
L-Lactate Treatment at 24 h and 48 h after Acute Experimental Stroke Is Neuroprotective via Activation of the L-Lactate Receptor HCA1

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
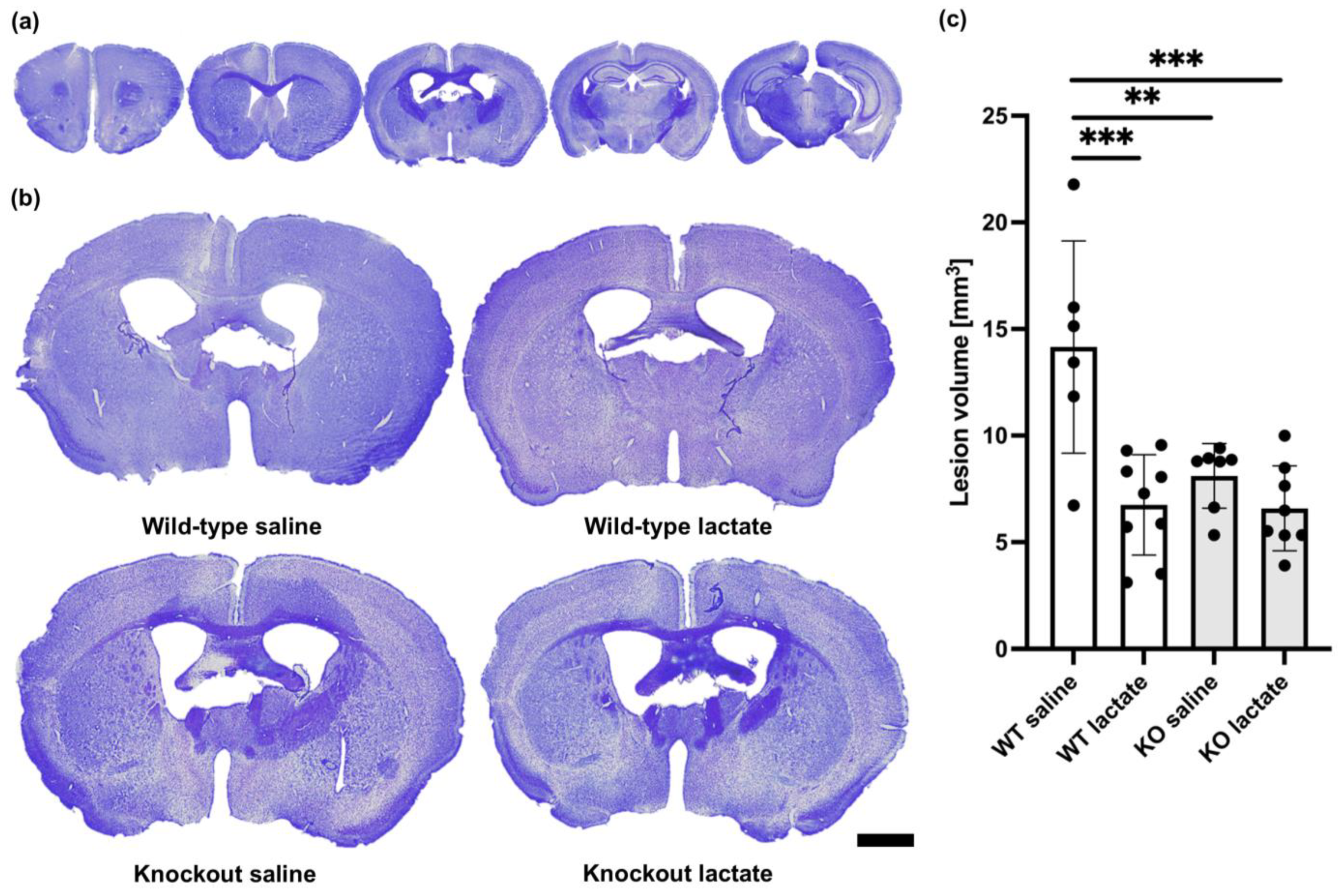
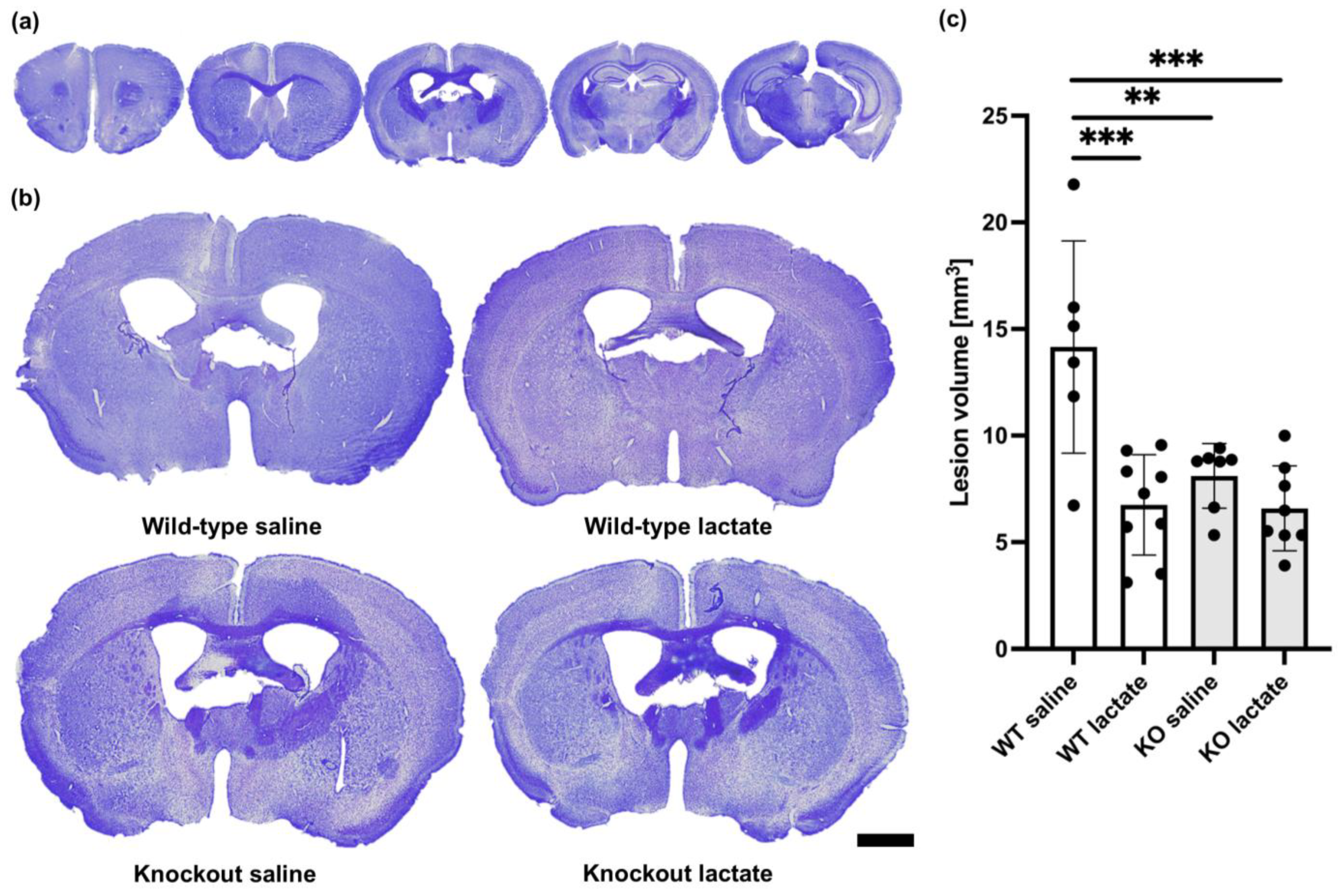
2.1. Lactate Treatment Reduced the Lesion Volume Three Weeks after Stroke Induction in Wild-Type Mice but Not in HCA1 Knockout Mice
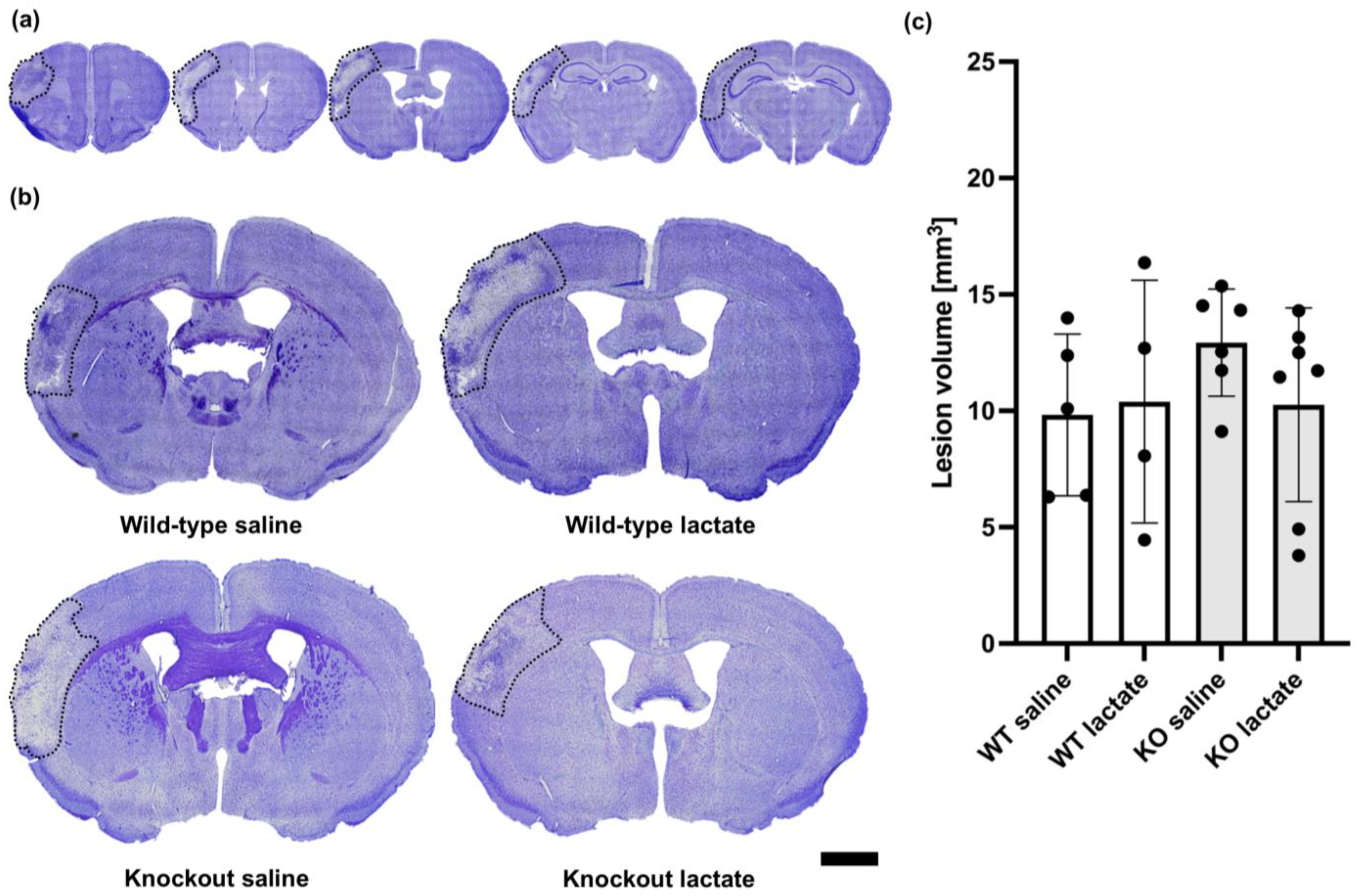
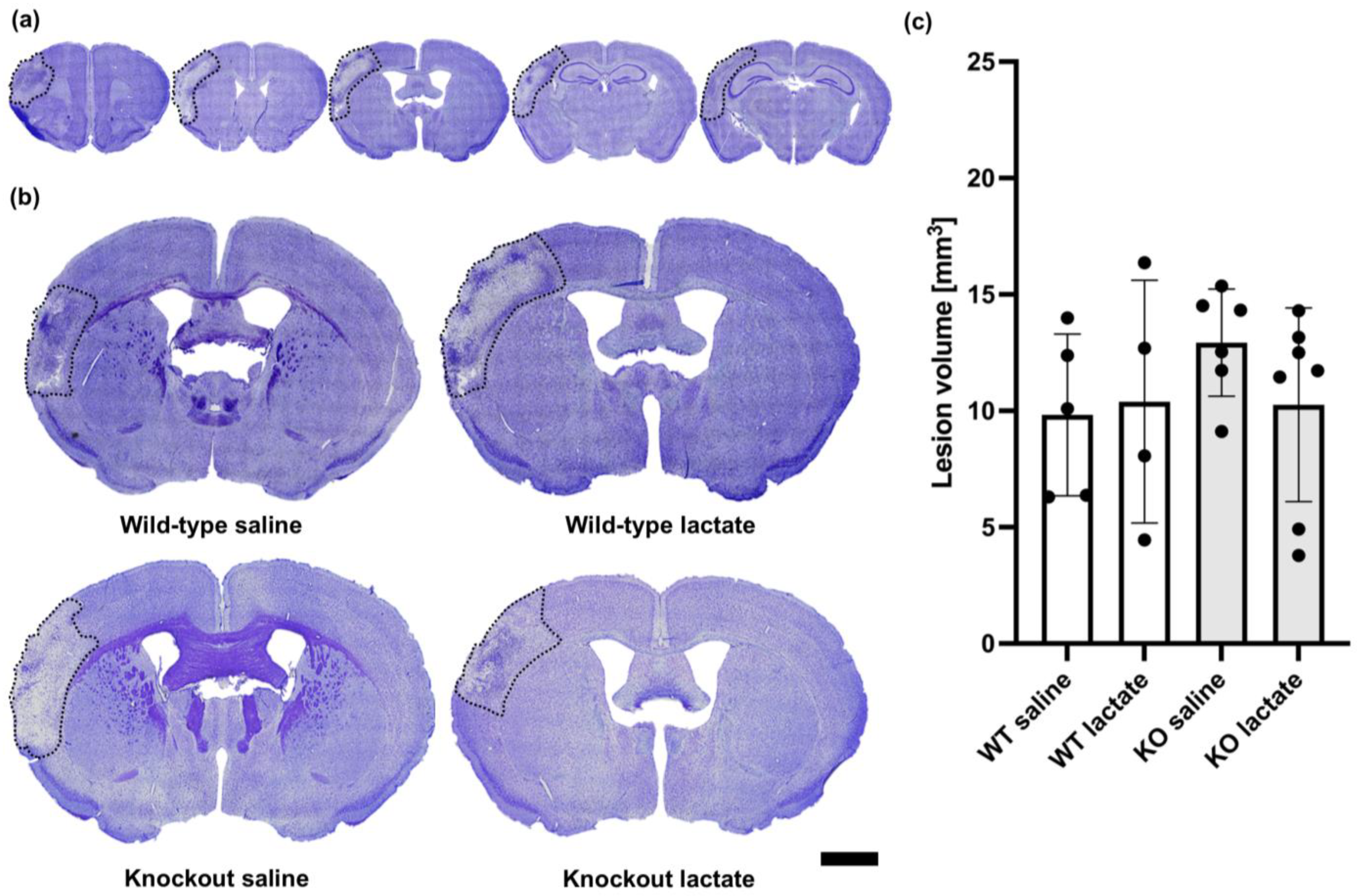
2.2. At One Week after dMCAO, the Lesion Volumes Were Unaffected by the Treatments or Genotypes
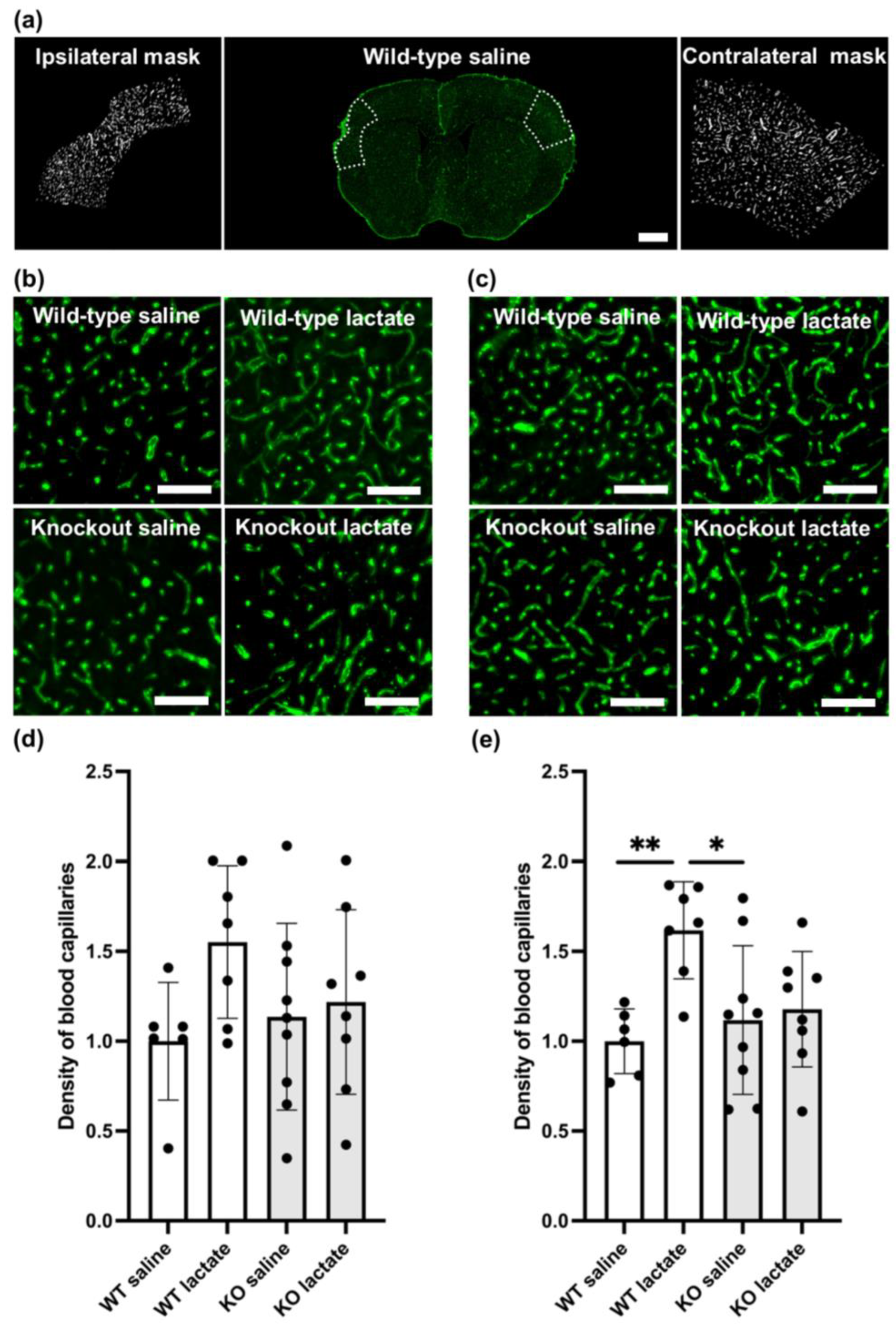
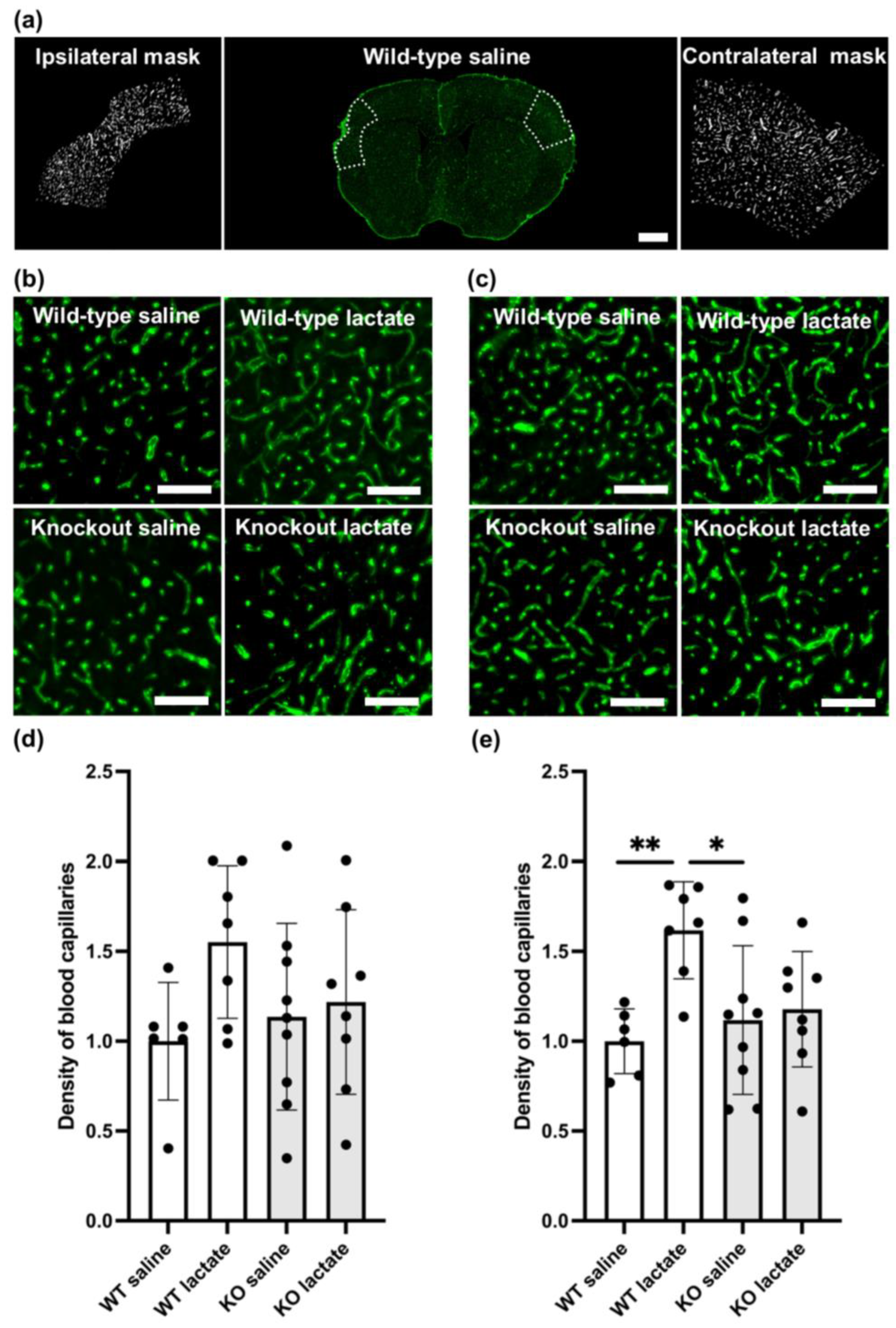
2.3. L-Lactate Treatment Increased Capillary Density Three Weeks after dMCAO in WT Mice, but Not in HCA1 KO Mice
2.4. At One Week after dMCAO, the Capillary Density Was Unaffected by the Treatments or Genotypes
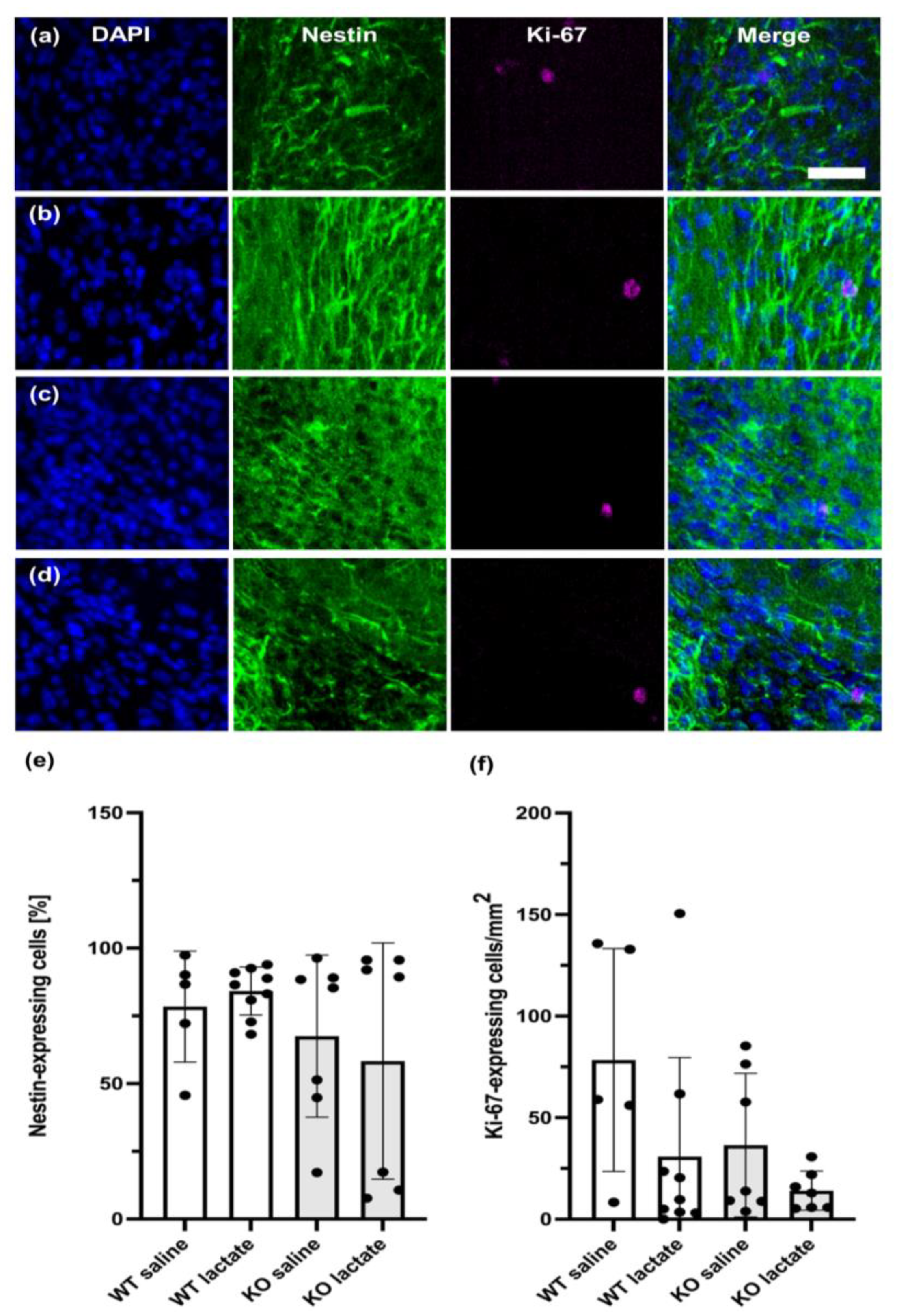
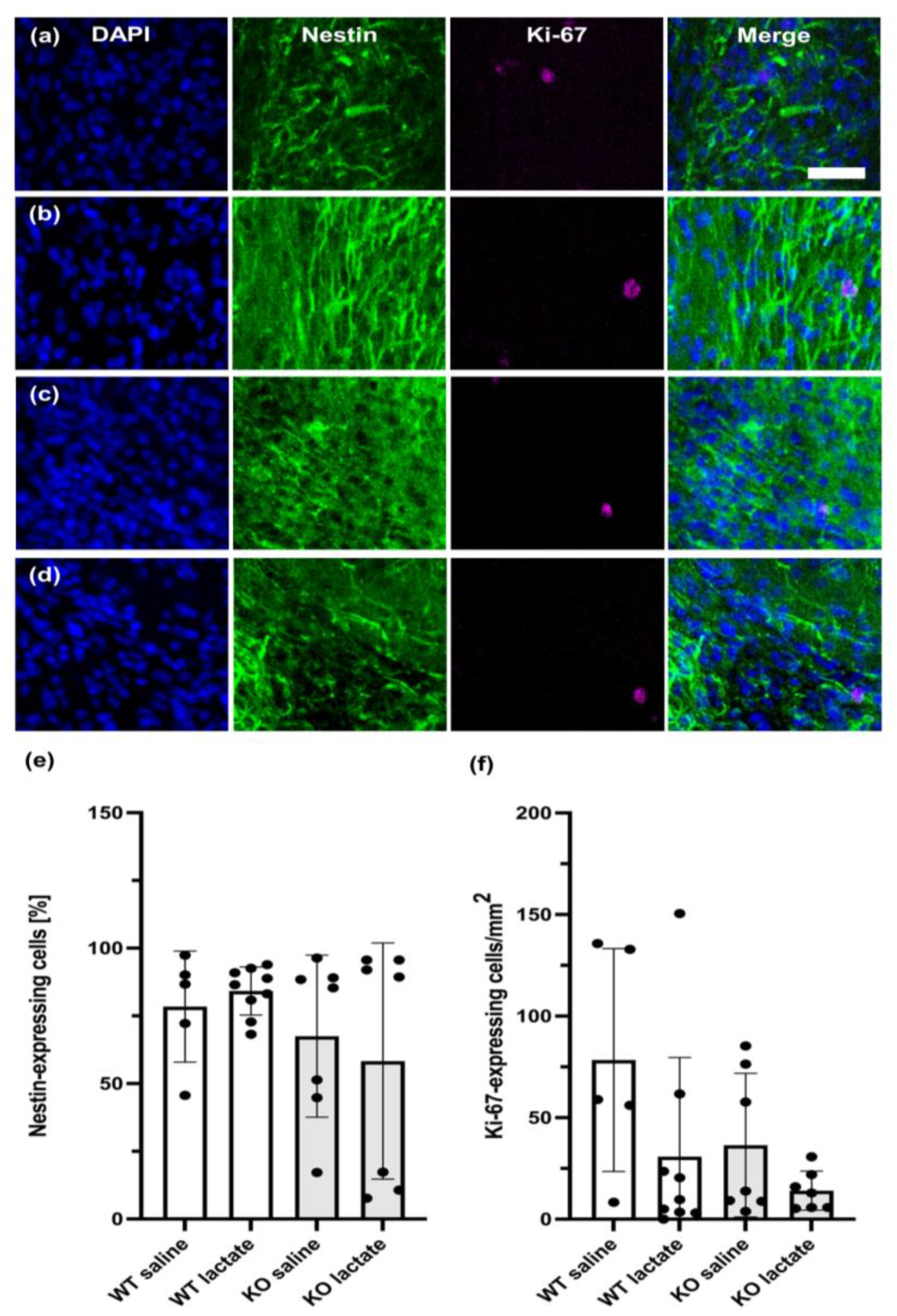
2.5. Neurogenesis Was Unaffected by the Treatments or Genotypes Both at Three Weeks and One Week after dMCAO
3. Discussion
3.1. HCA1-Dependent Neuroprotective Effect of L-Lactate
3.2. A Two-Sided Effect of HCA1 in Stroke Protection
3.3. The Stroke Model
3.4. L-Lactate
3.5. Potential for Translation into the Treatment of Stroke Patients
4. Materials and Methods
4.1. Animals
4.2. Permanent Occlusion of the Distal Medial Cerebral Artery
4.3. L-Lactate and Saline Injections
4.4. Exclusion Criteria
4.5. Tissue Preparation and Analysis of Stroke Volume
4.6. Staining with Cresyl Violet
4.7. Imaging and Lesion Volume Measurements
4.8. Immunohistochemistry
4.9. Image Acquisition and Quantification of Capillaries
4.10. Image Acquisition and Quantification of the Neurogenesis Markers Ki-67 and Nestin
4.11. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katan, M.; Luft, A. Global Burden of Stroke. Semin. Neurol. 2018, 38, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthi, R.V.; Feigin, V.L.; Forouzanfar, M.H.; Mensah, G.A.; Connor, M.; Bennett, D.A.; Moran, A.E.; Sacco, R.L.; Anderson, L.M.; Truelsen, T.; et al. Global and Regional Burden of First-Ever Ischaemic and Haemorrhagic Stroke during 1990–2010: Findings from the Global Burden of Disease Study 2010. Lancet Glob. Health 2013, 1, e259–e281. [Google Scholar] [CrossRef] [PubMed]
- Chrissobolis, S.; Sobey, C.G. 1—Vascular Biology and Atherosclerosis of Cerebral Vessels. In Stroke: Pathophysiology, Diagnosis, and Management; Grotta, J.C., Albers, G.W., Broderick, J.P., Kasner, S.E., Lo, E.H., Mendelow, A.D., Sacco, R.L., Wong, L.K.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 3–12. [Google Scholar]
- Auer, R.N. 4—Histopathology of Brain Tissue Response to Stroke and Injury. In Stroke: Pathophysiology, Diagnosis, and Management; Grotta, J.C., Albers, G.W., Broderick, J.P., Kasner, S.E., Lo, E.H., Mendelow, A.D., Sacco, R.L., Wong, L.K.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 47–59. [Google Scholar]
- Dirnagl, U.; Iadecola, C.; Moskowitz, M.A. Pathobiology of Ischaemic Stroke: An Integrated View. Trends Neurosci. 1999, 22, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Virani, S.S.; Callaway, C.W.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Chiuve, S.E.; Cushman, M.; Delling, F.N.; Deo, R.; et al. Heart Disease and Stroke Statistics-2018 Update: A Report from the American Heart Association. Circulation 2018, 137, e67–e492. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, L.; Jespersen, S.N.; Mouridsen, K.; Mikkelsen, I.K.; Jonsdottír, K.; Tietze, A.; Blicher, J.U.; Aamand, R.; Hjort, N.; Iversen, N.K.; et al. The Role of the Cerebral Capillaries in Acute Ischemic Stroke: The Extended Penumbra Model. J. Cereb. Blood Flow Metab. 2013, 33, 635–648. [Google Scholar] [CrossRef]
- Robinson, T.; Zaheer, Z.; Mistri, A.K. Thrombolysis in Acute Ischaemic Stroke: An Update. Ther. Adv. Chronic Dis. 2011, 2, 119–131. [Google Scholar] [CrossRef]
- Koh, S.H.; Park, H.H. Neurogenesis in Stroke Recovery. Transl. Stroke Res. 2017, 8, 3–13. [Google Scholar] [CrossRef]
- Zhao, L.R.; Willing, A. Enhancing Endogenous Capacity to Repair a Stroke-Damaged Brain: An Evolving Field for Stroke Research. Prog. Neurobiol. 2018, 163–164, 5–26. [Google Scholar] [CrossRef]
- Krupinski, J.; Kaluza, J.; Kumar, P.; Kumar, S.; Wang, J.M. Role of Angiogenesis in Patients with Cerebral Ischemic Stroke. Stroke 1994, 25, 1794–1798. [Google Scholar] [CrossRef]
- Boero, J.A.; Ascher, J.; Arregui, A.; Rovainen, C.; Woolsey, T.A. Increased Brain Capillaries in Chronic Hypoxia. J. Appl. Physiol. 1999, 86, 1211–1219. [Google Scholar] [CrossRef]
- Harik, N.; Harik, S.I.; Kuo, N.-T.; Sakai, K.; Przybylski, R.J.; LaManna, J.C. Time-Course and Reversibility of the Hypoxia-Induced Alterations in Cerebral Vascularity and Cerebral Capillary Glucose Transporter Density. Brain Res. 1996, 737, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Geiseler, S.J.; Morland, C. The Janus Face of VEGF in Stroke. Int. J. Mol. Sci. 2018, 19, 1362. [Google Scholar] [CrossRef] [PubMed]
- Ali, C.; Nicole, O.; Docagne, F.; Lesne, S.; MacKenzie, E.T.; Nouvelot, A.; Buisson, A.; Vivien, D. Ischemia-Induced Interleukin-6 as a Potential Endogenous Neuroprotective Cytokine against NMDA Receptor-Mediated Excitotoxicity in the Brain. J. Cereb. Blood Flow Metab. 2000, 20, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Grønhøj, M.H.; Clausen, B.H.; Fenger, C.D.; Lambertsen, K.L.; Finsen, B. Beneficial Potential of Intravenously Administered IL-6 in Improving Outcome after Murine Experimental Stroke. Brain Behav. Immun. 2017, 65, 296–311. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Zhang, J.C.; Shi, R.L.; Zhang, S.H.; Yuan, S.Y. Inhibition of Interleukin-6 Abolishes the Promoting Effects of Pair Housing on Post-Stroke Neurogenesis. Neuroscience 2015, 307, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Berthet, C.; Castillo, X.; Magistretti, P.J.; Hirt, L. New Evidence of Neuroprotection by Lactate after Transient Focal Cerebral Ischaemia: Extended Benefit after Intracerebroventricular Injection and Efficacy of Intravenous Administration. Cerebrovasc. Dis. 2012, 34, 329–335. [Google Scholar] [CrossRef]
- Roumes, H.; Dumont, U.; Sanchez, S.; Mazuel, L.; Blanc, J.; Raffard, G.; Chateil, J.-F.; Pellerin, L.; Bouzier-Sore, A.-K. Neuroprotective Role of Lactate in Rat Neonatal Hypoxia-Ischemia. J. Cereb. Blood Flow Metab. 2021, 41, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Morland, C.; Lauritzen, K.H.; Puchades, M.; Holm-Hansen, S.; Andersson, K.; Gjedde, A.; Attramadal, H.; Storm-Mathisen, J.; Bergersen, L.H. The Lactate Receptor, G-Protein-Coupled Receptor 81/Hydroxycarboxylic Acid Receptor 1: Expression and Action in Brain. J. Neurosci. Res. 2015, 93, 1045–1055. [Google Scholar] [CrossRef]
- Morland, C.; Andersson, K.A.; Haugen, Ø.P.; Hadzic, A.; Kleppa, L.; Gille, A.; Rinholm, J.E.; Palibrk, V.; Diget, E.H.; Kennedy, L.H.; et al. Exercise Induces Cerebral VEGF and Angiogenesis via the Lactate Receptor HCAR1. Nat. Commun. 2017, 8, 15557. [Google Scholar] [CrossRef]
- Lambertus, M.; Øverberg, L.T.; Andersson, K.A.; Hjelden, M.S.; Hadzic, A.; Haugen, Ø.P.; Storm-Mathisen, J.; Bergersen, L.H.; Geiseler, S.; Morland, C. L-lactate Induces Neurogenesis in the Mouse Ventricular-Subventricular Zone via the Lactate Receptor HCA(1). Acta Physiol. 2021, 231, e13587. [Google Scholar] [CrossRef]
- El-Brolosy, M.A.; Stainier, D.Y.R. Genetic Compensation: A Phenomenon in Search of Mechanisms. PLoS Genet. 2017, 13, e1006780. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Jiang, L.; Yuan, Y.; Deng, T.; Zheng, Y.R.; Zhao, Y.Y.; Li, W.L.; Wu, J.Y.; Gao, J.Q.; Hu, W.W.; et al. Inhibition of G Protein-Coupled Receptor 81 (GPR81) Protects against Ischemic Brain Injury. CNS Neurosci. Ther. 2015, 21, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Taggart, A.K.; Kero, J.; Gan, X.; Cai, T.Q.; Cheng, K.; Ippolito, M.; Ren, N.; Kaplan, R.; Wu, K.; Wu, T.J.; et al. (D)-β-Hydroxybutyrate Inhibits Adipocyte Lipolysis via the Nicotinic Acid Receptor PUMA-G. J. Biol. Chem. 2005, 280, 26649–26652. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Muhammad, S.; Khan, M.A.; Chen, H.; Ridder, D.A.; Müller-Fielitz, H.; Pokorná, B.; Vollbrandt, T.; Stölting, I.; Nadrowitz, R.; et al. The β-Hydroxybutyrate Receptor HCA2 Activates a Neuroprotective Subset of Macrophages. Nat. Commun. 2014, 5, 3944. [Google Scholar] [CrossRef] [PubMed]
- Llovera, G.; Roth, S.; Plesnila, N.; Veltkamp, R.; Liesz, A. Modeling Stroke in Mice: Permanent Coagulation of the Distal Middle Cerebral Artery. J. Vis. Exp. 2014, 89, e51729. [Google Scholar] [CrossRef]
- Howells, D.W.; Porritt, M.J.; Rewell, S.S.; O’Collins, V.; Sena, E.S.; van der Worp, H.B.; Traystman, R.J.; Macleod, M.R. Different Strokes for Different Folks: The Rich Diversity of Animal Models of Focal Cerebral Ischemia. J. Cereb. Blood Flow Metab. 2010, 30, 1412–1431. [Google Scholar] [CrossRef]
- Carmichael, S.T. Rodent Models of Focal Stroke: Size, Mechanism, and Purpose. NeuroRx 2005, 2, 396–409. [Google Scholar] [CrossRef]
- Stroke Therapy Academic Industry Roundtable. Recommendations for Standards Regarding Preclinical Neuroprotective and Restorative Drug Development. Stroke 1999, 30, 2752–2758. [Google Scholar] [CrossRef]
- McBride, D.W.; Zhang, J.H. Precision Stroke Animal Models: The Permanent MCAO Model Should Be the Primary Model, Not Transient MCAO. Transl. Stroke Res. 2017, 8, 397–404. [Google Scholar] [CrossRef]
- Hossmann, K.A. The Two Pathophysiologies of Focal Brain Ischemia: Implications for Translational Stroke Research. J. Cereb. Blood Flow Metab. 2012, 32, 1310–1316. [Google Scholar] [CrossRef]
- Jeon, S.B.; Koh, Y.; Choi, H.A.; Lee, K. Critical Care for Patients with Massive Ischemic Stroke. J. Stroke 2014, 16, 146–160. [Google Scholar] [CrossRef] [PubMed]
- de Castro Abrantes, H.; Briquet, M.; Schmuziger, C.; Restivo, L.; Puyal, J.; Rosenberg, N.; Rocher, A.B.; Offermanns, S.; Chatton, J.Y. The Lactate Receptor HCAR1 Modulates Neuronal Network Activity through the Activation of G(α) and G(βγ) Subunits. J. Neurosci. 2019, 39, 4422–4433. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Davenport, R.; Stafford, S.; Grosse, J.; Ogawa, K.; Cameron, J.; Parton, L.; Sykes, A.; Mack, S.; Bousba, S.; et al. Identification of a Novel GPR81-Selective agonist that Suppresses Lipolysis in Mice without Cutaneous Flushing. Eur. J. Pharmacol. 2014, 727, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR Drug Discovery: New Agents, Targets and Indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Pitts, F.N., Jr.; McClure, J.N., Jr. Lactate Metabolism in Anxiety Neurosis. N. Engl. J. Med. 1967, 277, 1329–1336. [Google Scholar] [CrossRef]
- Ahmed, K.; Tunaru, S.; Tang, C.; Müller, M.; Gille, A.; Sassmann, A.; Hanson, J.; Offermanns, S. An Autocrine Lactate Loop Mediates Insulin-Dependent Inhibition of Lipolysis through GPR81. Cell Metab. 2010, 11, 311–319. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE Guidelines 2.0: Updated Guidelines for Reporting Animal Research. J. Physiol. 2020, 598, 3793–3801. [Google Scholar] [CrossRef]
- Swanson, R.A.; Morton, M.T.; Tsao-Wu, G.; Savalos, R.A.; Davidson, C.; Sharp, F.R. A Semiautomated Method for Measuring Brain Infarct Volume. J. Cereb. Blood Flow Metab. 1990, 10, 290–293. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geiseler, S.J.; Hadzic, A.; Lambertus, M.; Forbord, K.M.; Sajedi, G.; Liesz, A.; Morland, C. L-Lactate Treatment at 24 h and 48 h after Acute Experimental Stroke Is Neuroprotective via Activation of the L-Lactate Receptor HCA1. Int. J. Mol. Sci. 2024, 25, 1232. https://doi.org/10.3390/ijms25021232
Geiseler SJ, Hadzic A, Lambertus M, Forbord KM, Sajedi G, Liesz A, Morland C. L-Lactate Treatment at 24 h and 48 h after Acute Experimental Stroke Is Neuroprotective via Activation of the L-Lactate Receptor HCA1. International Journal of Molecular Sciences. 2024; 25(2):1232. https://doi.org/10.3390/ijms25021232
Chicago/Turabian StyleGeiseler, Samuel J., Alena Hadzic, Marvin Lambertus, Karl Martin Forbord, Ghazal Sajedi, Arthur Liesz, and Cecilie Morland. 2024. "L-Lactate Treatment at 24 h and 48 h after Acute Experimental Stroke Is Neuroprotective via Activation of the L-Lactate Receptor HCA1" International Journal of Molecular Sciences 25, no. 2: 1232. https://doi.org/10.3390/ijms25021232
APA StyleGeiseler, S. J., Hadzic, A., Lambertus, M., Forbord, K. M., Sajedi, G., Liesz, A., & Morland, C. (2024). L-Lactate Treatment at 24 h and 48 h after Acute Experimental Stroke Is Neuroprotective via Activation of the L-Lactate Receptor HCA1. International Journal of Molecular Sciences, 25(2), 1232. https://doi.org/10.3390/ijms25021232