
From Beach to the Bedside: Harnessing Mitochondrial Function in Human Diseases Using New Marine-Derived Strategies


Abstract
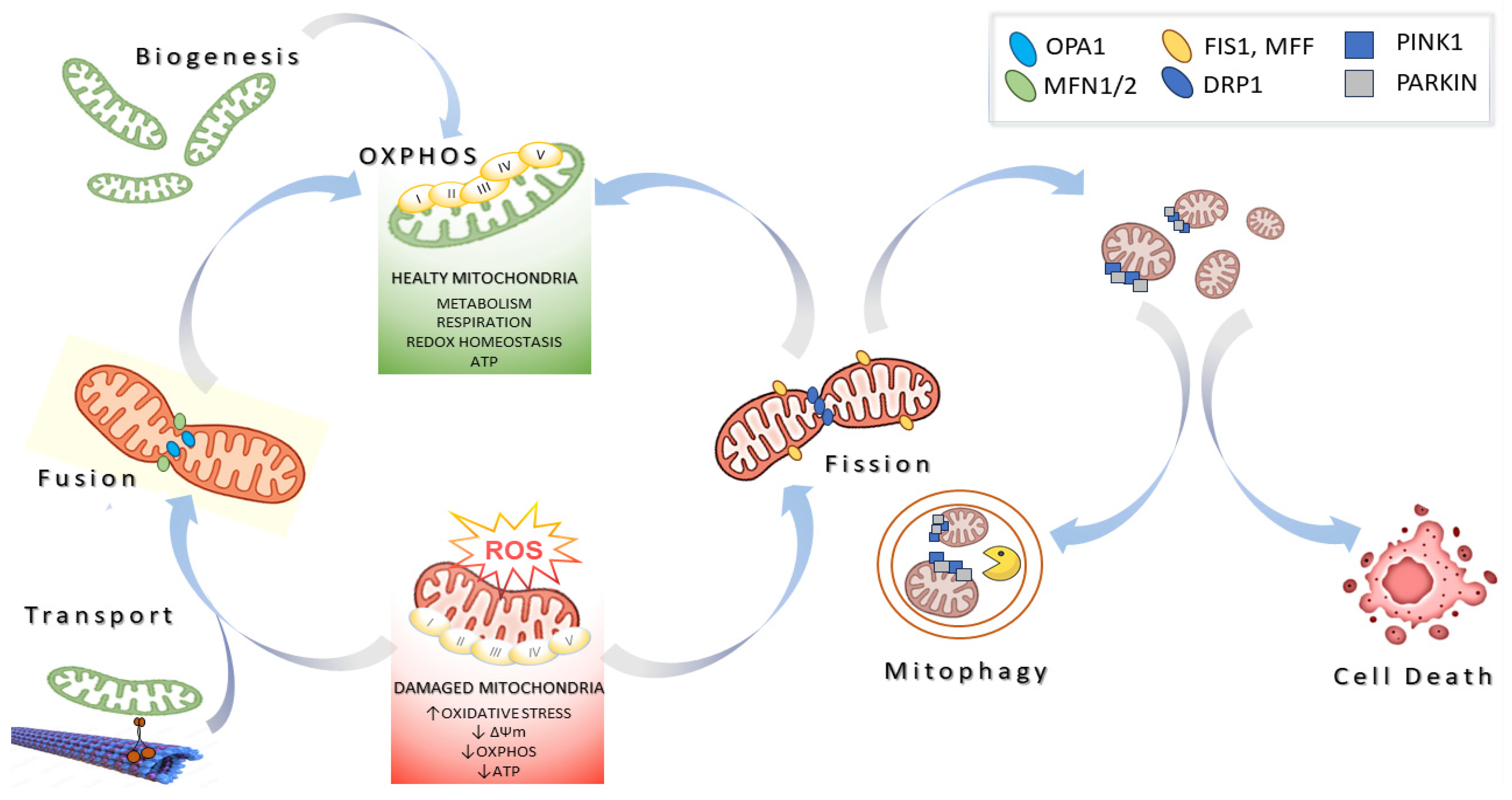
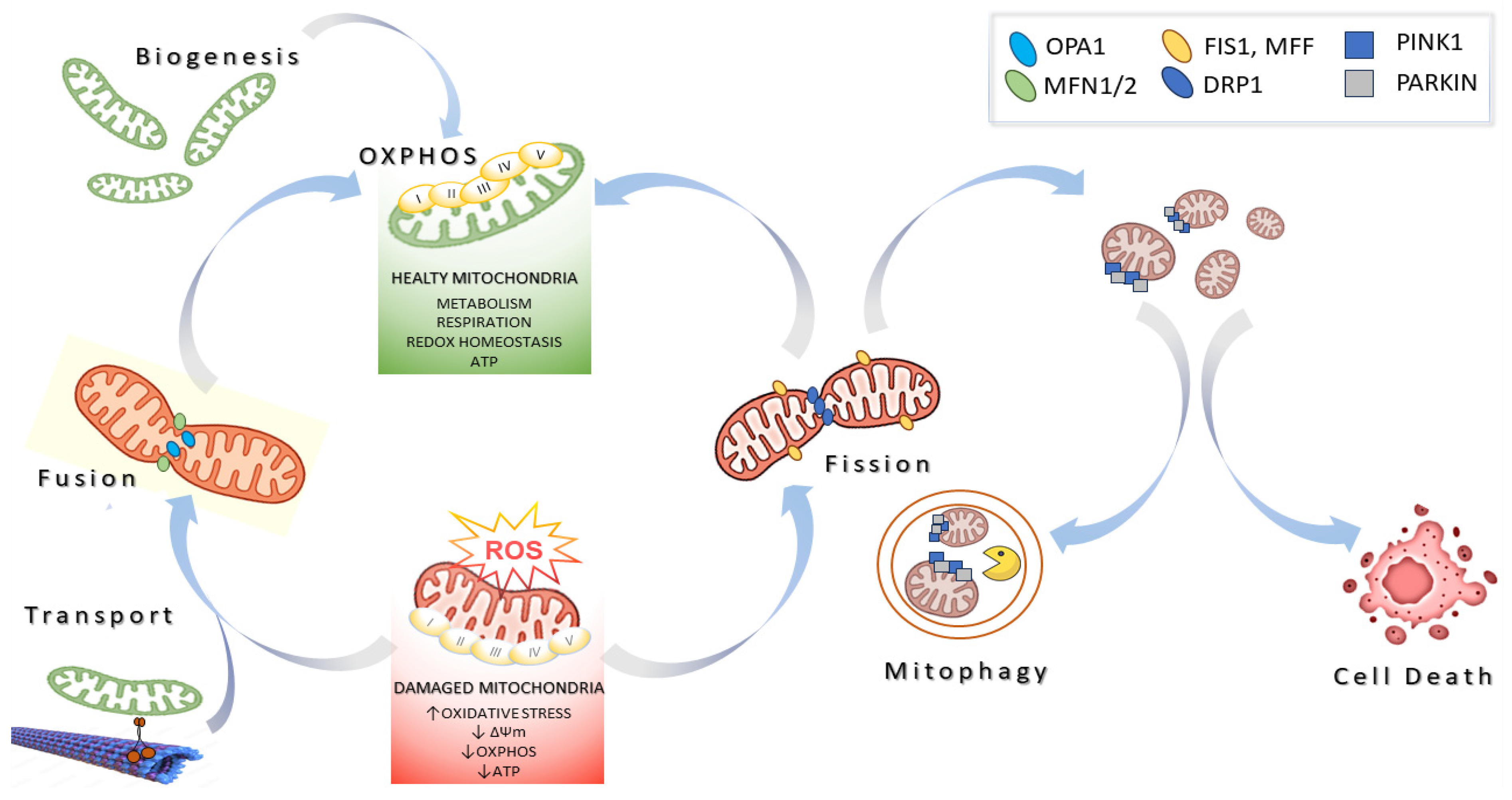
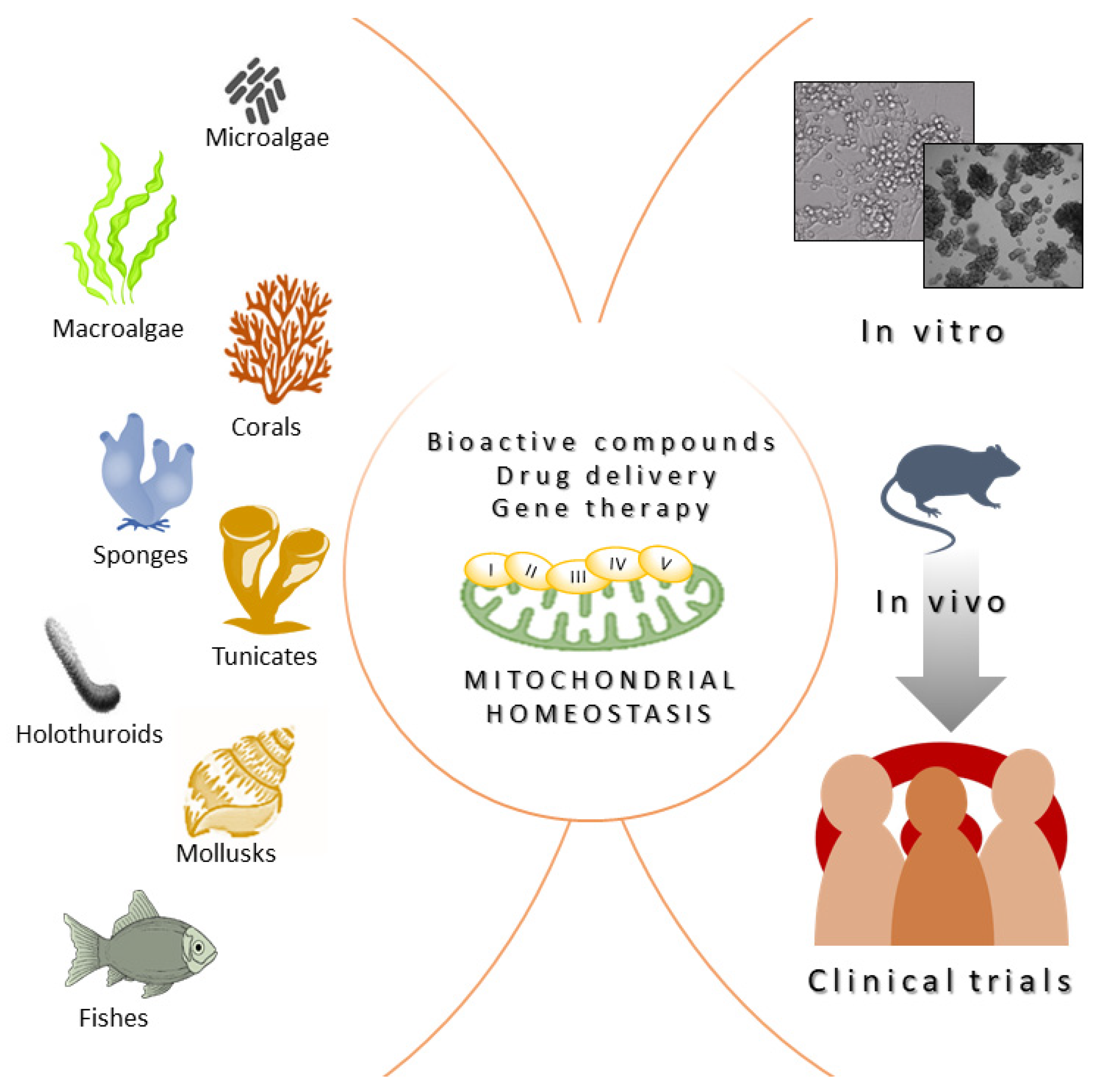
1. Introduction
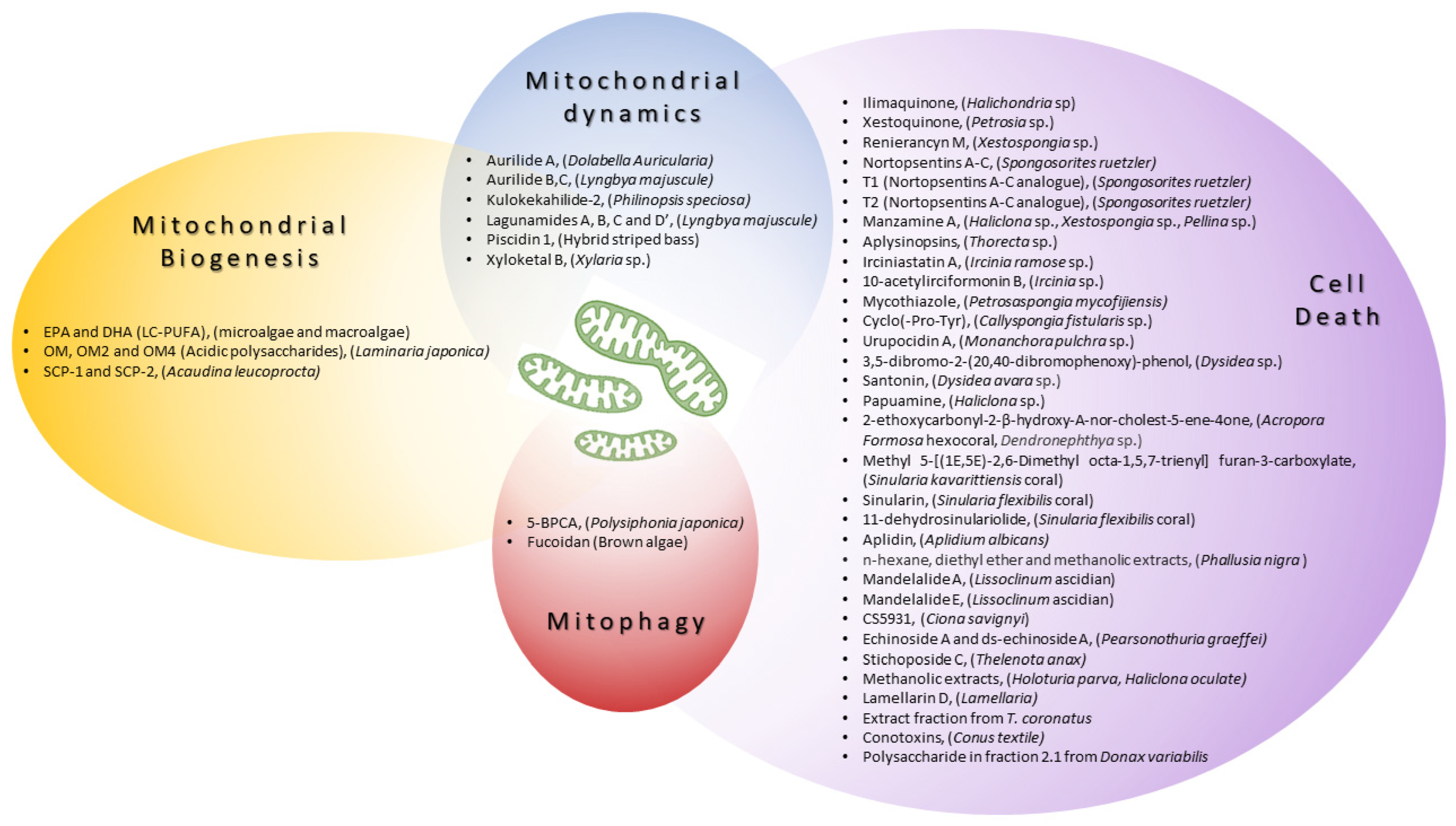
2. Modulators of Mitochondrial Biology Derived from Marine Resources
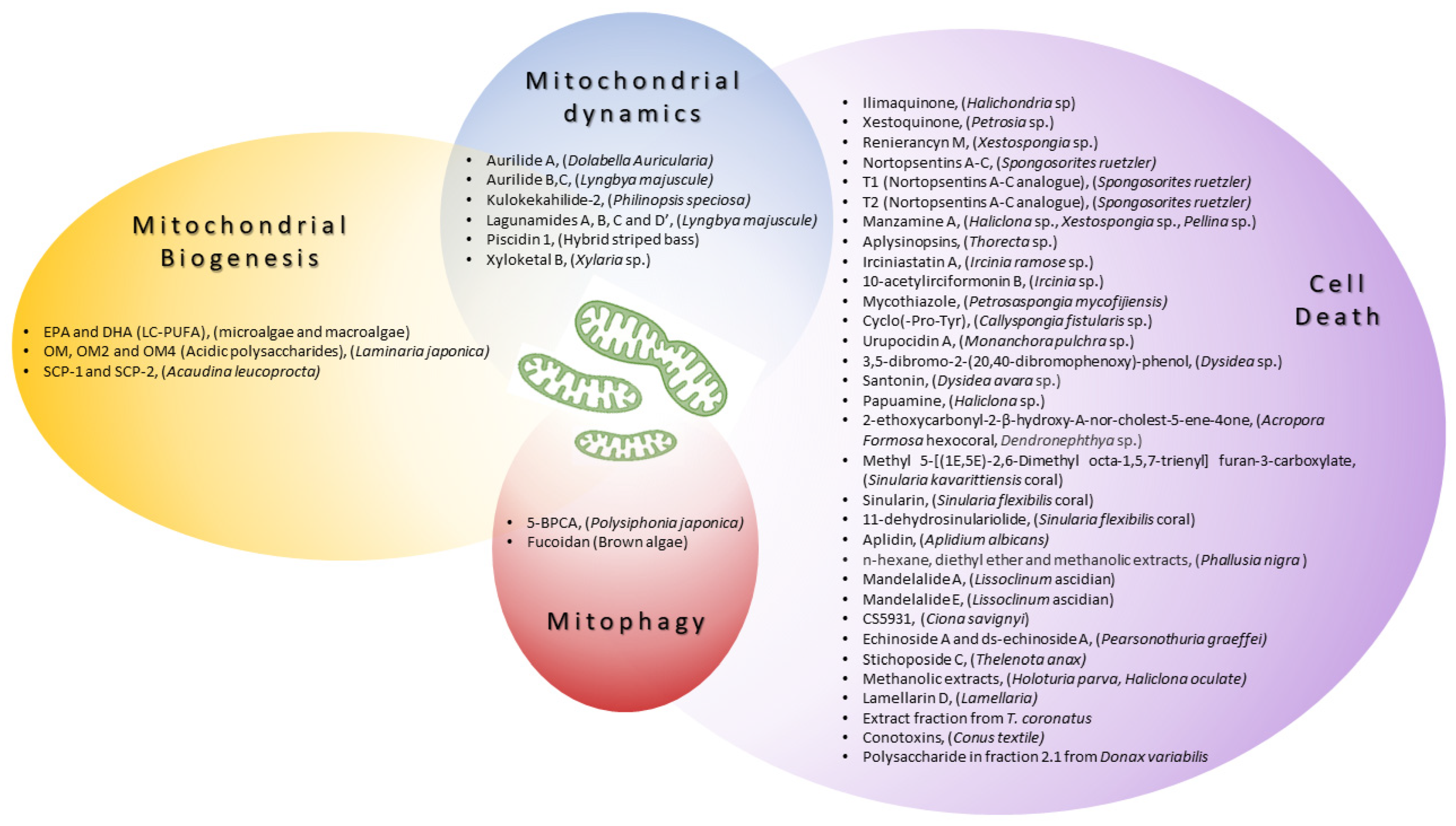
2.1. Enhancing Mitochondrial Biogenesis
2.2. Modulating Mitochondrial Dynamics
2.3. Inducing Mitophagy
2.4. The Regulation of Cell Death
2.4.1. Marine Sponges
2.4.2. Corals
2.4.3. Marine Tunicates
2.4.4. Holothuroids
2.4.5. Marine Mollusks
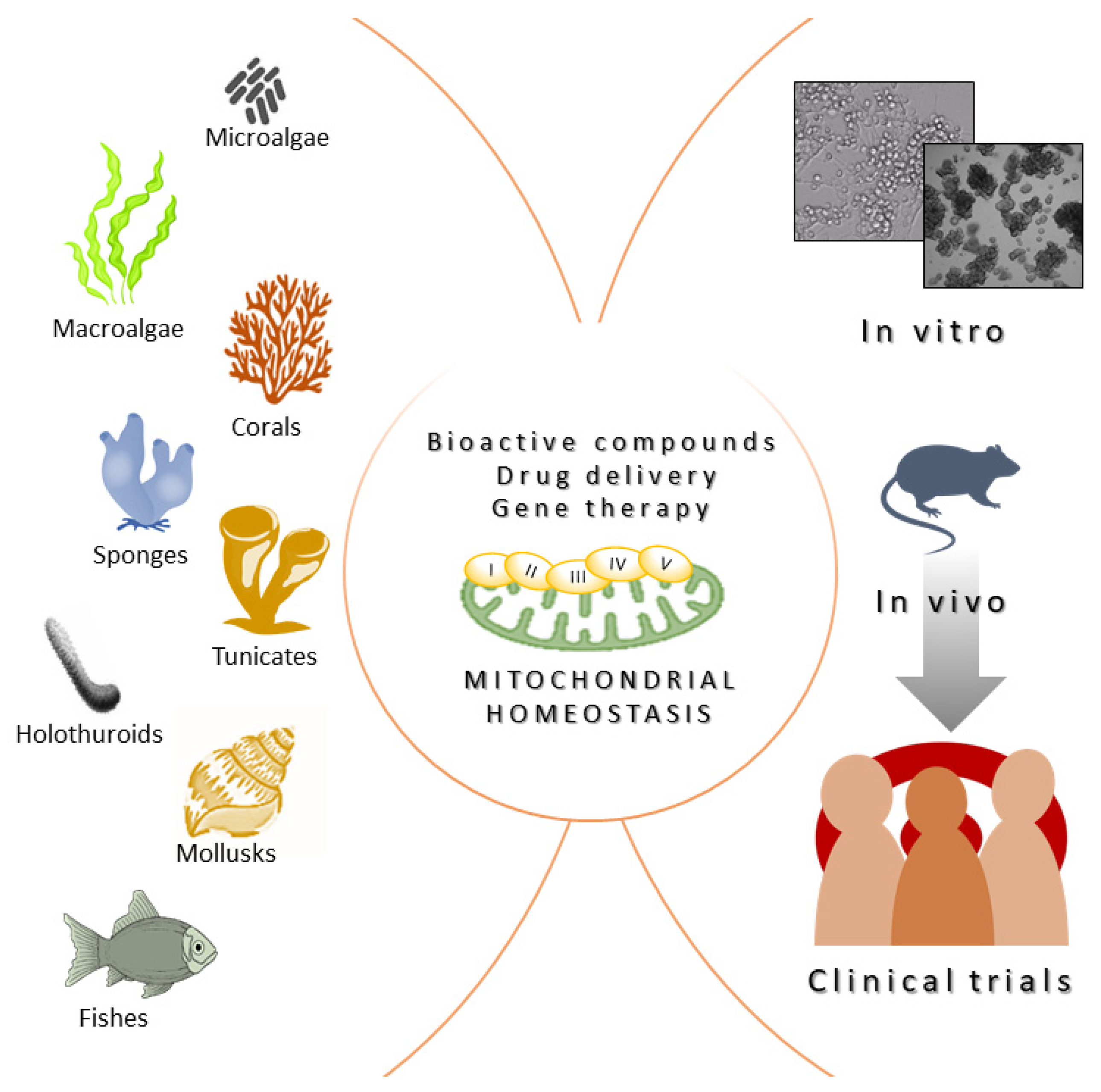
3. Gene Therapy Approaches Inspired by Marine Organism Strategies
4. Conclusions, Challenges, and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guan, S.; Zhao, L.; Peng, R. Mitochondrial Respiratory Chain Supercomplexes: From Structure to Function. Int. J. Mol. Sci. 2022, 23, 13880. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.Y.; Sun, L.; Chen, X.P.; Zhang, D.S. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; van Delft, M.F.; Dewson, G. Too much death can kill you: Inhibiting intrinsic apoptosis to treat disease. EMBO J. 2021, 40, e107341. [Google Scholar] [CrossRef] [PubMed]
- Means, R.E.; Katz, S.G. Balancing life and death: BCL-2 family members at diverse ER-mitochondrial contact sites. FEBS J. 2022, 289, 7075–7112. [Google Scholar] [CrossRef] [PubMed]
- Bock, F.J.; Tait, S.W.G. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef]
- Delettre, C.; Griffoin, J.M.; Kaplan, J.; Dollfus, H.; Lorenz, B.; Faivre, L.; Lenaers, G.; Belenguer, P.; Hamel, C.P. Mutation spectrum and splicing variants in the OPA1 gene. Hum. Genet. 2001, 109, 584–591. [Google Scholar] [CrossRef]
- Olichon, A.; ElAchouri, G.; Baricault, L.; Delettre, C.; Belenguer, P.; Lenaers, G. OPA1 alternate splicing uncouples an evolutionary conserved function in mitochondrial fusion from a vertebrate restricted function in apoptosis. Cell Death Differ. 2007, 14, 682–692. [Google Scholar] [CrossRef]
- Ban, T.; Ishihara, T.; Kohno, H.; Saita, S.; Ichimura, A.; Maenaka, K.; Oka, T.; Mihara, K.; Ishihara, N. Molecular basis of selective mitochondrial fusion by heterotypic action between OPA1 and cardiolipin. Nat. Cell Biol. 2017, 19, 856–863. [Google Scholar] [CrossRef]
- Gilkerson, R.; De La Torre, P.; St. Vallier, S. Mitochondrial OMA1 and OPA1 as Gatekeepers of Organellar Structure/Function and Cellular Stress Response. Front. Cell Dev. Biol. 2021, 9, 626117. [Google Scholar] [CrossRef]
- Consolato, F.; Maltecca, F.; Tulli, S.; Sambri, I.; Casari, G. m-AAA and i-AAA complexes coordinate to regulate OMA1, the stress-activated supervisor of mitochondrial dynamics. J. Cell Sci. 2018, 131, jcs213546. [Google Scholar] [CrossRef]
- Barrera, M.; Koob, S.; Dikov, D.; Vogel, F.; Reichert, A.S. OPA1 functionally interacts with MIC60 but is dispensable for crista junction formation. FEBS Lett. 2016, 590, 3309–3322. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.J.; Lampe, P.A.; Stojanovski, D.; Korwitz, A.; Anand, R.; Tatsuta, T.; Langer, T. Stress-induced OMA1 activation and autocatalytic turnover regulate OPA1-dependent mitochondrial dynamics. EMBO J. 2014, 33, 578–593. [Google Scholar] [CrossRef] [PubMed]
- Losón, O.C.; Song, Z.; Chen, H.; Chan, D.C. Fis1, Mff, MiD49, and MiD51 mediate Drp1 recruitment in mitochondrial fission. Mol. Biol. Cell 2013, 24, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Long, H.; Hou, L.; Feng, B.; Ma, Z.; Wu, Y.; Zeng, Y.; Cai, J.; Zhang, D.W.; Zhao, G. The mitophagy pathway and its implications in human diseases. Signal Transduct. Target. Ther. 2023, 8, 304. [Google Scholar] [CrossRef] [PubMed]
- Shutt, T.; Geoffrion, M.; Milne, R.; McBride, H.M. The intracellular redox state is a core determinant of mitochondrial fusion. EMBO Rep. 2012, 13, 909–915. [Google Scholar] [CrossRef]
- McWilliams, T.G.; Muqit, M.M. PINK1 and Parkin: Emerging themes in mitochondrial homeostasis. Curr. Opin. Cell Biol. 2017, 45, 83–91. [Google Scholar] [CrossRef]
- Jin, S.M.; Lazarou, M.; Wang, C.; Kane, L.A.; Narendra, D.P.; Youle, R.J. Mitochondrial membrane potential regulates PINK1 import and proteolytic destabilization by PARL. J. Cell Biol. 2010, 191, 933–942. [Google Scholar] [CrossRef]
- Lazarou, M.; Sliter, D.A.; Kane, L.A.; Sarraf, S.A.; Wang, C.; Burman, J.L.; Sideris, D.P.; Fogel, A.I.; Youle, R.J. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature 2015, 524, 309–314. [Google Scholar] [CrossRef]
- Yang, Y.; He, P.Y.; Zhang, Y.; Li, N. Natural Products Targeting the Mitochondria in Cancers. Molecules 2020, 26, 92. [Google Scholar] [CrossRef]
- Mitra, S.; Rauf, A.; Sutradhar, H.; Sadaf, S.; Hossain, M.J.; Soma, M.A.; Emran, T.B.; Ahmad, B.; Aljohani, A.S.M.; Al Abdulmonem, W.; et al. Potential candidates from marine and terrestrial resources targeting mitochondrial inhibition: Insights from the molecular approach. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2023, 264, 109509. [Google Scholar] [CrossRef]
- Duarte, C.M.; Agusti, S.; Barbier, E.; Britten, G.L.; Castilla, J.C.; Gattuso, J.P.; Fulweiler, R.W.; Hughes, T.P.; Knowlton, N.; Lovelock, C.E.; et al. Rebuilding marine life. Nature 2020, 580, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Lichota, A.; Gwozdzinski, K. Anticancer Activity of Natural Compounds from Plant and Marine Environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2022, 39, 1122–1171. [Google Scholar] [CrossRef] [PubMed]
- Popov, L.D. Mitochondrial biogenesis: An update. J. Cell. Mol. Med. 2020, 24, 4892–4899. [Google Scholar] [CrossRef] [PubMed]
- Halling, J.F.; Pilegaard, H. PGC-1α-mediated regulation of mitochondrial function and physiological implications. Appl. Physiol. Nutr. Metab. 2020, 45, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Ruan, D.; Shi, J.; Du, D.; Bian, C. The multifaceted roles of natural products in mitochondrial dysfunction. Front. Pharmacol. 2023, 14, 1093038. [Google Scholar] [CrossRef] [PubMed]
- Eze, C.N.; Onyejiaka, C.K.; Ihim, S.A.; Ayoka, T.O.; Aduba, C.C.; Ndukwe, J.K.; Nwaiwu, O.; Onyeaka, H. Bioactive compounds by microalgae and potentials for the management of some human disease conditions. AIMS Microbiol. 2023, 9, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Furse, S.; Virtue, S.; Snowden, S.G.; Vidal-Puig, A.; Stevenson, P.C.; Chiarugi, D.; Koulman, A. Dietary PUFAs drive diverse system-level changes in lipid metabolism. Mol. Metab. 2022, 59, 101457. [Google Scholar] [CrossRef]
- Flachs, P.; Horakova, O.; Brauner, P.; Rossmeisl, M.; Pecina, P.; Franssen-Van Hal, N.; Ruzickova, J.; Sponarova, J.; Drahota, Z.; Vlcek, C.; et al. Polyunsaturated fatty acids of marine origin upregulate mitochondrial biogenesis and induce beta-oxidation in white fat. Diabetologia 2005, 48, 2365–2375. [Google Scholar] [CrossRef]
- Xie, X.T.; Cheong, K.L. Recent advances in marine algae oligosaccharides: Structure, analysis, and potential prebiotic activities. Crit. Rev. Food Sci. Nutr. 2022, 62, 7703–7717. [Google Scholar] [CrossRef]
- Aziz, E.; Batool, R.; Khan, M.U.; Rauf, A.; Akhtar, W.; Heydari, M.; Rehman, S.; Shahzad, T.; Malik, A.; Mosavat, S.H.; et al. An overview on red algae bioactive compounds and their pharmaceutical applications. J. Complement. Integr. Med. 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Hao, C.; Zhang, L.; Liu, X.; Zhou, X.; Dun, Y.; Li, H.; Li, G.; Zhao, X.; An, Y.; et al. OM2, a Novel Oligomannuronate-Chromium(III) Complex, Promotes Mitochondrial Biogenesis and Lipid Metabolism in 3T3-L1 Adipocytes via the AMPK-PGC1α Pathway. PLoS ONE 2015, 10, e0131930. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wu, G.; Jiang, Y.; Li, B.; Feng, C.; Ge, Y.; Le, H.; Jiang, L.; Liu, H.; Shi, Y.; et al. Sea Cucumber Peptides Improved the Mitochondrial Capacity of Mice: A Potential Mechanism to Enhance Gluconeogenesis and Fat Catabolism during Exercise for Improved Antifatigue Property. Oxid. Med. Cell. Longev. 2020, 2020, 4604387. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Shi, J.; Zhong, H.; Abdullah; Zhuang, J.; Zhang, J.; Wang, J.; Zhang, X.; Feng, F. High-degree hydrolysis sea cucumber peptides improve exercise performance and exert antifatigue effect via activating the NRF2 and AMPK signaling pathways in mice. J. Funct. Foods 2021, 86, 104677. [Google Scholar] [CrossRef]
- Yang, D.; Ying, J.; Wang, X.; Zhao, T.; Yoon, S.; Fang, Y.; Zheng, Q.; Liu, X.; Yu, W.; Hua, F. Mitochondrial Dynamics: A Key Role in Neurodegeneration and a Potential Target for Neurodegenerative Disease. Front. Neurosci. 2021, 15, 654785. [Google Scholar] [CrossRef]
- Marin, W.; Marin, D.; Ao, X.; Liu, Y. Mitochondria as a therapeutic target for cardiac ischemia-reperfusion injury (Review). Int. J. Mol. Med. 2021, 47, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, K.; Mutou, T.; Shibata, T.; Itoh, T.; Fujita, T.; Takada, N.; Hayamizu, K.; Takagi, M.; Irifune, T.; Kigoshi, H.; et al. Aurilide, a cytotoxic depsipeptide from the sea hare Dolabella auricularia: Isolation, structure determination, synthesis, and biological activity. Tetrahedron 2004, 60, 8509–8527. [Google Scholar] [CrossRef]
- Suenaga, K.; Mutou, T.; Shibata, T.; Itoh, T.; Kigoshi, H.; Yamada, K. Isolation and stereostructure of aurilide, a novel cyclodepsipeptide from the Japanese sea hare Dolabella auricularia. Tetrahedron Lett. 1996, 37, 6771–6774. [Google Scholar] [CrossRef]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides B and C, cancer cell toxins from a Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef]
- Nakao, Y.; Yoshida, W.Y.; Takada, Y.; Kimura, J.; Yang, L.; Mooberry, S.L.; Scheuer, P.J. Kulokekahilide-2, a cytotoxic depsipeptide from a cephalaspidean mollusk Philinopsis speciosa. J. Nat. Prod. 2004, 67, 1332–1340. [Google Scholar] [CrossRef]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Chan, K.P.; Chen, D.Y.K.; Tan, L.T. Lagunamide C, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2011, 72, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Tan, L.T. Lagunamides A and B: Cytotoxic and antimalarial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 1810–1814. [Google Scholar] [CrossRef]
- Luo, D.; Putra, M.Y.; Ye, T.; Paul, V.J.; Luesch, H. Isolation, Structure Elucidation and Biological Evaluation of Lagunamide D: A New Cytotoxic Macrocyclic Depsipeptide from Marine Cyanobacteria. Mar. Drugs 2019, 17, 83. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, M.; Kawaguchi, S.; Fujii, N.; Ohno, H.; Oishi, S. Structure-Activity Relationship Study on Odoamide: Insights into the Bioactivities of Aurilide-Family Hybrid Peptide-Polyketides. ACS Med. Chem. Lett. 2018, 9, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Fang, W.; Leong, D.T.; Tan, L.T. Biochemical studies of the lagunamides, potent cytotoxic cyclic depsipeptides from the marine cyanobacterium Lyngbya majuscula. Mar. Drugs 2012, 10, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.I.; Murata, A.; Orihara, T.; Shirakawa, T.; Suenaga, K.; Kigoshi, H.; Uesugi, M. Marine natural product aurilide activates the OPA1-mediated apoptosis by binding to prohibitin. Chem. Biol. 2011, 18, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Semenzato, M.; Cogliati, S.; Scorrano, L. Prohibitin(g) cancer: Aurilide and killing by Opa1-dependent cristae remodeling. Chem. Biol. 2011, 18, 8–9. [Google Scholar] [CrossRef] [PubMed]
- Wasilewski, M.; Scorrano, L. The changing shape of mitochondrial apoptosis. Trends Endocrinol. Metab. 2009, 20, 287–294. [Google Scholar] [CrossRef]
- Michon, S.; Cavelier, F.; Salom-Roig, X.J. Synthesis and Biological Activities of Cyclodepsipeptides of Aurilide Family from Marine Origin. Mar. Drugs 2021, 19, 55. [Google Scholar] [CrossRef]
- Lin, H.J.; Huang, T.C.; Muthusamy, S.; Lee, J.F.; Duann, Y.F.; Lin, C.H. Piscidin-1, an antimicrobial peptide from fish (hybrid striped bass morone saxatilis x M. chrysops), induces apoptotic and necrotic activity in HT1080 cells. Zoolog. Sci. 2012, 29, 327–332. [Google Scholar] [CrossRef]
- Cheng, M.H.; Pan, C.Y.; Chen, N.F.; Yang, S.N.; Hsieh, S.; Wen, Z.H.; Chen, W.F.; Wang, J.W.; Lu, W.H.; Kuo, H.M. Piscidin-1 Induces Apoptosis via Mitochondrial Reactive Oxygen Species-Regulated Mitochondrial Dysfunction in Human Osteosarcoma Cells. Sci. Rep. 2020, 10, 5045. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, L.; Ling, C.; Li, J.; Pang, J.Y.; Lin, Y.C.; Liu, J.; Huang, R.; Wang, G.L.; Pei, Z.; et al. Marine compound Xyloketal B protects PC12 cells against OGD-induced cell damage. Brain Res. 2009, 1302, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Nigam, M.; Suleria, H.A.R.; Farzaei, M.H.; Mishra, A.P. Marine anticancer drugs and their relevant targets: A treasure from the ocean. Daru 2019, 27, 491–515. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.H.; Zhang, C.; Heo, S.J.; Jun, H.S. 5-Bromoprotocatechualdehyde Combats against Palmitate Toxicity by Inhibiting Parkin Degradation and Reducing ROS-Induced Mitochondrial Damage in Pancreatic β-Cells. Antioxidants 2021, 10, 264. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.I.; Coutinho, A.J.; Costa Lima, S.A.; Reis, S. Marine Polysaccharides in Pharmaceutical Applications: Fucoidan and Chitosan as Key Players in the Drug Delivery Match Field. Mar. Drugs 2019, 17, 654. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wei, Z.; Xue, C. Physicochemical properties of fucoidan and its applications as building blocks of nutraceutical delivery systems. Crit. Rev. Food Sci. Nutr. 2022, 62, 8935–8953. [Google Scholar] [CrossRef]
- Gao, X.; Yin, Y.; Liu, S.; Dong, K.; Wang, J.; Guo, C. Fucoidan-proanthocyanidins nanoparticles protect against cisplatin-induced acute kidney injury by activating mitophagy and inhibiting mtDNA-cGAS/STING signaling pathway. Int. J. Biol. Macromol. 2023, 245, 125541. [Google Scholar] [CrossRef]
- Zuo, W.; Kwok, H.F. Development of Marine-Derived Compounds for Cancer Therapy. Mar. Drugs 2021, 19, 342. [Google Scholar] [CrossRef]
- Nova, P.; Gomes, A.M.; Costa-Pinto, A.R. It comes from the sea: Macroalgae-derived bioactive compounds with anti-cancer potential. Crit. Rev. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Fakhri, S.; Abdian, S.; Moradi, S.Z.; Delgadillo, B.E.; Fimognari, C.; Bishayee, A. Marine Compounds, Mitochondria, and Malignancy: A Therapeutic Nexus. Mar. Drugs 2022, 20, 625. [Google Scholar] [CrossRef]
- Wang, E.; Sorolla, M.A.; Krishnan, P.D.G.; Sorolla, A. From Seabed to Bedside: A Review on Promising Marine Anticancer Compounds. Biomolecules 2020, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Torres, V.; Encinar, J.A.; Herranz-López, M.; Pérez-Sánchez, A.; Galiano, V.; Barrajón-Catalán, E.; Micol, V. An Updated Review on Marine Anticancer Compounds: The Use of Virtual Screening for the Discovery of Small-Molecule Cancer Drugs. Molecules 2017, 22, 1037. [Google Scholar] [CrossRef]
- Bai, L.Y.; Su, J.H.; Chiu, C.F.; Lin, W.Y.; Hu, J.L.; Feng, C.H.; Shu, C.W.; Weng, J.R. Antitumor Effects of a Sesquiterpene Derivative from Marine Sponge in Human Breast Cancer Cells. Mar. Drugs 2021, 19, 244. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Lu, M.C.; Hsu, K.C.; El-Shazly, M.; Shih, S.P.; Lien, S.T.; Kuo, F.W.; Yang, S.C.; Chen, C.L.; Yang, Y.C.S.H. The Antileukemic Effect of Xestoquinone, A Marine-Derived Polycyclic Quinone-Type Metabolite, Is Mediated through ROS-Induced Inhibition of HSP-90. Molecules 2021, 26, 7037. [Google Scholar] [CrossRef]
- Sirimangkalakitti, N.; Chamni, S.; Suwanborirux, K.; Chanvorachote, P. Renieramycin M Attenuates Cancer Stem Cell-like Phenotypes in H460 Lung Cancer Cells. Anticancer Res. 2017, 37, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Pinkhien, T.; Maiuthed, A.; Chamni, S.; Suwanborirux, K.; Saito, N.; Chanvorachote, P. Bishydroquinone Renieramycin M Induces Apoptosis of Human Lung Cancer Cells Through a Mitochondria-dependent Pathway. Anticancer Res. 2016, 36, 6327–6333. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Pennati, M.; Barraja, P.; Montalbano, A.; Parrino, B.; Spano, V.; Lopergolo, A.; Sbarra, S.; Doldi, V.; Zaffaroni, N.; et al. Synthesis and antiproliferative activity of substituted 3[2-(1H-indol-3-yl)-1,3-thiazol-4-yl]-1H-pyrrolo[3,2-b]pyridines, marine alkaloid nortopsentin analogues. Curr. Med. Chem. 2014, 21, 1654–1666. [Google Scholar] [CrossRef]
- Diana, P.; Carbone, A.; Barraja, P.; Martorana, A.; Gia, O.; DallaVia, L.; Cirrincione, G. 3,5-bis(3′-indolyl)pyrazoles, analogues of marine alkaloid nortopsentin: Synthesis and antitumor properties. Bioorg. Med. Chem. Lett. 2007, 17, 6134–6137. [Google Scholar] [CrossRef]
- Lin, L.C.; Kuo, T.T.; Chang, H.Y.; Liu, W.S.; Hsia, S.M.; Huang, T.C. Manzamine A Exerts Anticancer Activity against Human Colorectal Cancer Cells. Mar. Drugs 2018, 16, 252. [Google Scholar] [CrossRef]
- Song, S.; Kim, S.; El-Sawy, E.R.; Cerella, C.; Orlikova-Boyer, B.; Kirsch, G.; Christov, C.; Dicato, M.; Diederich, M. Anti-Leukemic Properties of Aplysinopsin Derivative EE-84 Alone and Combined to BH3 Mimetic A-1210477. Mar. Drugs 2021, 19, 285. [Google Scholar] [CrossRef]
- Chinen, T.; Nagumo, Y.; Watanabe, T.; Imaizumi, T.; Shibuya, M.; Kataoka, T.; Kanoh, N.; Iwabuchi, Y.; Usui, T. Irciniastatin A induces JNK activation that is involved in caspase-8-dependent apoptosis via the mitochondrial pathway. Toxicol. Lett. 2010, 199, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.B.; Mahdi, F.; Liu, Y.; Coothankandaswamy, V.; Jekabsons, M.B.; Johnson, T.A.; Sashidhara, K.V.; Crews, P.; Nagle, D.G.; Zhou, Y.D. The marine sponge metabolite mycothiazole: A novel prototype mitochondrial complex I inhibitor. Bioorg. Med. Chem. 2010, 18, 5988–5994. [Google Scholar] [CrossRef] [PubMed]
- Karanam, G.; Arumugam, M.K. Reactive oxygen species generation and mitochondrial dysfunction for the initiation of apoptotic cell death in human hepatocellular carcinoma HepG2 cells by a cyclic dipeptide Cyclo(-Pro-Tyr). Mol. Biol. Rep. 2020, 47, 3347–3359. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.A.; Kudryashova, E.K.; Kaune, M.; Makarieva, T.N.; Shubina, L.K.; Busenbender, T.; Denisenko, V.A.; Popov, R.S.; Hauschild, J.; Fedorov, S.N.; et al. Urupocidin C: A new marine guanidine alkaloid which selectively kills prostate cancer cells via mitochondria targeting. Sci. Rep. 2020, 10, 9764. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.; Shin, D.; Kamiya, K.; Ishida, R.; Setiawan, A.; Kotoku, N.; Kobayashi, M. Marine spongean polybrominated diphenyl ethers, selective growth inhibitors against the cancer cells adapted to glucose starvation, inhibits mitochondrial complex II. J. Nat. Med. 2017, 71, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Aghvami, M.; Keshavarz, A.; Nazemi, M.; Zarei, M.H.; Pourahmad, J. Selective Cytotoxicity of α-Santonin from the Persian Gulf Sponge Dysidea Avara on Pediatric ALL B-lymphocytes via Mitochondrial Targeting. Asian Pac. J. Cancer Prev. 2018, 19, 2149–2154. [Google Scholar] [CrossRef]
- Kanno, S.I.; Yomogida, S.; Tomizawa, A.; Yamazaki, H.; Ukai, K.; Mangindaan, R.E.P.; Namikoshi, M.; Ishikawa, M. Papuamine causes autophagy following the reduction of cell survival through mitochondrial damage and JNK activation in MCF-7 human breast cancer cells. Int. J. Oncol. 2013, 43, 1413–1419. [Google Scholar] [CrossRef]
- Vaikundamoorthy, R.; Sundaramoorthy, R.; Krishnamoorthy, V.; Vilwanathan, R.; Rajendran, R. Marine steroid derived from Acropora formosa enhances mitochondrial-mediated apoptosis in non-small cell lung cancer cells. Tumour Biol. 2016, 37, 10517–10531. [Google Scholar] [CrossRef]
- Arepalli, S.K.; Sridhar, V.; Rao, J.V.; Kennady, P.K.; Venkateswarlu, Y. Furano-sesquiterpene from soft coral, Sinularia kavarittiensis: Induces apoptosis via the mitochondrial-mediated caspase-dependent pathway in THP-1, leukemia cell line. Apoptosis 2009, 14, 729–740. [Google Scholar] [CrossRef]
- Su, T.R.; Tsai, F.J.; Lin, J.J.; Huang, H.H.; Chiu, C.C.; Su, J.H.; Yang, Y.T.; Chen, J.Y.F.; Wong, B.S.; Wu, Y.J. Induction of apoptosis by 11-dehydrosinulariolide via mitochondrial dysregulation and ER stress pathways in human melanoma cells. Mar. Drugs 2012, 10, 1883–1898. [Google Scholar] [CrossRef]
- Ko, C.Y.; Shih, P.C.; Huang, P.W.; Lee, Y.H.; Chen, Y.F.; Tai, M.H.; Liu, C.H.; Wen, Z.H.; Kuo, H.M. Sinularin, an Anti-Cancer Agent Causing Mitochondria-Modulated Apoptosis and Cytoskeleton Disruption in Human Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 3946. [Google Scholar] [CrossRef]
- Humeniuk, R.; Menon, L.G.; Mishra, P.J.; Saydam, G.; Longo-Sorbello, G.S.A.; Elisseyeff, Y.; Lewis, L.D.; Aracil, M.; Jimeno, J.; Bertino, J.R.; et al. Aplidin synergizes with cytosine arabinoside: Functional relevance of mitochondria in Aplidin-induced cytotoxicity. Leukemia 2007, 21, 2399–2405. [Google Scholar] [CrossRef] [PubMed]
- Taddei, M.L.; Chiarugi, P.; Cuevas, C.; Ramponi, G.; Raugei, G. Oxidation and inactivation of low molecular weight protein tyrosine phosphatase by the anticancer drug Aplidin. Int. J. Cancer 2006, 118, 2082–2088. [Google Scholar] [CrossRef] [PubMed]
- Arast, Y.; Razi, N.S.; Seydi, E.; Naserzadeh, P.; Nazemi, M.; Pourahmad, J. Selective Toxicity of Non Polar Bioactive Compounds of Persian Gulf Sea Squirt Phallusia Nigra on Skin Mitochondria Isolated from Rat Model of Melanoma. Asian Pac. J. Cancer Prev. 2017, 18, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Sikorska, J.; Hau, A.M.; Anklin, C.; Parker-Nance, S.; Davies-Coleman, M.T.; Ishmael, J.E.; McPhail, K.L. Mandelalides A–D, cytotoxic macrolides from a new Lissoclinum species of South African tunicate. J. Org. Chem. 2012, 77, 6066–6075. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.; Serrill, J.D.; Sikorska, J.; Ye, T.; Ishmael, J.E.; McPhail, K.L. Discovery of Mandelalide E and Determinants of Cytotoxicity for the Mandelalide Series. Org. Lett. 2016, 18, 1374–1377. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, C.; Liu, H.; Wang, F.; Zheng, L.; Zhao, J.; Chu, E.; Lin, X. A novel polypeptide extracted from Ciona savignyi induces apoptosis through a mitochondrial-mediated pathway in human colorectal carcinoma cells. Clin. Color. Cancer 2012, 11, 207–214. [Google Scholar] [CrossRef]
- Salimi, A.; Rahimitabar, N.; Vazirizadeh, A.; Adhami, V.; Pourahmad, J. Persian Gulf Snail Crude Venom (Conus textile): A Potential Source of Anti-Cancer Therapeutic Agents for Glioblastoma through Mitochondrial-Mediated Apoptosis. Asian Pac. J. Cancer Prev. 2021, 22, 49–57. [Google Scholar] [CrossRef]
- Zhao, Q.; Xue, Y.; Wang, J.-F.; Li, H.; Long, T.-T.; Li, Z.; Wang, Y.-M.; Dong, P.; Xue, C.-H. In vitro and in vivo anti-tumour activities of echinoside A and ds-echinoside A from Pearsonothuria graeffei. J. Sci. Food Agric. 2012, 92, 965–974. [Google Scholar] [CrossRef]
- Yun, S.H.; Park, E.S.; Shin, S.W.; Na, Y.W.; Han, J.Y.; Jeong, J.S.; Shastina, V.V.; Stonik, V.A.; Park, J.I.; Kwak, J.Y. Stichoposide C induces apoptosis through the generation of ceramide in leukemia and colorectal cancer cells and shows in vivo antitumor activity. Clin. Cancer Res. 2012, 18, 5934–5948. [Google Scholar] [CrossRef]
- Seydi, E.; Motallebi, A.; Dastbaz, M.; Dehghan, S.; Salimi, A.; Nazemi, M.; Pourahmad, J. Selective Toxicity of Persian Gulf Sea Cucumber (Holothuria parva) and Sponge (Haliclona oculata) Methanolic Extracts on Liver Mitochondria Isolated from an Animal Model of Hepatocellular Carcinoma. Hepat. Mon. 2015, 15, e33073. [Google Scholar] [CrossRef] [PubMed]
- Ballot, C.; Kluza, J.; Lancel, S.; Martoriati, A.; Hassoun, S.M.; Mortier, L.; Vienne, J.C.; Briand, G.; Formstecher, P.; Bailly, C.; et al. Inhibition of mitochondrial respiration mediates apoptosis induced by the anti-tumoral alkaloid lamellarin D. Apoptosis 2010, 15, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Ballot, C.; Kluza, J.; Martoriati, A.; Nyman, U.; Formstecher, P.; Joseph, B.; Bailly, C.; Marchetti, P. Essential role of mitochondria in apoptosis of cancer cells induced by the marine alkaloid Lamellarin D. Mol. Cancer Ther. 2009, 8, 3307–3317. [Google Scholar] [CrossRef] [PubMed]
- Zangeneh, F.; Vazirizadeh, A.; Mirshamsi, M.R.; Fakhri, A.; Faizi, M.; Pourahmad, J. Induction of apoptosis by an extract of persian gulf marine mollusc, Turbo coronatus through the production of reactive oxygen species in mouse melanoma cells. Asian Pac. J. Cancer Prev. 2018, 19, 3479–3488. [Google Scholar] [CrossRef] [PubMed]
- Sahayanathan, G.J.; Padmanaban, D.; Raja, K.; Chinnasamy, A. Anticancer effect of purified polysaccharide from marine clam Donax variabilis on A549 cells. J. Food Biochem. 2020, 44, e13486. [Google Scholar] [CrossRef] [PubMed]
- Shih, H.C.; El-Shazly, M.; Juan, Y.S.; Chang, C.Y.; Su, J.H.; Chen, Y.C.; Shih, S.P.; Chen, H.M.; Wu, Y.C.; Lu, M.C. Cracking the cytotoxicity code: Apoptotic induction of 10-acetylirciformonin B is mediated through ROS generation and mitochondrial dysfunction. Mar. Drugs 2014, 12, 3072–3090. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.F.; Chakraborty, C.; Sung, C.S.; Feng, C.W.; Jean, Y.H.; Lin, Y.Y.; Hung, H.C.; Huang, T.Y.; Huang, S.Y.; Su, T.M.; et al. Neuroprotection by marine-derived compound, 11-dehydrosinulariolide, in an in vitro Parkinson’s model: A promising candidate for the treatment of Parkinson’s disease. Naunyn. Schmiedebergs. Arch. Pharmacol. 2012, 385, 265–275. [Google Scholar] [CrossRef]
- Nazari, M.; Serrill, J.D.; Wan, X.; Nguyen, M.H.; Anklin, C.; Gallegos, D.A.; Smith, A.B.; Ishmael, J.E.; McPhail, K.L. New Mandelalides Expand a Macrolide Series of Mitochondrial Inhibitors. J. Med. Chem. 2017, 60, 7850–7862. [Google Scholar] [CrossRef]
- Wargasetia, T.L. Widodo Mechanisms of cancer cell killing by sea cucumber-derived compounds. Investig. New Drugs 2017, 35, 820–826. [Google Scholar] [CrossRef]
- Jacobs, H.T.; Ballard, J.W.O. What physiological role(s) does the alternative oxidase perform in animals? Biochim. Biophys. Acta-Bioenerg. 2022, 1863, 148556. [Google Scholar] [CrossRef]
- Dry, I.B.; Moore, A.L.; Day, D.A.; Wiskich, J.T. Regulation of alternative pathway activity in plant mitochondria: Nonlinear relationship between electron flux and the redox poise of the quinone pool. Arch. Biochem. Biophys. 1989, 273, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Hakkaart, G.A.J.; Dassa, E.P.E.P.; Jacobs, H.T.; Rustin, P. Allotopic expression of a mitochondrial alternative oxidase confers cyanide resistance to human cell respiration. EMBO Rep. 2006, 7, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.; Fernández-Ayala, D.J.M.; Stefanatos, R.K.A.; Jacobs, H.T. Mitochondrial ROS production correlates with, but does not directly regulate lifespan in Drosophila. Aging 2010, 2, 200–223. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ayala, D.J.M.; Sanz, A.; Vartiainen, S.; Kemppainen, K.K.; Babusiak, M.; Mustalahti, E.; Costa, R.; Tuomela, T.; Zeviani, M.; Chung, J.; et al. Expression of the Ciona intestinalis alternative oxidase (AOX) in Drosophila complements defects in mitochondrial oxidative phosphorylation. Cell Metab. 2009, 9, 449–460. [Google Scholar] [CrossRef]
- Humphrey, D.M.; Parsons, R.B.; Ludlow, Z.N.; Riemensperger, T.; Esposito, G.; Verstreken, P.; Jacobs, H.T.; Birman, S.; Hirth, F. Alternative oxidase rescues mitochondria-mediated dopaminergic cell loss in Drosophila. Hum. Mol. Genet. 2012, 21, 2698–2712. [Google Scholar] [CrossRef] [PubMed]
- Kemppainen, K.K.; Rinne, J.; Sriram, A.; Lakanmaa, M.; Zeb, A.; Tuomela, T.; Popplestone, A.; Singh, S.; Sanz, A.; Rustin, P.; et al. Expression of alternative oxidase in Drosophila ameliorates diverse phenotypes due to cytochrome oxidase deficiency. Hum. Mol. Genet. 2014, 23, 2078–2093. [Google Scholar] [CrossRef] [PubMed]
- El-Khoury, R.; Dufour, E.; Rak, M.; Ramanantsoa, N.; Grandchamp, N.; Csaba, Z.; Duvillié, B.; Bénit, P.; Gallego, J.; Gressens, P.; et al. Alternative oxidase expression in the mouse enables bypassing cytochrome c oxidase blockade and limits mitochondrial ROS overproduction. PLoS Genet. 2013, 9, e1003182. [Google Scholar] [CrossRef]
- Viscomi, C.; Moore, A.L.; Zeviani, M.; Szibor, M. Xenotopic expression of alternative oxidase (AOX) to study mechanisms of mitochondrial disease. Biochim. Biophys. Acta. Bioenerg. 2023, 1864, 148947. [Google Scholar] [CrossRef]
- Szibor, M.; Dhandapani, P.K.; Dufour, E.; Holmström, K.M.; Zhuang, Y.; Salwig, I.; Wittig, I.; Heidler, J.; Gizatullina, Z.; Gainutdinov, T.; et al. Broad AOX expression in a genetically tractable mouse model does not disturb normal physiology. Dis. Model. Mech. 2017, 10, 163–171. [Google Scholar] [CrossRef]
- Dogan, S.A.; Cerutti, R.; Benincá, C.; Brea-Calvo, G.; Jacobs, H.T.; Zeviani, M.; Szibor, M.; Viscomi, C. Perturbed Redox Signaling Exacerbates a Mitochondrial Myopathy. Cell Metab. 2018, 28, 764–775.e5. [Google Scholar] [CrossRef]
- Giordano, L.; Farnham, A.; Dhandapani, P.K.; Salminen, L.; Bhaskaran, J.; Voswinckel, R.; Rauschkolb, P.; Scheibe, S.; Sommer, N.; Beisswenger, C.; et al. Alternative Oxidase Attenuates Cigarette Smoke-induced Lung Dysfunction and Tissue Damage. Am. J. Respir. Cell Mol. Biol. 2019, 60, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Dhandapani, P.K.; Begines-Moreno, I.M.; Brea-Calvo, G.; Gärtner, U.; Graeber, T.G.; Javier Sanchez, G.; Morty, R.E.; Schönig, K.; Hoeve, J.T.; Wietelmann, A.; et al. Hyperoxia but not AOX expression mitigates pathological cardiac remodeling in a mouse model of inflammatory cardiomyopathy. Sci. Rep. 2019, 9, 12741. [Google Scholar] [CrossRef] [PubMed]
- Szibor, M.; Schreckenberg, R.; Gizatullina, Z.; Dufour, E.; Wiesnet, M.; Dhandapani, P.K.; Debska-Vielhaber, G.; Heidler, J.; Wittig, I.; Nyman, T.A.; et al. Respiratory chain signalling is essential for adaptive remodelling following cardiac ischaemia. J. Cell. Mol. Med. 2020, 24, 3534–3548. [Google Scholar] [CrossRef] [PubMed]
- Sommer, N.; Alebrahimdehkordi, N.; Pak, O.; Knoepp, F.; Strielkov, I.; Scheibe, S.; Dufour, E.; Andjelković, A.; Sydykov, A.; Saraji, A.; et al. Bypassing mitochondrial complex III using alternative oxidase inhibits acute pulmonary oxygen sensing. Sci. Adv. 2020, 6, eaba0694. [Google Scholar] [CrossRef] [PubMed]
- Visapää, I.; Fellman, V.; Vesa, J.; Dasvarma, A.; Hutton, J.L.; Kumar, V.; Payne, G.S.; Makarow, M.; Van Coster, R.; Taylor, R.W.; et al. GRACILE syndrome, a lethal metabolic disorder with iron overload, is caused by a point mutation in BCS1L. Am. J. Hum. Genet. 2002, 71, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Vizarra, E.; Zeviani, M. Nuclear gene mutations as the cause of mitochondrial complex III deficiency. Front. Genet. 2015, 6, 134. [Google Scholar] [CrossRef]
- Kotarsky, H.; Karikoski, R.; Mörgelin, M.; Marjavaara, S.; Bergman, P.; Zhang, D.L.; Smet, J.; van Coster, R.; Fellman, V. Characterization of complex III deficiency and liver dysfunction in GRACILE syndrome caused by a BCS1L mutation. Mitochondrion 2010, 10, 497–509. [Google Scholar] [CrossRef]
- Rajendran, J.; Purhonen, J.; Tegelberg, S.; Smolander, O.; Mörgelin, M.; Rozman, J.; Gailus-Durner, V.; Fuchs, H.; de Angelis, M.H.; Auvinen, P.; et al. Alternative oxidase-mediated respiration prevents lethal mitochondrial cardiomyopathy. EMBO Mol. Med. 2019, 11, e9456. [Google Scholar] [CrossRef]
- Purhonen, J.; Rajendran, J.; Tegelberg, S.; Smolander, O.P.; Pirinen, E.; Kallijärvi, J.; Fellman, V. NAD+ repletion produces no therapeutic effect in mice with respiratory chain complex III deficiency and chronic energy deprivation. FASEB J. 2018, 32, 5913–5926. [Google Scholar] [CrossRef]
- May, B.; Young, L.; Moore, A.L. Structural insights into the alternative oxidases: Are all oxidases made equal? Biochem. Soc. Trans. 2017, 45, 731–740. [Google Scholar] [CrossRef]
- Voultsiadou Eleni, E. Therapeutic properties and uses of marine invertebrates in the ancient Greek world and early Byzantium. J. Ethnopharmacol. 2010, 130, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhao, H.; Li, Y. Mitochondrial dynamics in health and disease: Mechanisms and potential targets. Signal Transduct. Target. Ther. 2023, 8, 333. [Google Scholar] [CrossRef] [PubMed]
- Saide, A.; Lauritano, C.; Ianora, A. A Treasure of Bioactive Compounds from the Deep Sea. Biomedicines 2021, 9, 1556. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Almeida, M.; Coelho, A.V.; Cutignano, A.; Gonçalves, L.G.; Hansen, E.; Khnykin, D.; Mass, T.; Ramšak, A.; Rocha, M.S.; et al. Biomaterials and Bioactive Natural Products from Marine Invertebrates: From Basic Research to Innovative Applications. Mar. Drugs 2022, 20, 219. [Google Scholar] [CrossRef] [PubMed]
- García-Poza, S.; Leandro, A.; Cotas, C.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. The Evolution Road of Seaweed Aquaculture: Cultivation Technologies and the Industry 4.0. Int. J. Environ. Res. Public Health 2020, 17, 6528. [Google Scholar] [CrossRef]
- Ding, Q.; Ye, C. Microbial cell factories based on filamentous bacteria, yeasts, and fungi. Microb. Cell Fact. 2023, 22, 20. [Google Scholar] [CrossRef]
- Fatehullah, A.; Tan, S.H.; Barker, N. Organoids as an in vitro model of human development and disease. Nat. Cell Biol. 2016, 18, 246–254. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Compound(s) | Marine Organism | Mechanism of Action Regarding Mitochondria | Cell Line or Model of Disease Used in Preclinical Studies |
|---|---|---|---|
| Mitochondrial Biogenesis | |||
| n-3 PUFA eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) | Microalgae and macroalgae | ↑PGC1-α, ↑NRF1, ↑mitochondrial biogenesis | C57BL/6J epididymal fat [24] |
| oligomannuronate (OM) and OM-chromium (III) complexes (OM2 and OM4) | Laminaria japonica | ↑PGC1-α, ↑mitochondrial function, ↑mitochondrial biogenesis | C2C12, 3T3-L1 [25] |
| SCP-1 and SCP-2 | Acaudina leucoprocta | ↑AMPK/PGC1-α, ↑NRF2, ↑mitochondrial biogenesis, ↓oxidative stress | Fatigue test in ICR mice [26] |
| Mitochondrial dynamics | |||
| Aurilide A | Dolabella auricularia | Accelerate OPA-1 processing, mitochondrial fragmentation, and the release of CytC [27] | HeLa S3, NCI60 panel [28] |
| Aurilide B, C | Lyngbya majuscule | HCT-8, P388, A549, SK-OV-3, PC-3 [29] | |
| Kulokekahilide-2 | Philinopsis speciosa | P388, SK-OV-3, MDA-MB-435 [30] | |
| Lagunamides A, B, C and D | L. majuscule | P388, A549, PC3, HCT8, SK-OV3, HCT8, MCF7 [31,32] | |
| Piscidin-1 | Hybrid striped bass | ↓MFN1, ↓MFN2, ↓OPA1, ↓OXPHOS, ↑DRP1, ↑FIS1, ↑mtROS, mitochondrial dysfunction, apoptosis | MG63 [33,34] |
| Xyloketal B | Xylaria sp. | ↑Drp1, ↓mitochondrial fregmentation, ↓mitochondrial superoxide production | In vitro model of ischemic stroke in PC12 [35] |
| Mitophagy | |||
| 5-BPCA | Polysiphonia japonica | The preservation of PARKIN expression and stabilization of mitochondrial morphology | Model of palmitate (PA)-induced lipotoxicity in a rat pancreatic β-cell line (Ins-1 cells) [36] |
| Fucoidan: treatment with Fucoidan nanoparticles loaded with proanthocyanidins | Brown algae | ↑PINK1, ↑PARKIN, ↓mtDNA release | A model of cisplatin-induced damage in vitro (HK-2 cells) and in vivo (Kunming mice) [37] |
| Compound(s) | Marine Organism | In Vitro/In Vivo Models | Mechanism of Action Regarding Mitochondria | Disease Area |
|---|---|---|---|---|
| Ilimaquinone | Halichondria sp. | MCF-7, MDA-MB-231 | caspase activation, ↑ROS, ↓Δψm | Breast cancer [38] |
| Xestoquinone | Petrosia sp. | Molt-4, K562, Sup-T1 | ↑ROS, ↓HSP90 | Leukemia [39] |
| Renieranycin M | Xestospongia sp. | H460 | ↑BAX, ↓MCL1, ↓BCL2, caspase activation | Lung cancer [40,41] |
| Nortopsentins A-C | Spongosorites ruetzler | P388 cells | caspase activation | Leukemia [42] |
| T1 (Nortopsentins A-C analogue) | Spongosorites ruetzleri | HCT-116 colorectal cancer cells | caspase activation, ↑mitochondrial trans-membrane potential | Colon cancer [43] |
| T2 (Nortopsentins A-C analogue) | Spongosorites ruetzleri | HCT-116 colorectal cancer cells | caspase activation, ↑mitochondrial trans-membrane potential | Colon cancer [42] |
| Manzamine A | Haliclona sp., Xestospongia sp., Pellina sp. | HCT116 cells | ↓BCL2, Δψm loss, ↑caspase activation, CytC release, | Colon cancer [44] |
| Aplysinopsins | Thorecta sp. | K562 cells | ↓BCL2, Δψm loss | Leukemia [45] |
| Irciniastatin A | Ircinia ramose sp. | Jurkat cells | ↑ROS, ↑JNK, ↑p38, apoptosis | Leukemia [46] |
| 10-acetylirciformonin B | Ircinia sp. | HL 60 cells | ↓BCL2, ↓Bcl-xL, ↑BAX, ↑ROS, CytC release, apoptosis | Leukemia [46] |
| Mycothiazole | Petrosaspongia mycofijiensis | T47D cells | ↓HIF-1 signaling, ↓mitochondrial function | Breast tumor [47] |
| Cyclo(-Pro-Tyr) | Callyspongia fistularis sp. | HepG2 cell | ↓BCL2, ↑BAX, ↑ROS, apoptosis | Hepatocellular carcinoma [48] |
| Urupocidin A | Monanchora pulchra sp. | PCa cells | Δψm loss, ↑ROS, CytC release, apoptosis | Prostate cancer [49] |
| 3,5-dibromo-2-(20,40-dibromophenoxy)-phenol | Dysidea sp. | PANC-1 | Complex II inhibition | Pancreatic carcinoma [50] |
| Santonin | Dysidea avara sp. | ALL B-lymphocytes | ↓Δψm, ↑ROS, CytC release, apoptosis | Acute lymphoblastic leukemia [51] |
| Papuamine | Haliclona sp. | MCF-7 | mitochondrial damage and JNK activation | Breast cancer [52] |
| 2-ethoxycarbonyl-2-β-hydroxy-A-nor-cholest-5-ene-4one | Acropora Formosa hexocoral, Dendronephthya sp. | A549 | ↓ TNF-α, ↓IL-8, ↓Bcl2, ↓MMP2, ↓MMP9, ↑ROS, ↑ BAX, ↑p21, CytC release | Lung cancer [53] |
| Methyl 5-[(1E,5E)-2,6-Dimethyl octa-1,5,7-trienyl] furan-3-carboxylate | Sinularia kavarittiensis coral | THP-1 | ↓Bcl-xL, ↑BAX, ↑ROS, ↓Δψm, CytC release, apoptosis | Leukemia [54] |
| Sinularin | Sinularia flexibilis coral | SK-HEP-1 | ↑ROS, ↓Δψm,↓OXPHOS, apoptosis | Liver cancer [55,56] |
| 11-dehydro-sinulariolide | Sinularia flexibilis coral | Ca9-22 | ∆Ψm loss, ↑caspase-3/-9 ↑Bax, ↓Bcl-2/Bcl-Xl, CytC release, apoptosis | Melanoma [55] |
| Aplidin | Aplidium albicans | MOLT-4, NIH3T3 | ↑ROS, ↓Δψm, ↓ATP, apoptosis | Leukemia, Lymphoma [57,58] |
| n-hexane, diethyl ether and methanolic extracts | Phallusia nigra | Isolated mitochondria from skin tissue of melanoma induced albino/Wistar rats | mitochondrial swelling, ↑ROS, ↓Δψm, CytC release, apoptosis | Melanoma [59] |
| Mandelalide A | Lissoclinum ascidian | NCI-H460, Neuro-2A, HeLa cells | complex V inhibition, apoptosis | Lung cancer, Neuroblastoma [60] |
| Mandelalide E | Lissoclinum ascidian | NCI-H460, HeLa, U87-MG, HCT116 | apoptosis | Lung cancer, Glioblastoma [61] |
| CS5931 | Ciona savignyi | HCT-8 | ↑caspase-3, ↑caspase-9, ↑Bax, ↓Δψm, CytC release, apoptosis | Colon cancer [62] |
| Echinoside A and ds-echinoside A | Pearsonothuria graeffei | HepG2, mice | apoptosis | Hepatocarcinoma [63] |
| Stichoposide C | Thelenota anax | HL-60, K562, THP-1, NB4, SNU-C4, HT-29, CT-26; mouse CT-26 subcutaneous tumor and HL-60 leukemia xenograft models | ↑Fas, ↑caspase-3, ↑caspase-8, cleavage of Bid, mitochondrial damage, apoptosis | Leukemia, Colorectal cancer [64] |
| Methanolic extracts | Holoturia parva, Haliclona oculate sp. | Mitochondria isolated from a rat model of hepatocellular carcinoma | ↑ROS, ↓Δψm, CytC release, ↑caspase-3, apoptosis | Hepatocellular carcinoma [65] |
| Lamellarin D | Lamellaria | p388 | ↓Bcl-2, ↓Δψm, ↑caspase-3, ↑caspase-9, apoptosis | Leukemia [66,67] |
| Extract fraction of T. coronatus | Turbo coronatus | EOC cells | ↑ROS, ↓Δψm, CytC release, mitochondrial swelling, apoptosis and necrosis | Epithelial ovarian cancer [68] |
| Conotoxins | Conus textile | U87MG | ↑ROS, ↓Δψm, CytC release, ↑caspase-3, ↑caspase-9, ↑Bax/Bcl-2 | Glioma [69] |
| Polysaccharide in fraction 2.1 | Donax variabilis | A549 | Mitochondrial disfunction, ↓Δψm, CytC release, ↑caspase-3, ↑caspase-9, ↑Bax/Bcl-2, apoptosis | Lung cancer cells [70] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirra, S.; Marfany, G. From Beach to the Bedside: Harnessing Mitochondrial Function in Human Diseases Using New Marine-Derived Strategies. Int. J. Mol. Sci. 2024, 25, 834. https://doi.org/10.3390/ijms25020834
Mirra S, Marfany G. From Beach to the Bedside: Harnessing Mitochondrial Function in Human Diseases Using New Marine-Derived Strategies. International Journal of Molecular Sciences. 2024; 25(2):834. https://doi.org/10.3390/ijms25020834
Chicago/Turabian StyleMirra, Serena, and Gemma Marfany. 2024. "From Beach to the Bedside: Harnessing Mitochondrial Function in Human Diseases Using New Marine-Derived Strategies" International Journal of Molecular Sciences 25, no. 2: 834. https://doi.org/10.3390/ijms25020834
APA StyleMirra, S., & Marfany, G. (2024). From Beach to the Bedside: Harnessing Mitochondrial Function in Human Diseases Using New Marine-Derived Strategies. International Journal of Molecular Sciences, 25(2), 834. https://doi.org/10.3390/ijms25020834