
Exosomal microRNA-92b Is a Diagnostic Biomarker in Breast Cancer and Targets Survival-Related MTSS1L to Promote Tumorigenesis

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
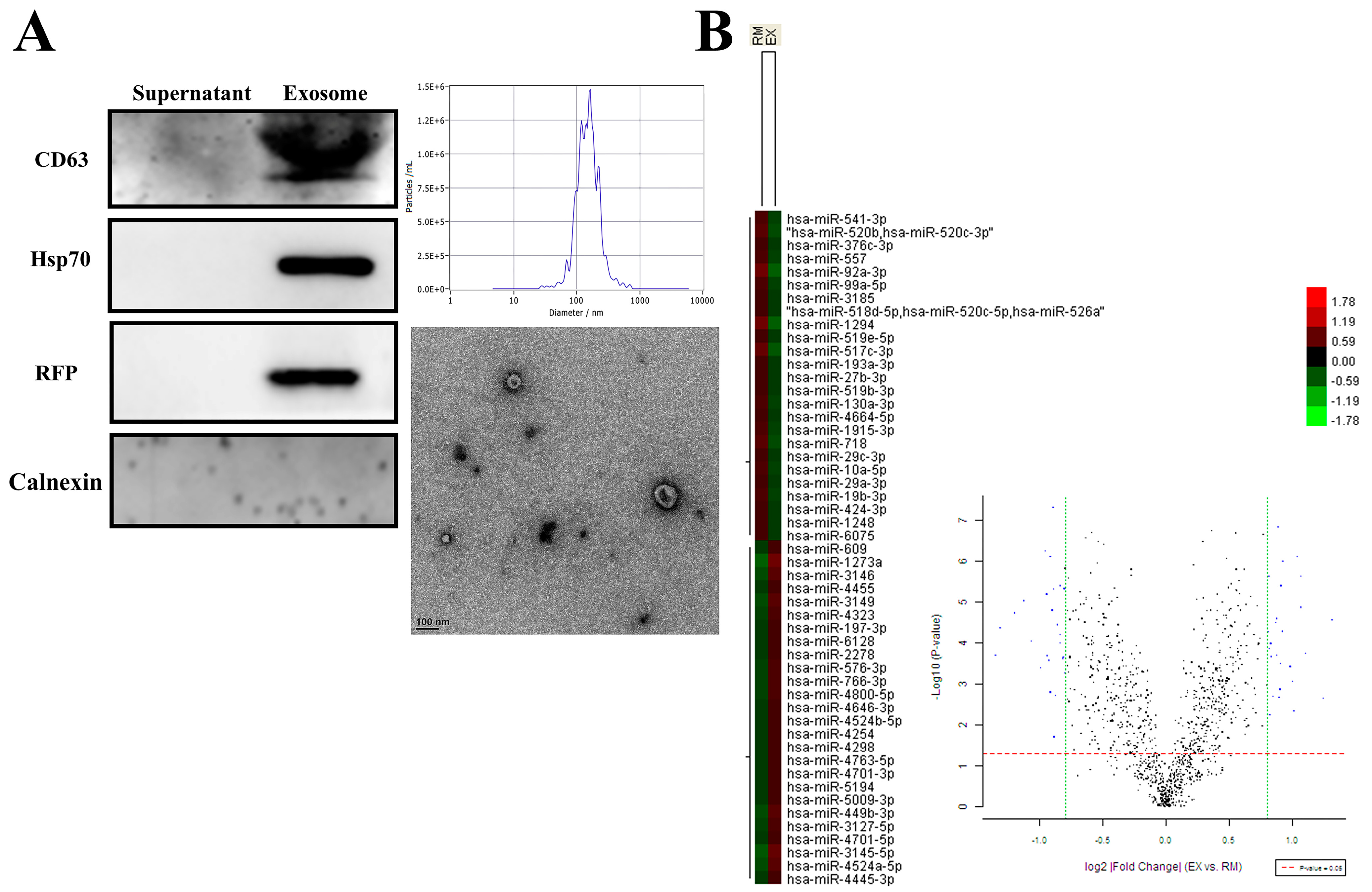
2.1. Characterization of Isolated Exosomes and Expression Profile of Exosomal miRNA
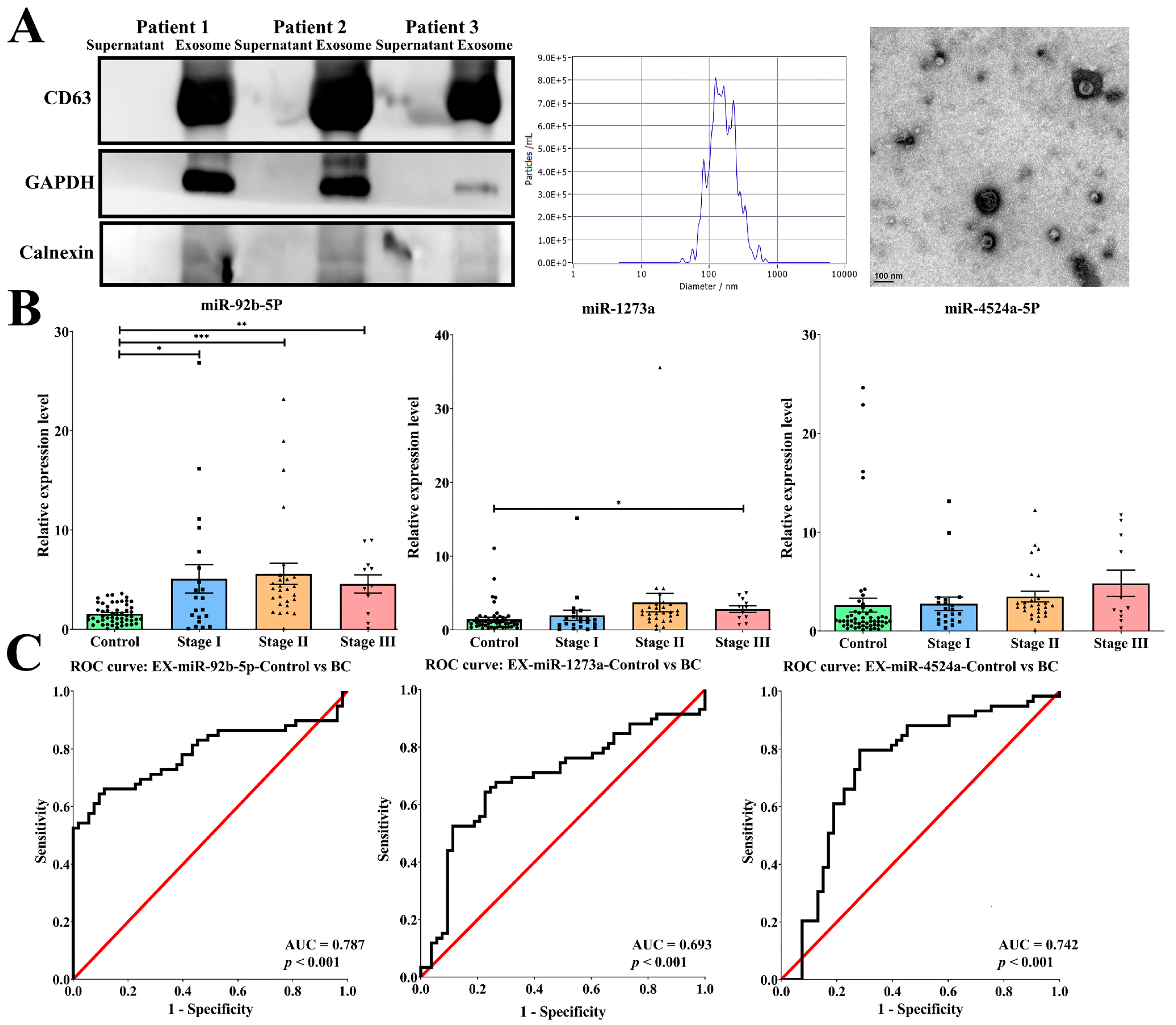
2.2. Exosomal miR-92b-5p Was Significantly Increased in a Different Stage of Breast Cancer Patients
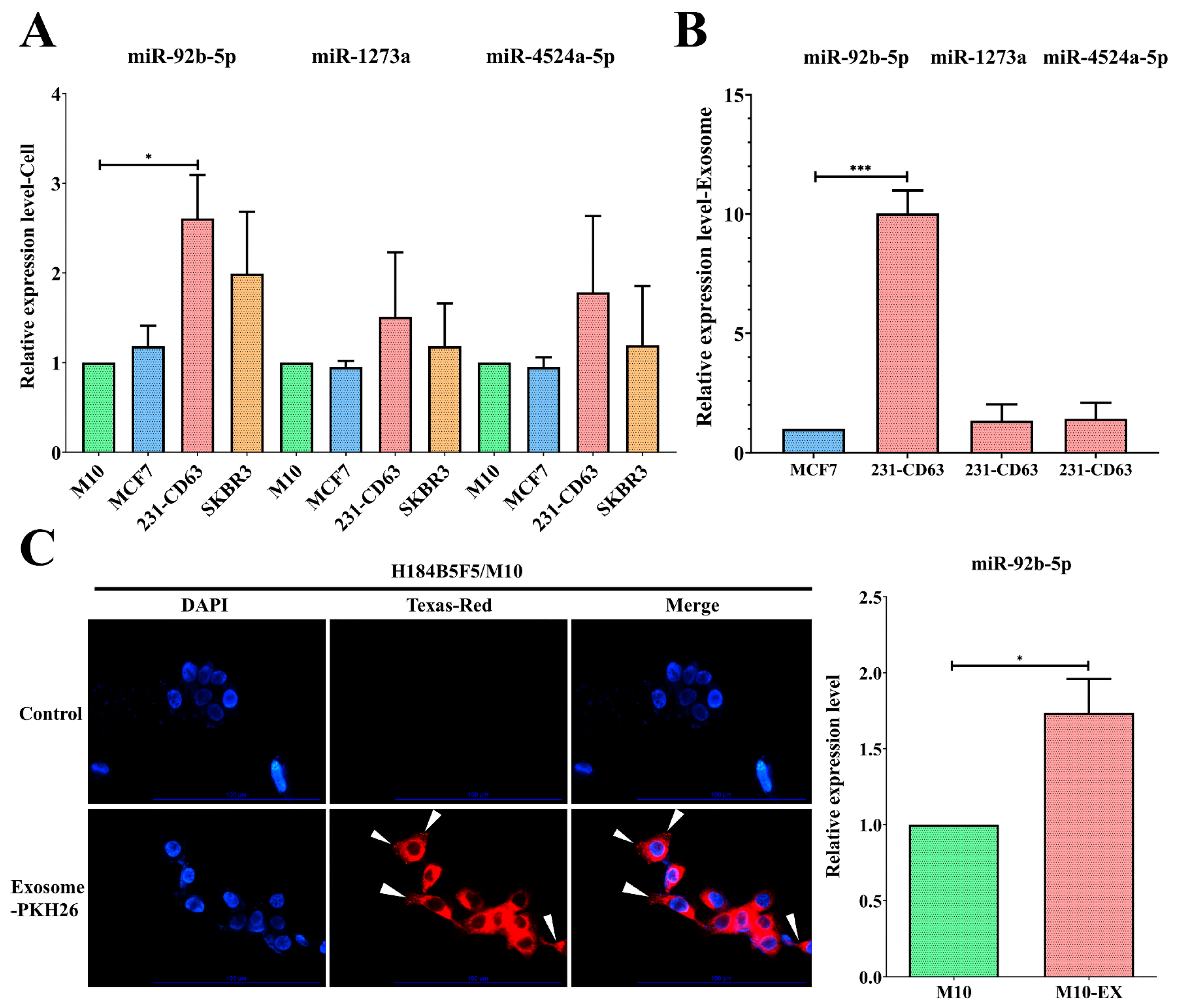
2.3. miR-92b-5p Was Upregulated in Both Cell Lines and Exosomes of 231-CD63
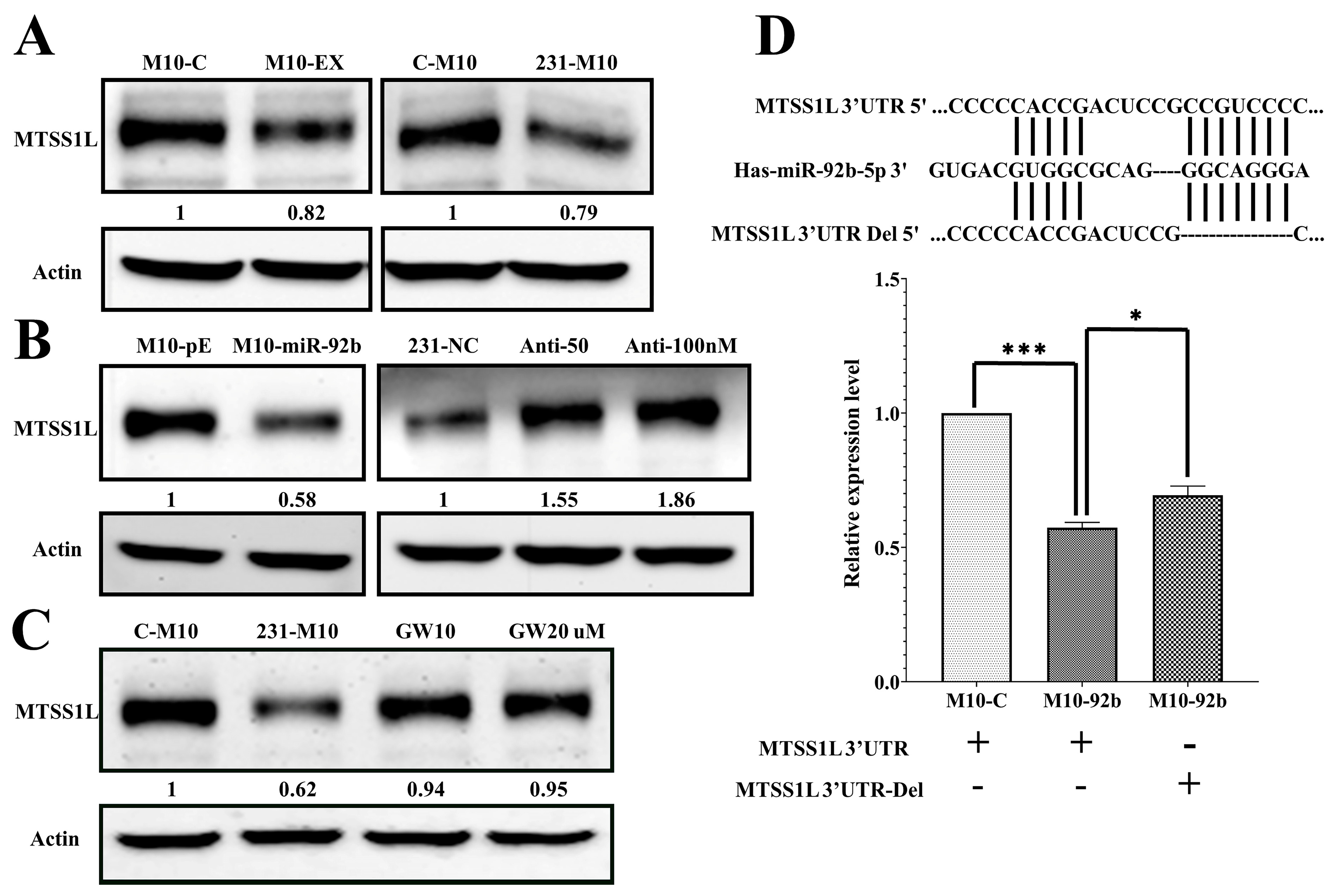
2.4. MTSS1L Was Targeted by miR-92b-5p
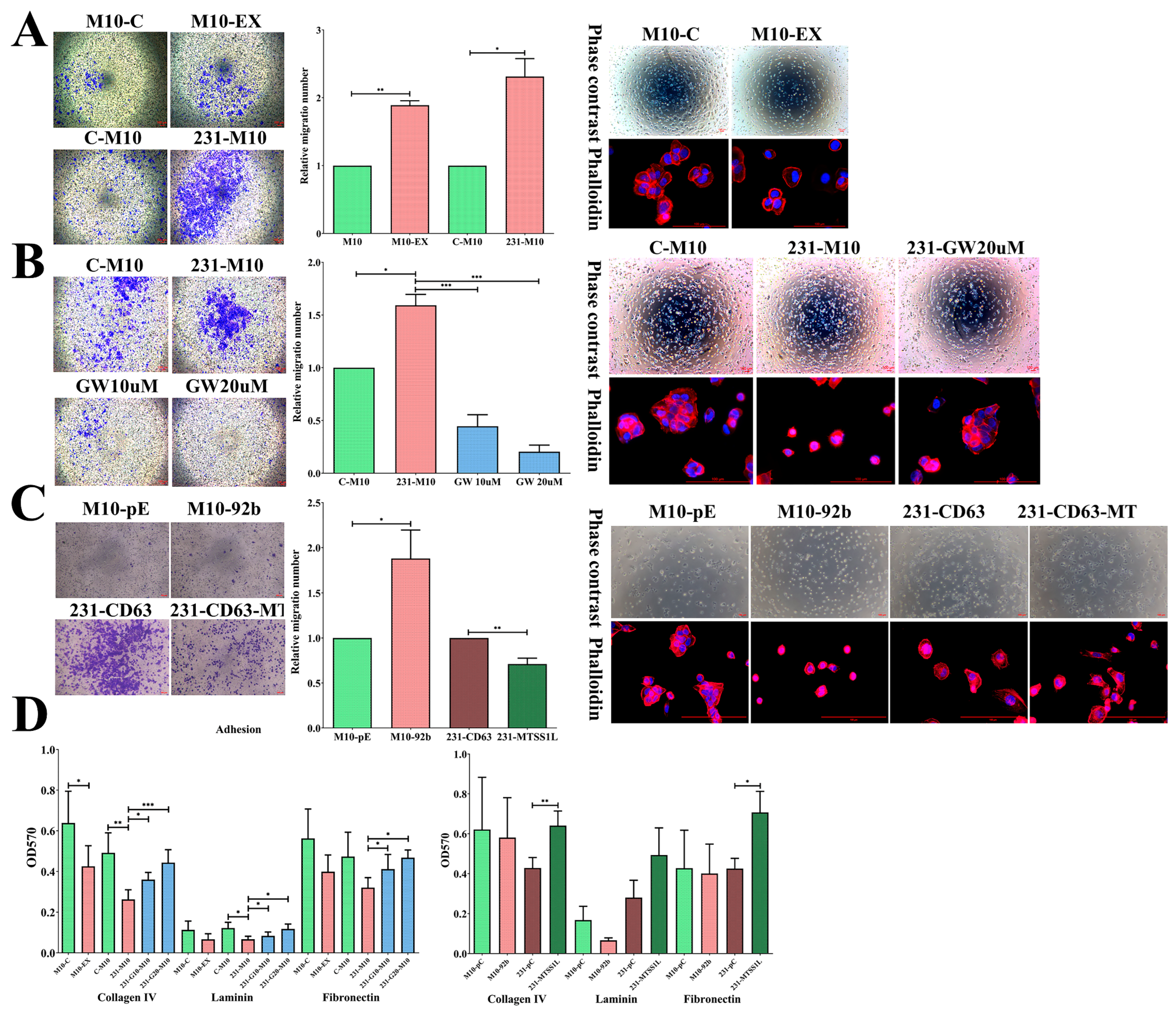
2.5. Exosomes Could Alter the Characteristics of the Recipient Cell
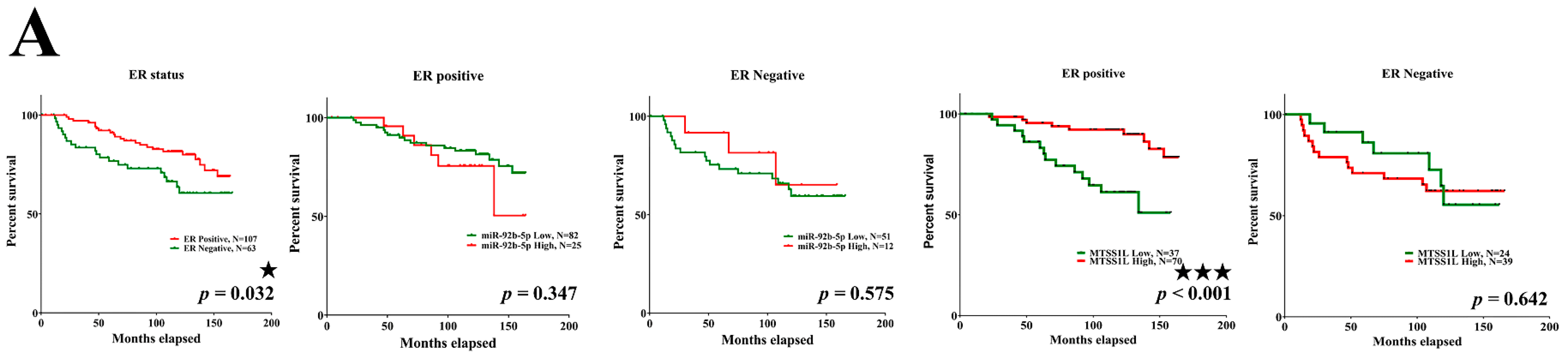
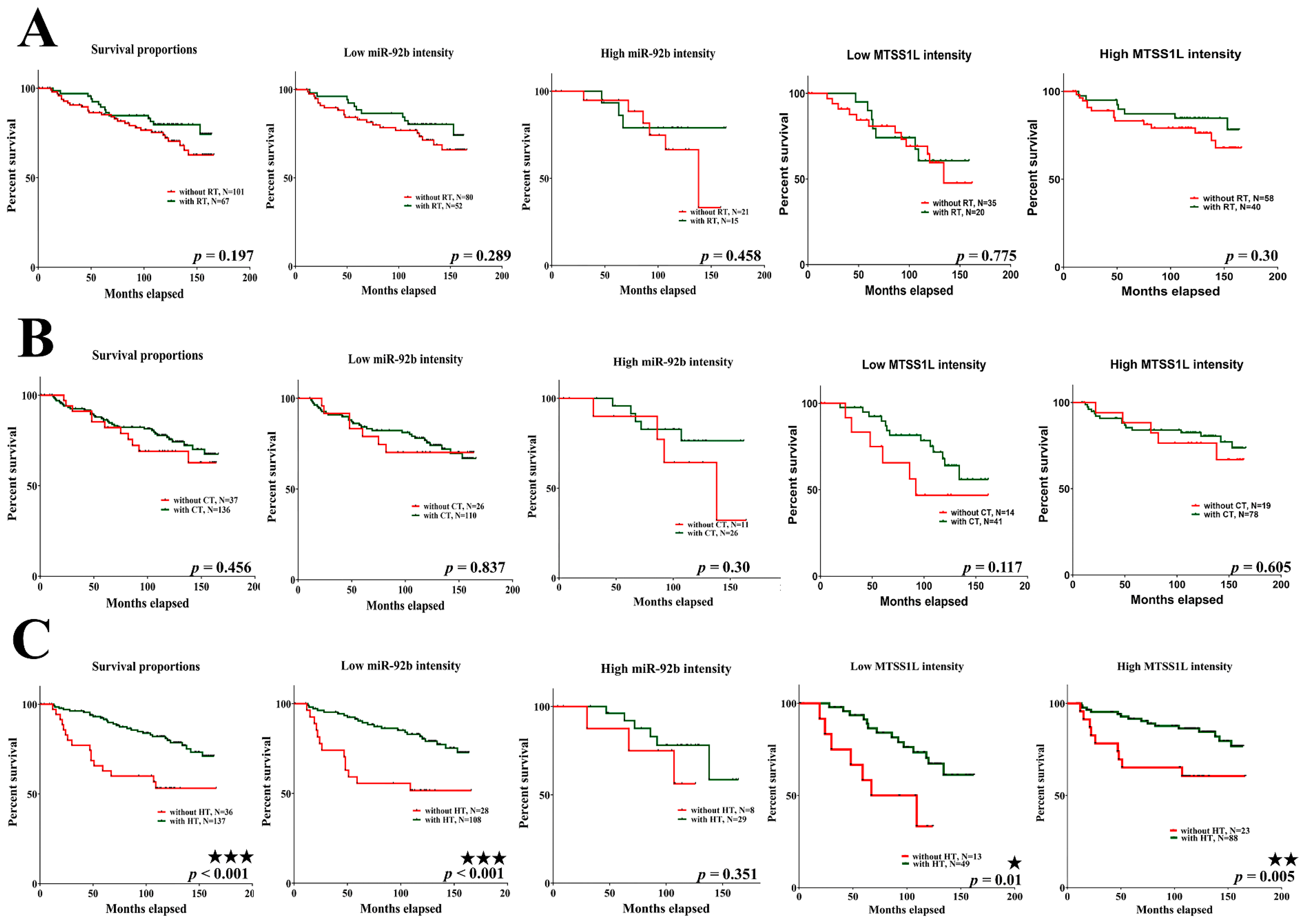
2.6. MTSS1L Expression Correlated with the Survival of Breast Cancer Patients
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. Study Population
4.3. Exosome Isolation and Labeling
4.4. Microarray Analysis
4.5. Western Blots
4.6. Exosome RNA Extraction and Real-Time PCR
4.7. Nanoparticle Tracking Analysis (NTA)
4.8. Transmission Electron Microscopy (TEM)
4.9. Co-Culture System
4.10. 3′-UTR Reporter and Dual-Luciferase Assay
4.11. Migration and Spreading Assay
4.12. Cell Adhesion Assay
4.13. In Situ Hybridization (ISH) and Immunohistochemical Staining (IHC)
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nordenskjold, A.E.; Fohlin, H.; Arnesson, L.G.; Einbeigi, Z.; Holmberg, E.; Albertsson, P.; Karlsson, P. Breast cancer survival trends in different stages and age groups–A population-based study 1989–2013. Acta Oncol. 2019, 58, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs–microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Garcia, I.; Miska, E.A. MicroRNA functions in animal development and human disease. Development 2005, 132, 4653–4662. [Google Scholar] [CrossRef]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed]
- Asiaf, A.; Ahmad, S.T.; Arjumand, W.; Zargar, M.A. MicroRNAs in Breast Cancer: Diagnostic and Therapeutic Potential. Methods Mol. Biol. 2018, 1699, 23–43. [Google Scholar]
- Fu, S.W.; Chen, L.; Man, Y.G. miRNA Biomarkers in Breast Cancer Detection and Management. J. Cancer 2011, 2, 116–122. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef]
- Eichelser, C.; Stuckrath, I.; Muller, V.; Milde-Langosch, K.; Wikman, H.; Pantel, K.; Schwarzenbach, H. Increased serum levels of circulating exosomal microRNA-373 in receptor-negative breast cancer patients. Oncotarget 2014, 5, 9650–9663. [Google Scholar] [CrossRef] [PubMed]
- Urbanelli, L.; Magini, A.; Buratta, S.; Brozzi, A.; Sagini, K.; Polchi, A.; Tancini, B.; Emiliani, C. Signaling pathways in exosomes biogenesis, secretion and fate. Genes 2013, 4, 152–170. [Google Scholar] [CrossRef] [PubMed]
- Cocucci, E.; Meldolesi, J. Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Joyce, D.P.; Kerin, M.J.; Dwyer, R.M. Exosome-encapsulated microRNAs as circulating biomarkers for breast cancer. Int. J. Cancer 2016, 139, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Schwille, P.; Brugger, B.; Simons, M. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef] [PubMed]
- Plantefaber, L.C.; Hynes, R.O. Changes in integrin receptors on oncogenically transformed cells. Cell 1989, 56, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, F.; Ding, H.; Wang, Y.; Li, P.; Wang, K. Emerging Function and Clinical Values of Exosomal MicroRNAs in Cancer. Mol. Ther. Nucleic Acids 2019, 16, 791–804. [Google Scholar] [CrossRef]
- He, Y.; Deng, F.; Yang, S.; Wang, D.; Chen, X.; Zhong, S.; Zhao, J.; Tang, J. Exosomal microRNA: A novel biomarker for breast cancer. Biomark. Med. 2018, 12, 177–188. [Google Scholar] [CrossRef]
- Sueta, A.; Yamamoto, Y.; Iwase, H. The role of exosomal microRNAs; focus on clinical applications in breast cancer. Cancer Drug Resist. 2019, 2, 847–861. [Google Scholar] [CrossRef]
- Rabinowits, G.; Gercel-Taylor, C.; Day, J.M.; Taylor, D.D.; Kloecker, G.H. Exosomal microRNA: A diagnostic marker for lung cancer. Clin. Lung Cancer 2009, 10, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Tavoosidana, G.; Ronquist, G.; Darmanis, S.; Yan, J.; Carlsson, L.; Wu, D.; Conze, T.; Ek, P.; Semjonow, A.; Eltze, E.; et al. Multiple recognition assay reveals prostasomes as promising plasma biomarkers for prostate cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 8809–8814. [Google Scholar] [CrossRef] [PubMed]
- Hannafon, B.N.; Trigoso, Y.D.; Calloway, C.L.; Zhao, Y.D.; Lum, D.H.; Welm, A.L.; Zhao, Z.J.; Blick, K.E.; Dooley, W.C.; Ding, W.Q. Plasma exosome microRNAs are indicative of breast cancer. Breast Cancer Res. 2016, 18, 90. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, Y.; Xia, T.; Zhou, X.; Huang, Z.; Zhang, H.; Zhu, W.; Ding, Q.; Wang, S. Circulating microRNAs from the miR-106a-363 cluster on chromosome X as novel diagnostic biomarkers for breast cancer. Breast Cancer Res. Treat. 2018, 170, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Zhang, Y.; Wang, R.; Zhou, W.; Zhao, Z.; Liang, H.; Qi, L.; Zhao, W.; Wang, H.; Wang, C.; et al. Identification of differentially expressed miRNAs in individual breast cancer patient and application in personalized medicine. Oncogenesis 2016, 5, e194. [Google Scholar] [CrossRef]
- Pena-Chilet, M.; Martinez, M.T.; Perez-Fidalgo, J.A.; Peiro-Chova, L.; Oltra, S.S.; Tormo, E.; Alonso-Yuste, E.; Martinez-Delgado, B.; Eroles, P.; Climent, J.; et al. MicroRNA profile in very young women with breast cancer. BMC Cancer 2014, 14, 529. [Google Scholar] [CrossRef]
- Wu, T.; Chen, Y.; Du, Y.; Tao, J.; Zhou, Z.; Yang, Z. Serum Exosomal MiR-92b-5p as a Potential Biomarker for Acute Heart Failure Caused by Dilated Cardiomyopathy. Cell Physiol. Biochem. 2018, 46, 1939–1950. [Google Scholar] [CrossRef]
- Wu, T.; Chen, Y.; Du, Y.; Tao, J.; Li, W.; Zhou, Z.; Yang, Z. Circulating exosomal miR-92b-5p is a promising diagnostic biomarker of heart failure with reduced ejection fraction patients hospitalized for acute heart failure. J. Thorac. Dis. 2018, 10, 6211–6220. [Google Scholar] [CrossRef]
- Peterlin, A.; Pocivavsek, K.; Petrovic, D.; Peterlin, B. The Role of microRNAs in Heart Failure: A Systematic Review. Front. Cardiovasc. Med. 2020, 7, 161. [Google Scholar] [CrossRef]
- Leidner, R.S.; Li, L.; Thompson, C.L. Dampening enthusiasm for circulating microRNA in breast cancer. PLoS ONE 2013, 8, e57841. [Google Scholar] [CrossRef]
- Mattila, P.K.; Pykalainen, A.; Saarikangas, J.; Paavilainen, V.O.; Vihinen, H.; Jokitalo, E.; Lappalainen, P. Missing-in-metastasis and IRSp53 deform PI(4,5)P2-rich membranes by an inverse BAR domain-like mechanism. J. Cell Biol. 2007, 176, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Machesky, L.M.; Johnston, S.A. MIM: A multifunctional scaffold protein. J. Mol. Med. 2007, 85, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Niu, S.; Yu, D.; Zhan, X.H.; Zeng, X.; Cui, B.; Chen, Y.; Yoon, J.; Martin, S.S.; Lu, X.; et al. Abba promotes PDGF-mediated membrane ruffling through activation of the small GTPase Rac1. Biochem. Biophys. Res. Commun. 2010, 401, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.C.; Luo, X.; Wang, S.X.; Zhan, X. Fibronectin-mediated cell spreading requires ABBA-Rac1 signaling. J. Cell Biochem. 2013, 114, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Chatzi, C.; Zhang, Y.; Hendricks, W.D.; Chen, Y.; Schnell, E.; Goodman, R.H.; Westbrook, G.L. Exercise-induced enhancement of synaptic function triggered by the inverse BAR protein, Mtss1L. eLife 2019, 8, e45920. [Google Scholar] [CrossRef]
- Aseervatham, J. Cytoskeletal Remodeling in Cancer. Biology 2020, 9, 385. [Google Scholar] [CrossRef]
- Padilla-Rodriguez, M.; Parker, S.S.; Adams, D.G.; Westerling, T.; Puleo, J.I.; Watson, A.W.; Hill, S.M.; Noon, M.; Gaudin, R.; Aaron, J.; et al. The actin cytoskeletal architecture of estrogen receptor positive breast cancer cells suppresses invasion. Nat. Commun. 2018, 9, 2980. [Google Scholar] [CrossRef]
- Sueta, A.; Yamamoto, Y.; Tomiguchi, M.; Takeshita, T.; Yamamoto-Ibusuki, M.; Iwase, H. Differential expression of exosomal miRNAs between breast cancer patients with and without recurrence. Oncotarget 2017, 8, 69934–69944. [Google Scholar] [CrossRef]
- Rabinowits, G.; Bowden, M.; Flores, L.M.; Verselis, S.; Vergara, V.; Jo, V.Y.; Chau, N.; Lorch, J.; Hammerman, P.S.; Thomas, T.; et al. Comparative Analysis of MicroRNA Expression among Benign and Malignant Tongue Tissue and Plasma of Patients with Tongue Cancer. Front. Oncol. 2017, 7, 191. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ct < 35 | Ct ≥ 35 | |||
|---|---|---|---|---|
| miRNA ID | miRNA ID | miRNA ID | miRNA ID | miRNA ID |
| miR-21 | miR-7 | miR-140 | miR-204-5p | miR-502-5p |
| miR-92a-3p | miR-10b | miR-141 | miR-204-3p | miR-503 |
| miR-92b-5p | miR-17 | miR-142-3p | miR-205 | miR-503-3p |
| miR-223-3p | miR-18a | miR-142-5p | miR-214 | miR-548d-3p |
| miR-320a | miR-19a-3p | miR-146a | miR-221 | miR-550a-5p |
| miR-483-3p | miR-20a | miR-148a | miR-222 | miR-602 |
| miR-1273a | miR-22-3p | miR-148b | miR-223-5p | miR-638 |
| miR-4524a-5p | miR-23a | miR-149 | miR-302a | miR-642a-3p |
| miR-4634 | miR-34a | miR-149-3p | miR-302b | miR-654-5p |
| miR-34b | miR-150 | miR-326 | miR-744-5p | |
| miR-92a-1-5p | miR-181a | miR-361 | miR-765 | |
| miR-96 | miR-181b | miR-371a-5p | miR-2278 | |
| miR-101-3p | miR-182 | miR-371a-3p | miR-3145-5p | |
| miR-101-5p | miR-183 | miR-372-5p | miR-3146 | |
| miR-105 | miR-186 | miR-372-3p | miR-3149 | |
| miR-122-5p | miR-197-3p | miR-373-5p | miR-3679-5p | |
| miR-125a | miR-199b | miR-373-3p | miR-4644 | |
| miR-128b | miR-200a | miR-449b-3p | miR-4653-3p | |
| miR-133a | miR-202-3p | miR-483-5p | miR-4725-3p | |
| miR-133b | miR-203 | miR-484 | ||
| Exosomal miR-92b-5p | p-Value | ||||
|---|---|---|---|---|---|
| Total | ≤2 | >2 | |||
| Stage | 0–I | 21 | 10 | 11 | 0.033 * |
| II–III | 38 | 8 | 30 | ||
| Lymph node metastasis | Positive | 24 | 5 | 19 | 0.181 |
| Negative | 35 | 13 | 22 | ||
| ER | Positive | 43 | 10 | 33 | 0.047 * |
| Negative | 16 | 8 | 8 | ||
| PR | Positive | 35 | 6 | 29 | 0.007 ** |
| Negative | 24 | 12 | 12 | ||
| HER-2 | ≥3 | 13 | 7 | 6 | 0.038 * |
| <3 | 46 | 11 | 35 | ||
| Subtypes | Non-TNBC | 52 | 15 | 37 | 0.45 |
| TNBC | 7 | 3 | 4 | ||
| Subtypes | ER-positive | 39 | 8 | 31 | 0.023 * |
| HER-2-positive | 13 | 7 | 6 | ||
| Tissue miR-92b-5p | p-Value | ||||
|---|---|---|---|---|---|
| Total | Low | High | |||
| MTSS1L | Low | 68 | 44 | 24 | 0.001 ** |
| High | 119 | 101 | 18 | ||
| Stage | 0–I | 70 | 51 | 19 | 0.235 |
| II–III | 117 | 94 | 23 | ||
| Lymph node 1 metastasis | Positive | 81 | 65 | 16 | 0.649 |
| Negative | 93 | 72 | 21 | ||
| ER 2 | Positive | 107 | 82 | 25 | 0.51 |
| Negative | 63 | 51 | 12 | ||
| PR 3 | Positive | 91 | 72 | 19 | 0.697 |
| Negative | 77 | 59 | 18 | ||
| HER-2 4 | ≥3 | 57 | 40 | 17 | 0.049 * |
| <3 | 108 | 90 | 18 | ||
| Subtypes | Non-TNBC | 130 | 99 | 31 | 0.28 |
| TNBC | 33 | 28 | 5 | ||
| Subtypes | ER-positive | 73 | 59 | 14 | 0.16 |
| HER-2-positive | 57 | 40 | 17 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kan, J.-Y.; Shih, S.-L.; Yang, S.-F.; Chu, P.-Y.; Chen, F.-M.; Li, C.-L.; Wu, Y.-C.; Yeh, Y.-T.; Hou, M.-F.; Chiang, C.-P. Exosomal microRNA-92b Is a Diagnostic Biomarker in Breast Cancer and Targets Survival-Related MTSS1L to Promote Tumorigenesis. Int. J. Mol. Sci. 2024, 25, 1295. https://doi.org/10.3390/ijms25021295
Kan J-Y, Shih S-L, Yang S-F, Chu P-Y, Chen F-M, Li C-L, Wu Y-C, Yeh Y-T, Hou M-F, Chiang C-P. Exosomal microRNA-92b Is a Diagnostic Biomarker in Breast Cancer and Targets Survival-Related MTSS1L to Promote Tumorigenesis. International Journal of Molecular Sciences. 2024; 25(2):1295. https://doi.org/10.3390/ijms25021295
Chicago/Turabian StyleKan, Jung-Yu, Shen-Liang Shih, Sheau-Fang Yang, Pei-Yi Chu, Fang-Ming Chen, Chung-Liang Li, Yi-Chia Wu, Yao-Tsung Yeh, Ming-Feng Hou, and Chih-Po Chiang. 2024. "Exosomal microRNA-92b Is a Diagnostic Biomarker in Breast Cancer and Targets Survival-Related MTSS1L to Promote Tumorigenesis" International Journal of Molecular Sciences 25, no. 2: 1295. https://doi.org/10.3390/ijms25021295
APA StyleKan, J.-Y., Shih, S.-L., Yang, S.-F., Chu, P.-Y., Chen, F.-M., Li, C.-L., Wu, Y.-C., Yeh, Y.-T., Hou, M.-F., & Chiang, C.-P. (2024). Exosomal microRNA-92b Is a Diagnostic Biomarker in Breast Cancer and Targets Survival-Related MTSS1L to Promote Tumorigenesis. International Journal of Molecular Sciences, 25(2), 1295. https://doi.org/10.3390/ijms25021295