
Chronic Morphine Treatment and Antiretroviral Therapy Exacerbate HIV-Distal Sensory Peripheral Neuropathy and Induce Distinct Microbial Alterations in the HIV Tg26 Mouse Model
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
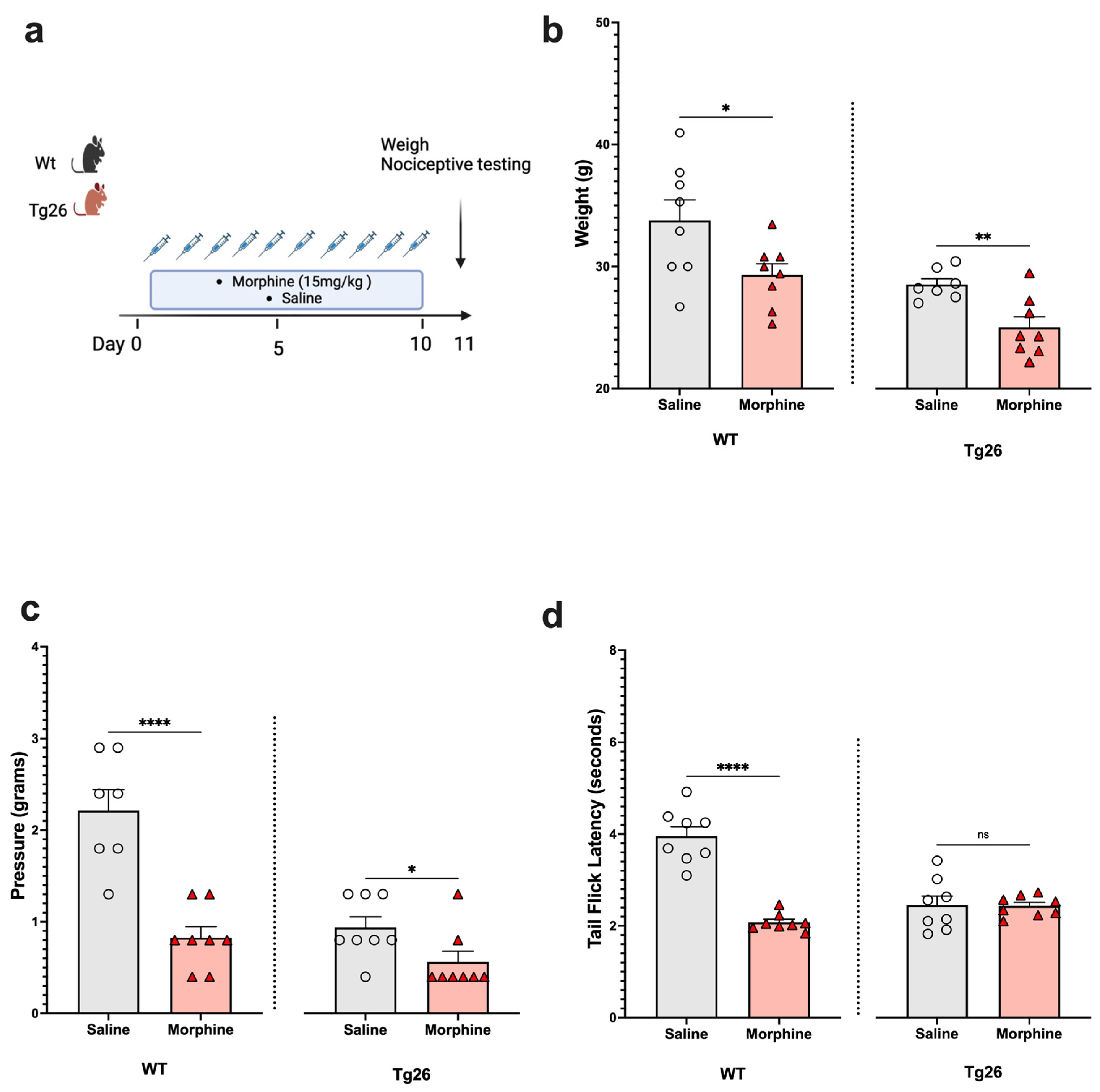
2.1. Chronic Morphine Treatment Worsened the Development of Peripheral Neuropathy through Mechanical Pain Pathways in Tg26 Mice Compared to WT
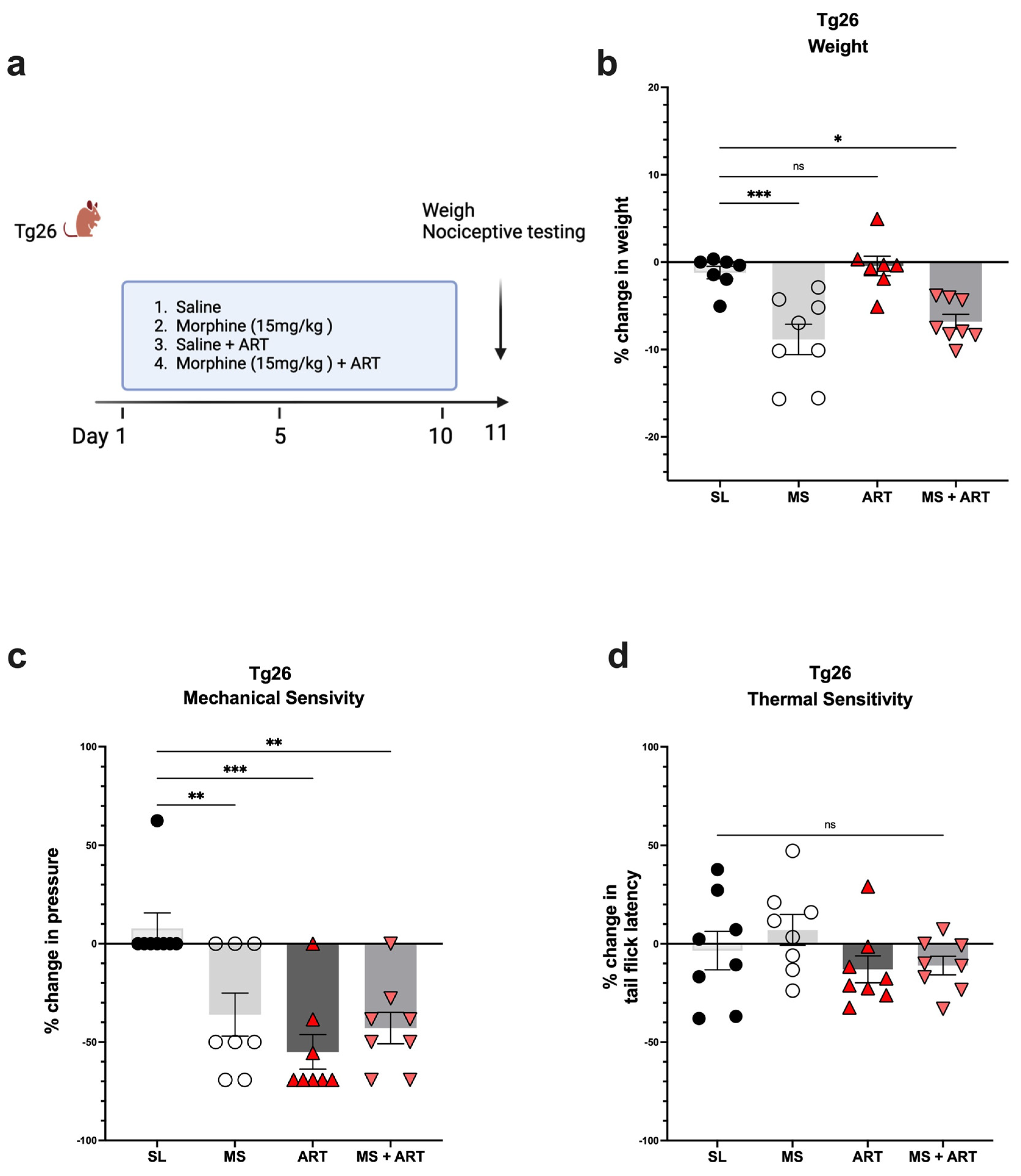
2.2. ART Induced Peripheral Neuropathy through Mechanical Pain Pathways and Worsened Analgesic Tolerance to Morphine in Tg26 Mice
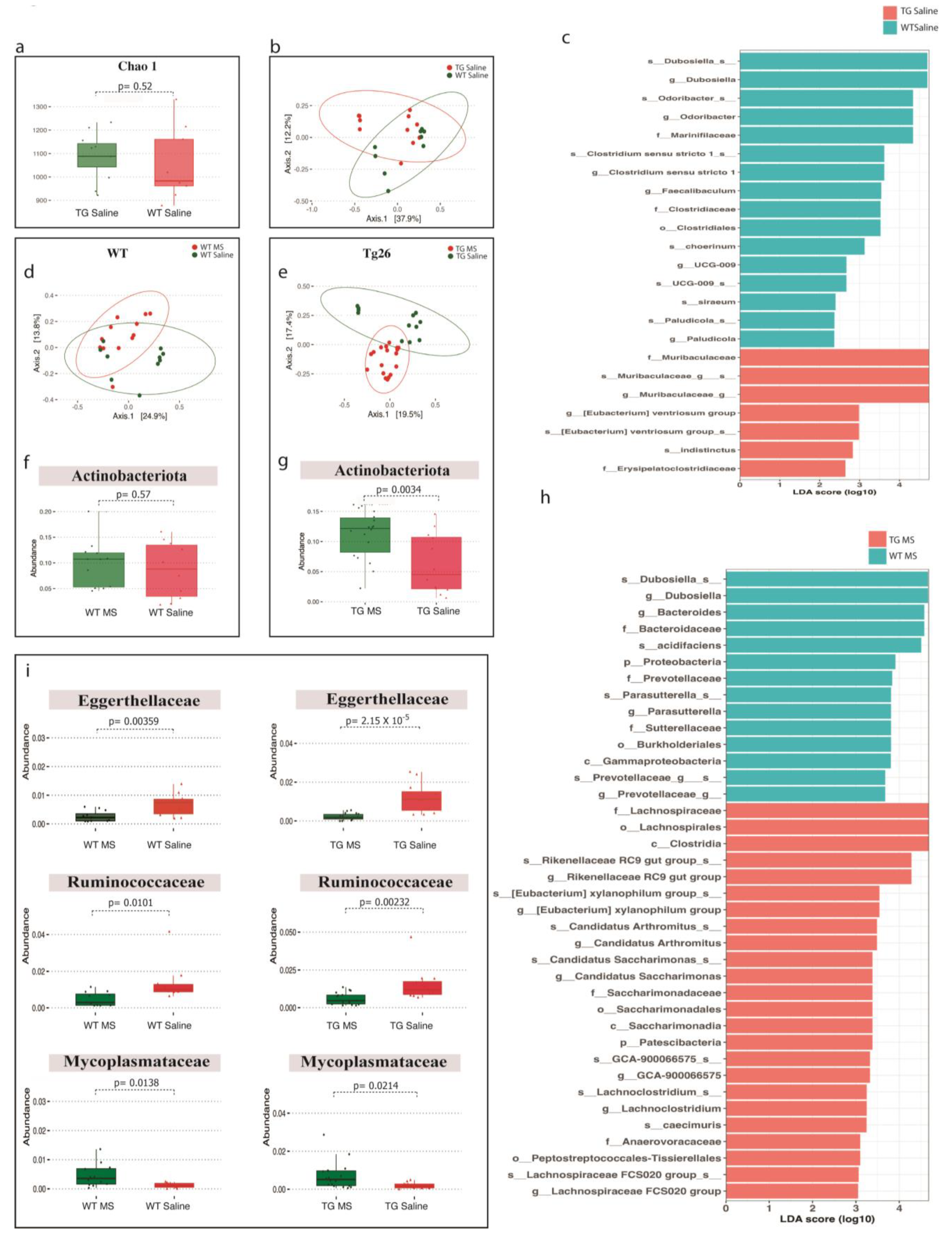
2.3. Distinct Bacterial Communities Are Observed between WT and Tg26 Mice before and after Morphine Treatment
2.4. ART Treatment Modulates the Gut Microbiome in WT and Tg26 Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Animal Treatments
4.2.1. Morphine Treatment
4.2.2. ART Treatment
4.3. Nociceptive Testing
4.3.1. Tail Flick
4.3.2. Von Frey
4.4. 16S rRNA Gene Sequencing
4.5. Microbiome Sequencing Data Analysis
4.6. Statistical Analysis and Diversity Metrics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centner, C.M.; Bateman, K.J.; Heckmann, J.M. Manifestations of HIV infection in the peripheral nervous system. Lancet Neurol. 2013, 12, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Aziz-Donnelly, A.; Harrison, T.B. Update of HIV-Associated Sensory Neuropathies. Curr. Treat. Options Neurol. 2017, 19, 36. [Google Scholar] [CrossRef] [PubMed]
- Robinson-Papp, J.; Schütz, S.G. HIV-related neuropathy: Current perspectives. HIV/AIDS Res. Palliat. Care 2013, 5, 243–251. [Google Scholar] [CrossRef]
- Anastasi, J.K.; Pakhomova, A.M. Assessment and Management of HIV Distal Sensory Peripheral Neuropathy: Understanding the Symptoms. J. Nurse Pract. 2020, 16, 276–280. [Google Scholar] [CrossRef]
- Tumusiime, D.K.; Venter, F.; Musenge, E.; Stewart, A. Prevalence of peripheral neuropathy and its associated demographic and health status characteristics, among people on antiretroviral therapy in Rwanda. BMC Public Health 2014, 14, 1306. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Rosario, D.; Clifford, D.B.; McArthur, J.C.; Simpson, D.; Alexander, T.; Gelman, B.B.; Vaida, F.; Collier, A.; Marra, C.M.; et al. Continued High Prevalence and Adverse Clinical Impact of Human Immunodeficiency Virus–Associated Sensory Neuropathy in the Era of Combination Antiretroviral Therapy. Arch. Neurol. 2010, 67, 552–558. [Google Scholar] [CrossRef]
- Dorsey, S.G.; Morton, P.G. HIV peripheral neuropathy: Pathophysiology and clinical implications. AACN Adv. Crit. Care 2006, 17, 30–36. [Google Scholar] [CrossRef]
- Phillips, T.J.C.; Cherry, C.L.; Cox, S.; Marshall, S.J.; Rice, A.S.C. Pharmacological Treatment of Painful HIV-Associated Sensory Neuropathy: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. PLoS ONE 2010, 5, e14433. [Google Scholar] [CrossRef]
- Krashin, D. Opioids in the Management of HIV-Related Pain. IBJ Plus 2012, 15, ES157–ES168. [Google Scholar] [CrossRef]
- Liu, B.; Liu, X.; Tang, S.-J. Interactions of Opioids and HIV Infection in the Pathogenesis of Chronic Pain. Front. Microbiol. 2016, 7, 103. [Google Scholar] [CrossRef]
- Satish, S.; Abu, Y.; Gomez, D.; Dutta, R.K.; Roy, S. HIV, opioid use, and alterations to the gut microbiome: Elucidating independent and synergistic effects. Front. Immunol. 2023, 14, 1156862. [Google Scholar] [CrossRef]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef]
- Kinross, J.M.; Darzi, A.W.; Nicholson, J.K. Gut microbiome-host interactions in health and disease. Genome Med. 2011, 3, 14. [Google Scholar] [CrossRef]
- Jalodia, R.; Kolli, U.; Braniff, R.G.; Tao, J.; Abu, Y.F.; Chupikova, I.; Moidunny, S.; Ramakrishnan, S.; Roy, S. Morphine mediated neutrophil infiltration in intestinal tissue play essential role in histological damage and microbial dysbiosis. Gut Microbes 2022, 14, 2143225. [Google Scholar] [CrossRef]
- Banerjee, S.; Sindberg, G.; Wang, F.; Meng, J.; Sharma, U.; Zhang, L.; Dauer, P.; Chen, C.; Dalluge, J.; Johnson, T.; et al. Opioid-induced gut microbial disruption and bile dysregulation leads to gut barrier compromise and sustained systemic inflammation. Mucosal Immunol. 2016, 9, 1418–1428. [Google Scholar] [CrossRef] [PubMed]
- Sindberg, G.M.; Callen, S.E.; Banerjee, S.; Meng, J.; Hale, V.L.; Hegde, R.; Cheney, P.D.; Villinger, F.; Roy, S.; Buch, S. Morphine Potentiates Dysbiotic Microbial and Metabolic Shifts in Acute SIV Infection. J. Neuroimmune Pharmacol. 2018, 14, 200–214. [Google Scholar] [CrossRef]
- Acharya, C.; Betrapally, N.S.; Gillevet, P.M.; Sterling, R.K.; Akbarali, H.; White, M.B.; Ganapathy, D.; Fagan, A.; Sikaroodi, M.; Bajaj, J.S. Chronic opioid use is associated with altered gut microbiota and predicts readmissions in patients with cirrhosis. Aliment. Pharmacol. Ther. 2016, 45, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xie, Z.; Wang, H.; Shen, Z.; Guo, Y.; Gao, Y.; Chen, X.; Wu, Q.; Li, X.; Wang, K. Bacterial Diversity of Intestinal Microbiota in Patients with Substance Use Disorders Revealed by 16S rRNA Gene Deep Sequencing. Sci. Rep. 2017, 7, 3628. [Google Scholar] [CrossRef]
- Meng, J.; Yu, H.; Ma, J.; Wang, J.; Banerjee, S.; Charboneau, R.; Barke, R.A.; Roy, S. Morphine Induces Bacterial Translocation in Mice by Compromising Intestinal Barrier Function in a TLR-Dependent Manner. PLoS ONE 2013, 8, e54040. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.; Wilck, N.; Haghikia, A.; Gold, R.; Mueller, D.N.; Linker, R.A. The role of the gut microbiota and microbial metabolites in neuroinflammation. Eur. J. Immunol. 2020, 50, 1863–1870. [Google Scholar] [CrossRef]
- Brunt, V.E.; LaRocca, T.J.; Bazzoni, A.E.; Sapinsley, Z.J.; Miyamoto-Ditmon, J.; Gioscia-Ryan, R.A.; Neilson, A.P.; Link, C.D.; Seals, D.R. The gut microbiome–derived metabolite trimethylamine N-oxide modulates neuroinflammation and cognitive function with aging. GeroScience 2020, 43, 377–394. [Google Scholar] [CrossRef]
- Quinn, M.; McMillin, M.; Galindo, C.; Frampton, G.; Pae, H.Y.; DeMorrow, S. Bile acids permeabilize the blood brain barrier after bile duct ligation in rats via Rac1-dependent mechanisms. Dig. Liver Dis. 2014, 46, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic-Cvijin, I.; Dunham, R.M.; Iwai, S.; Maher, M.C.; Albright, R.G.; Broadhurst, M.J.; Hernandez, R.D.; Lederman, M.M.; Huang, Y.; Somsouk, M.; et al. Dysbiosis of the Gut Microbiota Is Associated with HIV Disease Progression and Tryptophan Catabolism. Sci. Transl. Med. 2013, 5, 193ra91. [Google Scholar] [CrossRef]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Sandler, N.G.; Wand, H.; Roque, A.; Law, M.; Nason, M.C.; Nixon, D.E.; Pedersen, C.; Ruxrungtham, K.; Lewin, S.R.; Emery, S.; et al. Plasma Levels of Soluble CD14 Independently Predict Mortality in HIV Infection. J. Infect. Dis. 2011, 203, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Banerjee, S.; Zhang, L.; Sindberg, G.; Moidunny, S.; Li, B.; Robbins, D.J.; Girotra, M.; Segura, B.; Ramakrishnan, S.; et al. Opioids Impair Intestinal Epithelial Repair in HIV-Infected Humanized Mice. Front. Immunol. 2020, 10, 2999. [Google Scholar] [CrossRef]
- Ancuta, P.; Kamat, A.; Kunstman, K.J.; Kim, E.-Y.; Autissier, P.; Wurcel, A.; Zaman, T.; Stone, D.; Mefford, M.; Morgello, S.; et al. Microbial Translocation Is Associated with Increased Monocyte Activation and Dementia in AIDS Patients. PLoS ONE 2008, 3, e2516. [Google Scholar] [CrossRef] [PubMed]
- Kopp, J.B.; E Klotman, M.; Adler, S.H.; A Bruggeman, L.; Dickie, P.; Marinos, N.J.; Eckhaus, M.; Bryant, J.L.; Notkins, A.L.; E Klotman, P. Progressive glomerulosclerosis and enhanced renal accumulation of basement membrane components in mice transgenic for human immunodeficiency virus type 1 genes. Proc. Natl. Acad. Sci. USA 1992, 89, 1577–1581. [Google Scholar] [CrossRef]
- Barbe, M.F.; Loomis, R.; Lepkowsky, A.M.; Forman, S.; Zhao, H.; Gordon, J. A longitudinal characterization of sex-specific somatosensory and spatial memory deficits in HIV Tg26 heterozygous mice. PLoS ONE 2020, 15, e0244725. [Google Scholar] [CrossRef]
- Putatunda, R.; Zhang, Y.; Li, F.; Fagan, P.R.; Zhao, H.; Ramirez, S.H.; Praticò, D.; Barbe, M.F.; Hu, W. Sex-specific neurogenic deficits and neurocognitive disorders in middle-aged HIV-1 Tg26 transgenic mice. Brain Behav. Immun. 2019, 80, 488–499. [Google Scholar] [CrossRef]
- Sagen, J.; Hama, A.T.; Castellanos, D. Antinociceptive effects of topical mepivacaine in a rat model of HIV-associated peripheral neuropathic pain. J. Pain Res. 2016, 9, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Guindon, J.; Blanton, H.; Brauman, S.; Donckels, K.; Narasimhan, M.; Benamar, K. Sex Differences in a Rodent Model of HIV-1-Associated Neuropathic Pain. Int. J. Mol. Sci. 2019, 20, 1196. [Google Scholar] [CrossRef] [PubMed]
- Toma, W.; Paris, J.J.; Warncke, U.O.; Nass, S.R.; Caillaud, M.; McKiver, B.; Ondo, O.; Bagdas, D.; Bigbee, J.; Knapp, P.E.; et al. Persistent sensory changes and sex differences in transgenic mice conditionally expressing HIV-1 Tat regulatory protein. Exp. Neurol. 2022, 358, 114226. [Google Scholar] [CrossRef] [PubMed]
- Wodarski, R.; Bagdas, D.; Paris, J.J.; Pheby, T.; Toma, W.; Xu, R.; Damaj, M.I.; Knapp, P.E.; Rice, A.S.; Hauser, K.F. Reduced intraepidermal nerve fibre density, glial activation, and sensory changes in HIV type-1 Tat-expressing female mice: Involvement of Tat during early stages of HIV-associated painful sensory neuropathy. PAIN Rep. 2018, 3, e654. [Google Scholar] [CrossRef] [PubMed]
- Bagdas, D.; Paris, J.J.; Carper, M.; Wodarski, R.; Rice, A.S.C.; Knapp, P.E.; Hauser, K.F.; Damaj, M.I. Conditional expression of HIV-1 tat in the mouse alters the onset and progression of tonic, inflammatory and neuropathic hypersensitivity in a sex-dependent manner. Eur. J. Pain 2020, 24, 1609–1623. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Umlauf, A.; Letendre, S.; Franklin, D.; Bush, W.S.; Atkinson, J.H.; Keltner, J.; Ellis, R.J. Catechol-O-methyltransferase polymorphism Val158Met is associated with distal neuropathic pain in HIV-associated sensory neuropathy. AIDS 2019, 33, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yuan, S.; Tang, S.-J. Morphine and HIV-1 gp120 cooperatively promote pathogenesis in the spinal pain neural circuit. Mol. Pain 2019, 15, 1744806919868380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.J.; Qiu, Y.; Hua, Z. The Emerging Perspective of Morphine Tolerance: MicroRNAs. Pain Res. Manag. 2019, 2019, 9432965. [Google Scholar] [CrossRef]
- Volkow, N.D.; McLellan, A.T. Opioid Abuse in Chronic Pain—Misconceptions and Mitigation Strategies. N. Engl. J. Med. 2016, 374, 1253–1263. [Google Scholar] [CrossRef]
- Önen, N.F.; Barrette, E.; Shacham, E.; Taniguchi, T.; Donovan, M.; Overton, E.T. A Review of Opioid Prescribing Practices and Associations with Repeat Opioid Prescriptions in a Contemporary Outpatient HIV Clinic. Pain Pract. 2011, 12, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Smith, H. Treatment Considerations in Painful HIV-Related Neuropathy. IBJ Plus 2011, 14, E505–E524. [Google Scholar] [CrossRef]
- Ray, S.; Sil, S.; Kannan, M.; Periyasamy, P.; Buch, S. Role of the gut-brain axis in HIV and drug abuse-mediated neuroinflammation. Adv. Drug Alcohol Res. 2023, 3, 11092. [Google Scholar] [CrossRef]
- Li, L.; Zhao, X.; He, J.J. HIV Tat Expression and Cocaine Exposure Lead to Sex- and Age-Specific Changes of the Microbiota Composition in the Gut. Microorganisms 2023, 11, 799. [Google Scholar] [CrossRef]
- He, N.; Yang, Y.; Wang, H.; Liu, N.; Yang, Z.; Li, S. Unsaturated alginate oligosaccharides (UAOS) protects against dextran sulfate sodium-induced colitis associated with regulation of gut microbiota. J. Funct. Foods 2021, 83, 104536. [Google Scholar] [CrossRef]
- Quraishi, M.N.; Sergeant, M.; Kay, G.; Iqbal, T.; Chan, J.; Constantinidou, C.; Trivedi, P.; Ferguson, J.; Adams, D.H.; Pallen, M.; et al. The gut-adherent microbiota of PSC–IBD is distinct to that of IBD. Gut 2016, 66, 386–388. [Google Scholar] [CrossRef]
- Huang, F.; Pan, H.; Tan, Z.; Chen, L.; Li, T.; Liu, Y. Prevotella histicola Prevented Particle-Induced Osteolysis via Gut Microbiota-Dependent Modulation of Inflammation in Ti-Treated Mice. Probiotics Antimicrob. Proteins, 2023; ahead of print. [Google Scholar] [CrossRef]
- Camelo-Castillo, A.J.; Mira, A.; Pico, A.; Nibali, L.; Henderson, B.; Donos, N.; Tomã¡S, I. Subgingival microbiota in health compared to periodontitis and the influence of smoking. Front. Microbiol. 2015, 6, 119. [Google Scholar] [CrossRef]
- Ray, S.; Narayanan, A.; Giske, C.G.; Neogi, U.; Sönnerborg, A.; Nowak, P. Altered Gut Microbiome under Antiretroviral Therapy: Impact of Efavirenz and Zidovudine. ACS Infect. Dis. 2020, 7, 1104–1115. [Google Scholar] [CrossRef]
- Ishizaka, A.; Koga, M.; Mizutani, T.; Parbie, P.K.; Prawisuda, D.; Yusa, N.; Sedohara, A.; Kikuchi, T.; Ikeuchi, K.; Adachi, E.; et al. Unique Gut Microbiome in HIV Patients on Antiretroviral Therapy (ART) Suggests Association with Chronic Inflammation. Microbiol. Spectr. 2021, 9, e0070821. [Google Scholar] [CrossRef] [PubMed]
- Imahashi, M.; Ode, H.; Kobayashi, A.; Nemoto, M.; Matsuda, M.; Hashiba, C.; Hamano, A.; Nakata, Y.; Mori, M.; Seko, K.; et al. Impact of long-term antiretroviral therapy on gut and oral microbiotas in HIV-1-infected patients. Sci. Rep. 2021, 11, 960. [Google Scholar] [CrossRef] [PubMed]
- Villar-García, J.; Güerri-Fernández, R.; Moya, A.; González, A.; Hernández, J.J.; Lerma, E.; Guelar, A.; Sorli, L.; Horcajada, J.P.; Artacho, A.; et al. Impact of probiotic Saccharomyces boulardii on the gut microbiome composition in HIV-treated patients: A double-blind, randomised, placebo-controlled trial. PLoS ONE 2017, 12, e0173802. [Google Scholar] [CrossRef]
- Graf, J. The family rikenellaceae. In The Prokaryotes: Other Major Lineages of Bacteria and The Archaea; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Lin, H.; Meng, L.; Sun, Z.; Sun, S.; Huang, X.; Lin, N.; Zhang, J.; Lu, W.; Yang, Q.; Chi, J.; et al. Yellow Wine Polyphenolic Compound Protects Against Doxorubicin-Induced Cardiotoxicity by Modulating the Composition and Metabolic Function of the Gut Microbiota. Circ. Hearth Fail. 2021, 14, e008220. [Google Scholar] [CrossRef]
- Peters, B.A.; Shapiro, J.A.; Church, T.R.; Miller, G.; Trinh-Shevrin, C.; Yuen, E.; Friedlander, C.; Hayes, R.B.; Ahn, J. A taxonomic signature of obesity in a large study of American adults. Sci. Rep. 2018, 8, 9749. [Google Scholar] [CrossRef]
- Mutlu, E.A.; Keshavarzian, A.; Losurdo, J.; Swanson, G.; Siewe, B.; Forsyth, C.; French, A.; DeMarais, P.; Sun, Y.; Koenig, L.; et al. A Compositional Look at the Human Gastrointestinal Microbiome and Immune Activation Parameters in HIV Infected Subjects. PLoS Pathog. 2014, 10, e1003829. [Google Scholar] [CrossRef] [PubMed]
- Dinh, D.M.; Volpe, G.E.; Duffalo, C.; Bhalchandra, S.; Tai, A.K.; Kane, A.V.; Wanke, C.A.; Ward, H.D. Intestinal Microbiota, Microbial Translocation, and Systemic Inflammation in Chronic HIV Infection. J. Infect. Dis. 2014, 211, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Putatunda, R.; Zhang, Y.; Li, F.; Yang, X.-F.; Barbe, M.F.; Hu, W. Adult neurogenic deficits in HIV-1 Tg26 transgenic mice. J. Neuroinflammation 2018, 15, 287. [Google Scholar] [CrossRef]
- Dickie, P.; Felser, J.; Eckhaus, M.; Bryant, J.; Silver, J.; Marinos, N.; Notkins, A.L. HIV-associated nephropathy in transgenic mice expressing HIV-1 genes. Virology 1991, 185, 109–119. [Google Scholar] [CrossRef]
- Deuis, J.R.; Dvorakova, L.S.; Vetter, I. Methods Used to Evaluate Pain Behaviors in Rodents. Front. Mol. Neurosci. 2017, 10, 284. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2013, 42, D643–D648. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoine, D.; Chupikova, I.; Jalodia, R.; Singh, P.K.; Roy, S. Chronic Morphine Treatment and Antiretroviral Therapy Exacerbate HIV-Distal Sensory Peripheral Neuropathy and Induce Distinct Microbial Alterations in the HIV Tg26 Mouse Model. Int. J. Mol. Sci. 2024, 25, 1569. https://doi.org/10.3390/ijms25031569
Antoine D, Chupikova I, Jalodia R, Singh PK, Roy S. Chronic Morphine Treatment and Antiretroviral Therapy Exacerbate HIV-Distal Sensory Peripheral Neuropathy and Induce Distinct Microbial Alterations in the HIV Tg26 Mouse Model. International Journal of Molecular Sciences. 2024; 25(3):1569. https://doi.org/10.3390/ijms25031569
Chicago/Turabian StyleAntoine, Danielle, Irina Chupikova, Richa Jalodia, Praveen Kumar Singh, and Sabita Roy. 2024. "Chronic Morphine Treatment and Antiretroviral Therapy Exacerbate HIV-Distal Sensory Peripheral Neuropathy and Induce Distinct Microbial Alterations in the HIV Tg26 Mouse Model" International Journal of Molecular Sciences 25, no. 3: 1569. https://doi.org/10.3390/ijms25031569
APA StyleAntoine, D., Chupikova, I., Jalodia, R., Singh, P. K., & Roy, S. (2024). Chronic Morphine Treatment and Antiretroviral Therapy Exacerbate HIV-Distal Sensory Peripheral Neuropathy and Induce Distinct Microbial Alterations in the HIV Tg26 Mouse Model. International Journal of Molecular Sciences, 25(3), 1569. https://doi.org/10.3390/ijms25031569