
Electron-Transferring Flavoprotein and Its Dehydrogenase Required for Fungal Pathogenicity in Arthrobotrys oligospora



Abstract
1. Introduction
2. Results
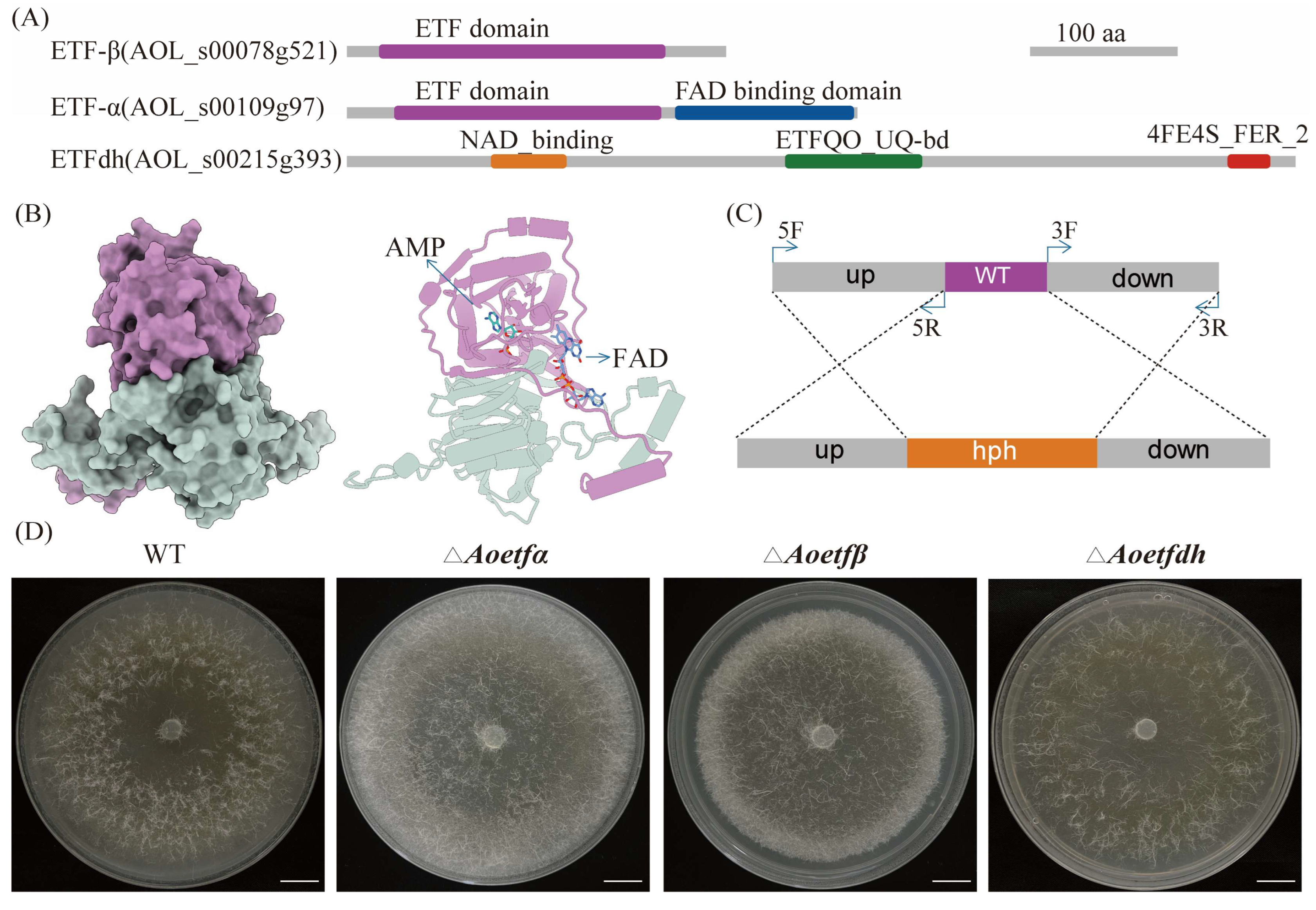
2.1. Analysis and Deletion of etf and etfdh in A. oligospora
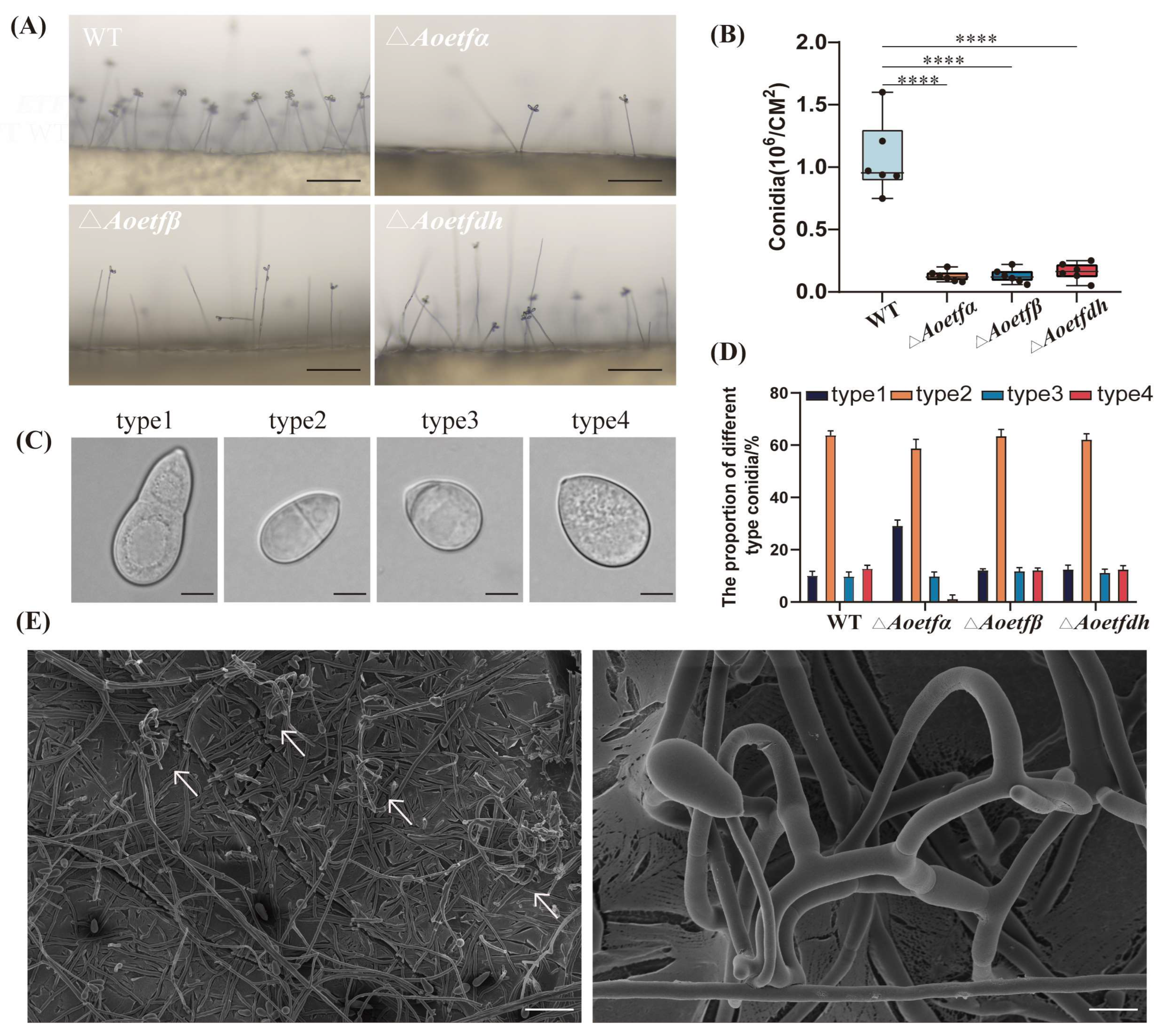
2.2. Aoetf and Aoetfdh Impair Conidiation in A. oligospora
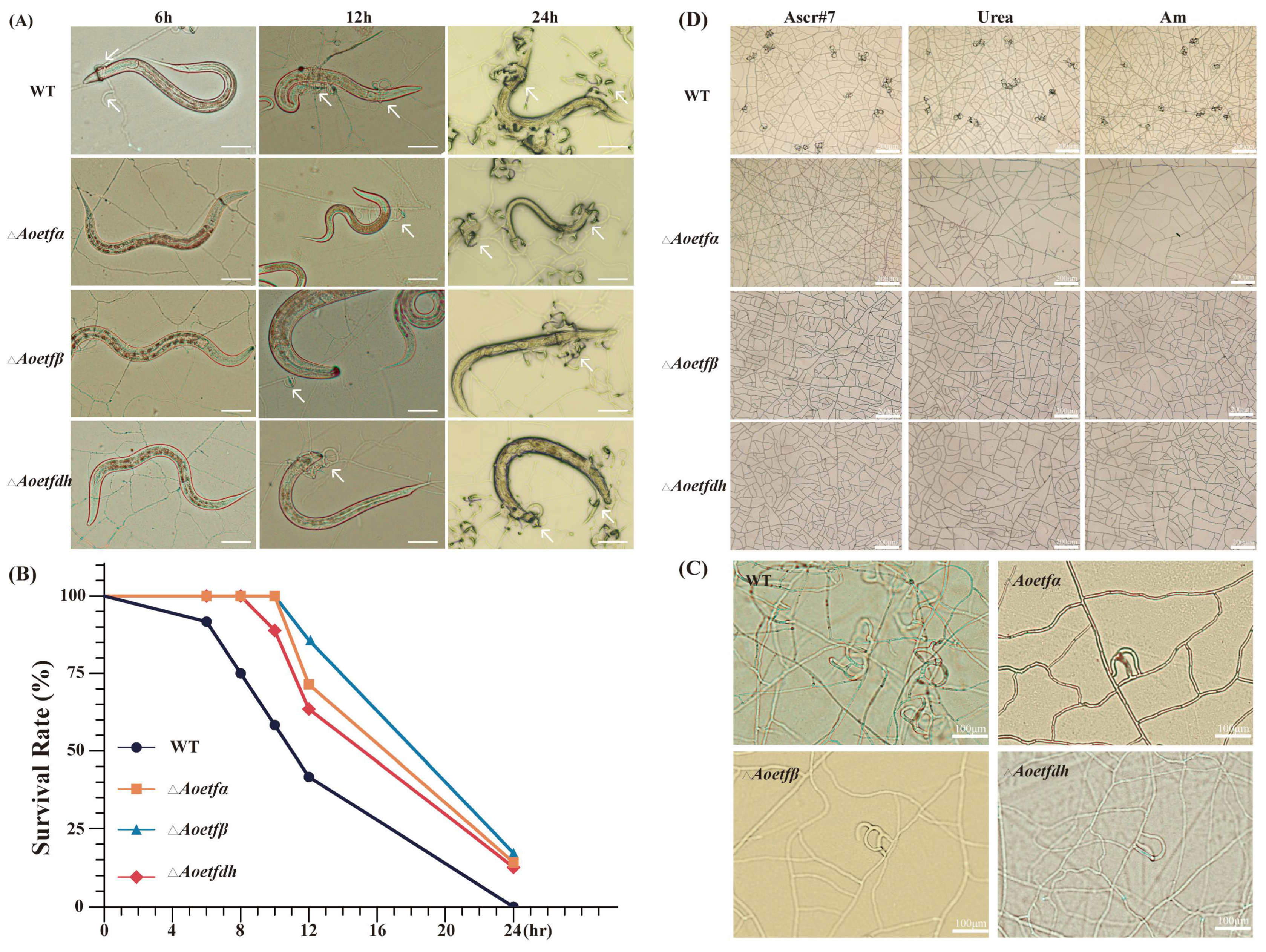
2.3. Aoetf and Aoetfdh Are Essential for the Formation of Traps in A. oligospora
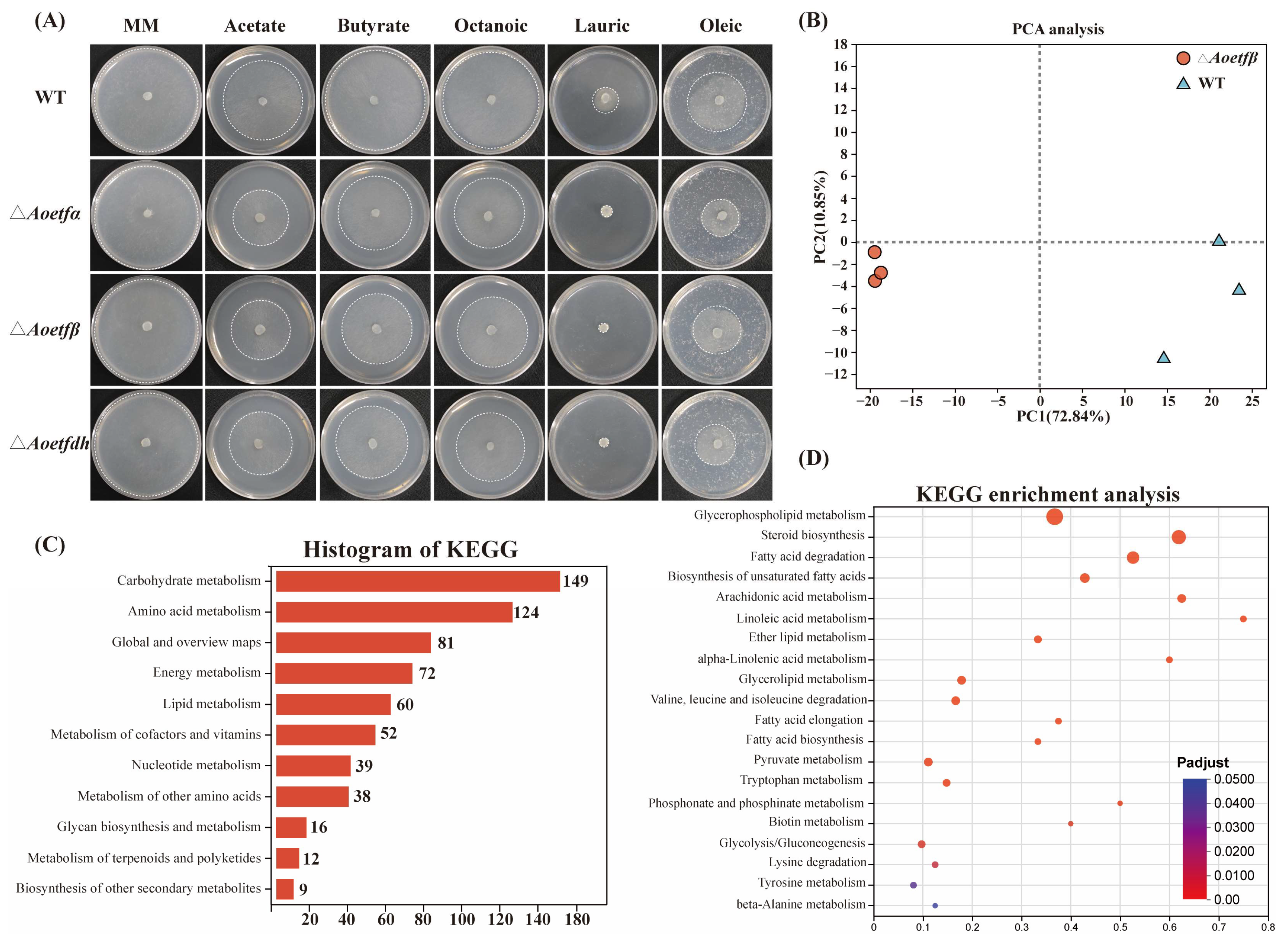
2.4. The Absence of Aoetf Affects Fatty Acid Metabolism in A. oligospora
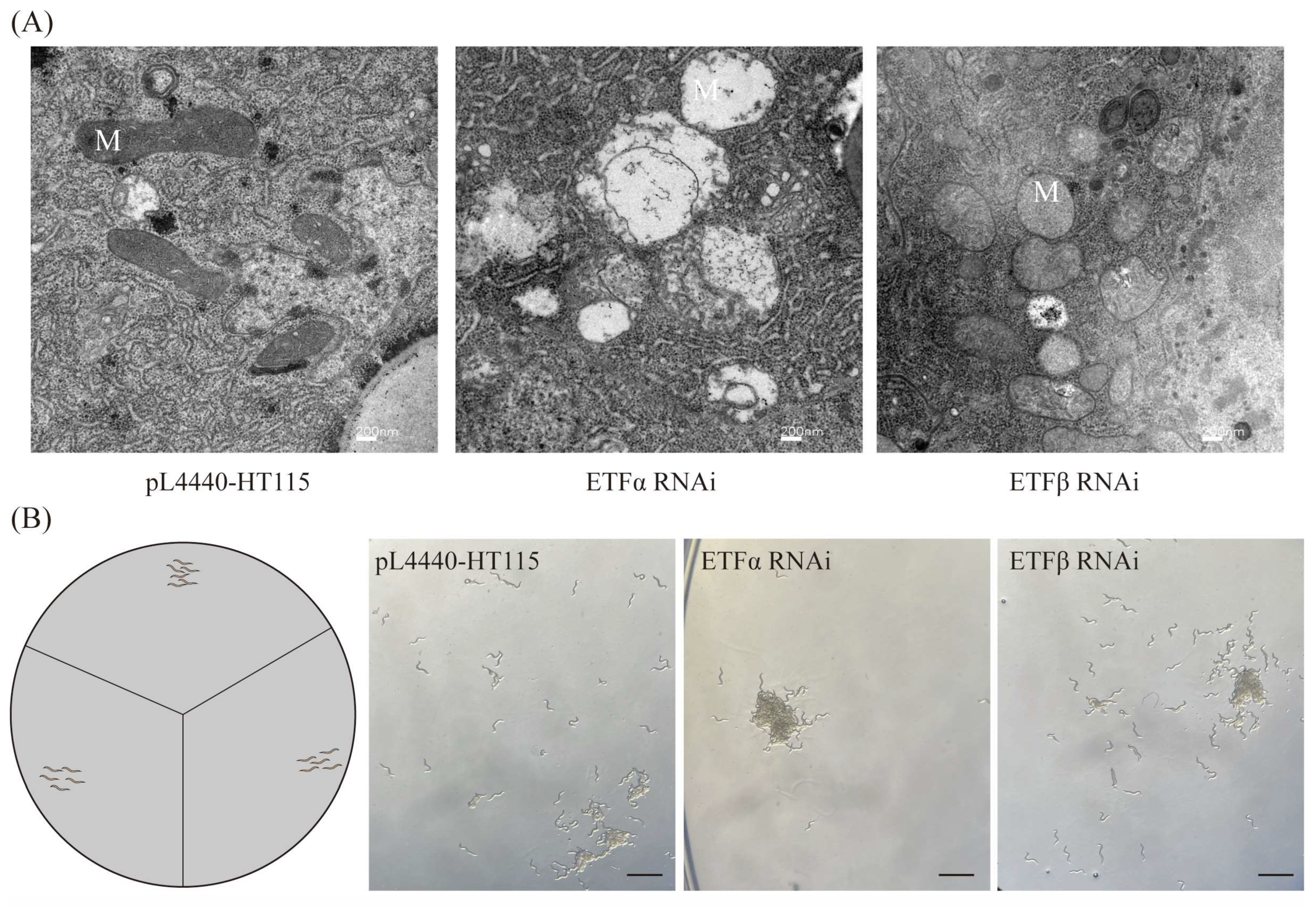
2.5. Aoetf Deletion Impairs Mitochondrial Function in A. oligospra
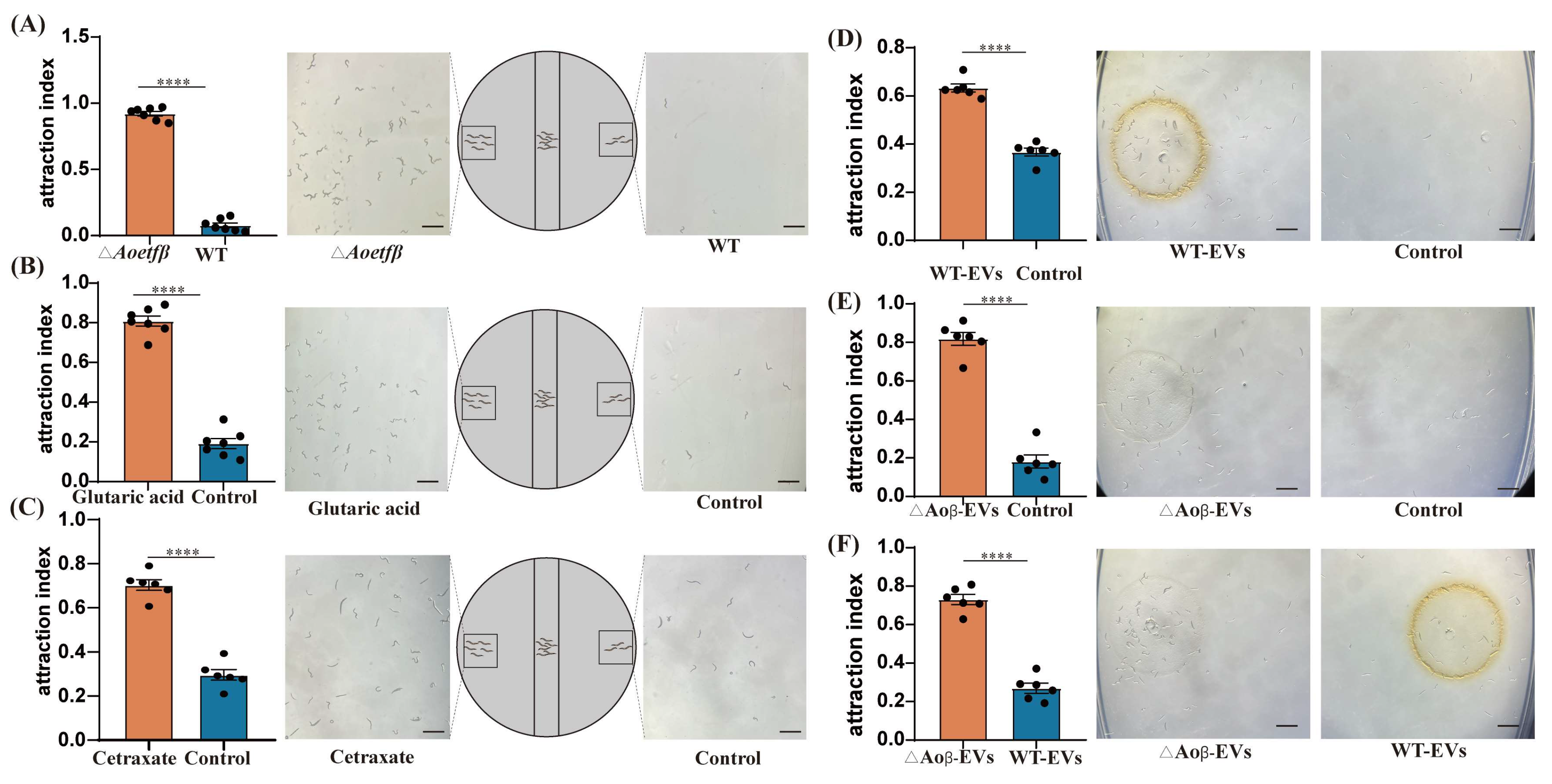
2.6. The Aoetf Mutant Strain Has the Ability to Attract C. elegans
2.7. Nematode ETF Exhibited Similar Roles in Mitochondrial Function to A. oligospora
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, and Culture Conditions
4.2. Targeted Gene Deletion
4.3. Phenotypic Characterization of Strains
4.4. Microscopic Observation
4.5. Difference in Trap Formation
4.6. Chemotaxis Assay
4.7. Exosome Assay
4.8. Transcriptome and Metabolome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Henriques, B.J.; Katrine Jentoft Olsen, R.; Gomes, C.M.; Bross, P. Electron transfer flavoprotein and its role in mitochondrial energy metabolism in health and disease. Gene 2021, 776, 145407. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wan, M.; Fan, Y. Electron-transferring flavoprotein and its dehydrogenase contributed to growth development and virulence in Beauveria bassiana. J. Invertebr. Pathol. 2024, 205, 108141. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, J.; Hu, J.; Meng, X.; Zhang, Q.; Zhu, K.; Chen, X.; Chen, X.; Li, G.; Wang, Z.; et al. Functional characterization of electron-transferring flavoprotein and its dehydrogenase required for fungal development and plant infection by the rice blast fungus. Sci. Rep. 2016, 6, 24911. [Google Scholar] [CrossRef]
- Toplak, M.; Brunner, J.; Tabib, C.R.; Macheroux, P. Closing the gap: Yeast electron-transferring flavoprotein links the oxidation of d-lactate and d-α-hydroxyglutarate to energy production via the respiratory chain. FEBS J. 2019, 286, 3611–3628. [Google Scholar] [CrossRef] [PubMed]
- Wakitani, S.; Torisu, S.; Yoshino, T.; Hattanda, K.; Yamato, O.; Tasaki, R.; Fujita, H.; Nishino, K. Multiple Acyl-CoA Dehydrogenation Deficiency (Glutaric Aciduria Type II) with a Novel Mutation of Electron Transfer Flavoprotein-Dehydrogenase in a Cat. JIMD Rep. 2014, 13, 43–51. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, M.S.; Zulfiqar, F.; Alam, M.M.; Fujita, M. Regulation of ROS Metabolism in Plants under Environmental Stress: A Review of Recent Experimental Evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef]
- Su, H.; Zhao, Y.; Zhou, J.; Feng, H.; Jiang, D.; Zhang, K.Q.; Yang, J. Trapping devices of nematode-trapping fungi: Formation, evolution, and genomic perspectives. Biol. Rev. Camb. Philos. Soc. 2017, 92, 357–368. [Google Scholar] [CrossRef]
- Soliman, M.S.; El-Deriny, M.M.; Ibrahim, D.S.S.; Zakaria, H.; Ahmed, Y. Suppression of root-knot nematode Meloidogyne incognita on tomato plants using the nematode trapping fungus Arthrobotrys oligospora Fresenius. J. Appl. Microbiol. 2021, 131, 2402–2415. [Google Scholar] [CrossRef]
- Pires, D.; Vicente, C.S.L.; Menéndez, E.; Faria, J.M.S.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The Fight against Plant-Parasitic Nematodes: Current Status of Bacterial and Fungal Biocontrol Agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef]
- Wang, X.; Li, G.H.; Zou, C.G.; Ji, X.L.; Liu, T.; Zhao, P.J.; Liang, L.M.; Xu, J.P.; An, Z.Q.; Zheng, X.; et al. Bacteria can mobilize nematode-trapping fungi to kill nematodes. Nat. Commun. 2014, 5, 5776. [Google Scholar] [CrossRef]
- Zhang, F.; Yang, Y.Q.; Zhou, F.P.; Xiao, W.; Boonmee, S.; Yang, X.Y. Morphological and Phylogenetic Characterization of Five Novel Nematode-Trapping Fungi (Orbiliomycetes) from Yunnan, China. J. Fungi 2023, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhu, X.; Sun, F.; Zhang, K.; Niu, S.; Huang, X. The roles of actin cytoskeleton and actin-associated protein Crn1p in trap formation of Arthrobotrys oligospora. Res. Microbiol. 2017, 168, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.L.; Chen, Y.H.; He, J.N.; Xue, H.X.; Yan, N.; Zeng, Z.J.; Bennett, J.W.; Zhang, K.Q.; Niu, X.M. Integrated Metabolomics and Morphogenesis Reveal Volatile Signaling of the Nematode-Trapping Fungus Arthrobotrys oligospora. Appl. Environ. Microbiol. 2018, 84, e02749. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Hu, X.; Pop, M.; Wernet, N.; Kirschhöfer, F.; Brenner-Weiß, G.; Keller, J.; Bunzel, M.; Fischer, R. Fatal attraction of Caenorhabditis elegans to predatory fungi through 6-methyl-salicylic acid. Nat. Commun. 2021, 12, 5462. [Google Scholar] [CrossRef] [PubMed]
- Youssar, L.; Wernet, V.; Hensel, N.; Yu, X.; Hildebrand, H.G.; Schreckenberger, B.; Kriegler, M.; Hetzer, B.; Frankino, P.; Dillin, A.; et al. Intercellular communication is required for trap formation in the nematode-trapping fungus Duddingtonia flagrans. PLoS Genet. 2019, 15, e1008029. [Google Scholar] [CrossRef]
- Jiang, X.; Xiang, M.; Liu, X. Nematode-Trapping Fungi. Microbiol. Spectr. 2017, 5, 963–974. [Google Scholar] [CrossRef]
- Heintz, C.E.; Pramer, D. Ultrastructure of nematode-trapping fungi. J. Bacteriol. 1972, 110, 1163–1170. [Google Scholar] [CrossRef]
- Veenhuis, M.; Harder, W.; Nordbring-Hertz, B. Occurrence and metabolic significance of microbodies in trophic hyphae of the nematophagous fungus Arthrobotrys oligospora. Antonie Leeuwenhoek. 1989, 56, 241–249. [Google Scholar] [CrossRef]
- Talbot, N.J.; Foster, A.J. Genetics and genomics of the rice blast fungus Magnaporthe grisea: Developing an experimental model for understanding fungal diseases of cereals. Adv. Bot. Res. 2001, 34, 263–287. [Google Scholar]
- Watmough, N.J.; Frerman, F.E. The electron transfer flavoprotein: Ubiquinone oxidoreductases. Biochim. Biophys. Acta 2010, 1797, 1910–1916. [Google Scholar] [CrossRef]
- Wang, Y.; Palmfeldt, J.; Gregersen, N.; Makhov, A.M.; Conway, J.F.; Wang, M.; McCalley, S.P.; Basu, S.; Alharbi, H.; St Croix, C.; et al. Mitochondrial fatty acid oxidation and the electron transport chain comprise a multifunctional mitochondrial protein complex. J. Biol. Chem. 2019, 294, 12380–12391. [Google Scholar] [CrossRef] [PubMed]
- Chokchaiwong, S.; Kuo, Y.T.; Hsu, S.P.; Hsu, Y.C.; Lin, S.H.; Zhong, W.B.; Lin, Y.F.; Kao, S.H. ETF-QO Mutants Uncoupled Fatty Acid β-Oxidation and Mitochondrial Bioenergetics Leading to Lipid Pathology. Cells 2019, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Salerno, K.M.; Domenico, J.; Le, N.Q.; Stiles, C.D.; Solov’yov, I.A.; Martino, C.F. Long-Time Oxygen Localization in Electron Transfer Flavoprotein. J. Chem. Inf. Model 2022, 62, 4191–4199. [Google Scholar] [CrossRef] [PubMed]
- Herrero Martín, J.C.; Salegi Ansa, B.; Álvarez-Rivera, G.; Domínguez-Zorita, S.; Rodríguez-Pombo, P.; Pérez, B.; Calvo, E.; Paradela, A.; Miguez, D.G.; Cifuentes, A.; et al. An ETFDH-driven metabolon supports OXPHOS efficiency in skeletal muscle by regulating coenzyme Q homeostasis. Nat. Metab. 2024, 6, 209–225. [Google Scholar] [CrossRef]
- Liu, Q.; Bai, N.; Duan, S.; Shen, Y.; Zhu, L.; Yang, J. Characterizing the Role of AosfgA and AofluG in Mycelial and Conidial Development in Arthrobotrys oligospora and Their Role in Secondary Metabolism. Microorganisms 2024, 12, 615. [Google Scholar] [CrossRef]
- Cheng, Y.Y.; Tang, Y.; Liu, A.J.; Wei, L.; Lin, L.; Zhang, J.; Zhi, L. Clinical features and ETFDH mutations of children with late-onset glutaric aciduria type II: A report of two cases. Zhongguo Dang Dai Er Ke Za Zhi = Chin. J. Contemp. Pediatr. 2017, 19, 975–978. [Google Scholar]
- Stotz, H.U.; Brotherton, D.; Inal, J. Communication is key: Extracellular vesicles as mediators of infection and defence during host-microbe interactions in animals and plants. FEMS Microbiol. Rev. 2022, 46, fuab044. [Google Scholar] [CrossRef]
- Macchione, F.; Salviati, L.; Bordugo, A.; Vincenzi, M.; Camilot, M.; Teofoli, F.; Pancheri, E.; Zordan, R.; Bertolin, C.; Rossi, S.; et al. Multiple acyl-COA dehydrogenase deficiency in elderly carriers. J. Neurol. 2020, 267, 1414–1419. [Google Scholar] [CrossRef]
- Brandao, S.R.; Ferreira, R.; Rocha, H. Exploring the contribution of mitochondrial dynamics to multiple acyl-CoA dehydrogenase deficiency-related phenotype. Arch. Physiol. Biochem. 2021, 127, 210–216. [Google Scholar] [CrossRef]
- Angelini, C.; Burlina, A.; Blau, N.; Ferreira, C.R. Clinical and biochemical footprints of inherited metabolic disorders: X. Metabolic myopathies. Mol. Genet. Metab. 2022, 137, 213–222. [Google Scholar] [CrossRef]
- Szrok-Jurga, S.; Czumaj, A.; Turyn, J.; Hebanowska, A.; Swierczynski, J.; Sledzinski, T.; Stelmanska, E. The Physiological and Pathological Role of Acyl-CoA Oxidation. Int. J. Mol. Sci. 2023, 24, 14857. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Astiazaran-Symonds, E.; Basu, S.; Kisling, M.; Scaglia, F.; Chapman, K.A.; Wang, Y.; Vockley, J.; Ferreira, C.R. Mitochondrial energetic impairment in a patient with late-onset glutaric acidemia Type 2. Am. J. Med. Genet. A 2020, 182, 2426–2431. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Zeng, Z.L.; Yang, S.; Li, A.; Zu, X.; Liu, J. Mitochondrial Stress in Metabolic Inflammation: Modest Benefits and Full Losses. Oxid. Med. Cell. Longev. 2022, 2022, 8803404. [Google Scholar] [CrossRef]
- Picard, M.; McEwen, B.S.; Epel, E.S.; Sandi, C. An energetic view of stress: Focus on mitochondria. Front. Neuroendocrinol. 2018, 49, 72–85. [Google Scholar] [CrossRef]
- Zhu, M.C.; Li, X.M.; Zhao, N.; Yang, L.; Zhang, K.Q.; Yang, J.K. Regulatory Mechanism of Trap Formation in the Nematode-Trapping Fungi. J. Fungi 2022, 8, 406. [Google Scholar] [CrossRef]
- Angelini, C.; Tavian, D.; Missaglia, S. Heterogeneous Phenotypes in Lipid Storage Myopathy Due to ETFDH Gene Mutations. JIMD Rep. 2018, 38, 33–40. [Google Scholar] [PubMed]
- Chen, S.A.; Lin, H.C.; Schroeder, F.C.; Hsueh, Y.P. Prey sensing and response in a nematode-trapping fungus is governed by the MAPK pheromone response pathway. Genetics 2021, 217, iyaa008. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zheng, X.; Tian, M.; Zhang, K. Ammonia and Nematode Ascaroside Are Synergistic in Trap Formation in Arthrobotrys oligospora. Pathogens 2023, 12, 1114. [Google Scholar] [CrossRef]
- Hu, X.; Hoffmann, D.S.; Wang, M.; Schuhmacher, L.; Stroe, M.C.; Schreckenberger, B.; Elstner, M.; Fischer, R. GprC of the nematode-trapping fungus Arthrobotrys flagrans activates mitochondria and reprograms fungal cells for nematode hunting. Nat. Microbiol. 2024, 9, 1752–1763. [Google Scholar] [CrossRef]
- Kuo, C.Y.; Tay, R.J.; Lin, H.C.; Juan, S.C.; Vidal-Diez de Ulzurrun, G.; Chang, Y.C.; Hoki, J.; Schroeder, F.C.; Hsueh, Y.P. The nematode-trapping fungus Arthrobotrys oligospora detects prey pheromones via G protein-coupled receptors. Nat. Microbiol. 2024, 9, 1738–1751. [Google Scholar] [CrossRef]
- Xie, M.; Yang, J.; Jiang, K.; Bai, N.; Zhu, M.; Zhu, Y.; Zhang, K.Q.; Yang, J. AoBck1 and AoMkk1 Are Necessary to Maintain Cell Wall Integrity, Vegetative Growth, Conidiation, Stress Resistance, and Pathogenicity in the Nematode-Trapping Fungus Arthrobotrys oligospora. Front. Microbiol. 2021, 12, 649582. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, M.; Cui, P.; Tian, M.; Xu, Y.; Zheng, X.; Zhang, K.; Li, G.; Wang, X. Arrestin-Coding Genes Regulate Endocytosis, Sporulation, Pathogenicity, and Stress Resistance in Arthrobotrys oligospora. Front. Cell Infect. Microbiol. 2022, 12, 754333. [Google Scholar] [CrossRef] [PubMed]
- Conte, D., Jr.; MacNeil, L.T.; Walhout, A.J.M.; Mello, C.C. RNA Interference in Caenorhabditis elegans. Curr. Protoc. Mol. Biol. 2015, 109, 26.23.21–26.23.30. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolism Name | FC | KEGG Pathway Description |
|---|---|---|
| Cetraxate | 47.8841 | - |
| S-Lactoylglutathione | 1.4921 | Pyruvate metabolism |
| Kynurenic acid | 1.2806 | Tryptophan metabolism |
| Gamma-glutamyl-L-putrescine | 1.3035 | Arginine and proline metabolism |
| 2-Isopropylmalic acid | 1.1205 | Biosynthesis of secondary metabolites |
| Valine, leucine and isoleucine biosynthesis | ||
| 2-Oxocarboxylic acid | ||
| Theophylline | 1.1008 | Biosynthesis of secondary metabolites |
| Caffeine metabolism | ||
| 6-Lactoyltetrahydropterin | 1.1103 | Folate biosynthesis |
| Homogentisic acid | 1.1126 | Ubiquinone and other terpenoid-quinone biosynthesis |
| Tyrosine metabolism | ||
| Biosynthesis of cofactors | ||
| Imidazole lactic acid | 1.1166 | Histidine metabolism |
| D-xylonic acid | 1.0826 | Pentose and glucuronate interconversions |
| Ascorbate and aldarate metabolism | ||
| N-carbamoylputrescine | 1.0977 | Arginine and proline metabolism |
| 5-methoxyindoleacetate | 1.0983 | Tryptophan metabolism |
| lactose | 1.0749 | ABC transporters |
| Galactose metabolism | ||
| Indole-3-acetaldehyde | 1.0808 | Tryptophan metabolism |
| Glucosamine | 1.0607 | Amino sugar and nucleotide sugar metabolism |
| Biosynthesis of nucleotide sugars | ||
| LL-2,6-diaminopimelic acid | 1.0692 | Biosynthesis of secondary metabolites |
| D-amino acid metabolism | ||
| Biosynthesis of amino acids | ||
| Lysine biosynthesis | ||
| Biotin | 1.0596 | Biotin metabolism;ABC transporters |
| Biosynthesis of cofactors | ||
| Kynurenine | 1.0558 | Tryptophan metabolism |
| Biosynthesis of cofactors |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, Z.; Liu, J.; Zhang, X.; Wang, X. Electron-Transferring Flavoprotein and Its Dehydrogenase Required for Fungal Pathogenicity in Arthrobotrys oligospora. Int. J. Mol. Sci. 2024, 25, 10934. https://doi.org/10.3390/ijms252010934
Liu Y, Li Z, Liu J, Zhang X, Wang X. Electron-Transferring Flavoprotein and Its Dehydrogenase Required for Fungal Pathogenicity in Arthrobotrys oligospora. International Journal of Molecular Sciences. 2024; 25(20):10934. https://doi.org/10.3390/ijms252010934
Chicago/Turabian StyleLiu, Yonglan, Zhangyu Li, Junjie Liu, Xiqi Zhang, and Xin Wang. 2024. "Electron-Transferring Flavoprotein and Its Dehydrogenase Required for Fungal Pathogenicity in Arthrobotrys oligospora" International Journal of Molecular Sciences 25, no. 20: 10934. https://doi.org/10.3390/ijms252010934
APA StyleLiu, Y., Li, Z., Liu, J., Zhang, X., & Wang, X. (2024). Electron-Transferring Flavoprotein and Its Dehydrogenase Required for Fungal Pathogenicity in Arthrobotrys oligospora. International Journal of Molecular Sciences, 25(20), 10934. https://doi.org/10.3390/ijms252010934