
Impact of Heavy Metals on Cold Acclimation of Salix viminalis Roots


 , ,
, ,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results
2.1. Plant Material
2.1.1. Chlorophyll Fluorescence
2.1.2. Root Frost Tolerance
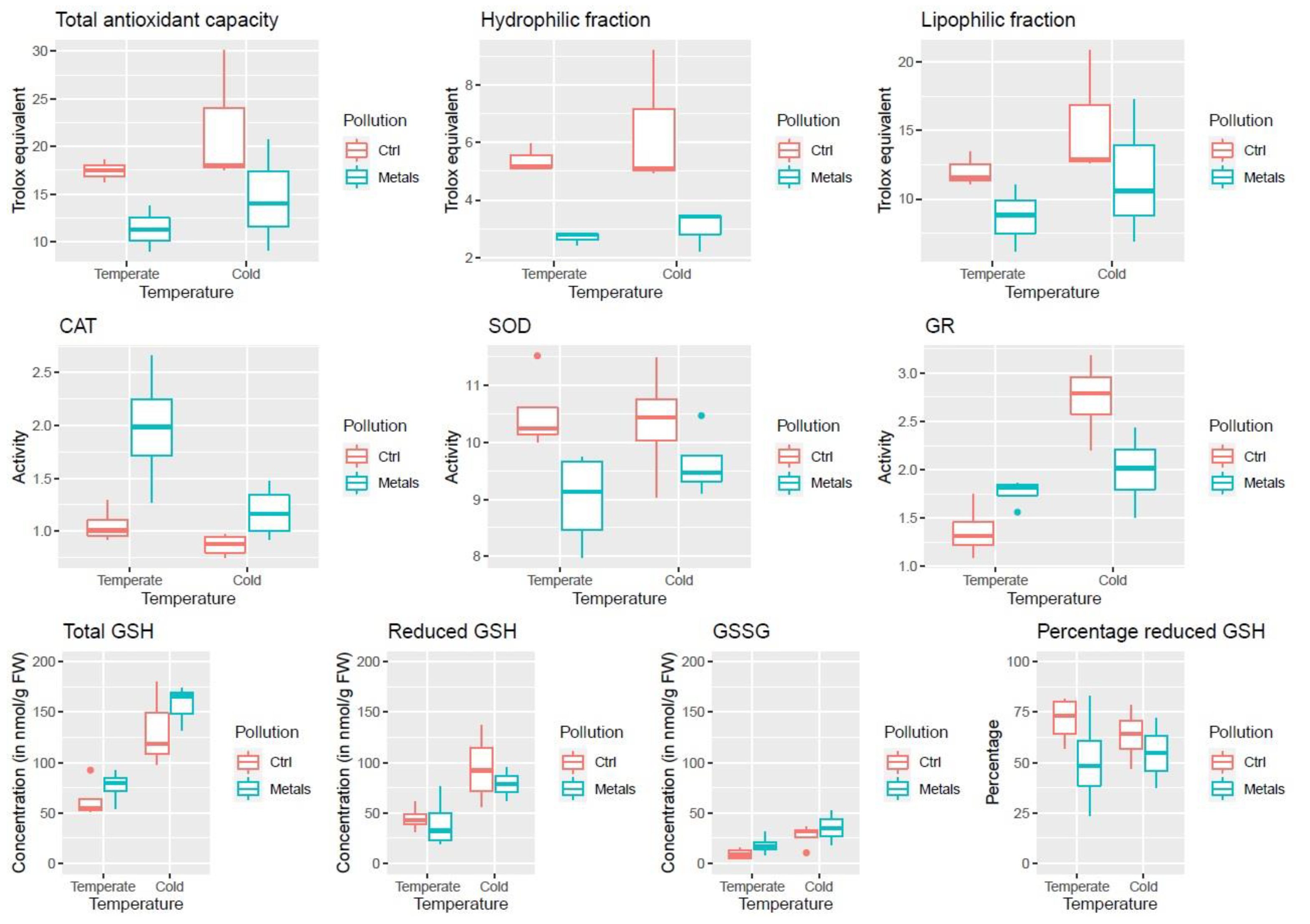
2.2. Impact on the Antioxidant System
2.2.1. Antioxidant Capacity
2.2.2. ROS Scavenging Enzymes Activity
2.2.3. Glutathione
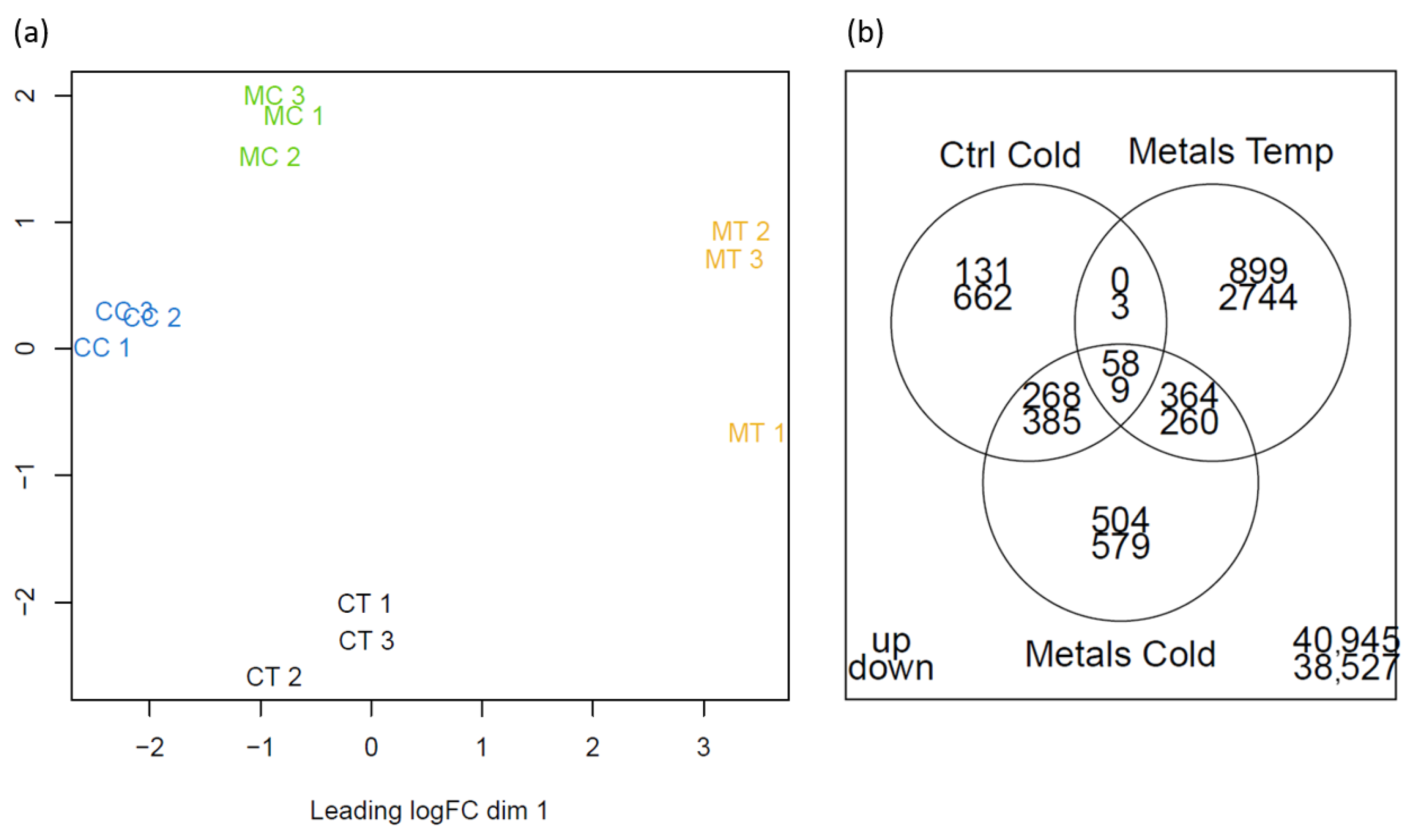
2.3. Transcriptomics
2.3.1. De Novo Transcriptome Assembly, Functional Annotation, and Mapping
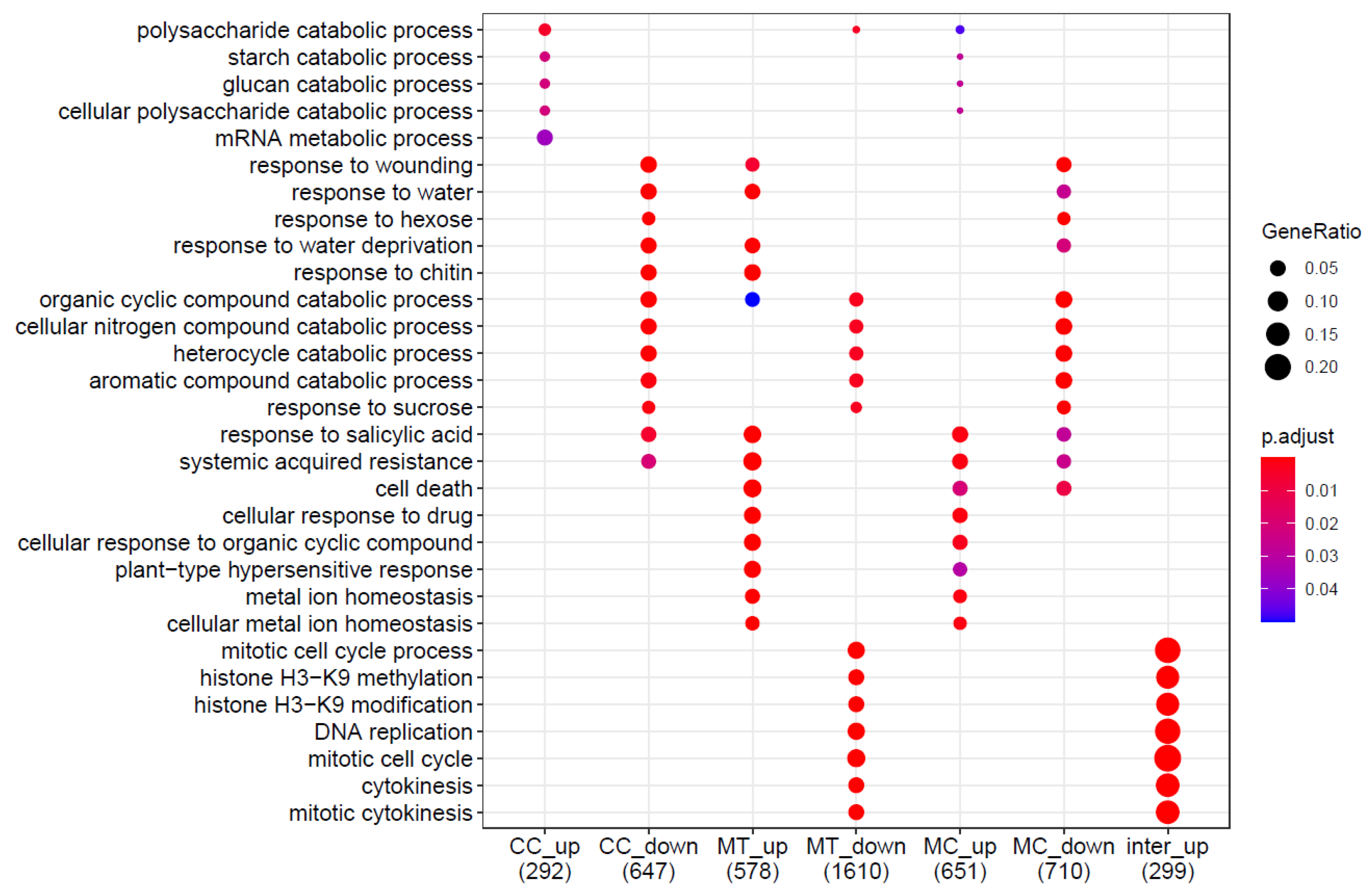
2.3.2. Differentially Expressed Genes
2.3.3. Gene Ontology Analysis
2.4. Proteomics
3. Discussion
3.1. Impact of Cold Acclimation
3.2. Impact of Long-Term Metal Exposure
3.3. Simultaneous Exposure to Cold and Metals
4. Materials and Methods
4.1. Plant Materials
4.2. Impact on the Antioxidant System
4.2.1. Antioxidant Capacity
4.2.2. ROS Scavenging Enzymes Activity
4.2.3. Glutathione Redox Couples
4.3. Transcriptomics
4.3.1. RNA extraction, Library Preparation, Sequencing, and De Novo Transcriptome Assembly
4.3.2. Differentially Expressed Genes and Gene Ontology Analysis
4.4. Proteomics
4.4.1. Protein Extraction and Gel-Free Proteomics
4.4.2. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lichtenthaler, H.K. The Stress Concept in Plants: An Introduction. Ann. N. Y. Acad. Sci. 1998, 851, 187–198. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When Defense Pathways Collide. The Response of Arabidopsis to a Combination of Drought and Heat Stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and Biotic Stress Combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Arruda, M.A.Z.; Azevedo, R.A. Metallomics and Chemical Speciation: Towards a Better Understanding of Metal-Induced Stress in Plants. Ann. Appl. Biol. 2009, 155, 301–307. [Google Scholar] [CrossRef]
- Hédiji, H.; Djebali, W.; Cabasson, C.; Maucourt, M.; Baldet, P.; Bertrand, A.; Boulila Zoghlami, L.; Deborde, C.; Moing, A.; Brouquisse, R.; et al. Effects of Long-Term Cadmium Exposure on Growth and Metabolomic Profile of Tomato Plants. Ecotoxicol. Environ. Saf. 2010, 73, 1965–1974. [Google Scholar] [CrossRef]
- Burke, M.J.; Gusta, L.V.; Quamme, H.A.; Weiser, C.J.; Li, P.H. Freezing and Injury in Plants. Plant Physiol. 1976, 27, 507–528. [Google Scholar] [CrossRef]
- Hossain, M.A.; Li, Z.G.; Hoque, T.S.; Burritt, D.J.; Fujita, M.; Munné-Bosch, S. Heat or Cold Priming-Induced Cross-Tolerance to Abiotic Stresses in Plants: Key Regulators and Possible Mechanisms. Protoplasma 2018, 255, 399–412. [Google Scholar] [CrossRef]
- Bielen, A.; Remans, T.; Vangronsveld, J.; Cuypers, A. The Influence of Metal Stress on the Availability and Redox State of Ascorbate, and Possible Interference with Its Cellular Functions. Int. J. Mol. Sci. 2013, 14, 6382–6413. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Dai, X.; Hu, Q.; Cai, Q.; Feng, K.; Ye, N.; Tuskan, G.A.; Milne, R.; Chen, Y.; Wan, Z.; Wang, Z.; et al. The Willow Genome and Divergent Evolution from Poplar after the Common Genome Duplication. Cell Res. 2014, 24, 1274–1277. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. Omi. A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Renaut, J.; Lutts, S.; Hoffmann, L.; Hausman, J.F. Responses of Poplar to Chilling Temperatures: Proteomic and Physiological Aspects. Plant Biol. 2004, 6, 81–90. [Google Scholar] [CrossRef]
- Bigras, F.J.; Paquin, R.; Rioux, J.A.; Therrien, H.P. Influence de La Photopériode et de La Température Sur l’évolution de La Tolérance Au Gel, de La Croissance et de La Teneur En Eau, Sucres, Amidon et Proline Des Rameaux et Des Racines de Genévrier (Juniperus Chinensis L. ’Pfitzerana’). Can. J. For. Res. 1989, 69, 305–316. [Google Scholar] [CrossRef]
- Sakai, A.; Larcher, W. (Eds.) Frost Survival of Plants: Responses and Adaptation to Freezing Stress; Sakai, A.; Larcher, W. (Eds.) Springer Science & Business Media: Berlin, Germany, 2012; ISBN 9783642717475. [Google Scholar]
- Ryyppö, A.; Repo, T.; Vapaavuori, E. Development of Freezing Tolerance in Roots and Shoots of Scots Pine Seedlings at Nonfreezing Temperatures. Can. J. For. Res. 1998, 28, 557–565. [Google Scholar] [CrossRef]
- Smit-Spinks, B.; Swanson, B.T.; Markhart III, A.H. Effect of Photoperiod and Thermoperiod on Cold Acclimation and Growth of Pinus Sylvestris. Can. J. For. Res. 1985, 15, 453–460. [Google Scholar] [CrossRef]
- Degand, H.; Faber, A.M.; Dauchot, N.; Mingeot, D.; Watillon, B.; Van Cutsem, P.; Morsomme, P.; Boutry, M. Proteomic Analysis of Chicory Root Identifies Proteins Typically Involved in Cold Acclimation. Proteomics 2009, 9, 2903–2907. [Google Scholar] [CrossRef] [PubMed]
- Regier, N.; Streb, S.; Zeeman, S.C.; Frey, B. Seasonal Changes in Starch and Sugar Content of Poplar (Populus Deltoides × Nigra Cv. Dorskamp) and the Impact of Stem Girdling on Carbohydrate Allocation to Roots. Tree Physiol. 2010, 30, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Ambroise, V.; Legay, S.; Guerriero, G.; Hausman, J.F.; Cuypers, A.; Sergeant, K. The Roots of Plant Frost Hardiness and Tolerance. Plant Cell Physiol. 2020, 61, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Ding, Y.; Shi, Y.; Zhang, X.; Gong, Z.; Yang, S. The Cbfs Triple Mutants Reveal the Essential Functions of CBFs in Cold Acclimation and Allow the Definition of CBF Regulons in Arabidopsis. New Phytol. 2016, 212, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Gilmour, S.J.; Grumet, R.; Thomashow, M.F. CBF-Dependent and CBF-Independent Regulatory Pathways Contribute to the Differences in Freezing Tolerance and Cold-Regulated Gene Expression of Two Arabidopsis Ecotypes Locally Adapted to Sites in Sweden and Italy. PLoS One 2018, 13, e0207723. [Google Scholar] [CrossRef] [PubMed]
- Kreps, J.A.; Wu, Y.; Chang, H.; Zhu, T.; Wang, X.; Harper, J.F.; Mesa, T.; Row, M.; Diego, S.; California, J.A.K.; et al. Transcriptome Changes for Arabidopsis in Response to Salt, Osmotic, and Cold Stress. Plant Physiol. 2002, 130, 2129–2141. [Google Scholar] [CrossRef] [PubMed]
- Scebba, F.; Sebastiani, L.; Vitagliano, C. Protective Enzymes against Activated Oxygen Species in Wheat (Triticum Aestivum L.) Seedlings: Responses to Cold Acclimation. J. Plant Physiol. 1999, 155, 762–768. [Google Scholar] [CrossRef]
- Nayyar, H.; Chander, S. Protective Effects of Polyamines against Oxidative Stress Induced by Water and Cold Stress in Chickpea. J. Agron. Crop Sci. 2004, 190, 335–365. [Google Scholar] [CrossRef]
- Zhao, S.; Blumwald, E. Changes in Oxidation-Redution State and Antioxidant Enzymes in the Root of Jack Pine Seeding during Cold Acclimation. Physiol. Plant. 1998, 104, 134–142. [Google Scholar] [CrossRef]
- Kieffer, P.; Dommes, J.; Hoffmann, L.; Hausman, J.F.; Renaut, J. Quantitative Changes in Protein Expression of Cadmium-Exposed Poplar Plants. Proteomics 2008, 8, 2514–2530. [Google Scholar] [CrossRef]
- Luo, Z.B.; He, J.; Polle, A.; Rennenberg, H. Heavy Metal Accumulation and Signal Transduction in Herbaceous and Woody Plants: Paving the Way for Enhancing Phytoremediation Efficiency. Biotechnol. Adv. 2016, 34, 1131–1148. [Google Scholar] [CrossRef]
- Di Baccio, D.; Galla, G.; Bracci, T.; Andreucci, A.; Barcaccia, G.; Tognetti, R.; Sebastiani, L. Transcriptome Analyses of Populus × Euramericana Clone I-214 Leaves Exposed to Excess Zinc. Tree Physiol. 2011, 31, 1293–1308. [Google Scholar] [CrossRef]
- Ambroise, V.; Legay, S.; Guerriero, G.; Hausman, J.F.; Cuypers, A.; Sergeant, K. Selection of Appropriate Reference Genes for Gene Expression Analysis under Abiotic Stresses in Salix Viminalis. Int. J. Mol. Sci. 2019, 20, 4210. [Google Scholar] [CrossRef]
- Evlard, A.; Sergeant, K.; Ferrandis, S.; Printz, B.; Renaut, J.; Guignard, C.; Paul, R.; Hausman, J.F.; Campanella, B. Physiological and Proteomic Responses of Different Willow Clones (Salix Fragilis × Alba) Exposed to Dredged Sediment Contaminated by Heavy Metals. Int. J. Phytoremediation 2014, 16, 1148–1169. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Singh, A.; Roychoudhury, A. Involvement of Sulfur in the Regulation of Abiotic Stress Tolerance in Plants. In Protective chemical agents in the amelioration of plant abiotic stress: Biochemical and Molecular Perspectives; Wiley: Hoboken, NJ, USA, 2020; pp. 437–466. [Google Scholar]
- Howarth, J.R.; Domínguez-Solís, J.R.; Gutiérrez-Alcalá, G.; Wray, J.L.; Romero, L.C.; Gotor, C. The Serine Acetyltransferase Gene Family in Arabidopsis Thaliana and the Regulation of Its Expression by Cadmium. Plant Mol. Biol. 2003, 51, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Samiullah; Singh, S.; Nazar, R. Activities of Antioxidative Enzymes, Sulphur Assimilation, Photosynthetic Activity and Growth of Wheat (Triticum Aestivum) Cultivars Differing in Yield Potential under Cadmium Stress. J. Agron. Crop Sci. 2007, 193, 435–444. [Google Scholar] [CrossRef]
- Monteiro, C.; Conceicao, S.; Pinho, S.; Oliveira, H.; Pedrosa, T.; Dias, M.C. Cadmium-Induced Cyto- and Genotoxicity Are Organ-Dependent in Lettuce. Chem. Res. Toxicol. 2012, 25, 1423–1434. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, S.; Keunen, E.; Mertens, A.I.G.; Beemster, G.T.S.; Vangronsveld, J.; Cuypers, A. Cell Cycle Regulation in Different Leaves of Arabidopsis Thaliana Plants Grown under Control and Cadmium-Exposed Conditions. Environ. Exp. Bot. 2018, 155, 441–452. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Barceló, J. Water Relations in Heavy Metal Stressed Plants. In Heavy Metal Stress in Plants; Springer: Berlin, Heidelberg, 1999; pp. 207–229. [Google Scholar]
- Cortés-Eslava, J.; Gómez-Arroyo, S.; Risueño, M.C.; Testillano, P.S. The Effects of Organophosphorus Insecticides and Heavy Metals on DNA Damage and Programmed Cell Death in Two Plant Models. Environ. Pollut. 2018, 240, 77–86. [Google Scholar] [CrossRef]
- Smeets, K.; Opdenakker, K.; Remans, T.; Van Sanden, S.; Van Belleghem, F.; Semane, B.; Horemans, N.; Guisez, Y.; Vangronsveld, J.; Cuypers, A. Oxidative Stress-Related Responses at Transcriptional and Enzymatic Levels after Exposure to Cd or Cu in a Multipollution Context. Plant Physiol. 2009, 166, 1982–1992. [Google Scholar] [CrossRef]
- Tauqeer, H.M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Iftikhar, U.; Ahmad, R.; Farid, M.; Abbasi, G.H. Phytoremediation of Heavy Metals by Alternanthera Bettzickiana: Growth and Physiological Response. Ecotoxicol. Environ. Saf. 2016, 126, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.P.; Shan, C.J.; Zhao, H.; Li, J.C.; Jia, G.L.; Jiang, H.; Wu, S.Q.; Wang, Q. The Difference in Antioxidant Capacity of Four Alfalfa Cultivars in Response to Zn. Ecotoxicol. Environ. Saf. 2015, 114, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Zeneli, L.; Daci-Ajvazi, M.; Daci, N.M.; Hoxha, D.; Shala, A. Environmental Pollution and Relationship between Total Antioxidant Capacity and Heavy Metals (Pb, Cd, Zn, Mn, and Fe) in Solanum Tuberosum L. and Allium Cepa L. Hum. Ecol. Risk Assess. 2013, 19, 1618–1627. [Google Scholar] [CrossRef]
- Gajewska, E.; Bernat, P.; Dlugoński, J.; Sklodowska, M. Effect of Nickel on Membrane Integrity, Lipid Peroxidation and Fatty Acid Composition in Wheat Seedlings. J. Agron. Crop Sci. 2012, 198, 286–294. [Google Scholar] [CrossRef]
- Taulavuori, K.; Prasad, M.N.V.; Taulavuori, E.; Laine, K. Metal Stress Consequences on Frost Hardiness of Plants at Northern High Latitudes: A Review and Hypothesis. Environ. Pollut. 2005, 135, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Talanova, V.V.; Titov, A.F.; Repkina, N.S.; Topchieva, L.V. Cold-Responsive COR/LEA Genes Participate in the Response of Wheat Plants to Heavy Metals Stress. Dokl. Biol. Sci. 2013, 448, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Barah, P.; Suarez-Rodriguez, M.C.; Bressendorff, S.; Friis, P.; Costantino, P.; Bones, A.M.; Nielsen, H.B.; Mundy, J. Transcriptome Responses to Combinations of Stresses in Arabidopsis. Plant Physiol. 2013, 161, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Noctor, G.; De Paepe, R.; Foyer, C.H. Mitochondrial Redox Biology and Homeostasis in Plants. Trends Plant Sci. 2007, 12, 125–134. [Google Scholar] [CrossRef]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 1446. [Google Scholar] [CrossRef]
- Domon, J.M.; Baldwin, L.; Acket, S.; Caudeville, E.; Arnoult, S.; Zub, H.; Gillet, F.; Lejeune-Hénaut, I.; Brancourt-Hulmel, M.; Pelloux, J.; et al. Cell Wall Compositional Modifications of Miscanthus Ecotypes in Response to Cold Acclimation. Phytochemistry 2013, 85, 51–61. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 1–22. [Google Scholar] [CrossRef]
- Gutsch, A.; Hendrix, S.; Guerriero, G.; Renaut, J.; Lutts, S.; Alseekh, S.; Fernie, A.R.; Hausman, J.F.; Vangronsveld, J.; Cuypers, A.; et al. Long-Term Cd Exposure Alters the Metabolite Profile in Stem Tissue of Medicago Sativa. Cells 2020, 9, 1–22. [Google Scholar] [CrossRef]
- Bruegmann, T.; Wetzel, H.; Hettrich, K.; Smeds, A.; Willför, S.; Kersten, B.; Fladung, M. Knockdown of PCBER1, a Gene of Neolignan Biosynthesis, Resulted in Increased Poplar Growth. Planta 2019, 249, 515–525. [Google Scholar] [CrossRef]
- Paniagua, C.; Bilkova, A.; Jackson, P.; Dabravolski, S.; Riber, W.; Didi, V.; Houser, J.; Gigli-Bisceglia, N.; Wimmerova, M.; Budínská, E.; et al. Dirigent Proteins in Plants: Modulating Cell Wall Metabolism during Abiotic and Biotic Stress Exposure. J. Exp. Bot. 2017, 68, 3287–3301. [Google Scholar] [CrossRef]
- Cunningham, S.M.; Nadeau, P.; Castonguay, Y.; Laberge, S.; Volenec, J.J. Raffinose and Stachyose Accumulation, Galactinol Synthase Expression, and Winter Injury of Contrasting Alfalfa Germplasms. Crop Sci. 2003, 43, 562–570. [Google Scholar] [CrossRef]
- Pena, L.B.; Azpilicueta, C.E.; Benavides, M.P.; Gallego, S.M. Protein Oxidative Modifications. In Metal Toxicity in Plants: Perception, Signaling and Remediation; Springer: Berlin/Heidelberg, Germany, 2012; pp. 207–225. [Google Scholar]
- Jacob, P.; Hirt, H.; Bendahmane, A. The Heat-Shock Protein/Chaperone Network and Multiple Stress Resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef]
- Sasikumar, A.N.; Perez, W.B.; Kinzy, T.G. The Many Roles of the Eukaryotic Elongation Factor 1 Complex. Wiley Interdiscip. Rev. RNA 2012, 3, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Momčilović, I.; Prasad, P.V.V. Roles of Protein Synthesis Elongation Factor EF-Tu in Heat Tolerance in Plants. J. Bot. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- McKay, H.M. Frost Hardiness and Cold-Storage Tolerance of the Root System of Picea Sitchensis, Pseudotsuga Menziesii, Larix Kaempferi and Pinus Sylvestris Bare-Root Seedlings. Scand. J. For. Res. 1994, 9, 203–213. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of ‘“Antioxidant Power”’: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. Superoxide Dismutase, an Enzymic Function for Erythrocuprein (Hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Queval, G.; Noctor, G. A Plate Reader Method for the Measurement of NAD, NADP, Glutathione, and Ascorbate in Tissue Extracts: Application to Redox Profiling during Arabidopsis Rosette Development. Anal. Biochem. 2007, 363, 58–69. [Google Scholar] [CrossRef]
- Legay, S.; Guerriero, G.; Deleruelle, A.; Lateur, M.; Evers, D.; André, C.M.; Hausman, J.F. Apple Russeting as Seen through the RNA-Seq Lens: Strong Alterations in the Exocarp Cell Wall. Plant Mol. Biol. 2015, 88, 21–40. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-Hit: A Fast Program for Clustering and Comparing Large Sets of Protein or Nucleotide Sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A Comparative Platform for Green Plant Genomics. Nucleic Acids Res. 2012, 40, 1178–1186. [Google Scholar] [CrossRef]
- Wang, W.; Scali, M.; Vignani, R.; Spadafora, A.; Sensi, E.; Mazzuca, S.; Cresti, M. Protein Extraction for Two-Dimensional Electrophoresis from Olive Leaf, a Plant Tissue Containing High Levels of Interfering Compounds. Electrophoresis 2003, 24, 2369–2375. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Pennycooke, J.C.; Cox, S.; Stushnoff, C. Relationship of Cold Acclimation, Total Phenolic Content and Antioxidant Capacity with Chilling Tolerance in Petunia (Petunia × Hybrida). Environ. Exp. Bot. 2005, 53, 225–232. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (1) | ||||
| Factor | TAC | Hydro. | Lipo. | |
| Temperature | 0.175 | 0.366 | 0.157 | |
| Heavy metals | 0.031 * | 0.002 * | 0.112 | |
| Interaction | 0.84 | 0.668 | 0.914 | |
| (2) | ||||
| Factor | CAT | SOD | GR | |
| Temperature | 0.017 | 0.562 | 0.003 * | |
| Heavy metals | 0.004 * | 0.015 * | 0.446 | |
| Interaction | 0.09 | 0.345 | 0.004 * | |
| (3) | ||||
| Factor | GSH Tot. | GSH Red. | GSSG | Perc. GSH |
| Temperature | <0.001 * | 0.007 * | 0.0127 * | 0.746 |
| Heavy metals | 0.258 | 0.529 | 0.1877 | 0.132 |
| Interaction | 0.723 | 0.704 | 0.939 | 0.58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambroise, V.; Legay, S.; Jozefczak, M.; Leclercq, C.C.; Planchon, S.; Hausman, J.-F.; Renaut, J.; Cuypers, A.; Sergeant, K. Impact of Heavy Metals on Cold Acclimation of Salix viminalis Roots. Int. J. Mol. Sci. 2024, 25, 1545. https://doi.org/10.3390/ijms25031545
Ambroise V, Legay S, Jozefczak M, Leclercq CC, Planchon S, Hausman J-F, Renaut J, Cuypers A, Sergeant K. Impact of Heavy Metals on Cold Acclimation of Salix viminalis Roots. International Journal of Molecular Sciences. 2024; 25(3):1545. https://doi.org/10.3390/ijms25031545
Chicago/Turabian StyleAmbroise, Valentin, Sylvain Legay, Marijke Jozefczak, Céline C. Leclercq, Sebastien Planchon, Jean-Francois Hausman, Jenny Renaut, Ann Cuypers, and Kjell Sergeant. 2024. "Impact of Heavy Metals on Cold Acclimation of Salix viminalis Roots" International Journal of Molecular Sciences 25, no. 3: 1545. https://doi.org/10.3390/ijms25031545
APA StyleAmbroise, V., Legay, S., Jozefczak, M., Leclercq, C. C., Planchon, S., Hausman, J.-F., Renaut, J., Cuypers, A., & Sergeant, K. (2024). Impact of Heavy Metals on Cold Acclimation of Salix viminalis Roots. International Journal of Molecular Sciences, 25(3), 1545. https://doi.org/10.3390/ijms25031545