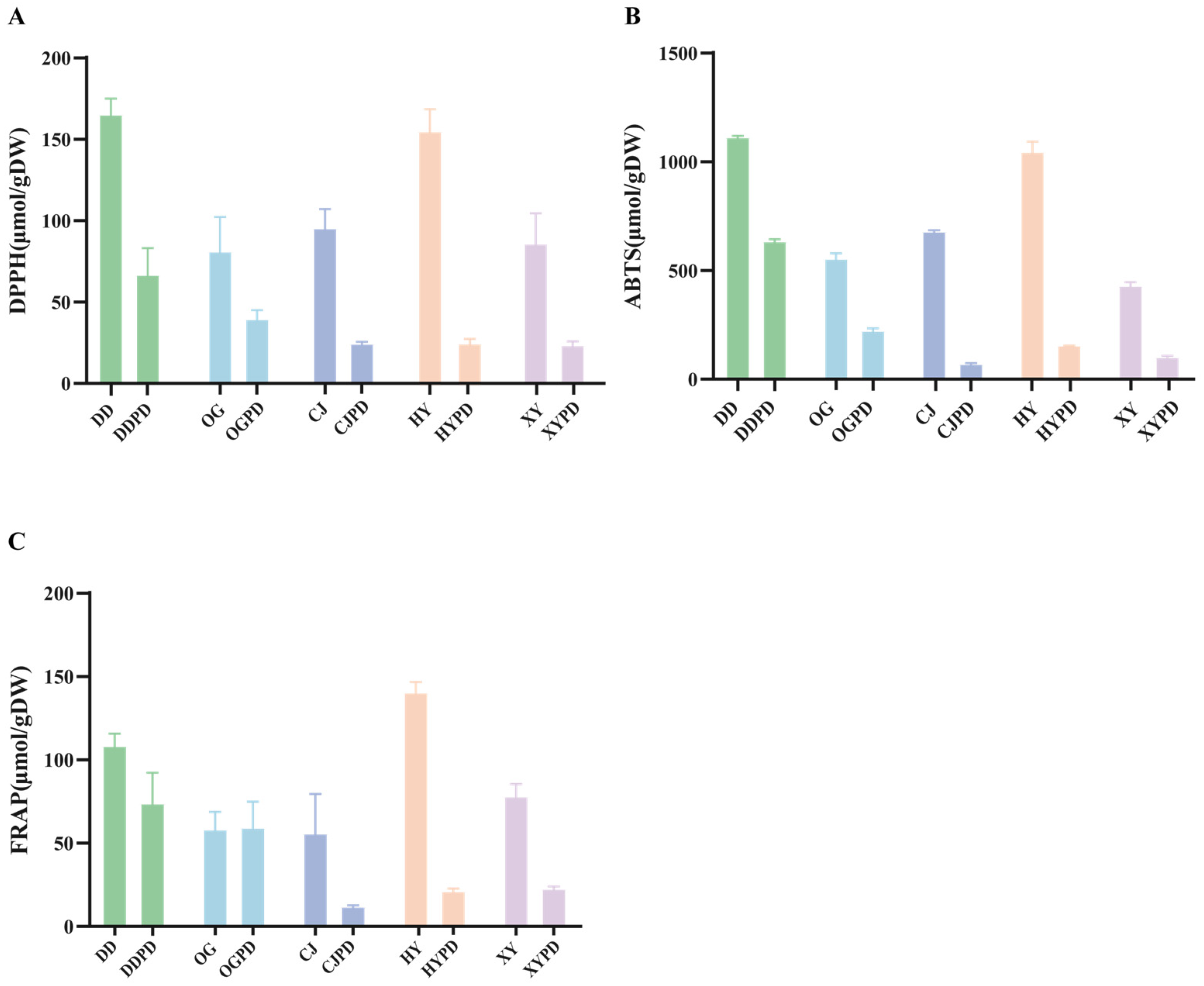
2.2. Antioxidant Activity in Citrus Fruits Extract
2.3. Untargeted Metabolomic Analysis of CPFDs
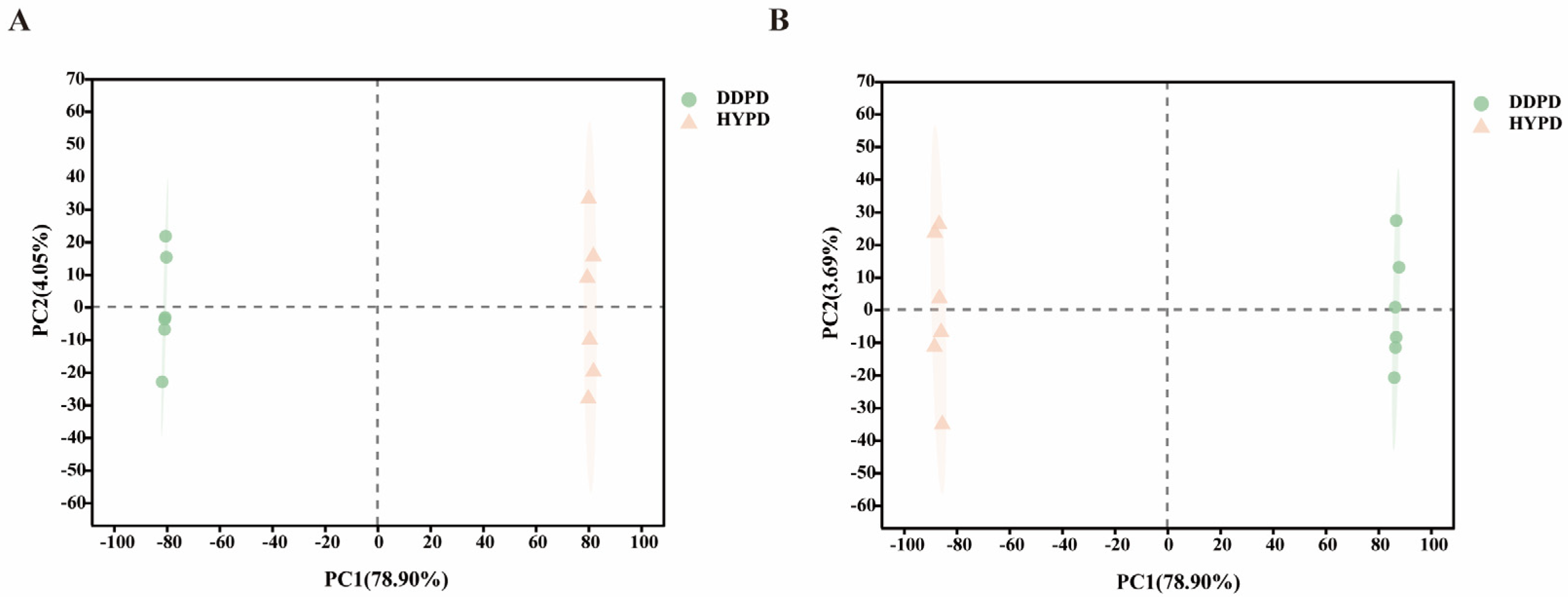
2.3.1. Principal Component Analysis of CPFD
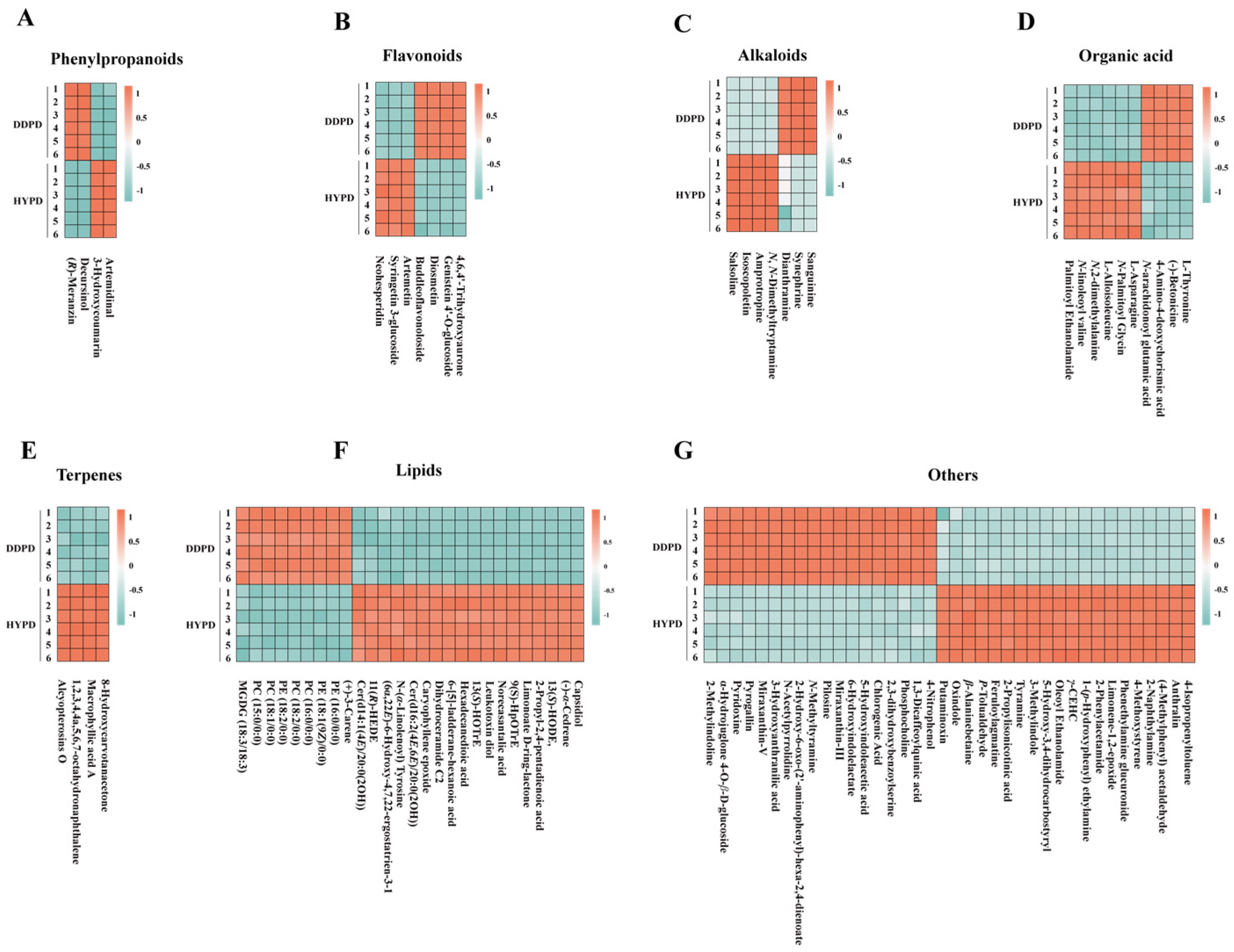
2.3.2. Screening and Identification of Candidate Differential Metabolites
- A.
- Phenylpropanoids
- B.
- Flavonoids
- C.
- Alkaloids
- D.
- Organic acid
- E.
- Terpenes
- F.
- Lipids
- G.
- Others
2.4. Antilipidemic Effects of CPFD
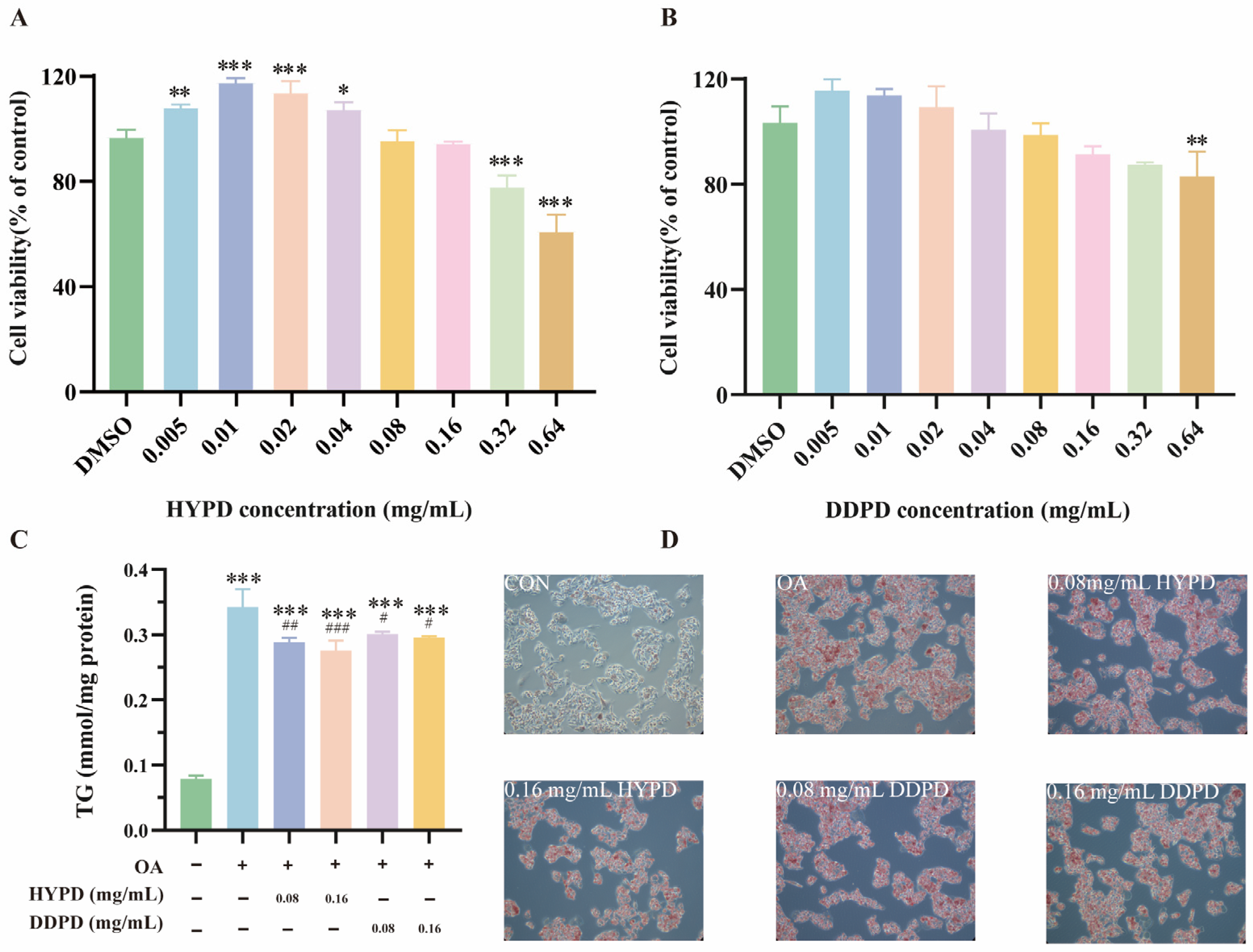
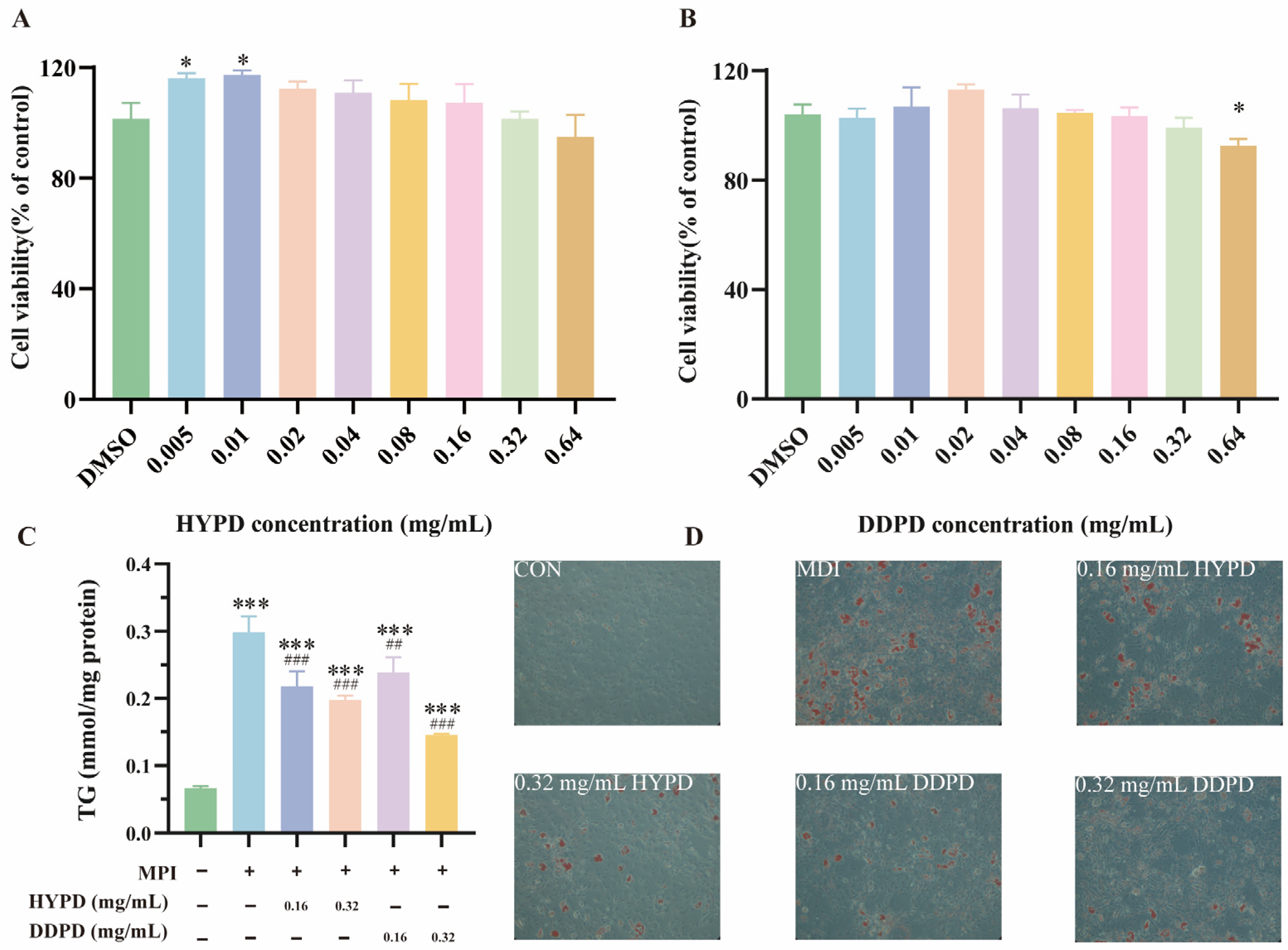
2.4.1. CPFD Alleviated OA-Induced Steatosis in HepG2
2.4.2. CPFD Reduced Lipid Accumulation in 3T3-L1 Adipocytes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Reagents
4.3. CPFD Samples and Extracts Preparation
4.4. Determinations of Phytochemical Composition
4.5. In Vitro Antioxidant Activity Assays of Citrus Fruit Extract
4.6. Untargeted Metabolomic Analysis of CPFD
4.7. Cell Culture
4.8. Cell Viability Assay
4.9. OA-Induced Steatosis in HepG2 Cells
4.10. Differentiation of 3T3-L1 Cells
4.11. Oil Red O Staining
4.12. Cellular Triglyceride Content
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jaacks, L.M.; Vandevijvere, S.; Pan, A.; McGowan, C.J.; Wallace, C.; Imamura, F.; Mozaffarian, D.; Swinburn, B.; Ezzati, M. The Obesity Transition: Stages of the Global Epidemic. Lancet Diabetes Endocrinol. 2019, 7, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Johnston, L.J.; Wu, C.; Ma, X. Gut Microbiota and Its Metabolites: Bridge of Dietary Nutrients and Obesity-Related Diseases. Crit. Rev. Food Sci. Nutr. 2023, 63, 3236–3253. [Google Scholar] [CrossRef] [PubMed]
- Nanadikar, M.S.; Vergel Leon, A.M.; Guo, J.; van Belle, G.J.; Jatho, A.; Philip, E.S.; Brandner, A.F.; Böckmann, R.A.; Shi, R.; Zieseniss, A.; et al. IDH3γ Functions as a Redox Switch Regulating Mitochondrial Energy Metabolism and Contractility in the Heart. Nat. Commun. 2023, 14, 2123. [Google Scholar] [CrossRef] [PubMed]
- Kobori, M.; Akimoto, Y.; Takahashi, Y.; Kimura, T. Combined Effect of Quercetin and Fish Oil on Oxidative Stress in the Liver of Mice Fed a Western-Style Diet. J. Agric. Food Chem. 2020, 68, 13267–13275. [Google Scholar] [CrossRef] [PubMed]
- Savini, I.; Catani, M.V.; Evangelista, D.; Gasperi, V.; Avigliano, L. Obesity-Associated Oxidative Stress: Strategies Finalized to Improve Redox State. Int. J. Mol. Sci. 2013, 14, 10497–10538. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef] [PubMed]
- Dufour, J.-F.; Anstee, Q.M.; Bugianesi, E.; Harrison, S.; Loomba, R.; Paradis, V.; Tilg, H.; Wong, V.W.-S.; Zelber-sagi, S. Current Therapies and New Developments in NASH. Gut 2022, 71, 2123–2134. [Google Scholar] [CrossRef]
- Lomonaco, R.; Leiva, E.G.; Bril, F.; Shrestha, S.; Mansour, L.; Budd, J.; Romero, J.P.; Schmidt, S.; Chang, K.-L.; Samraj, G.; et al. Advanced Liver Fibrosis Is Common in Patients With Type 2 Diabetes Followed in the Outpatient Setting: The Need for Systematic Screening. Diabetes Care 2021, 44, 399. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Setzer, W.N. Biological Activities and Safety of Citrus spp. Essential Oils. Int. J. Mol. Sci. 2018, 19, 1966. [Google Scholar] [CrossRef]
- Abdel-Naeem, H.H.S.; Elshebrawy, H.A.; Imre, K.; Morar, A.; Herman, V.; Pașcalău, R.; Sallam, K.I. Antioxidant and Antibacterial Effect of Fruit Peel Powders in Chicken Patties. Foods 2022, 11, 301. [Google Scholar] [CrossRef]
- García-Nicolás, M.; Ledesma-Escobar, C.A.; Priego-Capote, F. Spatial Distribution and Antioxidant Activity of Extracts from Citrus Fruits. Antioxidants 2023, 12, 781. [Google Scholar] [CrossRef]
- Al-Aubaidy, H.A.; Dayan, A.; Deseo, M.A.; Itsiopoulos, C.; Jamil, D.; Hadi, N.R.; Thomas, C.J. Twelve-Week Mediterranean Diet Intervention Increases Citrus Bioflavonoid Levels and Reduces Inflammation in People with Type 2 Diabetes Mellitus. Nutrients 2021, 13, 1133. [Google Scholar] [CrossRef]
- Liu, N.; Li, X.; Zhao, P.; Zhang, X.; Qiao, O.; Huang, L.; Guo, L.; Gao, W. A Review of Chemical Constituents and Health-Promoting Effects of Citrus Peels. Food Chem. 2021, 365, 130585. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Data. 2022. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 30 November 2023).
- Qi, T.; Ren, J.; Li, X.; An, Q.; Zhang, N.; Jia, X.; Pan, S.; Fan, G.; Zhang, Z.; Wu, K. Structural Characteristics and Gel Properties of Pectin from Citrus Physiological Premature Fruit Drop. Carbohydr. Polym. 2023, 309, 120682. [Google Scholar] [CrossRef] [PubMed]
- Chaix, A.; Zarrinpar, A.; Miu, P.; Panda, S. Time-Restricted Feeding Is a Preventative and Therapeutic Intervention against Diverse Nutritional Challenges. Cell Metab. 2014, 20, 991. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Qiao, L.; Shen, Y.; Jiang, P.; Chen, J.; Ye, X. Phytochemical Profile and Antioxidant Activity of Physiological Drop of Citrus Fruits. J. Food Sci. 2013, 78, C37–C42. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Schluesener, H. Health-Promoting Effects of the Citrus Flavanone Hesperidin. Crit. Rev. Food Sci. 2017, 57, 613–631. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.-J.; Chen, J.-B.; Cao, J.-P.; Li, X.; Sun, C.-D. Citrus Flavonoids and Their Antioxidant Evaluation. Crit. Rev. Food Sci. Nutr. 2022, 62, 3833–3854. [Google Scholar] [CrossRef]
- Lee, D.; Lee, J.H.; Kim, B.H.; Lee, S.; Kim, D.-W.; Kang, K.S. Phytochemical Combination (p-Synephrine, p-Octopamine Hydrochloride, and Hispidulin) for Improving Obesity in Obese Mice Induced by High-Fat Diet. Nutrients 2022, 14, 2164. [Google Scholar] [CrossRef]
- Talari, N.K.; Mattam, U.; Meher, N.K.; Paripati, A.K.; Mahadev, K.; Krishnamoorthy, T.; Sepuri, N.B.V. Lipid-Droplet Associated Mitochondria Promote Fatty-Acid Oxidation through a Distinct Bioenergetic Pattern in Male Wistar Rats. Nat. Commun. 2023, 14, 766. [Google Scholar] [CrossRef]
- Oh, C.-M.; Namkung, J.; Go, Y.; Shong, K.E.; Kim, K.; Kim, H.; Park, B.-Y.; Lee, H.W.; Jeon, Y.H.; Song, J.; et al. Regulation of Systemic Energy Homeostasis by Serotonin in Adipose Tissues. Nat. Commun. 2015, 6, 6794. [Google Scholar] [CrossRef]
- Venza, I.; Venza, M.; Visalli, M.; Lentini, G.; Teti, D.; d’Alcontres, F.S. ROS as Regulators of Cellular Processes in Melanoma. Oxidative Med. Cell. Longev. 2021, 2021, 1208690. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.D.; Ko, W.J.; Koh, K.S.; Jeon, Y.J.; Kim, S.H. Composition of Flavonoids and Antioxidative Activity from Juice of Jeju Native Citrus Fruits during Maturation. Korean J. Nutr. 2009, 42, 278–290. [Google Scholar] [CrossRef]
- Ruiz-Moreno, C.; Del Coso, J.; Giráldez-Costas, V.; González-García, J.; Gutiérrez-Hellín, J. Effects of P-Synephrine during Exercise: A Brief Narrative Review. Nutrients 2021, 13, 233. [Google Scholar] [CrossRef]
- Dugo, P.; Russo, M.; Sarò, M.; Carnovale, C.; Bonaccorsi, I.; Mondello, L. Multidimensional Liquid Chromatography for the Determination of Chiral Coumarins and Furocoumarins in Citrus Essential Oils. J. Sep. Sci. 2012, 35, 1828–1836. [Google Scholar] [CrossRef]
- Teng, W.-Y.; Chen, C.-C.; Chung, R.-S. HPLC Comparison of Supercritical Fluid Extraction and Solvent Extraction of Coumarins from the Peel of Citrus Maxima Fruit. Phytochem. Anal. 2005, 16, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Gao, Z.; Xia, J.; Ritenour, M.A.; Li, G.; Shan, Y. Comparative Analysis of Chemical Composition, Antimicrobial and Antioxidant Activity of Citrus Essential Oils from the Main Cultivated Varieties in China. LWT 2018, 97, 825–839. [Google Scholar] [CrossRef]
- Yuan, H.; Li, Y.; Ling, F.; Guan, Y.; Zhang, D.; Zhu, Q.; Liu, J.; Wu, Y.; Niu, Y. The Phytochemical Epigallocatechin Gallate Prolongs the Lifespan by Improving Lipid Metabolism, Reducing Inflammation and Oxidative Stress in High-fat Diet-fed Obese Rats. Aging Cell 2020, 19, e13199. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Flint, A.J.; Qi, Q.; van Dam, R.M.; Sampson, L.A.; Rimm, E.B.; Holmes, M.D.; Willett, W.C.; Hu, F.B.; Sun, Q. Whole Grain Intake and Mortality: Two Large Prospective Studies in U.S. Men and Women. JAMA Intern. Med. 2015, 175, 373. [Google Scholar] [CrossRef]
- Cremonini, E.; Daveri, E.; Iglesias, D.E.; Kang, J.; Wang, Z.; Gray, R.; Mastaloudis, A.; Kay, C.D.; Hester, S.N.; Wood, S.M.; et al. A Randomized Placebo-Controlled Cross-over Study on the Effects of Anthocyanins on Inflammatory and Metabolic Responses to a High-Fat Meal in Healthy Subjects. Redox Biol. 2022, 51, 102273. [Google Scholar] [CrossRef]
- Robert, V.; Farese, J.; Zechner, R.; Newgard, C.B.; Walther, T.C. The Problem of Establishing Relationships between Hepatic Steatosis and Hepatic Insulin Resistance. Cell Metab. 2012, 15, 570. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Sakurai, T.; Chen, Z.; Inoue, N.; Chiba, H.; Hui, S.-P. Lysophosphatidylethanolamine Affects Lipid Accumulation and Metabolism in a Human Liver-Derived Cell Line. Nutrients 2022, 14, 579. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Cai, N.; Lyu, Y.; Yao, L.; Wang, F.; Xu, H.; Hu, Z.; Li, H.; Xu, X. γ-Mangostin Ameliorates Free Fatty Acid-Induced Lipid Accumulation via the SIRT1/LKB1/AMPK Pathway in HepG2 and L02 Cells. J. Agric. Food Chem. 2019, 67, 13929–13938. [Google Scholar] [CrossRef] [PubMed]
- Mt, I.; Es, A.; Ms, M. Anti-Obesity Effect of Plant Diterpenes and Their Derivatives: A Review. Phytother. Res. PTR 2020, 34, 1216–1225. [Google Scholar] [CrossRef]
- Jia, S.; Hu, Y.; Zhang, W.; Zhao, X.; Chen, Y.; Sun, C.; Li, X.; Chen, K. Hypoglycemic and Hypolipidemic Effects of Neohesperidin Derived from Citrus aurantium L. in Diabetic KK-Ay Mice. Food Funct. 2015, 6, 878–886. [Google Scholar] [CrossRef]
- Bacquer, O.L.; Petroulakis, E.; Paglialunga, S.; Poulin, F.; Richard, D.; Cianflone, K.; Sonenberg, N. Elevated Sensitivity to Diet-Induced Obesity and Insulin Resistance in Mice Lacking 4E-BP1 and 4E-BP2. J. Clin. Investig. 2007, 117, 387. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kim, J.; Park, J.Y.; Kang, K.S.; Park, J.H.; Hwang, G.S. Processed Panax Ginseng, Sun Ginseng, Inhibits the Differentiation and Proliferation of 3T3-L1 Preadipocytes and Fat Accumulation in Caenorhabditis Elegans. J. Ginseng Res. 2017, 41, 257. [Google Scholar] [CrossRef]
- Kang, S.-I.; Shin, H.-S.; Kim, H.-M.; Hong, Y.-S.; Yoon, S.-A.; Kang, S.-W.; Kim, J.-H.; Kim, M.-H.; Ko, H.-C.; Kim, S.-J. Immature Citrus Sunki Peel Extract Exhibits Antiobesity Effects by β-Oxidation and Lipolysis in High-Fat Diet-Induced Obese Mice. Biol. Pharm. Bull. 2012, 35, 223–230. [Google Scholar] [CrossRef]
- Lee, D.; Kwak, H.J.; Kim, B.H.; Kim, S.H.; Kim, D.-W.; Kang, K.S. Combined Anti-Adipogenic Effects of Hispidulin and p-Synephrine on 3T3-L1 Adipocytes. Biomolecules 2021, 11, 1764. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Giménez, R.; Rufián-Henares, J.A.; Pastoriza, S. Effect of Brewing Time and Temperature on Antioxidant Capacity and Phenols of White Tea: Relationship with Sensory Properties. Food Chem. 2018, 248, 111–118. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, W.; Zhang, C.; Li, C.; Fang, Z.; Zeng, Z.; Hu, B.; Chen, H.; Wu, W.; Wang, T.; et al. Targeted and Untargeted Metabolomic Analyses and Biological Activity of Tibetan Tea. Food Chem. 2022, 384, 132517. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | VIP | Rt/min | m/z | Mode | CAS | Identification | Formula | Classification | Regulate |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2.4610 | 2.250766667 | 307.1760342 | pos | - | Feruloylagmatine | C15H22N4O3 | Others | down |
| 2 | 2.1676 | 4.89415 | 227.0697288 | pos | 1143-38-0 | Anthralin | C14H10O3 | Others | down |
| 3 | 2.0770 | 6.091066667 | 194.1173346 | pos | 89-31-6 | Salsoline | C11H15NO2 | Alkaloids | down |
| 4 | 2.0330 | 3.8725 | 138.0911662 | pos | 51-67-2 | Tyramine | C8H11NO | Others | down |
| 5 | 2.0154 | 7.645216667 | 163.0386991 | pos | 939-19-5 | 3-Hydroxycoumarin | C9H6O3 | Phenylpropanoids | down |
| 6 | 1.9001 | 1.9989 | 134.0962301 | pos | - | 2-Methylindoline | C9H11N | Others | up |
| 7 | 1.8681 | 3.3298 | 138.0911105 | pos | - | 1-(p-Hydroxyphenyl) ethylamine | C8H11NO | Others | down |
| 8 | 1.8654 | 7.123033333 | 193.0494018 | pos | 776-86-3 | Isoscopoletin | C10H8O4 | Alkaloids | down |
| 9 | 1.7830 | 6.108966667 | 205.194699 | pos | - | 2-Isopropenyl-4a,8-dimethyl-1,2,3,4,4a,5,6,7-octahydronaphthalene | C15H24 | Terpenes | down |
| 10 | 1.7008 | 4.309816667 | 308.2213803 | pos | 148-32-3 | Amprotropine | C18H29NO3 | Alkaloids | down |
| 11 | 1.6769 | 2.36035 | 226.0704863 | pos | - | 4-Amino-4-deoxychorismic acid | C10H11NO5 | Organic acid | up |
| 12 | 1.6710 | 4.109983333 | 593.1852356 | pos | - | Buddleoflavonoloside | C28H32O14 | Flavonoids | up |
| 13 | 1.6570 | 1.566566667 | 152.1066258 | pos | 370-98-9 | N-Methyltyramine | C9H13NO | Others | up |
| 14 | 1.6496 | 7.9102 | 434.2884372 | pos | - | N-arachidonoyl glutamic acid | C25H39NO5 | Organic acid | up |
| 15 | 1.6157 | 3.963116667 | 213.148295 | pos | 172104-03-9 | Putaminoxin | C12H20O3 | Others | down |
| 16 | 1.6154 | 1.78215 | 242.0653613 | pos | 7724-78-9 | 2,3-dihydroxybenzoylserine | C10H11NO6 | Others | up |
| 17 | 1.6147 | 2.415333333 | 140.0339558 | pos | 98-95-3 | 4-Nitrophenol | C6H5NO3 | Others | up |
| 18 | 1.6026 | 4.439366667 | 134.0599624 | pos | 59-48-3 | Oxindole | C8H7NO | Others | down |
| 19 | 1.5979 | 3.528816667 | 141.0907633 | pos | 72010-18-5 | 2-Propyl-2,4-pentadienoic acid | C8H12O2 | Lipids | down |
| 20 | 1.5921 | 5.166566667 | 169.1220375 | pos | - | 8-Hydroxycarvotanacetone | C10H16O2 | Terpenes | down |
| 21 | 1.5477 | 9.551733333 | 221.1895876 | pos | 1139-30-6 | Caryophyllene epoxide | C15H24O | Lipids | down |
| 22 | 1.5409 | 2.214966667 | 170.0807431 | pos | - | Pyridoxine | C8H11NO3 | Others | up |
| 23 | 1.5395 | 5.323516667 | 235.1688016 | pos | - | Alcyopterosins O | C15H22O2 | Terpenes | down |
| 24 | 1.5217 | 5.622416667 | 152.1067485 | pos | 104-09-6 | (4-Methylphenyl) acetaldehyde | C9H10O | Others | down |
| 25 | 1.4640 | 8.133916667 | 136.0755284 | pos | 103-81-1 | 2-Phenylacetamide | C8H9NO | Others | down |
| 26 | 1.4552 | 2.58045 | 222.0756095 | pos | - | 6-Hydroxyindolelactate | C11H11NO4 | Others | up |
| 27 | 1.4246 | 2.09015 | 132.0806876 | pos | 83-34-1 | 3-Methylindole | C9H9N | Others | down |
| 28 | 1.4186 | 6.86395 | 313.2354406 | neg | 125356-86-7 | Leukotoxin diol | C18H34O4 | Lipids | down |
| 29 | 1.4134 | 4.475616667 | 153.1271494 | pos | 1195-92-2 | Limonene-1,2-epoxide | C10H16O | Others | down |
| 30 | 1.4014 | 2.342383333 | 287.1384126 | pos | 13640-28-3 | Pilosine | C16H18N2O3 | Others | up |
| 31 | 1.3877 | 3.222066667 | 290.0654927 | pos | 136945-65-8 | Dianthramine | C14H11NO6 | Alkaloids | up |
| 32 | 1.3826 | 1.890766667 | 135.0803928 | pos | - | 4-Methoxystyrene | C9H10O | Others | down |
| 33 | 1.3678 | 3.2382 | 515.1144208 | neg | 30964-13-7 | 1,3-Dicaffeoylquinic acid | C25H24O12 | Others | up |
| 34 | 1.3282 | 1.24845 | 154.0493069 | pos | 548-93-6 | 3-Hydroxyanthranilic acid | C7H7NO3 | Others | up |
| 35 | 1.3248 | 1.11365 | 150.0909367 | pos | - | Synephrine | C9H13NO2 | Alkaloids | up |
| 36 | 1.3156 | 1.057266667 | 298.1280128 | pos | - | Phenethylamine glucuronide | C14H19NO6 | Others | down |
| 37 | 1.2997 | 9.073533333 | 411.3250925 | pos | 114020-59-6 | (6α,22E)-6-Hydroxy-4,7,22-ergostatrien-3-1 | C28H42O2 | Lipids | down |
| 38 | 1.2961 | 2.09015 | 189.1384483 | pos | - | N, N-Dimethyltryptamine | C12H16N2 | Alkaloids | down |
| 39 | 1.2921 | 8.676833333 | 380.3153089 | pos | - | N-linoleoyl valine | C23H41NO3 | Organic acid | down |
| 40 | 1.2772 | 8.154433333 | 184.0730015 | pos | 107-73-3 | Phosphocholine | C5H14NO4P | Others | up |
| 41 | 1.2761 | 8.536833333 | 366.2986914 | pos | - | Dihydroceramide C2 | C20H41NO3 | Lipids | down |
| 42 | 1.2722 | 7.951316667 | 205.1947673 | pos | - | (−)-α-Cedrene | C15H24 | Lipids | down |
| 43 | 1.2708 | 2.034916667 | 114.0913457 | pos | - | N-Acetylpyrrolidine | C6H11NO | Others | up |
| 44 | 1.2635 | 4.89415 | 261.1113648 | pos | - | (R)-Meranzin | C15H16O4 | Phenylpropanoids | up |
| 45 | 1.2584 | 2.672183333 | 234.0755659 | pos | - | 2-Hydroxy-6-oxo-(2′-aminophenyl)-hexa-2,4-dienoate | C12H11NO4 | Others | up |
| 46 | 1.2571 | 8.154433333 | 496.3388062 | pos | 17364-16-8 | PC (16:0/0:0) | C24H50NO7P | Lipids | up |
| 47 | 1.2396 | 5.039233333 | 137.1321248 | pos | 13466-78-9 | (+)-3-Carene | C10H16 | Lipids | up |
| 48 | 1.2385 | 5.97185 | 235.1687792 | pos | - | Macrophyllic acid A | C15H22O2 | Terpenes | down |
| 49 | 1.2343 | 5.3979 | 144.0805222 | pos | 91-59-8 | 2-Naphthylamine | C10H9N | Others | down |
| 50 | 1.2258 | 8.016083333 | 323.2559911 | neg | - | 11(R)-HEDE | C20H36O3 | Lipids | down |
| 51 | 1.2101 | 2.617466667 | 331.1280568 | pos | 5589-85-5 | Miraxanthin-III | C17H18N2O5 | Others | up |
| 52 | 1.2098 | 10.02308333 | 580.5293196 | pos | - | Cer(d16:2(4E,6E)/20:0(2OH)) | C36H69NO4 | Lipids | down |
| 53 | 1.2006 | 3.58255 | 433.1120777 | pos | - | Genistein 4′-O-glucoside | C21H20O10 | Flavonoids | up |
| 54 | 1.1945 | 5.686116667 | 309.2041751 | neg | 111004-08-1 | 9(S)-HpOTrE | C18H30O4 | Lipids | down |
| 55 | 1.1940 | 2.4533 | 347.1229536 | pos | 5375-64-4 | Miraxanthin-V | C17H18N2O6 | Others | up |
| 56 | 1.1890 | 3.944766667 | 271.0591982 | pos | - | 4,6,4′-Trihydroxyaurone | C15H10O5 | Flavonoids | up |
| 57 | 1.1814 | 7.526 | 133.0606782 | pos | 70-47-3 | L-Asparagine | C4H8N2O3 | Organic acid | down |
| 58 | 1.1805 | 8.676833333 | 118.0862446 | pos | 2566-34-9 | N,2-dimethylalanine | C5H11NO2 | Organic acid | down |
| 59 | 1.1795 | 7.78705 | 482.3232773 | pos | - | PC (15:0/0:0) | C23H48NO7P | Lipids | up |
| 60 | 1.1652 | 4.83305 | 387.104879 | neg | 479-90-3 | Artemetin | C20H20O8 | Flavonoids | down |
| 61 | 1.1645 | 8.133916667 | 138.0911354 | pos | 104-87-0 | P-Tolualdehyde | C8H8O | Others | down |
| 62 | 1.1630 | 4.639183333 | 489.2113061 | pos | 1180-71-8 | Limonoate D-ring-lactone | C26H32O9 | Lipids | down |
| 63 | 1.1538 | 6.091066667 | 219.1738452 | pos | 37208-05-2 | Capsidiol | C15H24O2 | Lipids | down |
| 64 | 1.1516 | 4.89415 | 175.0386137 | pos | 34328-51-3 | Artemidinal | C10H6O3 | Phenylpropanoids | down |
| 65 | 1.1503 | 2.143683333 | 274.1430781 | pos | - | Sanguinine | C16H19NO3 | Alkaloids | up |
| 66 | 1.1446 | 8.69595 | 326.304721 | pos | 111-58-0 | Oleoyl Ethanolamide | C20H39NO2 | Others | down |
| 67 | 1.1418 | 2.690183333 | 164.0703438 | pos | - | 5-Hydroxy-3,4-dihydrocarbostyryl | C9H9NO2 | Others | down |
| 68 | 1.1383 | 3.347783333 | 509.1282039 | pos | - | Syringetin 3-glucoside | C23H24O13 | Flavonoids | down |
| 69 | 1.1359 | 7.78705 | 520.3390665 | pos | - | PC (18:2/0:0) | C26H50NO7P | Lipids | up |
| 70 | 1.1268 | 5.003066667 | 181.1219295 | pos | 59300-52-6 | Norecasantalic acid | C11H16O2 | Lipids | down |
| 71 | 1.1218 | 3.365666667 | 247.0957688 | pos | 23458-02-8 | Decursinol | C14H14O4 | Phenylpropanoids | up |
| 72 | 1.1160 | 0.7165 | 132.1017484 | pos | β-Alaninebetaine | C6H13NO2 | Amino acids | up | |
| 73 | 1.1116 | 10.06556667 | 554.513563 | pos | - | Cer(d14:1(4E)/20:0(2OH)) | C34H67NO4 | Lipids | down |
| 74 | 1.1115 | 8.1742 | 454.2919431 | pos | 53862-35-4 | PE (16:0/0:0) | C21H44NO7P | Lipids | up |
| 75 | 1.1065 | 3.0777 | 339.1066468 | pos | 39015-63-9 | α-Hydrojuglone 4-O-β-D-glucoside | C16H18O8 | Others | up |
| 76 | 1.1059 | 8.457933333 | 166.0860751 | pos | - | 2-Propylisonicotinic acid | C9H11NO2 | Others | down |
| 77 | 1.1033 | 2.842416667 | 192.0651578 | pos | 54-16-0 | 5-Hydroxyindoleacetic acid | C10H9NO3 | Others | up |
| 78 | 1.1012 | 8.27685 | 480.3078345 | pos | - | PE (18:1(9Z)/0:0) | C23H46NO7P | Lipids | up |
| 79 | 1.0985 | 1.860016667 | 353.0843828 | neg | 327-97-9 | Chlorogenic Acid | C16H18O9 | Others | up |
| 80 | 1.0931 | 9.63365 | 797.5154884 | pos | - | MGDG (18:3/18:3) | C45H74O10 | Lipids | up |
| 81 | 1.0903 | 8.536833333 | 132.1017796 | pos | 1509-34-8 | L-Alloisoleucine | C6H13NO2 | Organic acid | down |
| 82 | 1.0854 | 7.465616667 | 279.2313458 | pos | - | 13(S)-HODE | C18H32O3 | Lipids | down |
| 83 | 1.0834 | 7.541266667 | 293.2095425 | neg | - | 13(S)-HOTrE | C18H30O3 | Lipids | down |
| 84 | 1.0788 | 6.025033333 | 263.1263265 | neg | 178167-75-4 | γ-CEHC | C15H20O4 | Others | down |
| 85 | 1.0757 | 7.848633333 | 442.2946433 | pos | - | N-(α-Linolenoyl) Tyrosine | C27H39NO4 | Lipids | down |
| 86 | 1.0600 | 5.603583333 | 275.1999899 | pos | - | 6-[5]-ladderane-hexanoic acid | C18H26O2 | Lipids | down |
| 87 | 1.0597 | 8.27685 | 522.3547486 | pos | - | PC (18:1/0:0) | C26H52NO7P | Lipids | up |
| 88 | 1.0574 | 4.785016667 | 301.0698558 | pos | 520-34-3 | Diosmetin | C16H12O6 | Flavonoids | up |
| 89 | 1.0526 | 8.594583333 | 300.2891351 | pos | 544-31-0 | Palmitoyl Ethanolamide | C18H37NO2 | Organic acid | down |
| 90 | 1.0239 | 3.2759 | 611.1962817 | pos | 13241-33-3 | Neohesperidin | C28H34O15 | Flavonoids | down |
| 91 | 1.0220 | 4.584333333 | 187.0385834 | pos | - | Pyrogallin | C11H8O4 | Others | up |
| 92 | 1.0214 | 7.227266667 | 285.2043948 | neg | 42150-38-9 | Hexadecanedioic acid | C16H30O4 | Lipids | down |
| 93 | 1.0207 | 0.68085 | 160.096405 | pos | 515-25-3 | (−)-Betonicine | C7H13NO3 | Organic acid | up |
| 94 | 1.0060 | 4.475616667 | 133.1010276 | pos | 1195-32-0 | 4-Isopropenyltoluene | C10H12 | Others | down |
| 95 | 1.0030 | 7.828316667 | 478.292056 | pos | - | PE (18:2/0:0) | C23H44NO7P | Lipids | up |
| 96 | 1.0011 | 2.672183333 | 274.106699 | pos | 1596-67-4 | L-Thyronine | C15H15NO4 | Organic acid | up |
| 97 | 1.0010 | 8.564383333 | 312.2514222 | neg | 158305-64-7 | N-Palmitoyl Glycine | C18H35NO3 | Organic acid | down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Peng, M.; Gao, Z.; Han, Q.; Fu, F.; Li, G.; Su, D.; Huang, L.; Guo, J.; Shan, Y. Untargeted Metabolomic Analyses and Antilipidemic Effects of Citrus Physiological Premature Fruit Drop. Int. J. Mol. Sci. 2024, 25, 1876. https://doi.org/10.3390/ijms25031876
Wang C, Peng M, Gao Z, Han Q, Fu F, Li G, Su D, Huang L, Guo J, Shan Y. Untargeted Metabolomic Analyses and Antilipidemic Effects of Citrus Physiological Premature Fruit Drop. International Journal of Molecular Sciences. 2024; 25(3):1876. https://doi.org/10.3390/ijms25031876
Chicago/Turabian StyleWang, Chao, Mingfang Peng, Zhipeng Gao, Qi Han, Fuhua Fu, Gaoyang Li, Donglin Su, Lvhong Huang, Jiajing Guo, and Yang Shan. 2024. "Untargeted Metabolomic Analyses and Antilipidemic Effects of Citrus Physiological Premature Fruit Drop" International Journal of Molecular Sciences 25, no. 3: 1876. https://doi.org/10.3390/ijms25031876
APA StyleWang, C., Peng, M., Gao, Z., Han, Q., Fu, F., Li, G., Su, D., Huang, L., Guo, J., & Shan, Y. (2024). Untargeted Metabolomic Analyses and Antilipidemic Effects of Citrus Physiological Premature Fruit Drop. International Journal of Molecular Sciences, 25(3), 1876. https://doi.org/10.3390/ijms25031876