
Cellular and Molecular Effects of Magnetic Fields




Abstract
1. Introduction
1.1. Magnetic Field Parameters
1.2. Magnetic Fields and Magnetosensitivity of Biological Systems
1.3. Geomagnetic Fields (GMFs)
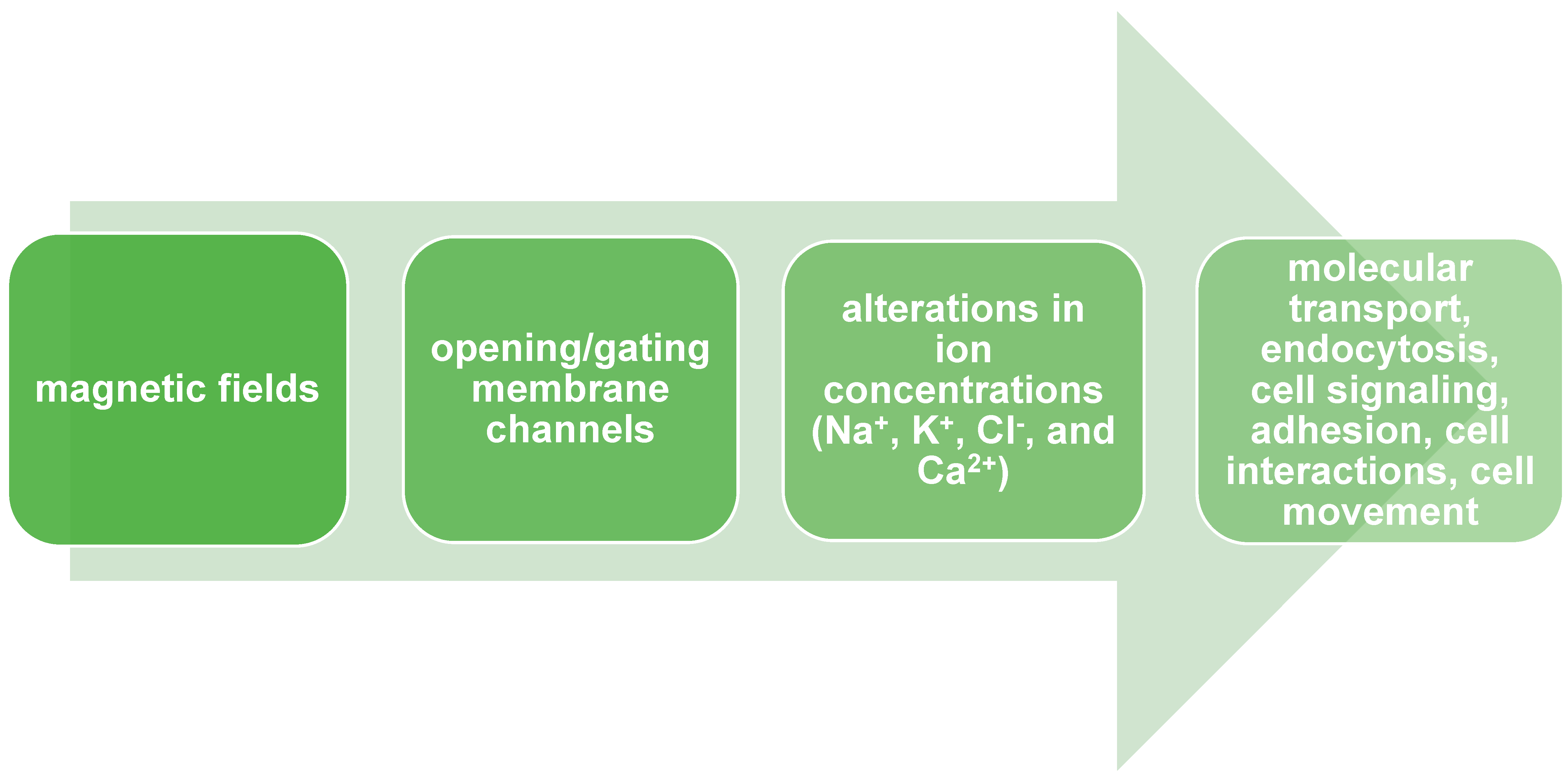
2. Biological Properties of Membranes
3. Cell Morphology and Motility
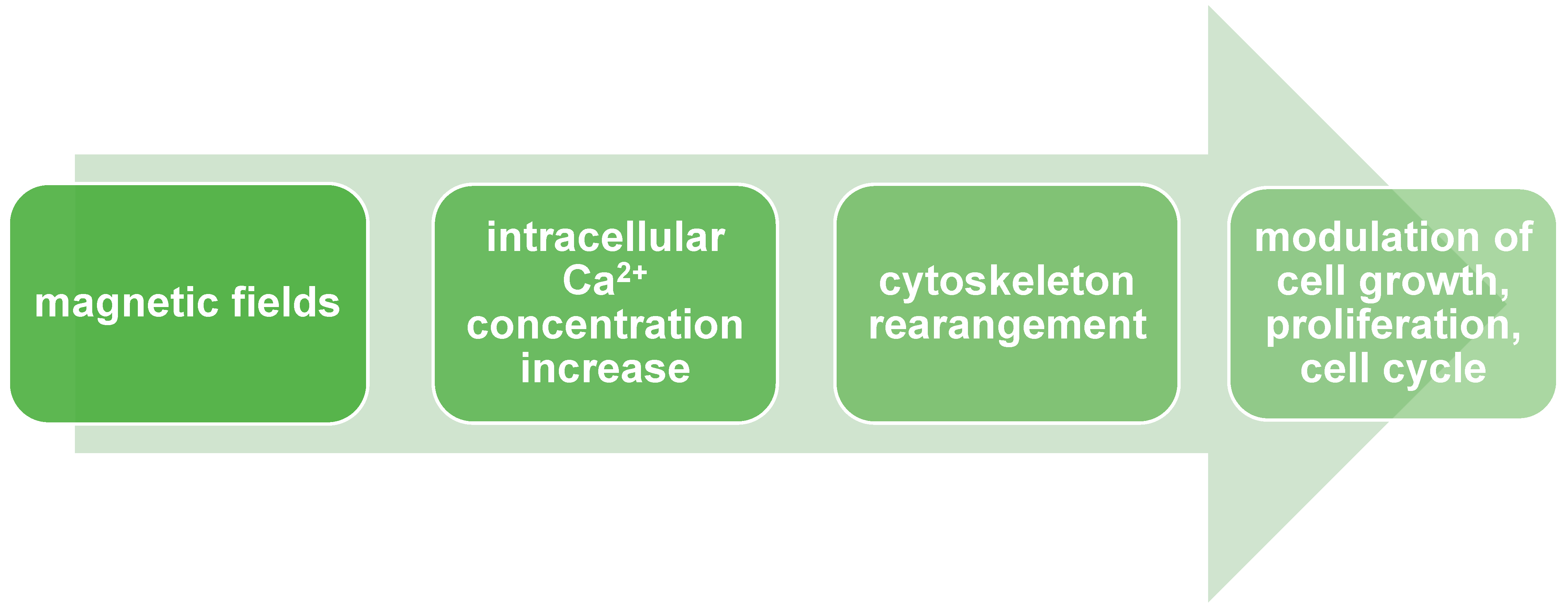
4. Cellular Differentiation and Proliferation
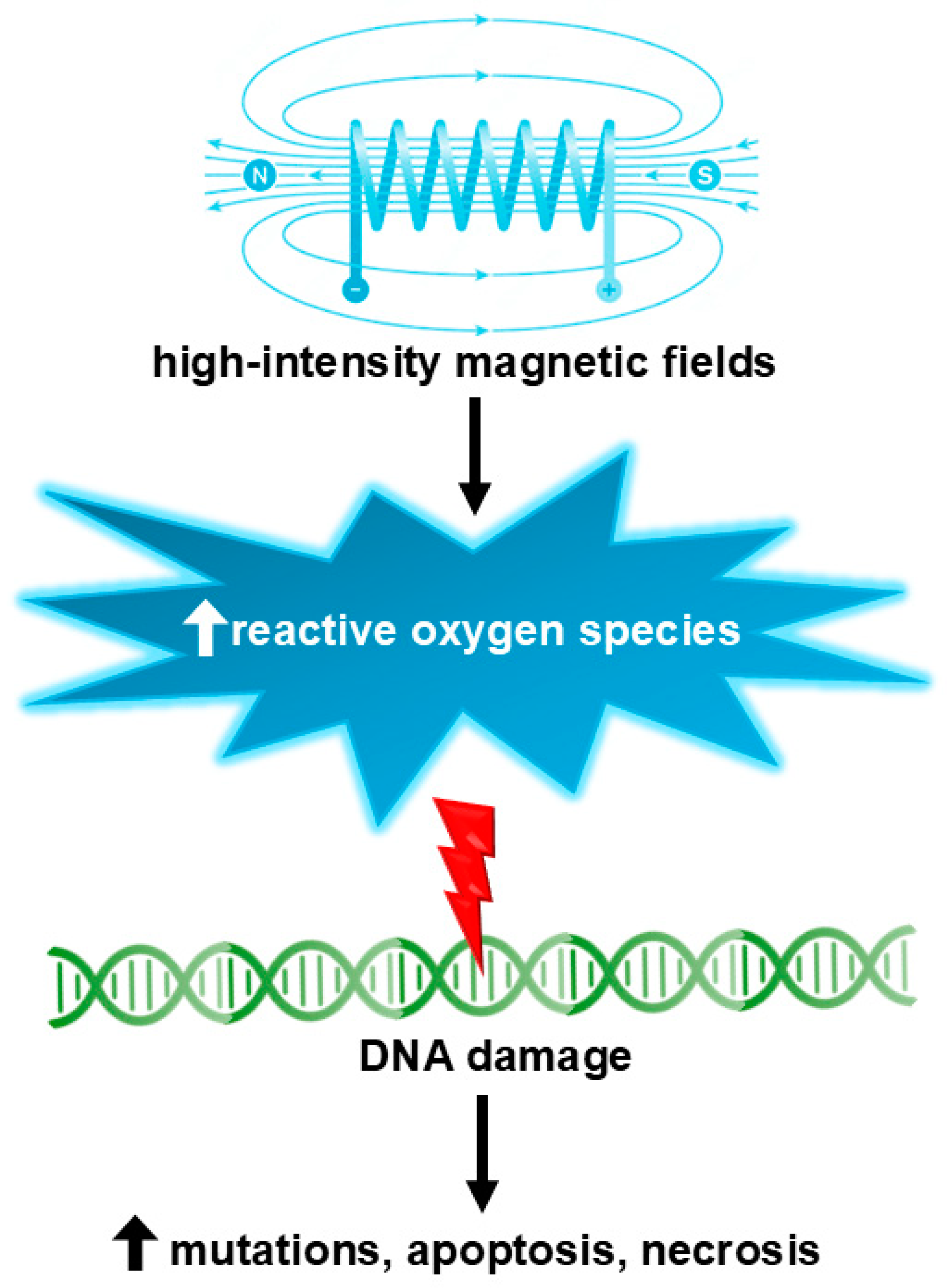
5. Potential Genotoxicity of Magnetic Fields
6. Neurological Effects
7. Potential Therapeutic Applications of Magnetic Fields
7.1. Transcranial Magnetic Stimulation (TMS)
7.2. Magnetic Seizure Therapy (MST)
8. Electromagnetic Sensing and Imaging
8.1. Magnetocardiography
8.2. Magnetoencaphalopathy
8.3. Magnetic Resonance Imaging (MRI) Enhancement
9. Biomedical Applications of Magnetic Fields
9.1. Magnetic Hyperthermia
9.2. Magnetic Targeting
9.3. Regenerative Medicine
10. Interactions with Nanoparticles
11. Conclusions
12. Future Directions and Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Yarema, K.; Xu, A. Biological Effects of Static Magnetic Fields; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–220. [Google Scholar] [CrossRef]
- Patruno, A.; Pesce, M.; Grilli, A.; Speranza, L.; Franceschelli, S.; De Lutiis, M.A.; Vianale, G.; Costantini, E.; Amerio, P.; Muraro, R.; et al. MTOR Activation by PI3K/Akt and ERK Signaling in Short ELF-EMF Exposed Human Keratinocytes. PLoS ONE 2015, 10, e0139644. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Chang, C.W.; Chen, C.R.; Chuang, C.Y.; Chiang, C.S.; Shu, W.Y.; Fan, T.C.; Hsu, I.C. Extremely Low-Frequency Electromagnetic Fields Cause G1 Phase Arrest through the Activation of the ATM-Chk2-P21 Pathway. PLoS ONE 2014, 9, e0104732. [Google Scholar] [CrossRef]
- Kim, J.; Ha, C.S.; Lee, H.J.; Song, K. Repetitive Exposure to a 60-Hz Time-Varying Magnetic Field Induces DNA Double-Strand Breaks and Apoptosis in Human Cells. Biochem. Biophys. Res. Commun. 2010, 400, 739–744. [Google Scholar] [CrossRef]
- Kim, J.; Yoon, Y.; Yun, S.; Park, G.S.; Lee, H.J.; Song, K. Time-Varying Magnetic Fields of 60 Hz at 7 MT Induce DNA Double-Strand Breaks and Activate DNA Damage Checkpoints without Apoptosis. Bioelectromagnetics 2012, 33, 383–393. [Google Scholar] [CrossRef]
- Benassi, B.; Filomeni, G.; Montagna, C.; Merla, C.; Lopresto, V.; Pinto, R.; Marino, C.; Consales, C. Extremely Low Frequency Magnetic Field (ELF-MF) Exposure Sensitizes SH-SY5Y Cells to the Pro-Parkinson’s Disease Toxin MPP(.). Mol. Neurobiol. 2016, 53, 4247–4260. [Google Scholar] [CrossRef] [PubMed]
- Osera, C.; Amadio, M.; Falone, S.; Fassina, L.; Magenes, G.; Amicarelli, F.; Ricevuti, G.; Govoni, S.; Pascale, A. Pre-Exposure of Neuroblastoma Cell Line to Pulsed Electromagnetic Field Prevents H2O2-Induced ROS Production by Increasing MnSOD Activity. Bioelectromagnetics 2015, 36, 219–232. [Google Scholar] [CrossRef]
- Zhou, A.; Liu, M.; Wang, X.; Yu, H.; Zhu, M.; Pang, L.; Wang, L.; Berg, H. Effects of ELF Inductively Coupled Weak Magnetic Fields on Proliferation of 6B1 Cells. Electro Magnetobiol. 1999, 18, 325–331. [Google Scholar] [CrossRef]
- Tokalov, S.V.; Gutzeit, H.O. Weak Electromagnetic Fields (50 Hz) Elicit a Stress Response in Human Cells. Environ. Res. 2004, 94, 145–151. [Google Scholar] [CrossRef]
- Srdjenovic, B.; Mrdjanovic, J.; Galovic, A.J.; Kladar, N.; Bozin, B.; Jurisic, V.; Bogdanovic, G. Effect of ELF-EMF on Antioxidant Status and Micronuclei in K562 Cells and Normal Lymphocytes. Cent. Eur. J. Biol. 2014, 9, 931–940. [Google Scholar] [CrossRef]
- Khavari, B.; Ahmadian, S.; Bolouri, B. The Effects of 50Hz, 0.6 MT Extremely Low Frequency (ELF) Electromagnetic Field (EMF) on Proliferation of the Prostate Cancer Cell Line, DU-145. Clin. Biochem. 2011, 44, S165. [Google Scholar] [CrossRef]
- Yoshizawa, H.; Tsuchiya, T.; Mizoe, H.; Ozeki, H.; Kanao, S.; Yomori, H.; Sakane, C.; Hasebe, S.; Motomura, T.; Yamakawa, T.; et al. No Effect of Extremely Low-Frequency Magnetic Field Observed on Cell Growth or Initial Response of Cell Proliferation in Human Cancer Cell Lines. Bioelectromagnetics 2002, 23, 355–368. [Google Scholar] [CrossRef]
- Zablotskii, V.; Polyakova, T.; Lunov, O.; Dejneka, A. How a High-Gradient Magnetic Field Could Affect Cell Life. Sci. Rep. 2016, 6, 37407. [Google Scholar] [CrossRef] [PubMed]
- Zablotskii, V.; Syrovets, T.; Schmidt, Z.W.; Dejneka, A.; Simmet, T. Modulation of Monocytic Leukemia Cell Function and Survival by High Gradient Magnetic Fields and Mathematical Modeling Studies. Biomaterials 2014, 35, 3164–3171. [Google Scholar] [CrossRef] [PubMed]
- Zablotskii, V.; Lunov, O.; Dejneka, A.; Jastrabk, L.; Polyakova, T.; Syrovets, T.; Simmet, T. Nanomechanics of Magnetically Driven Cellular Endocytosis. Appl. Phys. Lett. 2011, 99, 183701. [Google Scholar] [CrossRef]
- Scarfi, M.R.; Lioi, M.B.; Zeni, O.; Della Noce, M.; Franceschi, C.; Bersani, F. Micronucleus Frequency and Cell Proliferation in Human Lymphocytes Exposed to 50 Hz Sinusoidal Magnetic Fields. Health Phys. 1999, 76, 244–250. [Google Scholar] [CrossRef]
- Hore, P.J. Are Biochemical Reactions Affected by Weak Magnetic Fields? Proc. Natl. Acad. Sci. USA 2012, 109, 1357. [Google Scholar] [CrossRef]
- Leyland-Jones, B. Targeted Drug Delivery. Semin Oncol. 2013, 20, 181–234. [Google Scholar] [CrossRef]
- Kaufman, A.A.; Hansen, R.O.; Kleinberg, R.L.K. Chapter 6 Paramagnetism, Diamagnetism, and Ferromagnetism. Methods Geochem. Geophys. 2008, 42, 207–254. [Google Scholar] [CrossRef]
- Albuquerque, W.W.C.; Costa, R.M.P.B.; de Salazar e Fernandes, T.; Porto, A.L.F. Evidences of the Static Magnetic Field Influence on Cellular Systems. Prog. Biophys. Mol. Biol. 2016, 121, 16–28. [Google Scholar] [CrossRef]
- Lezcano, A.G.; Nocera, F.; Caponetto, R.G.; Sinčák, M.; Sedlakova-Kadukova, J. Hypomagnetic Fields and Their Multilevel Effects on Living Organisms. Processes 2023, 11, 282. [Google Scholar] [CrossRef]
- Bowles, J.F.W. Encyclopedia of Geology, 2nd ed.; Oxides; Academic Press: Cambridge, MA, USA, 2021; pp. 428–441. [Google Scholar]
- Panovska, S.; Korte, M.; Constable, C.G. One Hundred Thousand Years of Geomagnetic Field Evolution. Rev. Geophys. 2019, 57, 1289–1337. [Google Scholar] [CrossRef]
- Erdmann, W.; Kmita, H.; Kosicki, J.Z.; Kaczmarek, Ł. How the Geomagnetic Field Influences Life on Earth—An Integrated Approach to Geomagnetobiology. Orig. Life Evol. Biosph. 2021, 51, 231–257. [Google Scholar] [CrossRef] [PubMed]
- Zadeh-Haghighi, H.; Rishabh, R.; Simon, C. Hypomagnetic Field Effects as a Potential Avenue for Testing the Radical Pair Mechanism in Biology. Front. Phys. 2023, 11, 1026460. [Google Scholar] [CrossRef]
- Binhi, V.N.; Prato, F.S. Biological Effects of the Hypomagnetic Field: An Analytical Review of Experiments and Theories. PLoS ONE 2017, 12, e0179340. [Google Scholar] [CrossRef]
- Xu, C.; Li, Y.; Yu, Y.; Zhang, Y.; Wei, S. Suppression of Arabidopsis Flowering by Near-Null Magnetic Field Is Affected by Light. Bioelectromagnetics 2015, 36, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xue, Y.; Yang, J.; Shang, P.; Yuan, X. Biological Effects of Hypomagnetic Field: Ground-Based Data for Space Exploration. Bioelectromagnetics 2021, 42, 516–531. [Google Scholar] [CrossRef]
- Zhang, B.; Tian, L. Reactive Oxygen Species: Potential Regulatory Molecules in Response to Hypomagnetic Field Exposure. Bioelectromagnetics 2020, 41, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Martino, C.F.; Portelli, L.; McCabe, K.; Hernandez, M.; Barnes, F. Reduction of the Earth’s Magnetic Field Inhibits Growth Rates of Model Cancer Cell Lines. Bioelectromagnetics 2010, 31, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Ali, Y.F.; Luo, W.; Liu, C.; Zhou, G.; Liu, N.A. Biological Effects of Space Hypomagnetic Environment on Circadian Rhythm. Front. Physiol. 2021, 12, 643943. [Google Scholar] [CrossRef]
- Zadeh-Haghighi, H.; Simon, C. Magnetic Field Effects in Biology from the Perspective of the Radical Pair Mechanism. J. R. Soc. Interface 2022, 19, 20220325. [Google Scholar] [CrossRef] [PubMed]
- Goychuk, I. Sensing Magnetic Fields with Magnetosensitive Ion Channels. Sensors 2018, 18, 728. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, C.T.; Bazylinski, D.A. Ecology, Diversity, and Evolution of Magnetotactic Bacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 497–526. [Google Scholar] [CrossRef] [PubMed]
- Faivre, D.; Schüler, D. Magnetotactic Bacteria and Magnetosomes. Chem. Rev. 2008, 108, 4875–4898. [Google Scholar] [CrossRef] [PubMed]
- Wiltschko, W.; Wiltschko, R. Magnetic Orientation and Magnetoreception in Birds and Other Animals. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 2005, 191, 675–693. [Google Scholar] [CrossRef]
- Foley, L.E.; Gegear, R.J.; Reppert, S.M. Human Cryptochrome Exhibits Light-Dependent Magnetosensitivity. Nat. Commun. 2011, 2, 356. [Google Scholar] [CrossRef]
- Gorobets, O.; Gorobets, S.; Polyakova, T.; Zablotskii, V. Modulation of Calcium Signaling and Metabolic Pathways in Endothelial Cells with Magnetic Fields. Nanoscale Adv. 2024, 6, 1163–1182. [Google Scholar] [CrossRef] [PubMed]
- Gieré, R. Magnetite in the Human Body: Biogenic vs. Anthropogenic. Proc. Natl. Acad. Sci. USA 2016, 113, 11986–11987. [Google Scholar] [CrossRef]
- Kadir, L.A.; Stacey, M.; Barrett-Jolley, R. Emerging Roles of the Membrane Potential: Action Beyond the Action Potential. Front. Physiol. 2018, 9, 1661. [Google Scholar] [CrossRef]
- Yang, M.; Brackenbury, W.J. Membrane Potential and Cancer Progression. Front. Physiol. 2013, 4, 185. [Google Scholar] [CrossRef] [PubMed]
- Ramahi, A.A.; Ruff, R.L. Membrane Potential. In Encyclopedia of the Neurological Sciences; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1034–1035. [Google Scholar] [CrossRef]
- Escobar, J.F.; Vaca-González, J.J.; Guevara, J.M.; Garzón-Alvarado, D.A. Effect of Magnetic and Electric Fields on Plasma Membrane of Single Cells: A Computational Approach. Eng. Rep. 2020, 2, e12125. [Google Scholar] [CrossRef]
- Rubio Ayala, M.; Syrovets, T.; Hafner, S.; Zablotskii, V.; Dejneka, A.; Simmet, T. Spatiotemporal Magnetic Fields Enhance Cytosolic Ca2+ Levels and Induce Actin Polymerization via Activation of Voltage-Gated Sodium Channels in Skeletal Muscle Cells. Biomaterials 2018, 163, 174–184. [Google Scholar] [CrossRef]
- Yan, J.; Dong, L.; Zhang, B.; Qi, N. Effects of Extremely Low-Frequency Magnetic Field on Growth and Differentiation of Human Mesenchymal Stem Cells. Electromagn. Biol. Med. 2010, 29, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Zablotskii, V.; Polyakova, T.; Dejneka, A. Modulation of the Cell Membrane Potential and Intracellular Protein Transport by High Magnetic Fields. Bioelectromagnetics 2021, 42, 27–36. [Google Scholar] [CrossRef]
- Moorhouse, A.J. Membrane Potential: Concepts. Encycl. Cell Biol. 2016, 1, 218–236. [Google Scholar] [CrossRef]
- Bauréus Koch, C.L.M.; Sommarin, M.; Persson, B.R.R.; Salford, L.G.; Eberhardt, J.L. Interaction between Weak Low Frequency Magnetic Fields and Cell Membranes. Bioelectromagnetics 2003, 24, 395–402. [Google Scholar] [CrossRef]
- Wang, Z.; Che, P.L.; Du, J.; Ha, B.; Yarema, K.J. Static Magnetic Field Exposure Reproduces Cellular Effects of the Parkinson’s Disease Drug Candidate ZM241385. PLoS ONE 2010, 5, e13883. [Google Scholar] [CrossRef] [PubMed]
- Bekhite, M.M.; Figulla, H.R.; Sauer, H.; Wartenberg, M. Static Magnetic Fields Increase Cardiomyocyte Differentiation of Flk-1+ Cells Derived from Mouse Embryonic Stem Cells via Ca2+ Influx and ROS Production. Int. J. Cardiol. 2013, 167, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Bootman, M.D.; Bultynck, G. Fundamentals of Cellular Calcium Signaling: A Primer. Cold Spring Harb. Perspect Biol. 2020, 12, a038802. [Google Scholar] [CrossRef]
- Wu, H.; Li, C.; Masood, M.; Zhang, Z.; González-Almela, E.; Castells-Garcia, A.; Zou, G.; Xu, X.; Wang, L.; Zhao, G.; et al. Static Magnetic Fields Regulate T-Type Calcium Ion Channels and Mediate Mesenchymal Stem Cells Proliferation. Cells 2022, 11, 2460. [Google Scholar] [CrossRef] [PubMed]
- Chiaramello, E.; Fiocchi, S.; Bonato, M.; Gallucci, S.; Benini, M.; Tognola, G.; Ravazzani, P.; Parazzini, M. Contactless Cell Permeabilization by Time-Varying Magnetic Fields: Modelling Transmembrane Potential and Mechanical Stress in In-Vitro Experimental Set-Up. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2021, 2021, 4303–4305. [Google Scholar] [CrossRef]
- Shankayi, Z.; Firoozabadi, S.M.P.; Mansurian, M.G. The Effect of Pulsed Magnetic Field on the Molecular Uptake and Medium Conductivity of Leukemia Cell. Cell Biochem. Biophys. 2013, 65, 211–216. [Google Scholar] [CrossRef]
- Novickij, V.; Dermol, J.; Grainys, A.; Kranjc, M.; Miklavčič, D. Membrane Permeabilization of Mammalian Cells Using Bursts of High Magnetic Field Pulses. PeerJ 2017, 5, e3267. [Google Scholar] [CrossRef] [PubMed]
- Novickij, V.; Girkontaite, I.; Zinkevičiene, A.; Švediene, J.; Lastauskiene, E.; Paškevičius, A.; Markovskaja, S.; Novickij, J. Reversible Permeabilization of Cancer Cells by High Sub-Microsecond Magnetic Field. IEEE Trans. Magn. 2017, 53, 2001904. [Google Scholar] [CrossRef]
- Ye, H.; Cotic, M.; Kang, E.E.; Fehlings, M.G.; Carlen, P.L. Transmembrane Potential Induced on the Internal Organelle by a Time-Varying Magnetic Field: A Model Study. J. Neuroeng. Rehabil. 2010, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Saotome, M.; Katoh, H.; Satoh, T.; Hasan, P.; Ohtani, H.; Satoh, H.; Hayashi, H.; Maekawa, Y. Glycogen Synthase Kinase-3β Opens Mitochondrial Permeability Transition Pore through Mitochondrial Hexokinase II Dissociation. J. Physiol. Sci. 2018, 68, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Qiu, L.; Ye, C.; Chen, L.; Fu, Y.; Sun, W. Exposure to a 50-Hz Magnetic Field Induced Mitochondrial Permeability Transition through the ROS/GSK-3β Signaling Pathway. Int. J. Radiat. Biol. 2016, 92, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Franco-Obregón, A. Harmonizing Magnetic Mitohormetic Regenerative Strategies: Developmental Implications of a Calcium-Mitochondrial Axis Invoked by Magnetic Field Exposure. Bioengineering 2023, 10, 1176. [Google Scholar] [CrossRef]
- Fletcher, D.A.; Mullins, R.D. Cell Mechanics and the Cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef]
- Gartzke, J.; Lange, K. Cellular Target of Weak Magnetic Fields: Ionic Conduction along Actin Filaments of Microvilli. Am. J. Physiol. Cell Physiol. 2002, 283, C1333–C1346. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.C.; Zhang, Z.J.; Wang, D.L.; Liu, Y.; Bartlett, P.F.; He, R.Q. Shielding of the Geomagnetic Field Alters Actin Assembly and Inhibits Cell Motility in Human Neuroblastoma Cells. Sci. Rep. 2016, 6, 22624. [Google Scholar] [CrossRef]
- Zablotskii, V.; Dejneka, A.; Kubinová, Š.; Le-Roy, D.; Dumas-Bouchiat, F.; Givord, D.; Dempsey, N.M.; Syková, E. Life on Magnets: Stem Cell Networking on Micro-Magnet Arrays. PLoS ONE 2013, 8, 70416. [Google Scholar] [CrossRef]
- Lei, H.; Pan, Y.; Wu, R.; Lv, Y. Innate Immune Regulation Under Magnetic Fields with Possible Mechanisms and Therapeutic Applications. Front. Immunol. 2020, 11, 582772. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, Y.; Li, Z.; Ji, X.; Wang, Z.; Wang, H.; Tian, X.; Yu, F.; Yang, Z.; Pi, L.; et al. 27 T Ultra-High Static Magnetic Field Changes Orientation and Morphology of Mitotic Spindles in Human Cells. Elife 2017, 6, 22911. [Google Scholar] [CrossRef] [PubMed]
- Prosser, S.L.; Pelletier, L. Mitotic Spindle Assembly in Animal Cells: A Fine Balancing Act. Nat. Rev. Mol. Cell Biol. 2017, 18, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Dantas, M.; Lima, J.T.; Fer, J.G. Nucleus-Cytoskeleton Crosstalk During Mitotic Entry. Front. Cell Dev. Biol. 2021, 9, 649899. [Google Scholar] [CrossRef] [PubMed]
- Dini, L.; Abbro, L. Bioeffects of Moderate-Intensity Static Magnetic Fields on Cell Cultures. Micron 2005, 36, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ma, K.; Yang, L.; Chen, Y.; Qu, Y.; Wang, Y.; Wang, X.; Yang, F.; Sun, Q.; Song, Z.; et al. Influence of Magnetic Field on Morphological Structures and Physiological Characteristics of BEnd.3 Cells Cultured on Polypyrrole Substrates. RSC Adv. 2019, 9, 40887–40894. [Google Scholar] [CrossRef] [PubMed]
- Chionna, A.; Dwikat, M.; Panzarini, E.; Tenuzzo, B.; Carlà, E.C.; Verri, T.; Pagliara, P.; Abbro, L.; Dini, L. Cell Shape and Plasma Membrane Alterations after Static Magnetic Fields Exposure. Eur. J. Histochem. 2009, 47, 299–308. [Google Scholar] [CrossRef]
- Perez, J.E.; Ravasi, T.; Kosel, J. Corrigendum: Mesenchymal Stem Cells Cultured on Magnetic Nanowire Substrates (2017 Nanotechnology 28 055703). Nanotechnology 2017, 28, 055703. [Google Scholar] [CrossRef]
- Wosik, J.; Chen, W.; Qin, K.; Ghobrial, R.M.; Kubiak, J.Z.; Kloc, M. Magnetic Field Changes Macrophage Phenotype. Biophys. J. 2018, 114, 2001–2013. [Google Scholar] [CrossRef]
- Ji, X.; Tian, X.; Feng, S.; Zhang, L.; Wang, J.; Guo, R.; Zhu, Y.; Yu, X.; Zhang, Y.; Du, H.; et al. Intermittent F-Actin Perturbations by Magnetic Fields Inhibit Breast Cancer Metastasis. Research 2023, 6, 0080. [Google Scholar] [CrossRef]
- Tian, X.; Wang, D.; Zha, M.; Yang, X.; Ji, X.; Zhang, L.; Zhang, X. Magnetic Field Direction Differentially Impacts the Growth of Different Cell Types. Electromagn. Biol. Med. 2018, 37, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Maxson, S.; Lopez, E.A.; Yoo, D.; Danilkovitch-Miagkova, A.; LeRoux, M.A. Concise Review: Role of Mesenchymal Stem Cells in Wound Repair. Stem Cells Transl. Med. 2012, 1, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Liu, J.; Zhen, C.; Wang, Y.; Wei, Y.; Ren, W.; Shang, P. Magnetic Fields as a Potential Therapy for Diabetic Wounds Based on Animal Experiments and Clinical Trials. Cell Prolif. 2021, 54, 12982. [Google Scholar] [CrossRef]
- Kim, S.J.; Jang, Y.W.; Hyung, K.E.; Lee, D.K.; Hyun, K.H.; Jeong, S.H.; Min, K.H.; Kang, W.; Jeong, J.H.; Park, S.Y.; et al. Extremely Low-Frequency Electromagnetic Field Exposure Enhances Inflammatory Response and Inhibits Effect of Antioxidant in RAW 264.7 Cells. Bioelectromagnetics 2017, 38, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Lew, W.Z.; Huang, Y.C.; Huang, K.Y.; Lin, C.T.; Tsai, M.T.; Huang, H.M. Static Magnetic Fields Enhance Dental Pulp Stem Cell Proliferation by Activating the P38 Mitogen-Activated Protein Kinase Pathway as Its Putative Mechanism. J. Tissue Eng. Regen. Med. 2018, 12, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Marędziak, M.; Tomaszewski, K.; Polinceusz, P.; Lewandowski, D.; Marycz, K. Static Magnetic Field Enhances the Viability and Proliferation Rate of Adipose Tissue-Derived Mesenchymal Stem Cells Potentially through Activation of the Phosphoinositide 3-Kinase/Akt (PI3K/Akt) Pathway. Electromagn. Biol. Med. 2017, 36, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Uzieliene, I.; Bernotas, P.; Mobasheri, A.; Bernotiene, E. The Role of Physical Stimuli on Calcium Channels in Chondrogenic Differentiation of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2018, 19, 2998. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Yang, J.; Zhang, G.; Huyan, T.; Shang, P. 16 T High Static Magnetic Field Inhibits Receptor Activator of Nuclear Factor Kappa-Β Ligand-Induced Osteoclast Differentiation by Regulating Iron Metabolism in Raw264.7 Cells. J. Tissue Eng. Regen. Med. 2019, 13, 2181–2190. [Google Scholar] [CrossRef]
- Varani, K.; Vincenzi, F.; Pasquini, S.; Blo, I.; Salati, S.; Cadossi, M.; De Mattei, M. Pulsed Electromagnetic Field Stimulation in Osteogenesis and Chondrogenesis: Signaling Pathways and Therapeutic Implications. Int. J. Mol. Sci. 2021, 22, 809. [Google Scholar] [CrossRef]
- Zhang, K.; Ge, W.B.; Luo, S.T.; Zhou, Z.; Liu, Y.L. Static Magnetic Field Promotes Proliferation, Migration, Differentiation, and AKT Activation of Periodontal Ligament Stem Cells. Cells Tissues Organs 2023, 212, 317–326. [Google Scholar] [CrossRef]
- Zhang, L.; Ji, X.; Yang, X.; Zhang, X. Cell Type- and Density-Dependent Effect of 1 T Static Magnetic Field on Cell Proliferation. Oncotarget 2017, 8, 13126–13141. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, X.; Liu, J.; Luo, Y.; Li, Z.; Ji, X.; Wang, W.; Zhang, X. 1 T Moderate Intensity Static Magnetic Field Affects Akt/MTOR Pathway and Increases the Antitumor Efficacy of MTOR Inhibitors in CNE-2Z Cells. Sci. Bull. 2015, 60, 2120–2128. [Google Scholar] [CrossRef]
- Ashdown, C.P.; Johns, S.C.; Aminov, E.; Unanian, M.; Connacher, W.; Friend, J.; Fuster, M.M. Pulsed Low-Frequency Magnetic Fields Induce Tumor Membrane Disruption and Altered Cell Viability. Biophys. J. 2020, 118, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Tong, Y.; Jia, Y.; Jia, X.; Wang, H.; Chen, Y.; Wu, J.; Jin, W.; Ma, Z.; Cao, K.; et al. Effects of Extremely Low Frequency Electromagnetic Fields on the Tumor Cell Inhibition and the Possible Mechanism. Sci. Rep. 2023, 13, 6989. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Hu, P.; Xiang, L.; Liu, Y.; He, R.; Lu, T. A Static Magnetic Field Inhibits the Migration and Telomerase Function of Mouse Breast Cancer Cells. Biomed. Res. Int. 2020, 2020, 7472618. [Google Scholar] [CrossRef] [PubMed]
- Saunders, R. Static Magnetic Fields: Animal Studies. Prog. Biophys. Mol. Biol. 2005, 87, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Ghodbane, S.; Lahbib, A.; Sakly, M.; Abdelmelek, H. Bioeffects of Static Magnetic Fields: Oxidative Stress, Genotoxic Effects, and Cancer Studies. Biomed. Res. Int. 2013, 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, S.; Ushiyama, A.; Wada, K.; Suzuki, Y.; Hattori, K. In Vivo Genotoxicity of High-Intensity Intermediate Frequency Magnetic Fields in Somatic Cells and Germ Cells. J. Radiat. Res. 2023, 64, 250–260. [Google Scholar] [CrossRef]
- Amara, S.; Douki, T.; Ravanat, J.L.; Garrel, C.; Guiraud, P.; Favier, A.; Sakly, M.; Rhouma, K.B.; Abdelmelek, H. Influence of a Static Magnetic Field (250 MT) on the Antioxidant Response and DNA Integrity in THP1 Cells. Phys. Med. Biol. 2007, 52, 889. [Google Scholar] [CrossRef]
- Potenza, L.; Martinelli, C.; Polidori, E.; Zeppa, S.; Calcabrini, C.; Stocchi, L.; Sestili, P.; Stocchi, V. Effects of a 300 MT Static Magnetic Field on Human Umbilical Vein Endothelial Cells. Bioelectromagnetics 2010, 31, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Herrala, M.; Kumari, K.; Koivisto, H.; Luukkonen, J.; Tanila, H.; Naarala, J.; Juutilainen, J. Genotoxicity of Intermediate Frequency Magnetic Fields in Vitro and in Vivo. Env. Res. 2018, 167, 759–769. [Google Scholar] [CrossRef]
- Takashima, Y.; Miyakoshi, J.; Ikehata, M.; Iwasaka, M.; Ueno, S.; Koana, T. Genotoxic Effects of Strong Static Magnetic Fields in DNA-Repair Defective Mutants of Drosophila Melanogaster. J. Radiat. Res. 2004, 45, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Kim, M.S.; Kim, Y.J.; Choi, Y.J.; Lee, Y.; Chung, H.W. Genotoxic Effects of 3 T Magnetic Resonance Imaging in Cultured Human Lymphocytes. Bioelectromagnetics 2011, 32, 535–542. [Google Scholar] [CrossRef]
- Zafari, J.; Rastegar-Pouyani, N.; Javani Jouni, F.; Najjar, N.; Azarshin, S.Z.; Jafarzadeh, E.; Abdolmaleki, P.; Hoseini Shirazi, F. Static Magnetic Field Reduces Cisplatin Resistance via Increasing Apoptosis Pathways and Genotoxicity in Cancer Cell Lines. Sci. Rep. 2024, 14, 5792. [Google Scholar] [CrossRef] [PubMed]
- Caruso, L.; Wunderle, T.; Lewis, C.M.; Valadeiro, J.; Trauchessec, V.; Trejo Rosillo, J.; Amaral, J.P.; Ni, J.; Jendritza, P.; Fermon, C.; et al. In Vivo Magnetic Recording of Neuronal Activity. Neuron 2017, 95, 1283–1291.e4. [Google Scholar] [CrossRef]
- Xiong, J.; Fox, P.T.; Gao, J.H. Directly Mapping Magnetic Field Effects of Neuronal Activity by Magnetic Resonance Imaging. Hum. Brain Mapp. 2003, 20, 41–49. [Google Scholar] [CrossRef]
- Toyomaki, A.; Yamamoto, T. Observation of Changes in Neural Activity Due to the Static Magnetic Field of an MRI Scanner. J. Magn. Reson. Imaging 2007, 26, 1216–1221. [Google Scholar] [CrossRef]
- Ahmed, Z.; Wieraszko, A. The Mechanism of Magnetic Field-Induced Increase of Excitability in Hippocampal Neurons. Brain Res. 2008, 1221, 30–40. [Google Scholar] [CrossRef]
- Wang, C.X.; Hilburn, I.A.; Wu, D.A.; Mizuhara, Y.; Cousté, C.P.; Abrahams, J.N.H.; Bernstein, S.E.; Matani, A.; Shimojo, S.; Kirschvink, J.L. Transduction of the Geomagnetic Field as Evidenced from Alpha-Band Activity in the Human Brain. eNeuro 2019, 6, ENEURO.0483-18. [Google Scholar] [CrossRef]
- Ye, H.; Hendee, J.; Ruan, J.; Zhirova, A.; Ye, J.; Dima, M. Neuron Matters: Neuromodulation with Electromagnetic Stimulation Must Consider Neurons as Dynamic Identities. J. Neuroeng. Rehabil. 2022, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- Hernando, A.; Galvez, F.; García, M.A.; Soto-León, V.; Alonso-Bonilla, C.; Aguilar, J.; Oliviero, A. Effects of Moderate Static Magnetic Field on Neural Systems Is a Non-Invasive Mechanical Stimulation of the Brain Possible Theoretically? Front. Neurosci. 2020, 14, 419. [Google Scholar] [CrossRef] [PubMed]
- Prina-Mello, A.; Farrell, E.; Prendergast, P.J.; Campbell, V.; Coey, J.M.D. Influence of Strong Static Magnetic Fields on Primary Cortical Neurons. Bioelectromagnetics 2006, 27, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Rotem, A.; Moses, E. Magnetic Stimulation of One-Dimensional Neuronal Cultures. Biophys. J. 2008, 94, 5065–5078. [Google Scholar] [CrossRef] [PubMed]
- Aguila, J.; Cudeiro, J.; Rivadulla, C. Effects of Static Magnetic Fields on the Visual Cortex: Reversible Visual Deficits and Reduction of Neuronal Activity. Cereb Cortex 2016, 26, 628–638. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, M.G.; Anikeeva, P. Magnetic Fields for Modulating the Nervous System. Phys. Today 2021, 74, 28–34. [Google Scholar] [CrossRef]
- Christiansen, M.G.; Senko, A.W.; Anikeeva, P. Magnetic Strategies for Nervous System Control. Annu. Rev. Neurosci. 2019, 42, 271–293. [Google Scholar] [CrossRef] [PubMed]
- Turi, Z.; Hananeia, N.; Shirinpour, S.; Opitz, A.; Jedlicka, P.; Vlachos, A. Dosing Transcranial Magnetic Stimulation of the Primary Motor and Dorsolateral Prefrontal Cortices with Multi-Scale Modeling. Front. Neurosci. 2022, 16, 929814. [Google Scholar] [CrossRef]
- Lefaucheur, J.P. Repetitive Transcranial Magnetic Stimulation (RTMS): Insights into the Treatment of Parkinson’s Disease by Cortical Stimulation. Neurophysiol. Clin. 2006, 36, 125–133. [Google Scholar] [CrossRef]
- Klomjai, W.; Katz, R.; Lackmy-Vallée, A. Basic Principles of Transcranial Magnetic Stimulation (TMS) and Repetitive TMS (RTMS). Ann. Phys. Rehabil. Med. 2015, 58, 208–213. [Google Scholar] [CrossRef]
- Ueno, S.; Sekino, M. Figure-Eight Coils for Magnetic Stimulation: From Focal Stimulation to Deep Stimulation. Front. Hum. Neurosci. 2021, 15, 805971. [Google Scholar] [CrossRef]
- Minzenberg, M.J.; Yoon, J.H. Transcranial Magnetic Stimulation: A Clinical Primer for Nonexperts. J. Psychiatr. Pr. 2020, 26, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Edinoff, A.N.; Hegefeld, T.L.; Petersen, M.; Patterson, J.C.; Yossi, C.; Slizewski, J.; Osumi, A.; Cornett, E.M.; Kaye, A.; Kaye, J.S.; et al. Transcranial Magnetic Stimulation for Post-Traumatic Stress Disorder. Front. Psychiatry 2022, 13, 701348. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Toll, B.A.; Carpenter, M.J.; Nietert, P.J.; Dancy, M.; George, M.S. Repetitive Transcranial Magnetic Stimulation for Tobacco Treatment in Cancer Patients: A Preliminary Report of a One-Week Treatment. J. Smok. Cessat. 2022, 2022, e9. [Google Scholar] [CrossRef] [PubMed]
- Pell, G.S.; Harmelech, T.; Zibman, S.; Roth, Y.; Tendler, A.; Zangen, A. Efficacy of Deep TMS with the H1 Coil for Anxious Depression. J. Clin. Med. 2022, 11, 1015. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, F.; Fichera, M.; Chieffo, R.; Dalla Costa, G.; Pisa, M.; Volonté, M.A.; Falautano, M.; Zangen, A.; Comi, G.; Leocani, L. Bilateral Repetitive Transcranial Magnetic Stimulation with the H-Coil in Parkinson’s Disease: A Randomized, Sham-Controlled Study. Front. Neurol. 2021, 11, 584713. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Chang, M.C. Effect of Repetitive Transcranial Magnetic Stimulation on Pain Management: A Systematic Narrative Review. Front. Neurol. 2020, 11, 507027. [Google Scholar] [CrossRef] [PubMed]
- Dionísio, A.; Duarte, I.C.; Patrício, M.; Castelo-Branco, M. The Use of Repetitive Transcranial Magnetic Stimulation for Stroke Rehabilitation: A Systematic Review. J. Stroke Cerebrovasc. Dis. 2018, 27, 1–31. [Google Scholar] [CrossRef]
- Fisicaro, F.; Lanza, G.; Grasso, A.A.; Pennisi, G.; Bella, R.; Paulus, W.; Pennisi, M. Repetitive Transcranial Magnetic Stimulation in Stroke Rehabilitation: Review of the Current Evidence and Pitfalls. Ther. Adv. Neurol. Disord 2019, 12, 1756286419878317. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Lin, T.; Wu, M.; Cai, G.; Ding, Q.; Xu, J.; Li, W.; Wu, C.; Chen, H.; Lan, Y. Effects of Repetitive Transcranial Magnetic Stimulation on Upper-Limb and Finger Function in Stroke Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Front. Neurol. 2022, 13, 940467. [Google Scholar] [CrossRef]
- Lerner, A.J.; Wassermann, E.M.; Tamir, D.I. Seizures from Transcranial Magnetic Stimulation 2012-2016: Results of a Survey of Active Laboratories and Clinics. Clin. Neurophysiol. 2019, 130, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Classen, J.; Witte, O.W.; Schlaug, G.; Seitz, R.J.; Holthausen, H.; Benecke, R. Epileptic Seizures Triggered Directly by Focal Transcranial Magnetic Stimulation. Electroencephalogr. Clin. Neurophysiol. 1995, 94, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Stultz, D.J.; Osburn, S.; Burns, T.; Pawlowska-Wajswol, S.; Walton, R. Transcranial Magnetic Stimulation (TMS) Safety with Respect to Seizures: A Literature Review. Neuropsychiatr. Dis. Treat. 2020, 16, 2989. [Google Scholar] [CrossRef]
- Trifu, S.; Sevcenco, A.; Stănescu, M.; Drăgoi, A.; Cristea, M. Efficacy of Electroconvulsive Therapy as a Potential First-Choice Treatment in Treatment-Resistant Depression (Review). Exp. Ther. Med. 2021, 22, 10716. [Google Scholar] [CrossRef]
- Andrade, C.; Arumugham, S.S.; Thirthalli, J. Adverse Effects of Electroconvulsive Therapy. Psychiatr. Clin. N. Am. 2016, 39, 513–530. [Google Scholar] [CrossRef] [PubMed]
- Tang, V.M.; Blumberger, D.M.; McClintock, S.M.; Kaster, T.S.; Rajji, T.K.; Downar, J.; Fitzgerald, P.B.; Daskalakis, Z.J. Magnetic Seizure Therapy in Treatment-Resistant Schizophrenia: A Pilot Study. Front. Psychiatry 2018, 8, 325491. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, R.; Prakash, J.; Chatterjee, K. Magnetic Seizure Therapy. Ind. Psychiatry J. 2021, 30, S320. [Google Scholar] [CrossRef] [PubMed]
- Weissman, C.R.; Blumberger, D.M.; Dimitrova, J.; Throop, A.; Voineskos, D.; Downar, J.; Mulsant, B.H.; Rajji, T.K.; Fitzgerald, P.B.; Daskalakis, Z.J. Magnetic Seizure Therapy for Suicidality in Treatment-Resistant Depression. JAMA Netw. Open 2020, 3, e207434. [Google Scholar] [CrossRef]
- Sun, Y.; Farzan, F.; Mulsant, B.H.; Rajji, T.K.; Fitzgerald, P.B.; Barr, M.S.; Downar, J.; Wong, W.; Blumberger, D.M.; Daskalakis, Z.J. Indicators for Remission of Suicidal Ideation Following Magnetic Seizure Therapy in Patients with Treatment-Resistant Depression. JAMA Psychiatry 2016, 73, 337–345. [Google Scholar] [CrossRef]
- Daskalakis, Z.J.; Dimitrova, J.; McClintock, S.M.; Sun, Y.; Voineskos, D.; Rajji, T.K.; Goldbloom, D.S.; Wong, A.H.C.; Knyahnytska, Y.; Mulsant, B.H.; et al. Magnetic Seizure Therapy (MST) for Major Depressive Disorder. Neuropsychopharmacology 2019, 45, 276–282. [Google Scholar] [CrossRef]
- Lone, P.A.; Wani, N.A.; Ul Ain, Q.; Heer, A.; Devi, R.; Mahajan, S. Common Postoperative Complications after General Anesthesia in Oral and Maxillofacial Surgery. Natl. J. Maxillofac. Surg. 2021, 12, 206–210. [Google Scholar] [CrossRef]
- Allan, C.L.; Ebmeier, K.P. The Use of ECT and MST in Treating Depression. Int. Rev. Psychiatry 2011, 23, 400–412. [Google Scholar] [CrossRef]
- Daskalakis, Z.J.; Tamminga, C.; Throop, A.; Palmer, L.; Dimitrova, J.; Farzan, F.; Thorpe, K.E.; McClintock, S.M.; Blumberger, D.M. Confirmatory Efficacy and Safety Trial of Magnetic Seizure Therapy for Depression (CREST-MST): Study Protocol for a Randomized Non-Inferiority Trial of Magnetic Seizure Therapy versus Electroconvulsive Therapy. Trials 2021, 22, 786. [Google Scholar] [CrossRef]
- Camm, A.J.; Henderson, R.; Brisinda, D.; Body, R.; Charles, R.G.; Varcoe, B.; Fenici, R. Clinical Utility of Magnetocardiography in Cardiology for the Detection of Myocardial Ischemia. J. Electrocardiol. 2019, 57, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Goernig, M.; Liehr, M.; Tute, C.; Schlosser, M.; Haueisen, J.; Figulla, H.R.; Leder, U. Magnetocardiography Based Spatiotemporal Correlation Analysis Is Superior to Conventional ECG Analysis for Identifying Myocardial Injury. Ann. Biomed. Eng. 2009, 37, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Mäntynen, V.; Konttila, T.; Stenroos, M. Investigations of Sensitivity and Resolution of ECG and MCG in a Realistically Shaped Thorax Model. Phys. Med. Biol. 2014, 59, 7141–7158. [Google Scholar] [CrossRef] [PubMed]
- Her, A.Y.; Dischl, D.; Kim, Y.H.; Kim, S.W.; Shin, E.S. Magnetocardiography for the Detection of Myocardial Ischemia. Front. Cardiovasc. Med. 2023, 10, 1242215. [Google Scholar] [CrossRef] [PubMed]
- Brala, D.; Thevathasan, T.; Grahl, S.; Barrow, S.; Violano, M.; Bergs, H.; Golpour, A.; Suwalski, P.; Poller, W.; Skurk, C.; et al. Application of Magnetocardiography to Screen for Inflammatory Cardiomyopathy and Monitor Treatment Response. J. Am. Heart Assoc. 2023, 12, e027619. [Google Scholar] [CrossRef] [PubMed]
- Chaikovsky, I.; Li, T.; Zhang, W.; Kazmirchyk, A.; Mjasnikov, G.; Lutay, M.; Lomakovsky, O.; Wenming, J. Value of Magnetocardiography in Chronic Coronary Disease Detection: Results of Multicenter Trial. Eur. Heart J. 2021, 42, ehab724-1171. [Google Scholar] [CrossRef]
- Gussmann, A.W.; Strasburger, J.F.; Wakai, R.T.; Strasburger, J.F. Contribution of Fetal Magnetocardiography to Diagnosis, Risk Assessment, and Treatment of Fetal Arrhythmia. J. Am. Heart Assoc. 2022, 11, 25224. [Google Scholar] [CrossRef]
- Strasburger, J.F.; Cheulkar, B.; Wakai, R.T. Magnetocardiography for Fetal Arrhythmias. Heart Rhythm Off. J. Heart Rhythm Soc. 2008, 5, 1073. [Google Scholar] [CrossRef]
- Bagić, A.I.; Funke, M.E.; Kirsch, H.E.; Tenney, J.R.; Zillgitt, A.J.; Burgess, R.C. The 10 Common Evidence-Supported Indications for MEG in Epilepsy Surgery: An Illustrated Compendium. J. Clin. Neurophysiol. 2020, 37, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, S.M.; Pang, E.W.; Huang, M.; Papanicolaou, A.C.; Lee, R.R. Presurgical Functional Mapping with Magnetoencephalography. Neuroimaging Clin. N. Am. 2020, 30, 159–174. [Google Scholar] [CrossRef]
- Ahlfors, S.P.; Mody, M. Overview of MEG. Organ Res. Methods 2019, 22, 95–115. [Google Scholar] [CrossRef]
- Dean Deyle, G. The Role of MRI in Musculoskeletal Practice: A Clinical Perspective. J. Man. Manip. Ther. 2011, 19, 152. [Google Scholar] [CrossRef] [PubMed]
- Rochwerger, A.; Mattei, J.C. Management of Soft Tissue Tumors of the Musculoskeletal System. Orthop. Traumatol. Surg. Res. 2018, 104, S9–S17. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Federici, M.; Castaldi, A. Imaging of Cranial Nerves: A Pictorial Overview. Insights Imaging 2019, 10, 33. [Google Scholar] [CrossRef]
- Clough, T.J.; Jiang, L.; Wong, K.L.; Long, N.J. Ligand Design Strategies to Increase Stability of Gadolinium-Based Magnetic Resonance Imaging Contrast Agents. Nat. Commun. 2019, 10, 1420. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.S. Nephrogenic Systemic Fibrosis: A Serious Adverse Reaction to Gadolinium-1997-2006-2016. Part 2. Acta Radiol. 2016, 57, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Acharya, S.; Anwar, S.; Madala, S.; Thapa, S.; Maroun, R. Delayed Anaphylaxis Due to Gadolinium- A Rare Clinical Scenario. Radiol. Case Rep. 2021, 16, 2522–2525. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.S.S.R.; Mohammad, F. Magnetic Nanomaterials for Hyperthermia-Based Therapy and Controlled Drug Delivery. Adv. Drug. Deliv. Rev. 2011, 63, 789–808. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Jang, B.; Issadore, D.; Tsourkas, A. Use of Magnetic Fields and Nanoparticles to Trigger Drug Release and Improve Tumor Targeting. Wiley Interdiscip Rev. Nanomed. Nanobiotechnol. 2019, 11, e1571. [Google Scholar] [CrossRef]
- Court, K.A.; Hatakeyama, H.; Wu, S.Y.; Lingegowda, M.S.; Rodríguez-Aguayo, C.; López-Berestein, G.; Ju-Seog, L.; Rinaldi, C.; Juan, E.J.; Sood, A.K.; et al. HSP70 Inhibition Synergistically Enhances the Effects of Magnetic Fluid Hyperthermia in Ovarian Cancer. Mol. Cancer Ther. 2017, 16, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Hilger, I.; Kaiser, W.A. Iron Oxide-Based Nanostructures for MRI and Magnetic Hyperthermia. Nanomedicine 2012, 7, 1443–1459. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, S.; Karska, J.; Tota, M.; Skinderowicz, K.; Kulbacka, J.; Drąg-Zalesińska, M. Natural Compounds in Non-Melanoma Skin Cancer: Prevention and Treatment. Molecules 2024, 29, 728. [Google Scholar] [CrossRef] [PubMed]
- Legge, C.J.; Colley, H.E.; Lawson, M.A.; Rawlings, A.E. Targeted Magnetic Nanoparticle Hyperthermia for the Treatment of Oral Cancer. J. Oral Pathol. Med. 2019, 48, 803–809. [Google Scholar] [CrossRef]
- Taheri-Ledari, R.; Zolfaghari, E.; Zarei-Shokat, S.; Kashtiaray, A.; Maleki, A. A Magnetic Antibody-Conjugated Nano-System for Selective Delivery of Ca(OH)2 and Taxotere in Ovarian Cancer Cells. Commun. Biol. 2022, 5, 995. [Google Scholar] [CrossRef]
- Pi, Y.; Liang, H.; Yu, Q.; Yin, Y.; Xu, H.; Lei, Y.; Han, Z.; Tian, J. Low-frequency Pulsed Electromagnetic Field Inhibits RANKL-induced Osteoclastic Differentiation in RAW264.7 Cells by Scavenging Reactive Oxygen Species. Mol. Med. Rep. 2019, 19, 4129–4136. [Google Scholar] [CrossRef]
- Bekhite, M.M.; Finkensieper, A.; Abou-Zaid, F.A.; El-Shourbagy, I.K.; Omar, K.M.; Figulla, H.R.; Sauer, H.; Wartenberg, M. Static Electromagnetic Fields Induce Vasculogenesis and Chondro-Osteogenesis of Mouse Embryonic Stem Cells by Reactive Oxygen Species-Mediated up-Regulation of Vascular Endothelial Growth Factor. Stem Cells Dev. 2010, 19, 731–743. [Google Scholar] [CrossRef]
- Pesqueira, T.; Costa-Almeida, R.; Gomes, M.E. Uncovering the Effect of Low-Frequency Static Magnetic Field on Tendon-Derived Cells: From Mechanosensing to Tenogenesis. Sci. Rep. 2017, 7, 10948. [Google Scholar] [CrossRef]
- Enoch, S.; Leaper, D.J. Basic Science of Wound Healing. Surgery 2008, 26, 31–37. [Google Scholar] [CrossRef]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur. Surg. Res. 2017, 58, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.N.; Hardman, M.J. Wound Healing: Cellular Mechanisms and Pathological Outcomes. Adv. Surg. Med. Spec. 2023, 10, 341–370. [Google Scholar] [CrossRef]
- Li, J.; Chen, J.; Kirsner, R. Pathophysiology of Acute Wound Healing. Clin. Dermatol. 2007, 25, 9–18. [Google Scholar] [CrossRef]
- Krishnaswamy, V.R.; Mintz, D.; Sagi, I. Matrix Metalloproteinases: The Sculptors of Chronic Cutaneous Wounds. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2220–2227. [Google Scholar] [CrossRef] [PubMed]
- Spampinato, S.F.; Caruso, G.I.; De Pasquale, R.; Sortino, M.A.; Merlo, S. The Treatment of Impaired Wound Healing in Diabetes: Looking among Old Drugs. Pharmaceuticals 2020, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Martino, C.F.; Perea, H.; Hopfner, U.; Ferguson, V.L.; Wintermantel, E. Effects of Weak Static Magnetic Fields on Endothelial Cells. Bioelectromagnetics 2010, 31, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive Oxygen Species (ROS) and Wound Healing: The Functional Role of ROS and Emerging ROS-Modulating Technologies for Augmentation of the Healing Process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Van Huizen, A.V.; Morton, J.M.; Kinsey, L.J.; Von Kannon, D.G.; Saad, M.A.; Birkholz, T.R.; Czajka, J.M.; Cyrus, J.; Barnes, F.S.; Beane, W.S. Weak Magnetic Fields Alter Stem Cell-Mediated Growth. Sci. Adv. 2019, 5, eaau7201. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X. Magnetic Fields and Reactive Oxygen Species. Int. J. Mol. Sci. 2017, 18, 2175. [Google Scholar] [CrossRef] [PubMed]
- Poniedziałek, B.; Rzymski, P.; Karczewski, J.; Jaroszyk, F.; Wiktorowicz, K. Reactive Oxygen Species (ROS) Production in Human Peripheral Blood Neutrophils Exposed in Vitro to Static Magnetic Field. Electromagn. Biol. Med. 2013, 32, 560–568. [Google Scholar] [CrossRef]
- Castello, P.R.; Hill, I.; Sivo, F.; Portelli, L.; Barnes, F.; Usselman, R.; Martino, C.F. Inhibition of Cellular Proliferation and Enhancement of Hydrogen Peroxide Production in Fibrosarcoma Cell Line by Weak Radio Frequency Magnetic Fields. Bioelectromagnetics 2014, 35, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Gurhan, H.; Barnes, F. Impact of Weak Radiofrequency and Static Magnetic Fields on Key Signaling Molecules, Intracellular PH, Membrane Potential, and Cell Growth in HT-1080 Fibrosarcoma Cells. Sci. Rep. 2023, 13, 14223. [Google Scholar] [CrossRef] [PubMed]
- Romeo, S.; Sannino, A.; Scarfì, M.R.; Massa, R.; Dangelo, R.; Zeni, O. Lack of Effects on Key Cellular Parameters of MRC-5 Human Lung Fibroblasts Exposed to 370 MT Static Magnetic Field. Sci. Rep. 2016, 6, 19398. [Google Scholar] [CrossRef]
- Saliev, T.; Mustapova, Z.; Kulsharova, G.; Bulanin, D.; Mikhalovsky, S. Therapeutic Potential of Electromagnetic Fields for Tissue Engineering and Wound Healing. Cell Prolif. 2014, 47, 485–493. [Google Scholar] [CrossRef]
- Haberal, M.; Aydogan, C.; Balcik, C.; Ekici, Y.; Haberal, N.; Kirnap, M.; Moray, G. Effect of Static Magnetic Field on Experimental Dermal Wound Strength. Indian J. Plast. Surg. 2012, 45, 215–219. [Google Scholar] [CrossRef]
- Henry, S.L.; Concannon, M.J.; Yee, G.J. The Effect of Magnetic Fields on Wound Healing: Experimental Study and Review of the Literature. Eplasty 2008, 8, e40. [Google Scholar]
- Feng, C.; Yu, B.; Song, C.; Wang, J.; Zhang, L.; Ji, X.; Wang, Y.; Fang, Y.; Liao, Z.; Wei, M.; et al. Static Magnetic Fields Reduce Oxidative Stress to Improve Wound Healing and Alleviate Diabetic Complications. Cells 2022, 11, 443. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Salazar, M.; Gonzalez-Galofre, Z.N.; Casamitjana, J.; Crisan, M.; James, A.W.; Péault, B. Five Decades Later, Are Mesenchymal Stem Cells Still Relevant? Front. Bioeng. Biotechnol. 2020, 8, 148. [Google Scholar] [CrossRef]
- Musiał-Wysocka, A.; Kot, M.; Majka, M. The Pros and Cons of Mesenchymal Stem Cell-Based Therapies. Cell Transplant. 2019, 28, 801–812. [Google Scholar] [CrossRef]
- Mikhaylov, G.; Vasiljeva, O. Promising Approaches in Using Magnetic Nanoparticles in Oncology. Biol. Chem. 2011, 392, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.R.; Davis, T.P.; Glover, A.L.; Nikles, D.E.; Brazel, C.S. Impact of Magnetic Field Parameters and Iron Oxide Nanoparticle Properties on Heat Generation for Use in Magnetic Hyperthermia. J. Magn. Magn. Mater. 2015, 387, 96–106. [Google Scholar] [CrossRef]
- Kobayashi, T. Cancer Hyperthermia Using Magnetic Nanoparticles. Biotechnol. J. 2011, 6, 1342–1347. [Google Scholar] [CrossRef]
- Dürr, S.; Janko, C.; Lyer, S.; Tripal, P.; Schwarz, M.; Zaloga, J.; Tietze, R.; Alexiou, C. Magnetic Nanoparticles for Cancer Therapy. Nanotechnol. Rev. 2013, 2, 395–409. [Google Scholar] [CrossRef]
- Tao, T.; Li, Z.; Xu, S.; Rehman, S.U.; Chen, R.; Xu, H.; Xia, H.; Zhang, J.; Zhao, H.; Wang, J.; et al. Boosting SARS-CoV-2 Enrichment with Ultrasmall Immunomagnetic Beads Featuring Superior Magnetic Moment. Anal. Chem. 2023, 95, 11542–11549. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.; Webster, T.J. Magnetic Nanoparticles: Biomedical Applications and Challenges. J. Mater. Chem. 2010, 20, 8760–8767. [Google Scholar] [CrossRef]
- Monzel, C.; Vicario, C.; Piehler, J.; Coppey, M.; Dahan, M. Magnetic Control of Cellular Processes Using Biofunctional Nanoparticles. Chem. Sci. 2017, 8, 7330–7338. [Google Scholar] [CrossRef]
- Seo, D.; Southard, K.M.; Kim, J.W.; Lee, H.J.; Farlow, J.; Lee, J.U.; Litt, D.B.; Haas, T.; Alivisatos, A.P.; Cheon, J.; et al. A Mechanogenetic Toolkit for Interrogating Cell Signaling in Space and Time. Cell 2016, 165, 1507–1518. [Google Scholar] [CrossRef]
- Bharde, A.A.; Palankar, R.; Fritsch, C.; Klaver, A.; Kanger, J.S.; Jovin, T.M.; Arndt-Jovin, D.J. Magnetic Nanoparticles as Mediators of Ligand-Free Activation of EGFR Signaling. PLoS ONE 2013, 8, e68879. [Google Scholar] [CrossRef]
- Wee, P.; Wang, Z. Epidermal Growth Factor Receptor Cell Proliferation Signaling Pathways. Cancers 2017, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.H.; Lee, E.J.; Son, M.; Lee, J.H.; Yoo, D.; Kim, J.W.; Park, S.W.; Shin, J.S.; Cheon, J. A Magnetic Switch for the Control of Cell Death Signalling in in Vitro and in Vivo Systems. Nat. Mater. 2012, 11, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhang, Q.; Hu, H.; Yang, F.; Li, K.; Zhang, Y.; Shi, C. Enhancing Cellular Morphological Changes and Ablation of Cancer Cells via the Interaction of Drug Co-Loaded Magnetic Nanosystems in Weak Rotating Magnetic Fields. RSC Adv. 2020, 10, 14471–14481. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Magnetic Field Parameters | Cell Line | Biological Effects | Ref. |
|---|---|---|---|
| 50 Hz, 1 mT for 1 h | HaCaT keratinocyte | Induced proliferation via mTOR (PI3K/Akt) and ERK signaling pathways | [2] |
| 60 Hz, 1.5 mT for 144 h | HaCaT keratinocyte | Inhibited cell growth, activated ATM-Chk2-p21 pathway (G1 arrest) | [3] |
| 60 Hz, 6 mT for 30 min every 24 h for 3 days | IMR90, HeLa | Induced DNA double-strand breaks (DSBs) and apoptosis via p38 activation; increased micronucleus formation and chromosomal aberrations. | [4] |
| 60 Hz, 7 mT for 10–60 min | IMR90, HeLa | Induced DNA DSBs without apoptosis, activated DNA damage checkpoint without ROS production; increased DNA repair foci formation | [5] |
| 50 Hz, 1 mT for 24/48/72 h | SH-SY5Y | Significantly increased ROS levels | [6] |
| 2 ± 0.2 mT; 75 ± 2 Hz for 10 min, 4 times/week | SH-SY5Y | Decreased H2O2-induced ROS | [7] |
| 30 Hz, 0.8 mT for 1 h | 6B 1 hybridoma | Inhibited proliferation; variable effects at different frequencies | [8] |
| 60 Hz, 20, and 200 μT for various times up to 30 h | Human fibroblasts | Increased G1 phase length at lower intensities, no effect at higher flux densities | [9] |
| 50 Hz, 100 μT, 0.6 mT, 24 h, 48 h | K562, DU145 cells | Significantly increased cell proliferation; micronucleus formation observed | [10,11] |
| 50, 60 Hz, 2, 20, 100, 500 μT, 24 h, 48 h | K562 cells | No effect on cell proliferation | [12] |
| (104–105) Tm−1, static | Cancer cells enriched by Fe | Tumor arrest | [13] |
| (103–105) Tm−1, static | HeLa cells, other cancerous cells with low membrane tension | Magnetically assisted cell division | [13] |
| (102–103) Tm−1, static | PC-3 cells and fibroblasts | Magnetically assisted endocytosis | [14] |
| 103 Tm−1, static | THP-1 cells | Cell swelling | [15] |
| 50 Hz, 1 mT for 12 h/day for 3 days | Human lymphocytes | Increased ROS production and DNA fragmentation; increased micronucleus frequency and chromosomal aberrations | [16] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tota, M.; Jonderko, L.; Witek, J.; Novickij, V.; Kulbacka, J. Cellular and Molecular Effects of Magnetic Fields. Int. J. Mol. Sci. 2024, 25, 8973. https://doi.org/10.3390/ijms25168973
Tota M, Jonderko L, Witek J, Novickij V, Kulbacka J. Cellular and Molecular Effects of Magnetic Fields. International Journal of Molecular Sciences. 2024; 25(16):8973. https://doi.org/10.3390/ijms25168973
Chicago/Turabian StyleTota, Maciej, Laura Jonderko, Julia Witek, Vitalij Novickij, and Julita Kulbacka. 2024. "Cellular and Molecular Effects of Magnetic Fields" International Journal of Molecular Sciences 25, no. 16: 8973. https://doi.org/10.3390/ijms25168973
APA StyleTota, M., Jonderko, L., Witek, J., Novickij, V., & Kulbacka, J. (2024). Cellular and Molecular Effects of Magnetic Fields. International Journal of Molecular Sciences, 25(16), 8973. https://doi.org/10.3390/ijms25168973