
Nucleus Accumbens Associated Protein 1 in Cancers—The Real Value

 , , ,
, , ,  and
and
Abstract
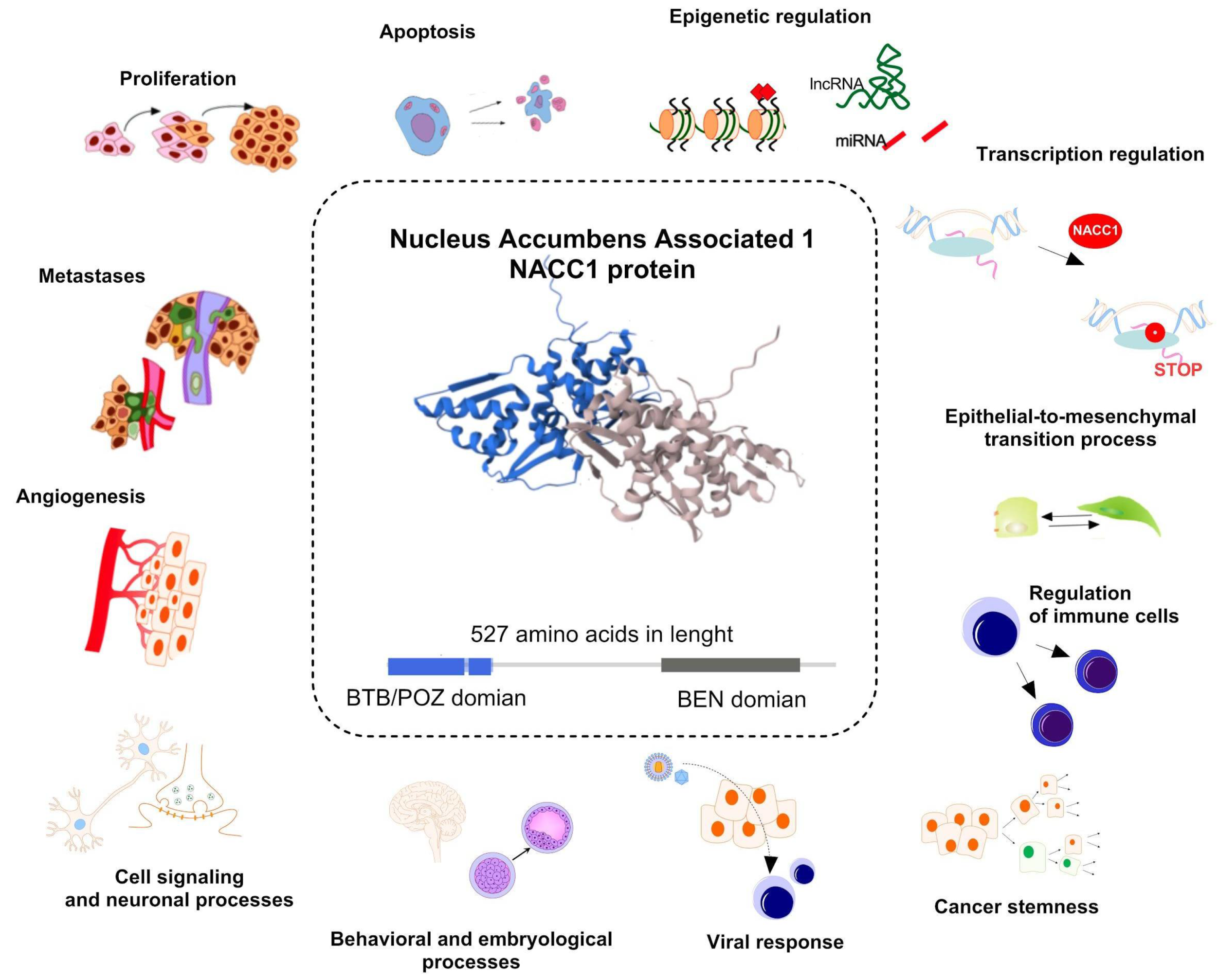
1. Introduction
2. Results
2.1. Expression Levels of NACC1 in Cancers Based on the Cancer Genome Atlas
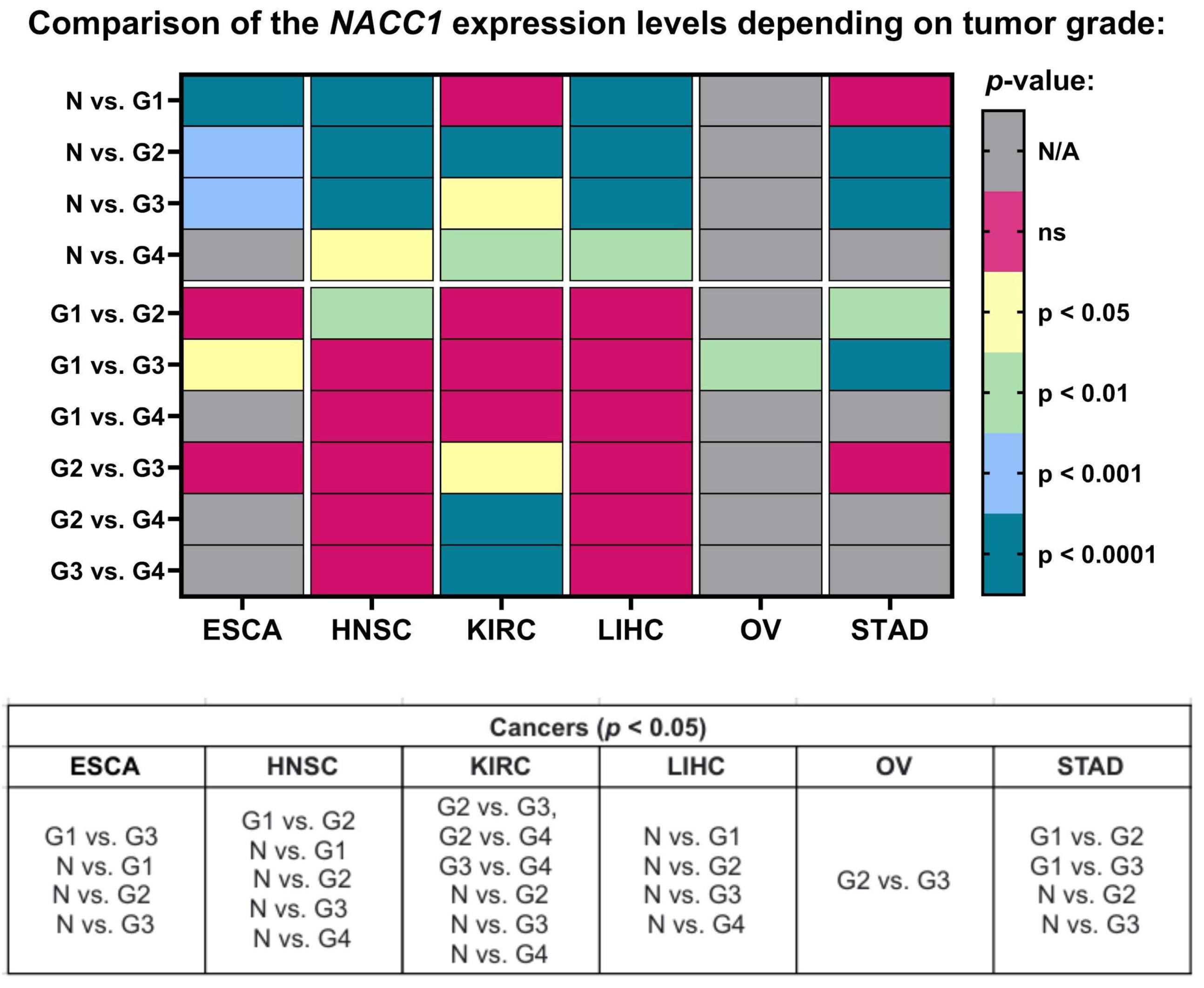
2.2. Tumor Grade
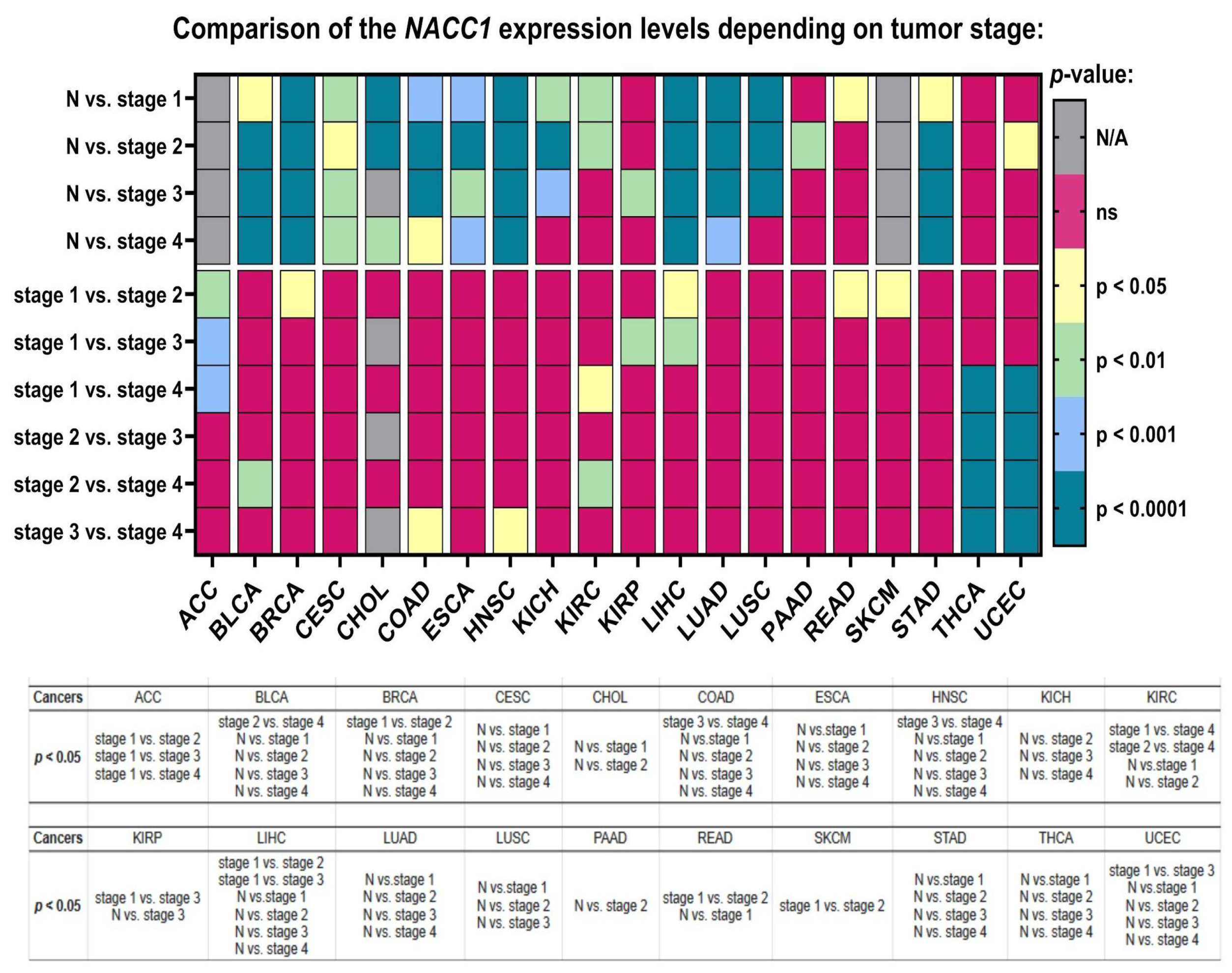
2.3. Cancer Stage
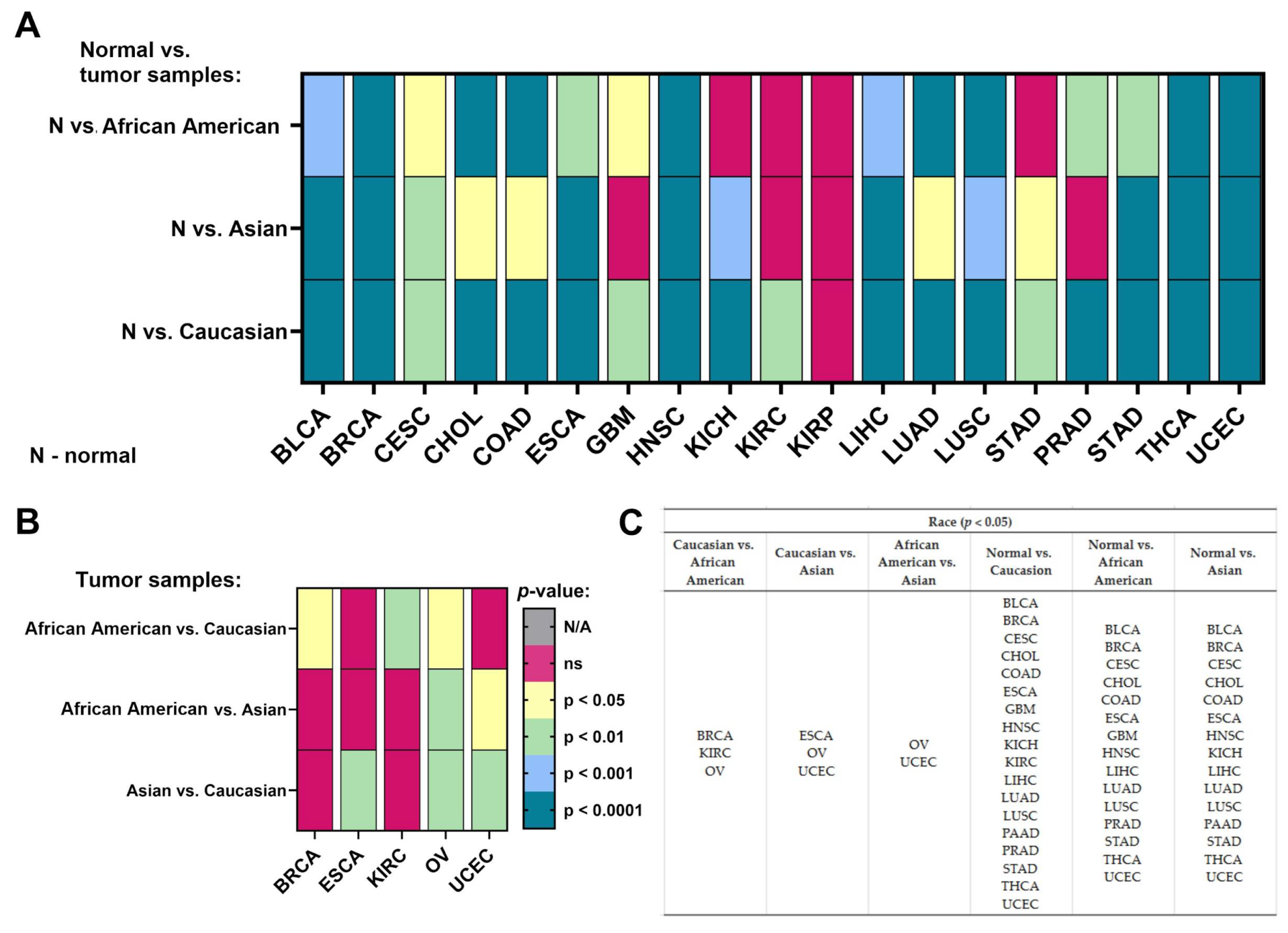
2.4. Race
2.5. Gender
2.6. Age
2.7. Weight
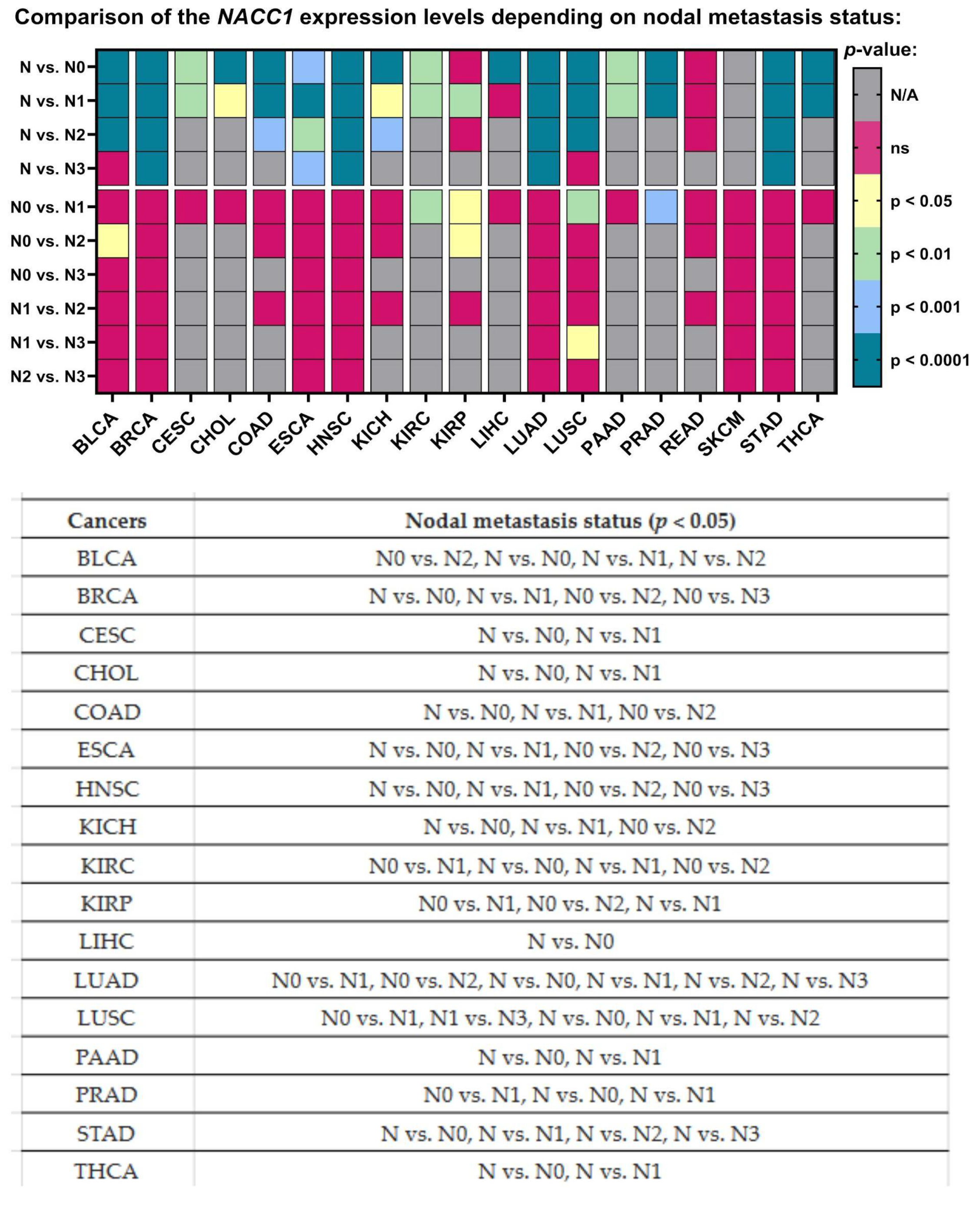
2.8. Nodal Metastasis
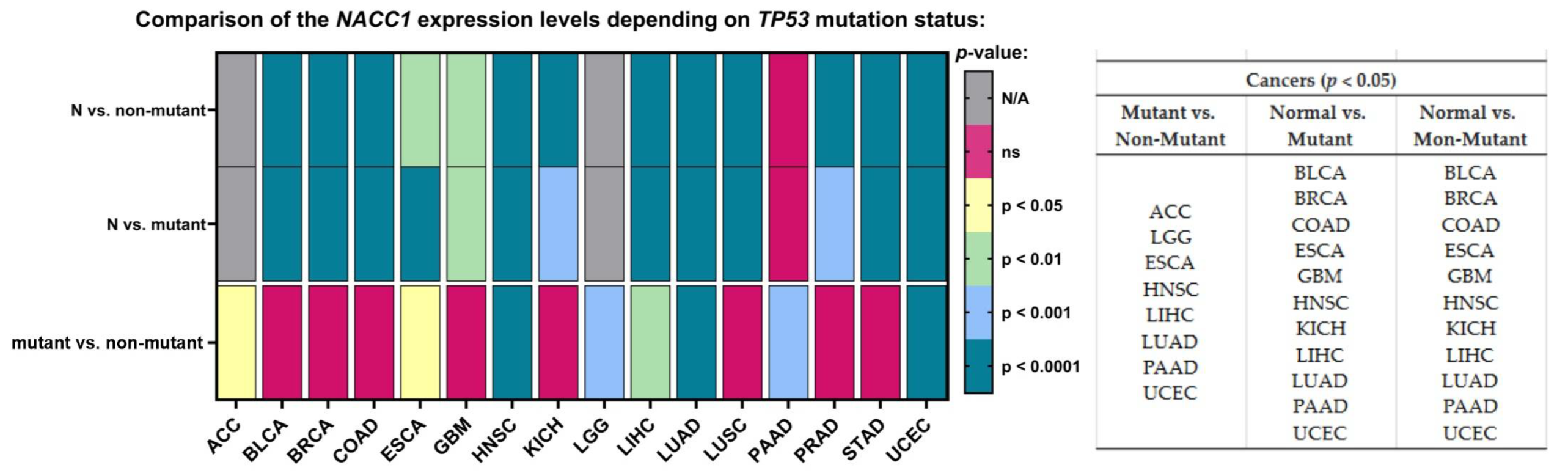
2.9. TP-53 Mutation
3. Discussion
4. Material and Methods
4.1. Databases
4.2. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef] [PubMed]
- Prager, G.W.; Braga, S.; Bystricky, B.; Qvortrup, C.; Criscitiello, C.; Esin, E.; Sonke, G.S.; Martínez, G.A.; Frenel, J.-S.; Karamouzis, M.; et al. Global cancer control: Responding to the growing burden, rising costs and inequalities in access. ESMO Open 2018, 3, e000285. [Google Scholar] [CrossRef] [PubMed]
- Dyzmann-Sroka, A. Assessment of educational services by Patient Target Group participating in the National Skin Cancer Prevention Programme (OPPNS) based on the example of the Wielkopolska region. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2024, 29, 294–299. [Google Scholar] [CrossRef]
- Bonet, M.; Recalde, E.; Soto, A.; Martinez, A.; Murcia, M.; Mases, J.; Fernandez, M.N.; Yufera, J.C.; Alvarez, A.; Eraso, M.A.; et al. Use of social media in radiation oncology: Multicenter data from the GOCO Group. Rep. Pr. Oncol. Radiother. 2024, 29, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Understanding Cancer Prognosis—National Cancer Institute; National Cancer Institute: Bethesda, MD, USA, 2019.
- Frick, C.; Rumgay, H.; Vignat, J.; Ginsburg, O.; Nolte, E.; Bray, F.; Soerjomataram, I. Isabelle Soerjomataram, Quantitative estimates of preventable and treatable deaths from 36 cancers worldwide: A population-based study. Lancet Glob. Health 2023, 11, e1700–e1712. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Tong, C.; Xiong, X. An overview of the co-transcription factor NACC1: Beyond its pro-tumor effects. Life Sci. 2024, 336, 122314. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ji, C.; Zhang, H.; Shan, Y.; Ren, Y.; Hu, Y.; Shi, L.; Guo, L.; Zhu, W.; Xia, Y.; et al. Identification of a small-molecule compound that inhibits homodimerization of oncogenic NAC1 protein and sensitizes cancer cells to anticancer agents. J. Biol. Chem. 2019, 294, 10006–10017. [Google Scholar] [CrossRef] [PubMed]
- Chaharbakhshi, E.; Jemc, J.C. Broad-complex, tramtrack, and bric-à-brac (BTB) proteins: Critical regulators of development. Genesis 2016, 54, 505–518. [Google Scholar] [CrossRef]
- Nakayama, N.; Kato, H.; Sakashita, G.; Nariai, Y.; Nakayama, K.; Kyo, S.; Urano, T. Protein complex formation and intranuclear dynamics of NAC1 in cancer cells. Arch. Biochem. Biophys. 2016, 606, 10–15. [Google Scholar] [CrossRef]
- Ruan, Y.; He, J.; Wu, W.; He, P.; Tian, Y.; Xiao, L.; Liu, G.; Wang, J.; Cheng, Y.; Zhang, S.; et al. Nac1 promotes self-renewal of embryonic stem cells through direct transcriptional regulation of c-Myc. Oncotarget 2017, 8, 47607–47618. [Google Scholar] [CrossRef]
- Nakayama, N.; Sakashita, G.; Nagata, T.; Kobayashi, N.; Yoshida, H.; Park, S.-Y.; Nariai, Y.; Kato, H.; Obayashi, E.; Nakayama, K.; et al. Nucleus Accumbens-Associated Protein 1 Binds DNA Directly through the BEN Domain in a Sequence-Specific Manner. Biomedicines 2020, 8, 608. [Google Scholar] [CrossRef]
- Available online: https://www.proteinatlas.org/ENSG00000160877-NACC1/cancer (accessed on 10 December 2024).
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M. Global Cancer Observatory: Cancer Today; International Agency for Research on Cancer: Lyon, France, 2021; Available online: https://gco.iarc.fr/today (accessed on 29 September 2021).
- Ngule, C.; Shi, R.; Ren, X.; Jia, H.; Oyelami, F.; Li, D.; Park, Y.; Kim, J.; Hemati, H.; Zhang, Y.; et al. NAC1 Promotes Stemness and Regulates Myeloid-derived Cell Status in Triple-Negative Breast Cancer. Mol. Cancer 2024, 23, 188. [Google Scholar] [CrossRef]
- Ngule, C.M.; Hemati, H.; Ren, X.; Obaleye, O.; Akinyemi, A.O.; Oyelami, F.F.; Xiong, X.; Song, J.; Liu, X.; Yang, J.-M. Identification of a NACC1-Regulated Gene Signature Implicated in the Features of Triple-Negative Breast Cancer. Biomedicines 2023, 11, 1223. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Ruan, S.; Jin, L.; Chen, Z.; Han, H.; Zhang, Y.; Jian, Z.; Lin, Y.; Shi, N.; Jin, H. CDK1, CCNB1, and CCNB2 are Prognostic Biomarkers and Correlated with Immune Infiltration in Hepatocellular Carcinoma. Med Sci. Monit. 2020, 26, e925289-1–e925289-14. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Sun, T.; Liu, R. NACC-1 regulates hepatocellular carcinoma cell malignancy and is targeted by miR-760. Acta Biochim. Biophys. Sin. 2020, 52, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Argani, P.; Palsgrove, D.N.; Anders, R.A.; Smith, S.C.; Saoud, C.; Kwon, R.; Voltaggio, L.; Assarzadegan, N.; Oshima, K.; Rooper, L.; et al. A Novel NIPBL-NACC1 Gene Fusion Is Characteristic of the Cholangioblastic Variant of Intrahepatic Cholangiocarcinoma. Am. J. Surg. Pathol. 2021, 45, 1550–1560. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Fujii, T.; Itami, H.; Uchiyama, T.; Nakai, T.; Hatakeyama, K.; Sugimoto, A.; Miyake, M.; Nakai, Y.; Tanaka, N.; et al. NACC1, as a Target of MicroRNA-331-3p, Regulates Cell Proliferation in Urothelial Carcinoma Cells. Cancers 2018, 10, 347. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.; Berner, A.; Trope, C.G.; Wang, T.-L.; Shih, I.-M. Expression and clinical role of the bric-a-brac tramtrack broad complex/poxvirus and zinc protein NAC-1 in ovarian carcinoma effusions. Hum. Pathol. 2007, 38, 1030–1036. [Google Scholar] [CrossRef]
- Gao, M.; Wu, R.-C.; Herlinger, A.L.; Yap, K.; Kim, J.-W.; Wang, T.-L.; Shih, I.-M. Identification of the NAC1-Regulated Genes in Ovarian Cancer. Am. J. Pathol. 2013, 184, 133–140. [Google Scholar] [CrossRef]
- Dong, S.; Wang, X.; Yang, S.; Guo, F.; Zhang, J.; Ji, C.; Shi, L.; Cheng, Y.; Hu, Y.; Li, Z.; et al. Mechanistic insights of NAC1 nuclear export and its role in ovarian cancer resistance to docetaxel. Biochem. Pharmacol. 2023, 211, 115533. [Google Scholar] [CrossRef]
- Rahman, M.T.; Nakayama, K.; Katagiri, H.; Katagiri, A.; Ishibashi, T.; Ishikawa, M.; Iida, K.; Nakayama, N.; Otsuki, Y.; Nakayama, S.; et al. Fatty acid synthase expression associated with NAC1 is a potential therapeutic target in ovarian clear cell carcinomas. Br. J. Cancer 2012, 107, 300–307. [Google Scholar] [CrossRef]
- Kolenda, T.; Poter, P.; Guglas, K.; Kozłowska-Masłoń, J.; Braska, A.; Kazimierczak, U.; Teresiak, A. Biological role and diagnostic utility of ribosomal protein L23a pseudogene 53 in cutaneous melanoma. Rep. Pr. Oncol. Radiother. 2023, 28, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, T.; Śmiełowska, M.; Lipowicz, J.; Ostapowicz, J.; Pacześna, P.; Rosochowicz, M.; Poter, P.; Kozłowska-Masłoń, J.; Guglas, K.; Dudek, K.; et al. The RNA world: From experimental laboratory to “in silico” approach. Part 1: User friendly RNA expression databases portals. Rep. Pract. Oncol. Radiother. 2024, 29, 245–257. [Google Scholar] [CrossRef]
- Daniel, J.A.; Elizarova, S.; Shaib, A.H.; Chouaib, A.A.; Magnussen, H.M.; Wang, J.; Brose, N.; Rhee, J.; Tirard, M. An intellectual-disability-associated mutation of the transcriptional regulator NACC1 impairs glutamatergic neurotransmission. Front. Mol. Neurosci. 2023, 16, 1115880. [Google Scholar] [CrossRef] [PubMed]
- Deehan, M.A.; Kothuis, J.M.; Sapp, E.; Chase, K.; Ke, Y.; Seeley, C.; Iuliano, M.; Kim, E.; Kennington, L.; Miller, R.; et al. Nacc1 Mutation in Mice Models Rare Neurodevelopmental Disorder with Underlying Synaptic Dysfunction. J. Neurosci. 2024, 44. [Google Scholar] [CrossRef]
- Ren, Y.; Kumar, A.; Das, J.; Peng, H.-J.; Wang, L.; Balllard, D.; Xiong, X.; Ren, X.; Zhang, Y.; Yang, J.-M.; et al. Tumorous expression of NAC1 restrains antitumor immunity through the LDHA-mediated immune evasion. J. Immunother. Cancer 2022, 10, e004856. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Peng, H.Y.; Kishore Das, J.; Kumar, A.; Ren, Y.; Ballard, D.J.; Xiong, X.; Yang, W.; Ren, X.; de Figueiredo, P.; et al. NAC1 confines virus-specific memory formation of CD4+ T cells through the ROCK1-mediated pathway. J. Med. Virol. 2023, 95, e28957. [Google Scholar] [CrossRef] [PubMed]
- Maćkowiak, B.; Ostrowska, K.; Kulcenty, K.; Kaźmierska, J.; Ostapowicz, J.; Nowicka, H.; Szewczyk, M.; Książek, K.; Suchorska, W.M.; Golusiński, W. The impact of XPC gene single nucleotide polymorphism rs2228001 on head and neck cancer patients’ response to radiotherapy treatment. Rep. Pr. Oncol. Radiother. 2024, 29, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-X.; Tan, X.-R.; Wei, W.; Li, M.-Q.; Zhang, W.-N.; Gong, Z.-C.; Zhang, Y.; Zhao, H.-R. A radiobiological perspective on radioresistance or/and radiosensitivity of head and neck squamous cell carcinoma. Rep. Pr. Oncol. Radiother. 2024, 28, 809–822. [Google Scholar] [CrossRef]
- Kozłowska-Masłoń, J.; Guglas, K.; Kolenda, T.; Lamperska, K.; Makałowska, I. miRNA in head and neck squamous cell carcinomas: Promising but still distant future of personalized oncology. Rep. Pr. Oncol. Radiother. 2023, 28, 681–697. [Google Scholar] [CrossRef]
- Regulska, K.; Kolenda, T.; Michalak, M.; Stanisz, B.J. Impact of ramipril nitroso-metabolites on cancer incidence — in silico and in vitro safety evaluation. Rep. Pr. Oncol. Radiother. 2023, 28, 612–622. [Google Scholar] [CrossRef]
- Nanda, R.; Katke, A.; B, T.; Giri, G.V.; Pawar, Y.; V, M.M.; Babu, G. Ten years survival results of randomized study comparing weekly vs. triweekly cisplatin with concurrent radiation in locally advanced carcinoma cervix. Rep. Pr. Oncol. Radiother. 2023, 28, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Chen, H.; Mei, X.; Li, X. Silencing of NACC1 inhibits the proliferation, migration and invasion of nasopharyngeal carcinoma cells via regulating the AKT/mTOR signaling pathway. Oncol. Lett. 2021, 22, 828. [Google Scholar] [CrossRef]
- Yang, J.-M.; Ren, Y.; Kumar, A.; Xiong, X.; Das, J.K.; Peng, H.-Y.; Wang, L.; Ren, X.; Zhang, Y.; Ji, C.; et al. NAC1 Modulates Autoimmunity by Suppressing Regulatory T Cell-Mediated Tolerance. Sci. Adv. 2022, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Kumar, A.; Das, J.K.; Ren, Y.; Peng, H.-Y.; Ballard, D.J.; Xiong, X.; Davis, J.R.; Ren, X.; Yang, J.-M.; et al. Expression of NAC1 Restrains the Memory Formation of CD8+ T Cells during Viral Infection. Viruses 2022, 14, 1713. [Google Scholar] [CrossRef]
- Tsunoda, K.; Oikawa, H.; Tada, H.; Tatemichi, Y.; Muraoka, S.; Miura, S.; Shibazaki, M.; Maeda, F.; Takahashi, K.; Akasaka, T.; et al. Nucleus Accumbens-Associated 1 Contributes to Cortactin Deacetylation and Augments the Migration of Melanoma Cells. J. Investig. Dermatol. 2011, 131, 1710–1719. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Ren, X.; Ngule, C.; Xiong, X.; Song, J.; Li, Z.; Yang, J.-M. Co-Targeting Nucleus Accumbens Associate 1 and NF-κB Signaling Synergistically Inhibits Melanoma Growth. Biomedicines 2023, 11, 2221. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overexpression of NACC1 in Cancers |
|---|
| bladder urothelial carcinoma breast invasive carcinoma cervical squamous cell carcinoma cholangiocarcinoma colon adenocarcinoma esophageal carcinoma glioblastoma multiforme head and neck squamous cell carcinoma kidney chromophobe kidney renal clear cell carcinoma liver hepatocellular carcinoma lung adenocarcinoma lung squamous cell carcinoma pancreatic adenocarcinoma prostate adenocarcinoma stomach adenocarcinoma thyroid carcinoma uterine corpus endometrial carcinoma |
| No. | TCGA Dataset | Number of Samples | Female | Men |
|---|---|---|---|---|
| 1. | Adenocritical carcinoma | 79 | 48 | 31 |
| 2. | Bladder urothelial carcinoma | 402 | 297 | 105 |
| 3. | Brain lower-grade glioma | 514 | 229 | 285 |
| 4. | Breast invasive carcinoma | 1087 | 1075 | 12 |
| 5. | Cervical squamous cell carcinoma | 305 | 305 | 0 |
| 6. | Cholangiocarcinoma | 36 | 20 | 16 |
| 7. | Colon adenocarcinoma | 283 | 127 | 156 |
| 8. | Esophageal carcinoma | 183 | 26 | 157 |
| 9. | Glioblastoma multiforme | 155 | 54 | 101 |
| 10. | Head and neck squamous cell carcinoma | 519 | 136 | 383 |
| 11. | Kidney chromophobe | 65 | 27 | 38 |
| 12. | Kidney renal clear cell carcinoma | 533 | 188 | 345 |
| 13. | Kidney renal papillary cell carcinoma | 290 | 76 | 214 |
| 14. | Liver hepatocellular carcinoma | 362 | 117 | 245 |
| 15. | Lung adenocarcinoma | 514 | 276 | 238 |
| 16. | Lung squamous cell carcinoma | 494 | 128 | 366 |
| 17. | Mesothelioma | 87 | 16 | 71 |
| 18. | Ovarian serous cystadenocarcinoma | 302 | 302 | 0 |
| 19. | Pancreatic adenocarcinoma | 177 | 80 | 97 |
| 20. | Pheochromocytoma and paraganglioma | 179 | 101 | 78 |
| 21. | Prostate adenocarcinoma | 497 | 0 | 497 |
| 22. | Rectum adenocarcinoma | 165 | 75 | 90 |
| 23. | Sarcoma | 259 | 118 | 141 |
| 24. | Skin cutaneous melanoma | 461 | 175 | 286 |
| 25. | Stomach adenocarcinoma | 415 | 147 | 268 |
| 26. | Testicular germ cell tumors | 128 | 0 | 128 |
| 27. | Thymoma | 120 | 57 | 63 |
| 28. | Thyroid carcinoma | 505 | 369 | 136 |
| 29. | Uterine corpus endometrial carcinoma | 546 | 546 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janiczek-Polewska, M.; Kolenda, T.; Poter, P.; Jagiełło, I.; Kozłowska-Masłoń, J.; Regulska, K.; Malicki, J.; Marszałek, A. Nucleus Accumbens Associated Protein 1 in Cancers—The Real Value. Int. J. Mol. Sci. 2024, 25, 13632. https://doi.org/10.3390/ijms252413632
Janiczek-Polewska M, Kolenda T, Poter P, Jagiełło I, Kozłowska-Masłoń J, Regulska K, Malicki J, Marszałek A. Nucleus Accumbens Associated Protein 1 in Cancers—The Real Value. International Journal of Molecular Sciences. 2024; 25(24):13632. https://doi.org/10.3390/ijms252413632
Chicago/Turabian StyleJaniczek-Polewska, Marlena, Tomasz Kolenda, Paulina Poter, Inga Jagiełło, Joanna Kozłowska-Masłoń, Katarzyna Regulska, Julian Malicki, and Andrzej Marszałek. 2024. "Nucleus Accumbens Associated Protein 1 in Cancers—The Real Value" International Journal of Molecular Sciences 25, no. 24: 13632. https://doi.org/10.3390/ijms252413632
APA StyleJaniczek-Polewska, M., Kolenda, T., Poter, P., Jagiełło, I., Kozłowska-Masłoń, J., Regulska, K., Malicki, J., & Marszałek, A. (2024). Nucleus Accumbens Associated Protein 1 in Cancers—The Real Value. International Journal of Molecular Sciences, 25(24), 13632. https://doi.org/10.3390/ijms252413632