
amer1 Regulates Zebrafish Craniofacial Development by Interacting with the Wnt/β-Catenin Pathway
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
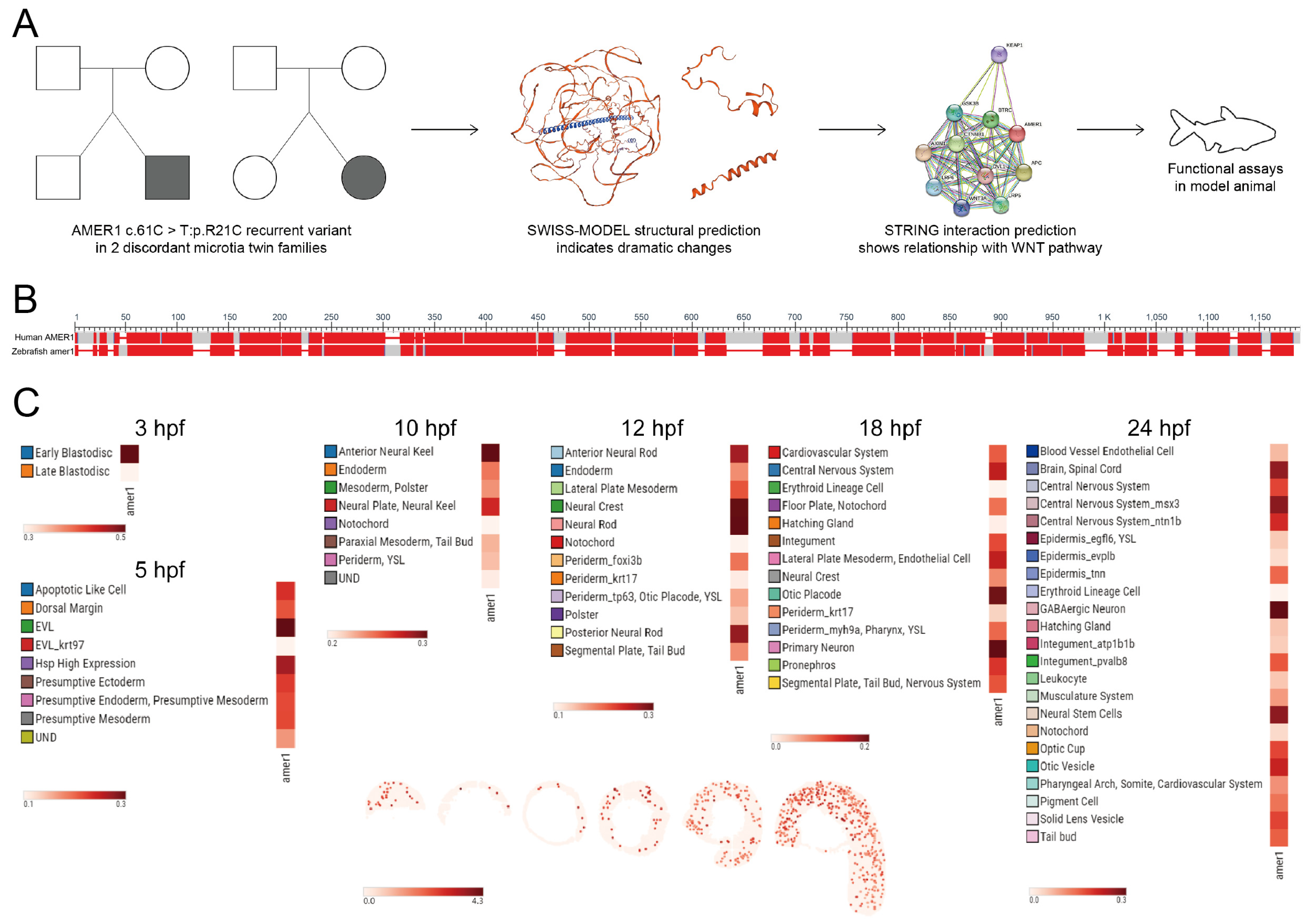
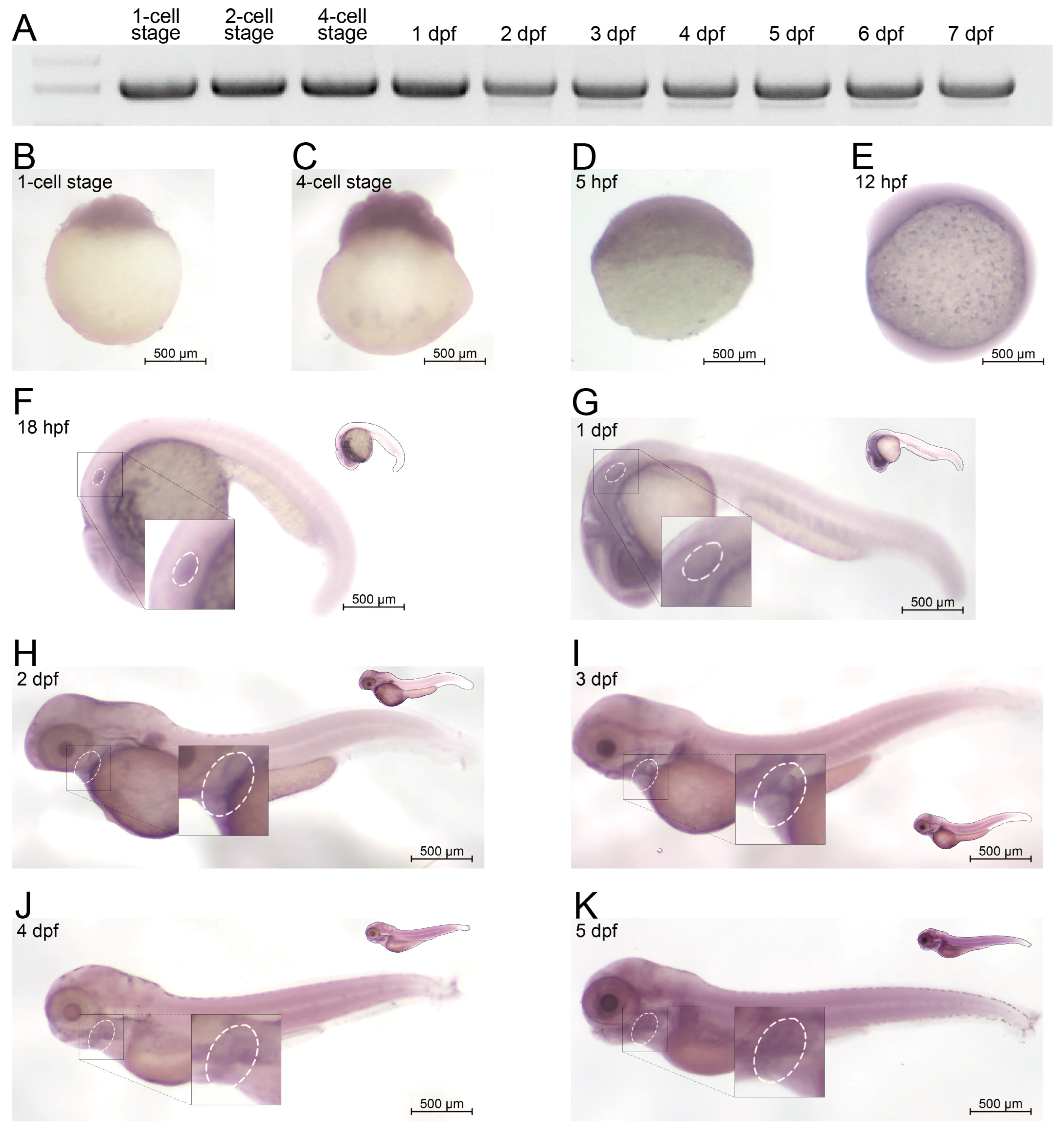
2.1. Homology and Expression Profile of AMER1 in Zebrafish
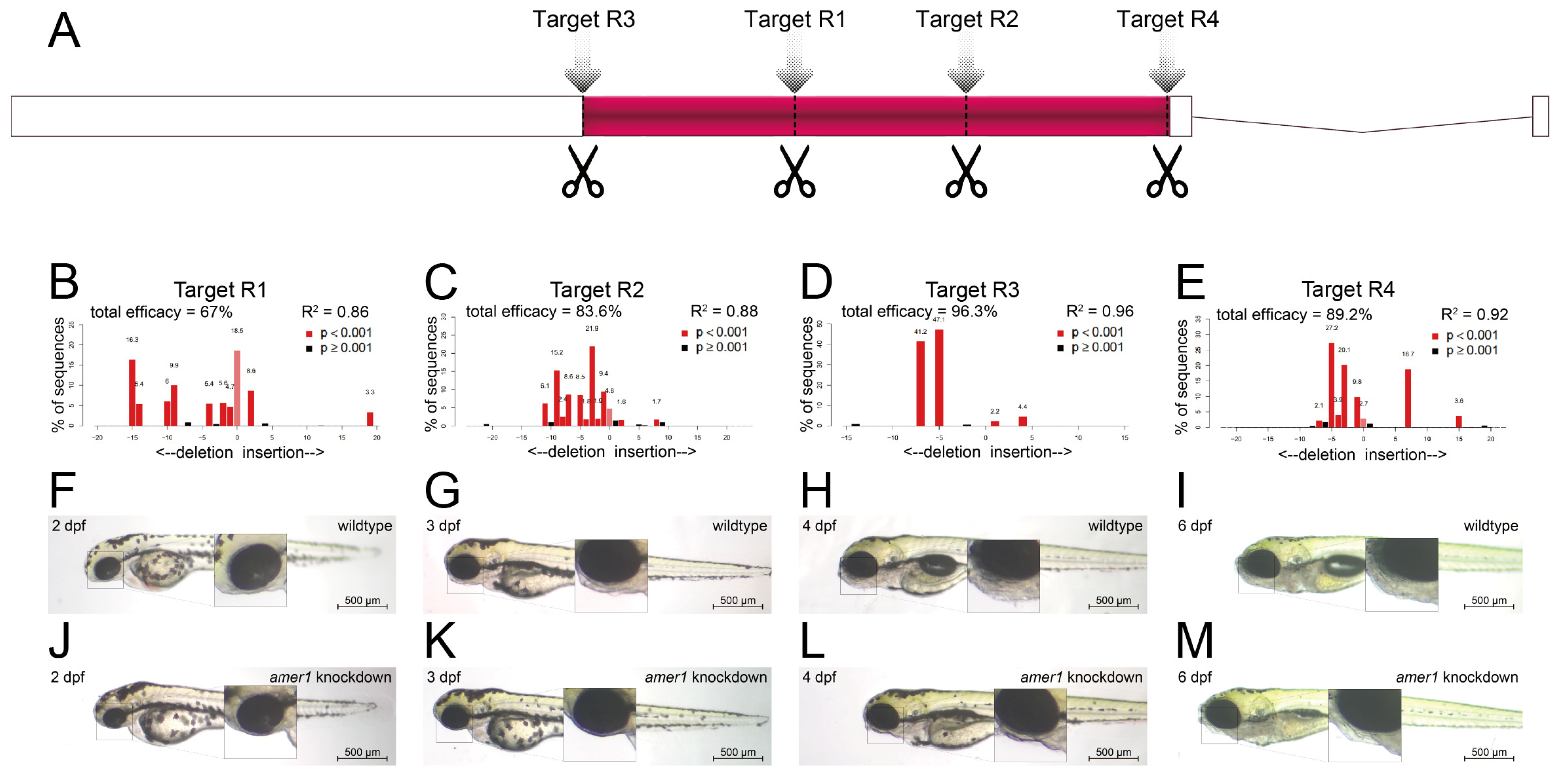
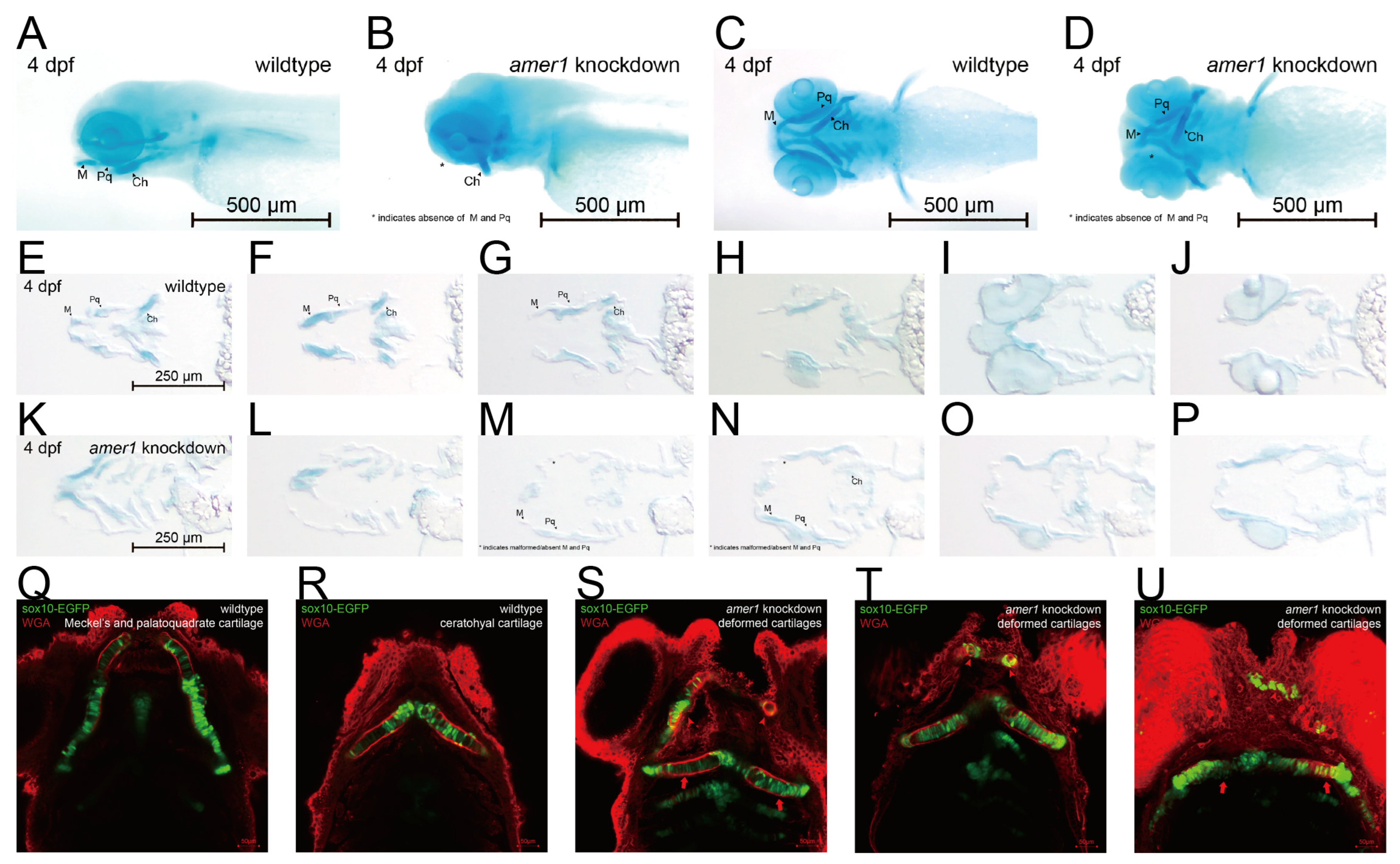
2.2. Knockdown of amer1 Results in Severe Pharyngeal Malformation
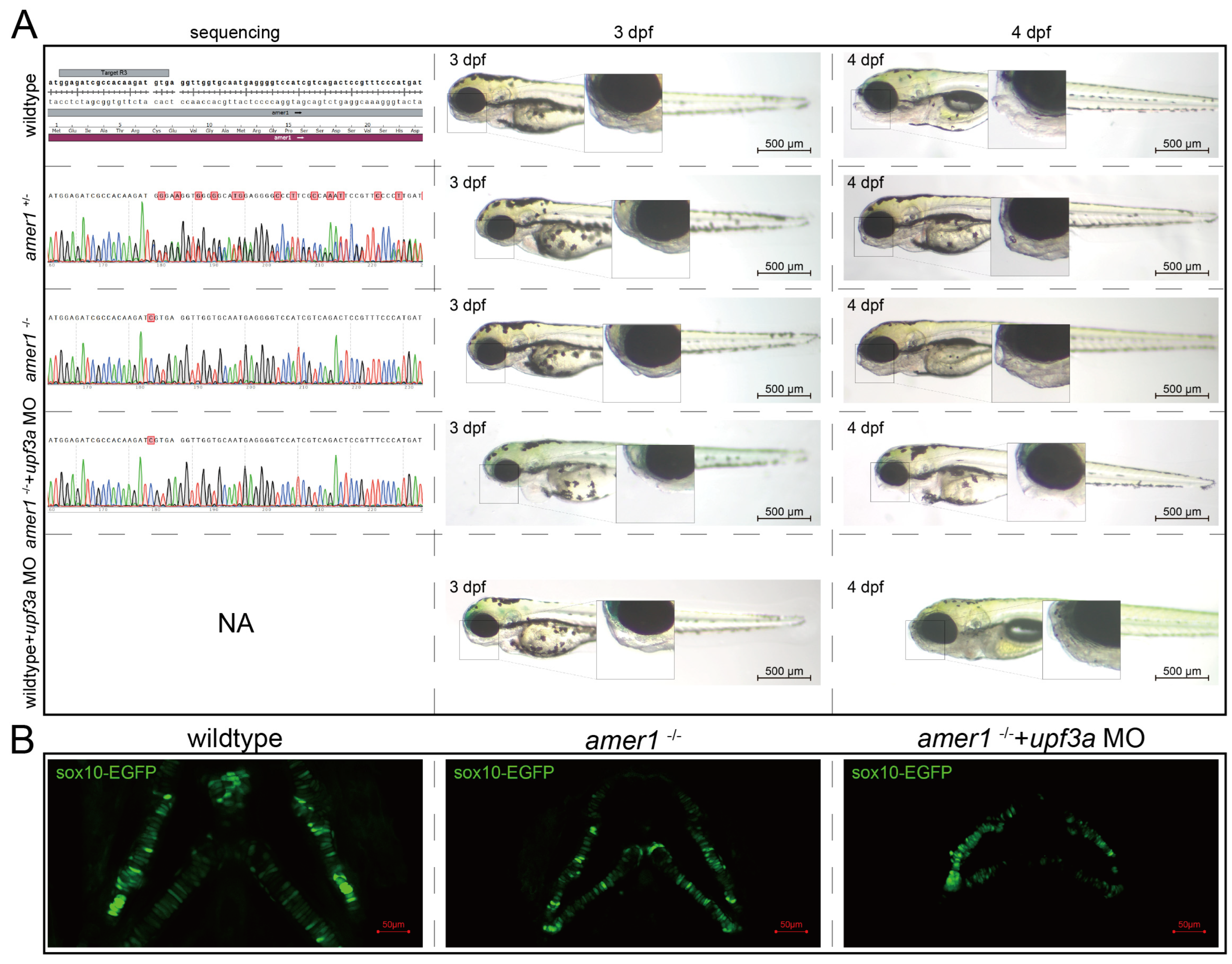
2.3. Inbred Homozygous Zebrafish Demonstrates Similar Phenotype after Excluding Potential Compensation
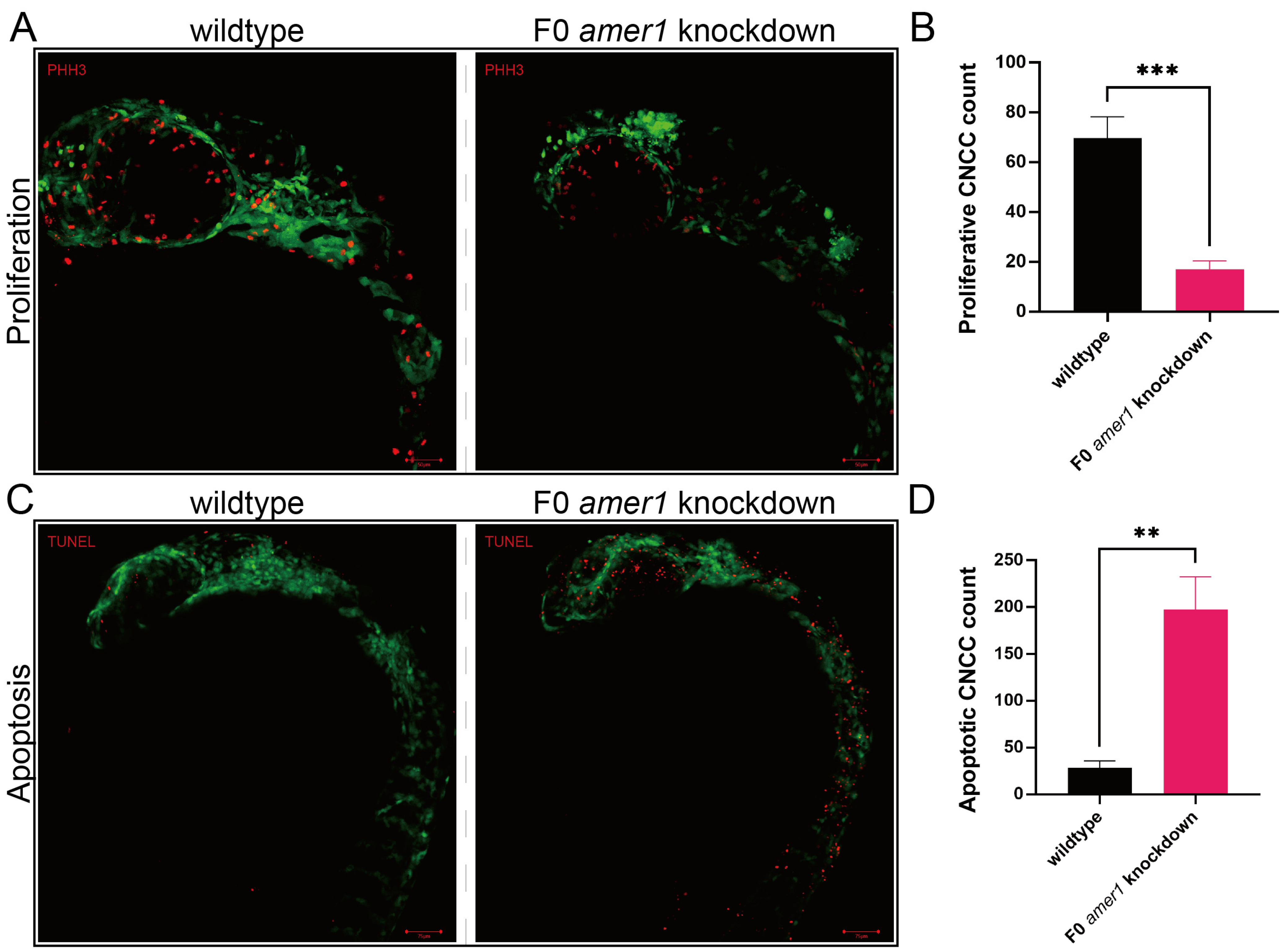
2.4. Knockdown of amer1 Affects Chondrocyte Differentiation, Proliferation, and Apoptosis
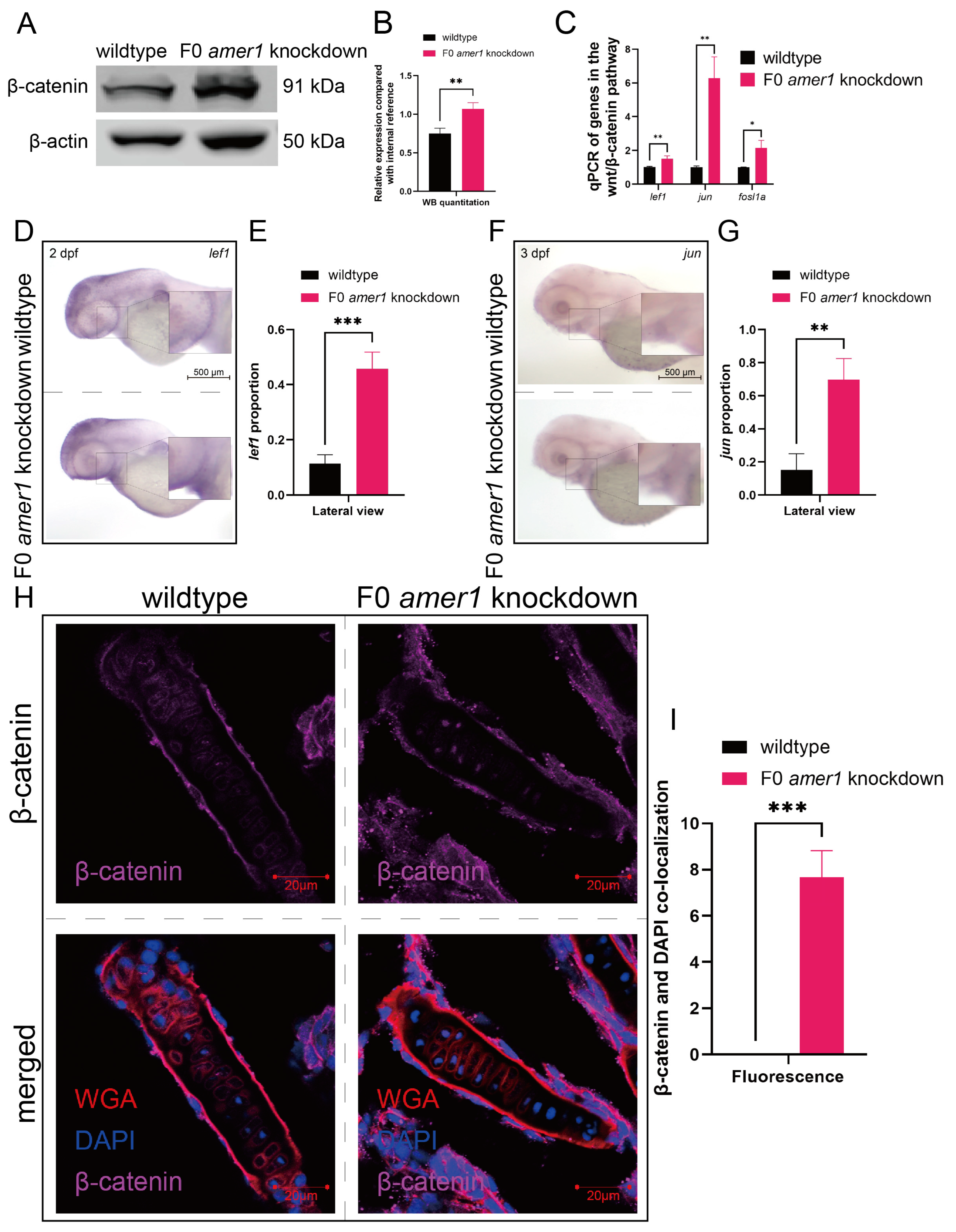
2.5. Amer1 Exerts Its Effects via the Wnt/β-Catenin Signaling Pathway
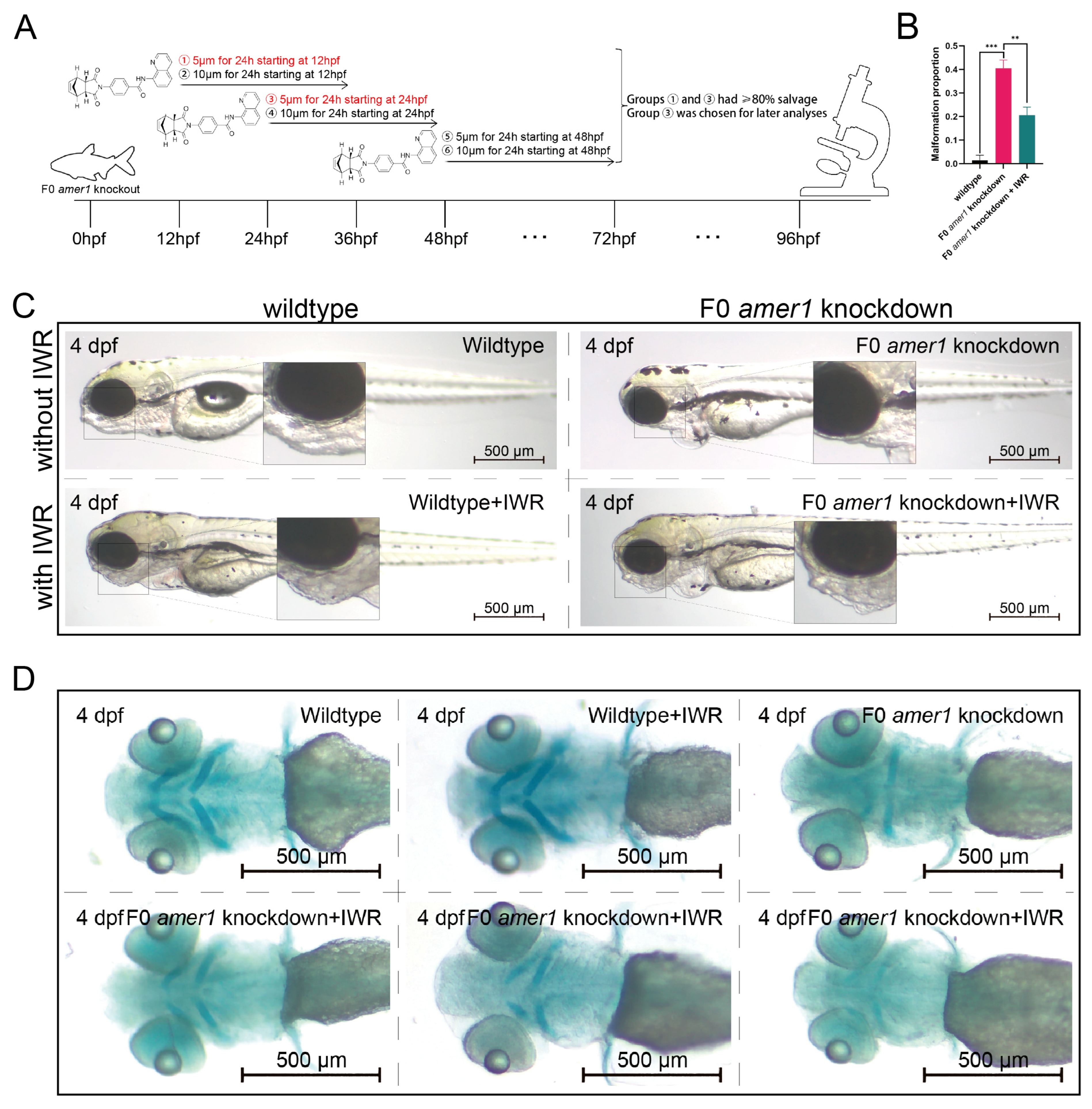
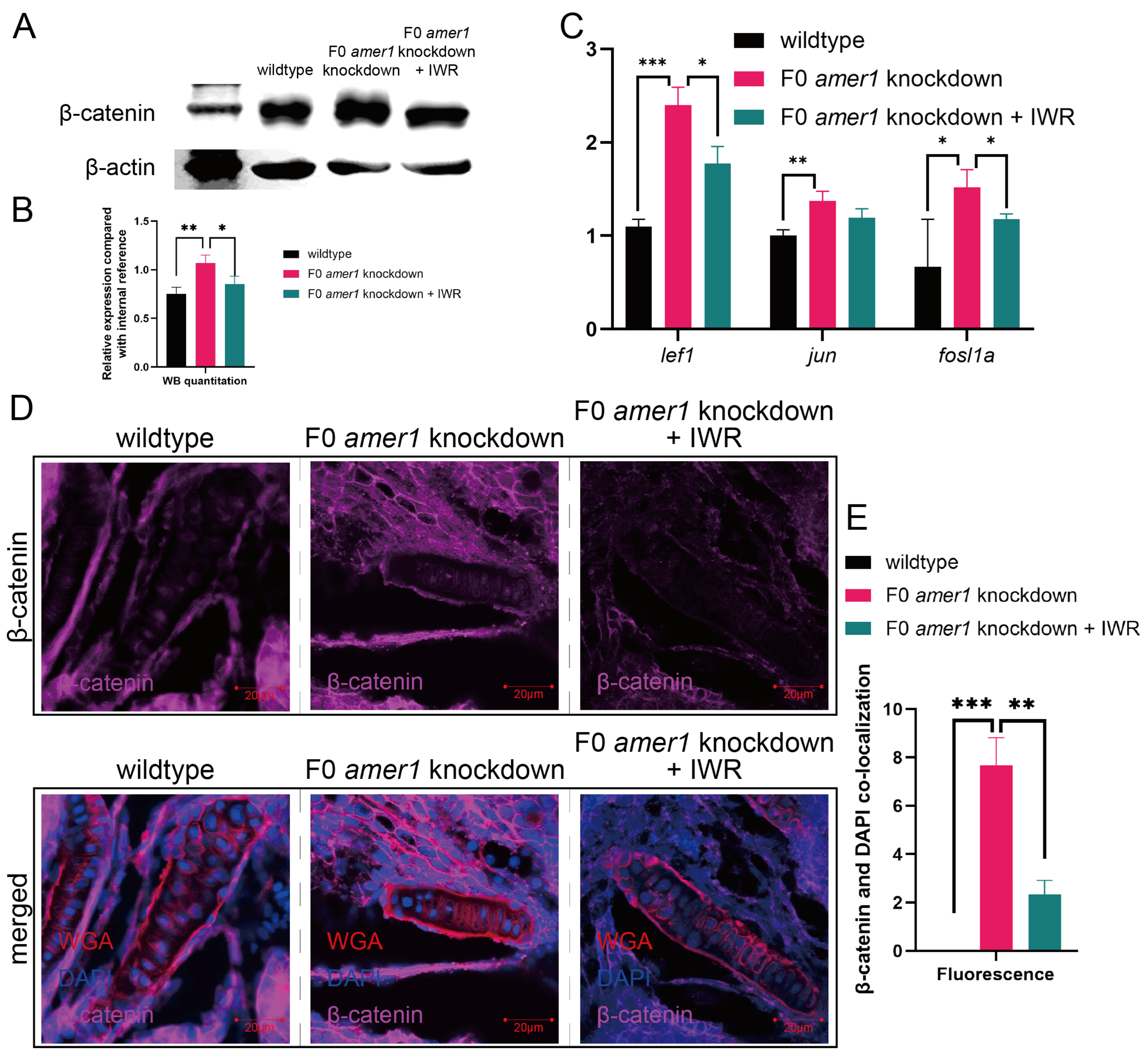
2.6. An Inhibitor of the Wnt/β-Catenin Signaling Pathway Rescues the Malformation of amer1 Knockdown Embryos
3. Discussion
4. Materials and Methods
4.1. Zebrafish and Embryos
4.2. Photography and Image Processing
4.3. Bioinformatic Prediction of amer1 Expression Profile in Zebrafish
4.4. Whole-Mount In Situ Hybridization
4.5. Genetic Manipulation of amer1
4.6. Cartilage Staining and Immunofluorescence Staining
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deng, K.; Dai, L.; Yi, L.; Deng, C.; Li, X.; Zhu, J. Epidemiologic characteristics and time trend in the prevalence of anotia and microtia in China. Birth Defects Res. A Clin. Mol. Teratol. 2016, 106, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Cabrejo, R.; Persing, J.; Alperovich, M. Epidemiologic Assessment of Microtia in Over 23 Million Consecutive United States Births. J. Craniofacial Surg. 2019, 30, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Bartel-Friedrich, S. Congenital Auricular Malformations: Description of Anomalies and Syndromes. Facial Plast. Surg. 2015, 31, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Ping, L.; Sun, H.; Chen, Y.; Wang, P.; Liu, T.; Jiang, R.; Zhang, X.; Chen, X. Whole-Exome Sequencing of Discordant Monozygotic Twin Families for Identification of Candidate Genes for Microtia-Atresia. Front. Genet. 2020, 11, 568052. [Google Scholar] [CrossRef] [PubMed]
- Arora, V.; Bijarnia-Mahay, S.; Saxena, K.K.; Suman, P.; Kukreja, S. Osteopathia Striata with Cranial Sclerosis: A Face-to-Radiograph-to-Gene Diagnosis. J. Pediatr. Genet. 2022, 11, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, Z.A.; van Kogelenberg, M.; Morgan, T.; Jeffs, A.; Fukuzawa, R.; Pearl, E.; Thaller, C.; Hing, A.V.; Porteous, M.E.; Garcia-Miñaur, S.; et al. Germline mutations in WTX cause a sclerosing skeletal dysplasia but do not predispose to tumorigenesis. Nat. Genet. 2009, 41, 95–100. [Google Scholar] [CrossRef]
- Li, J.; Rodriguez, G.; Han, X.; Janečková, E.; Kahng, S.; Song, B.; Chai, Y. Regulatory Mechanisms of Soft Palate Development and Malformations. J. Dent. Res. 2019, 98, 959–967. [Google Scholar] [CrossRef]
- Yin, X.; Li, J.; Salmon, B.; Huang, L.; Lim, W.H.; Liu, B.; Hunter, D.J.; Ransom, R.C.; Singh, G.; Gillette, M.; et al. Wnt Signaling and Its Contribution to Craniofacial Tissue Homeostasis. J. Dent. Res. 2015, 94, 1487–1494. [Google Scholar] [CrossRef]
- Sisson, B.E.; Dale, R.M.; Mui, S.R.; Topczewska, J.M.; Topczewski, J. A role of glypican4 and wnt5b in chondrocyte stacking underlying craniofacial cartilage morphogenesis. Mech. Dev. 2015, 138 Pt 3, 279–290. [Google Scholar] [CrossRef]
- Boutet, A.; Comai, G.; Schedl, A. The WTX/AMER1 gene family: Evolution, signature and function. BMC Evol. Biol. 2010, 10, 280. [Google Scholar] [CrossRef]
- Regimbald-Dumas, Y.; He, X. Wnt signalling: What The X@# is WTX? EMBO J. 2011, 30, 1415–1417. [Google Scholar] [PubMed]
- Fukuzawa, R.; Anaka, M.R.; Weeks, R.J.; Morison, I.M.; Reeve, A.E. Canonical WNT signalling determines lineage specificity in Wilms tumour. Oncogene 2009, 28, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, L.; Liu, L.; Wang, S.; Yang, Y.; Ding, Y.; Jeevan, D.; Li, X.; Ding, Y. LMP1 antagonizes WNT/β-catenin signalling through inhibition of WTX and promotes nasopharyngeal dysplasia but not tumourigenesis in LMP1(B95-8) transgenic mice. J. Pathol. 2011, 223, 574–583. [Google Scholar]
- Große, A.; Perner, B.; Naumann, U.; Englert, C. Zebrafish Wtx is a negative regulator of Wnt signaling but is dispensable for embryonic development and organ homeostasis. Dev. Dyn. 2019, 248, 866–881. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.S.; Lam, I.I.; Clay, H.; Duong, D.N.; Deo, R.C.; Coughlin, S.R. A Rapid Method for Directed Gene Knockout for Screening in G0 Zebrafish. Dev. Cell 2018, 46, 112–125.e4. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhu, P.; Shi, H.; Guo, L.; Zhang, Q.; Chen, Y.; Chen, S.; Zhang, Z.; Peng, J.; Chen, J. PTC-bearing mRNA elicits a genetic compensation response via Upf3a and COMPASS components. Nature 2019, 568, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Moisan, A.; Rivera, M.N.; Lotinun, S.; Akhavanfard, S.; Coffman, E.J.; Cook, E.B.; Stoykova, S.; Mukherjee, S.; Schoonmaker, J.A.; Burger, A.; et al. The WTX tumor suppressor regulates mesenchymal progenitor cell fate specification. Dev. Cell 2011, 20, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Major, M.B.; Camp, N.D.; Berndt, J.D.; Yi, X.; Goldenberg, S.J.; Hubbert, C.; Biechele, T.L.; Gingras, A.C.; Zheng, N.; Maccoss, M.J.; et al. Wilms tumor suppressor WTX negatively regulates WNT/beta-catenin signaling. Science 2007, 316, 1043–1046. [Google Scholar] [CrossRef]
- Chen, Q.; Zhao, Y.; Shen, G.; Dai, J. Etiology and Pathogenesis of Hemifacial Microsomia. J. Dent. Res. 2018, 97, 1297–1305. [Google Scholar] [CrossRef]
- Lefebvre, V.; de Crombrugghe, B. Toward understanding SOX9 function in chondrocyte differentiation. Matrix Biol. 1998, 16, 529–540. [Google Scholar] [CrossRef]
- Zhao, Q.; Eberspaecher, H.; Lefebvre, V.; De Crombrugghe, B. Parallel expression of Sox9 and Col2a1 in cells undergoing chondrogenesis. Dev. Dyn. 1997, 209, 377–386. [Google Scholar] [CrossRef]
- Tanneberger, K.; Pfister, A.S.; Kriz, V.; Bryja, V.; Schambony, A.; Behrens, J. Structural and functional characterization of the Wnt inhibitor APC membrane recruitment 1 (Amer1). J. Biol. Chem. 2011, 286, 19204–19214. [Google Scholar] [CrossRef] [PubMed]
- Tanneberger, K.; Pfister, A.S.; Brauburger, K.; Schneikert, J.; Hadjihannas, M.V.; Kriz, V.; Schulte, G.; Bryja, V.; Behrens, J. Amer1/WTX couples Wnt-induced formation of PtdIns(4,5)P2 to LRP6 phosphorylation. EMBO J. 2011, 30, 1433–1443. [Google Scholar] [CrossRef] [PubMed]
- Hans, S.; Westerfield, M. Changes in retinoic acid signaling alter otic patterning. Development 2007, 134, 2449–2458. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Mao, A.; Wang, P.; Ning, G.; Cao, Y.; Wang, Q. Endodermal pouch-expressed dmrt2b is important for pharyngeal cartilage formation. Biol. Open 2018, 7, bio035444. [Google Scholar]
- Liu, C.; Li, R.; Li, Y.; Lin, X.; Zhao, K.; Liu, Q.; Wang, S.; Yang, X.; Shi, X.; Ma, Y.; et al. Spatiotemporal mapping of gene expression landscapes and developmental trajectories during zebrafish embryogenesis. Dev. Cell 2022, 57, 1284–1298.e5. [Google Scholar] [CrossRef]
- Mm, W. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish Danio (Brachydanio) Rerio; University of Oregon Press: Eugene, Oregon, 1994. [Google Scholar]
- Montague, T.G.; Cruz, J.M.; Gagnon, J.A.; Church, G.M.; Valen, E. CHOPCHOP: A CRISPR/Cas9 and TALEN web tool for genome editing. Nucleic Acids Res. 2014, 42, W401–W407. [Google Scholar] [CrossRef]
- Chang, N.; Sun, C.; Gao, L.; Zhu, D.; Xu, X.; Zhu, X.; Xiong, J.W.; Xi, J.J. Genome editing with RNA-guided Cas9 nuclease in zebrafish embryos. Cell Res. 2013, 23, 465–472. [Google Scholar] [CrossRef]
- Shum, E.Y.; Jones, S.H.; Shao, A.; Dumdie, J.; Krause, M.D.; Chan, W.K.; Lou, C.H.; Espinoza, J.L.; Song, H.W.; Phan, M.H.; et al. The Antagonistic Gene Paralogs Upf3a and Upf3b Govern Nonsense-Mediated RNA Decay. Cell 2016, 165, 382–395. [Google Scholar] [CrossRef]
- Wang, Y.; Ping, L.; Luan, X.; Chen, Y.; Fan, X.; Li, L.; Liu, Y.; Wang, P.; Zhang, S.; Zhang, B.; et al. A Mutation in VWA1, Encoding von Willebrand Factor A Domain-Containing Protein 1, Is Associated With Hemifacial Microsomia. Front. Cell Dev. Biol. 2020, 8, 571004. [Google Scholar] [CrossRef] [PubMed]
- Dale, R.M.; Topczewski, J. Identification of an evolutionarily conserved regulatory element of the zebrafish col2a1a gene. Dev. Biol. 2011, 357, 518–531. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Ping, L.; Fan, X.; Fan, Y.; Zhang, B.; Chen, X. amer1 Regulates Zebrafish Craniofacial Development by Interacting with the Wnt/β-Catenin Pathway. Int. J. Mol. Sci. 2024, 25, 734. https://doi.org/10.3390/ijms25020734
Sun L, Ping L, Fan X, Fan Y, Zhang B, Chen X. amer1 Regulates Zebrafish Craniofacial Development by Interacting with the Wnt/β-Catenin Pathway. International Journal of Molecular Sciences. 2024; 25(2):734. https://doi.org/10.3390/ijms25020734
Chicago/Turabian StyleSun, Le, Lu Ping, Xinmiao Fan, Yue Fan, Bo Zhang, and Xiaowei Chen. 2024. "amer1 Regulates Zebrafish Craniofacial Development by Interacting with the Wnt/β-Catenin Pathway" International Journal of Molecular Sciences 25, no. 2: 734. https://doi.org/10.3390/ijms25020734
APA StyleSun, L., Ping, L., Fan, X., Fan, Y., Zhang, B., & Chen, X. (2024). amer1 Regulates Zebrafish Craniofacial Development by Interacting with the Wnt/β-Catenin Pathway. International Journal of Molecular Sciences, 25(2), 734. https://doi.org/10.3390/ijms25020734