
Mesenchymal Stem Cells from Mouse Hair Follicles Inhibit the Development of Type 1 Diabetes
 , , , ,
, , , ,  and
and
Abstract

1. Introduction
2. Results
2.1. moMSCORS Inhibit T Cell Proliferation and Activation In Vitro
2.2. moMSCORS Ameliorates T1D in Mice
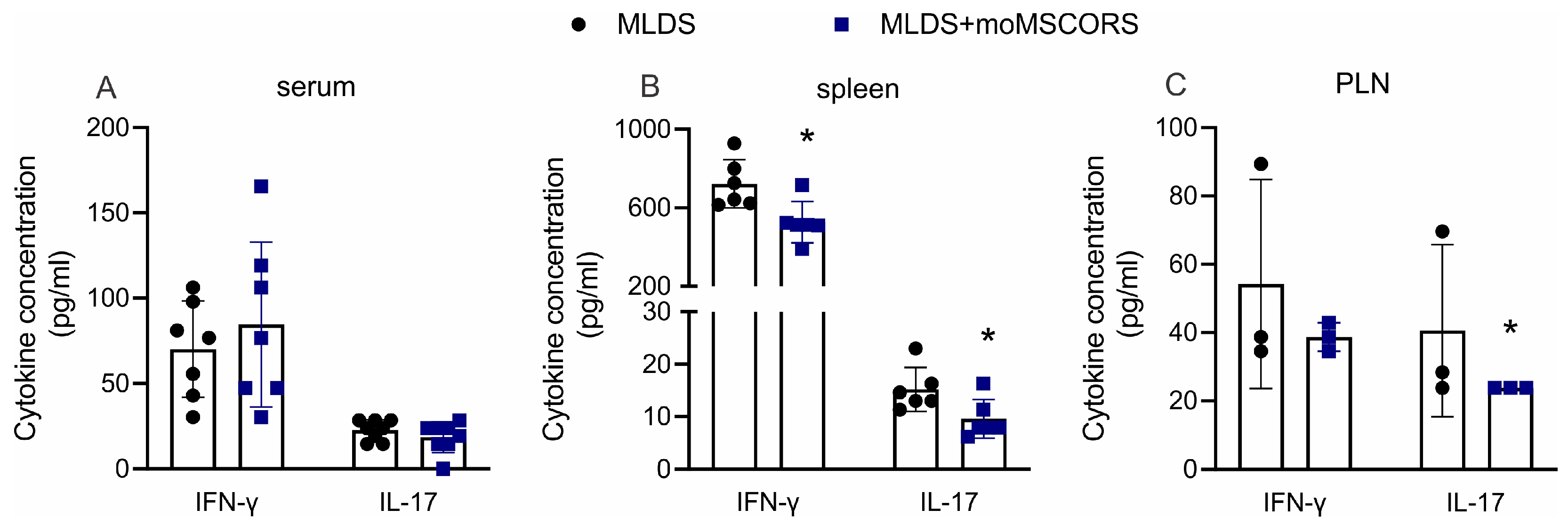
2.3. moMSCORS Treatment Decreases the Production of Pro-Inflammatory Cytokines
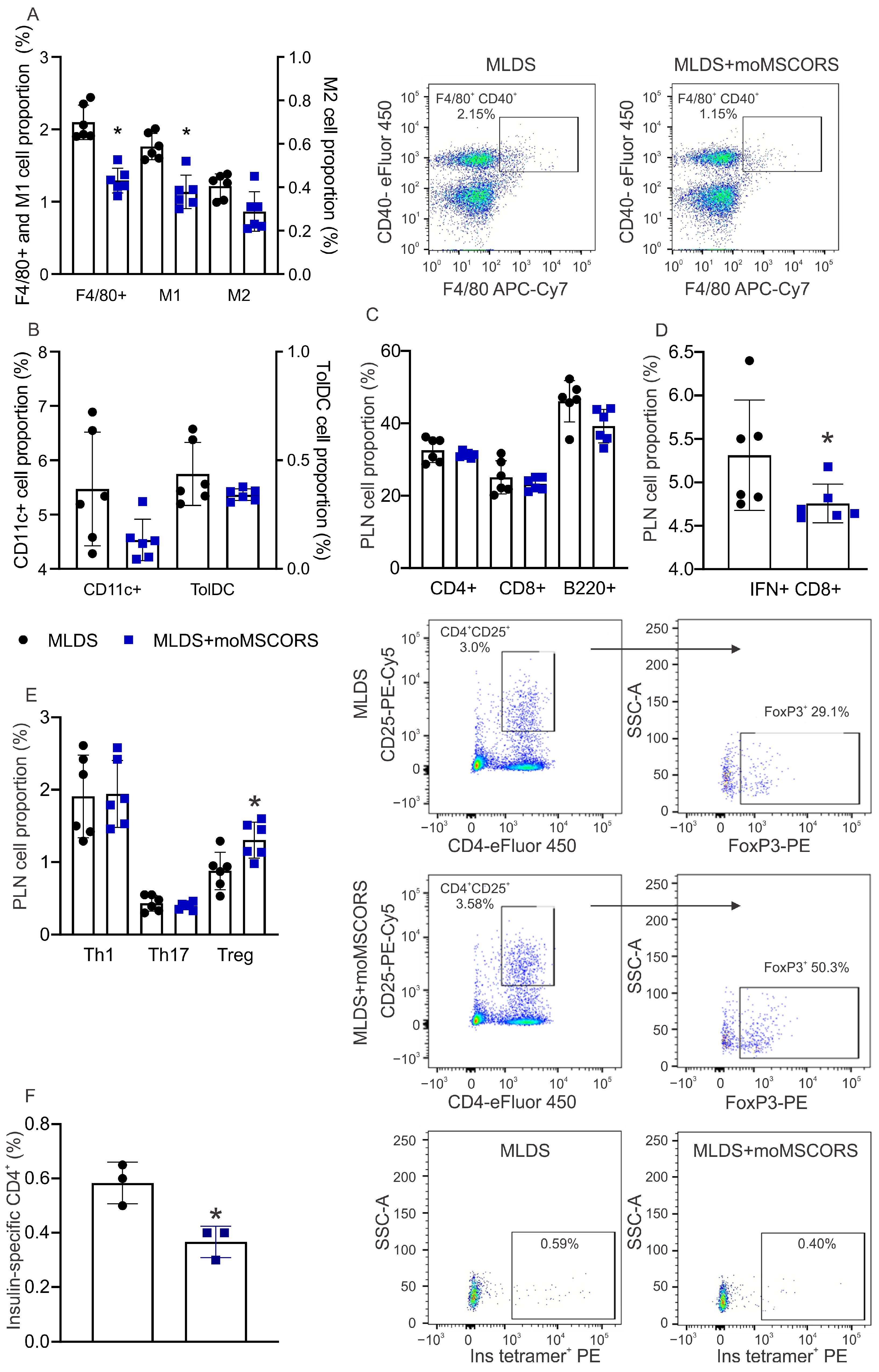
2.4. moMSCORS Treatment Affects the Proportion of Immune Cells in the Draining Lymph Node
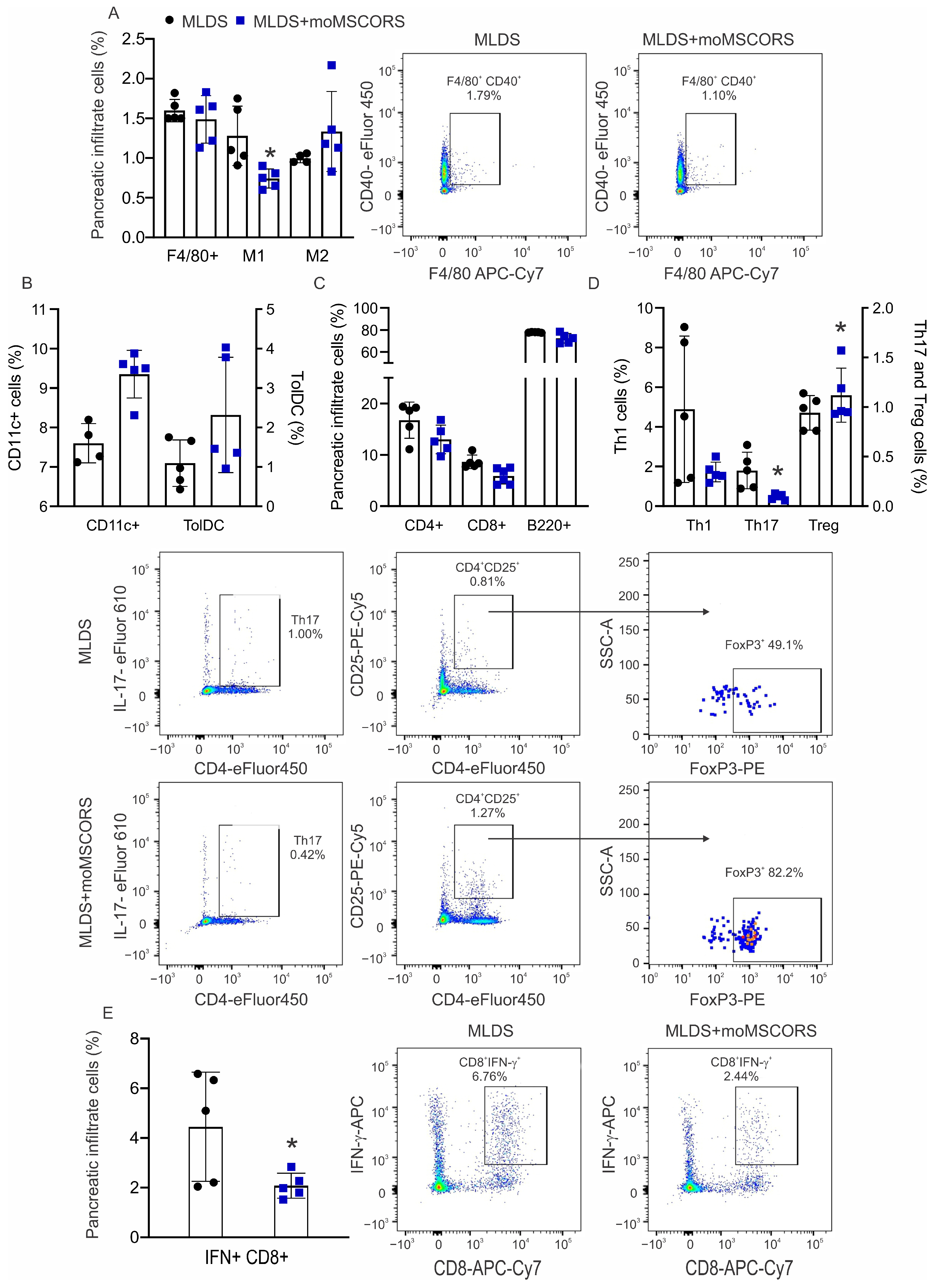
2.5. moMSCORS Treatment Affects the Proportion of Immune Cells in the Pancreas
3. Discussion
4. Materials and Methods
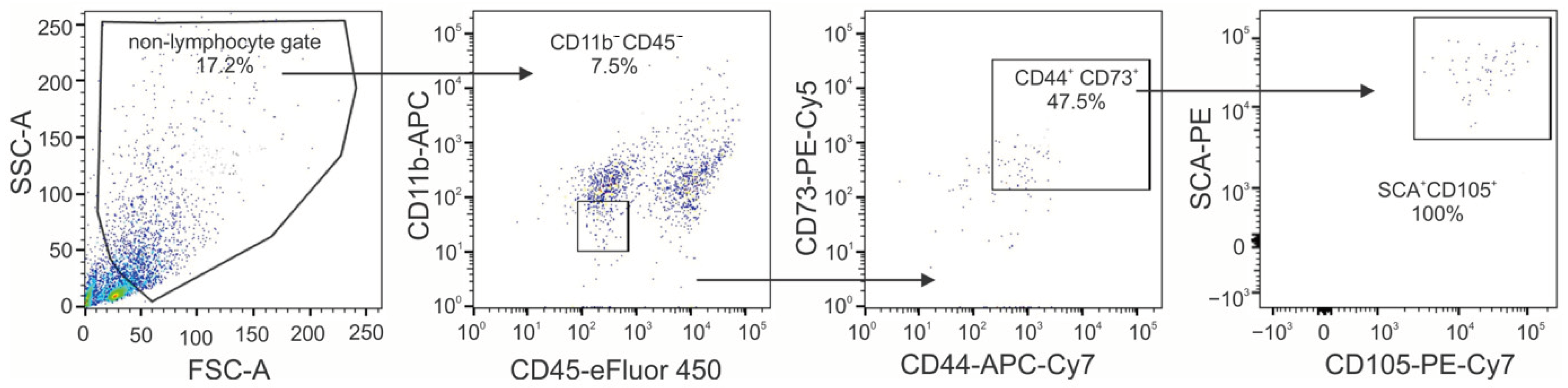
4.1. Isolation of Mesenchymal Stem Cells from Mouse Whisker Hair Follicle Outer Root Sheath (moMSCORS)
4.2. T1D Induction and moMSCORS Treatment
4.3. Histological Analysis
4.4. Cell Isolation
4.5. In Vitro Suppression Assay
4.6. Detection of Extracellular and Intracellular Markers Using Flow Cytometry
4.7. ELISA
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katsarou, A.; Gudbjörnsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, Å. Type 1 diabetes mellitus. Nat. Rev. Dis. Primers 2017, 3, 17016. [Google Scholar] [CrossRef] [PubMed]
- Willcox, A.; Richardson, S.J.; Bone, A.J.; Foulis, A.K.; Morgan, N.G. Analysis of islet inflammation in human type 1 diabetes. Clin. Exp. Immunol. 2009, 155, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.A.; Wong, F.S.; Wen, L. The importance of the Non Obese Diabetic (NOD) mouse model in autoimmune diabetes. J. Autoimmun. 2016, 66, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Radenković, M.; Stojanović, M.; Prostran, M. Experimental diabetes induced by alloxan and streptozotocin: The current state of the art. J. Pharmacol. Toxicol. Methods 2016, 78, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.C.; Shyu, W.C.; Lin, S.Z. Mesenchymal stem cells. Cell Transplant. 2011, 20, 5–14. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Ouryazdanpanah, N.; Dabiri, S.; Derakhshani, A.; Vahidi, R.; Farsinejad, A. Peripheral Blood-Derived Mesenchymal Stem Cells: Growth Factor-Free Isolation, Molecular Characterization and Differentiation. Iran. J. Pathol. 2018, 13, 461–466. [Google Scholar]
- Reh, T.A.; Fischer, A.J. Retinal stem cells. Methods Enzymol. 2006, 419, 52–73. [Google Scholar] [CrossRef] [PubMed]
- Witkowska-Zimny, M.; Wrobel, E. Perinatal sources of mesenchymal stem cells: Wharton’s jelly, amnion, and chorion. Cell. Mol. Biol. Lett. 2011, 16, 493–514. [Google Scholar] [CrossRef]
- Torre, P.; Flores, A.I. Current Status and Future Prospects of Perinatal Stem Cells. Genes 2020, 12, 6. [Google Scholar] [CrossRef]
- Schosserer, M.; Reynoso, R.; Wally, V.; Jug, B.; Kantner, V.; Weilner, S.; Buric, I.; Grillari, J.; Bauer, J.W.; Grillari-Voglauer, R. Urine is a novel source of autologous mesenchymal stem cells for patients with epidermolysis bullosa. BMC Res. Notes 2015, 8, 767. [Google Scholar] [CrossRef] [PubMed]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, e264–e277. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Scholtemeijer, M.; Shah, K. Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential. Trends Pharmacol. Sci. 2020, 41, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Huang, W.; Liu, J.; Tian, J.; Wang, S.; Rui, K. Effects of Mesenchymal Stem Cell-Derived Exosomes on Autoimmune Diseases. Front. Immunol. 2021, 12, 749192. [Google Scholar] [CrossRef] [PubMed]
- Nagy, G.; Szekely, T.E.; Somogyi, A.; Herold, M.; Herold, Z. New therapeutic approaches for type 1 diabetes: Disease-modifying therapies. World J. Diabetes 2022, 13, 835–850. [Google Scholar] [CrossRef]
- Kawada-Horitani, E.; Kita, S.; Okita, T.; Nakamura, Y.; Nishida, H.; Honma, Y.; Fukuda, S.; Tsugawa-Shimizu, Y.; Kozawa, J.; Sakaue, T.; et al. Human adipose-derived mesenchymal stem cells prevent type 1 diabetes induced by immune checkpoint blockade. Diabetologia 2022, 65, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Bassi, Ê.J.; Moraes-Vieira, P.M.; Moreira-Sá, C.S.; Almeida, D.C.; Vieira, L.M.; Cunha, C.S.; Hiyane, M.I.; Basso, A.S.; Pacheco-Silva, A.; Câmara, N.O. Immune regulatory properties of allogeneic adipose-derived mesenchymal stem cells in the treatment of experimental autoimmune diabetes. Diabetes 2012, 61, 2534–2545. [Google Scholar] [CrossRef] [PubMed]
- Shigemoto-Kuroda, T.; Oh, J.Y.; Kim, D.K.; Jeong, H.J.; Park, S.Y.; Lee, H.J.; Park, J.W.; Kim, T.W.; An, S.Y.; Prockop, D.J.; et al. MSC-derived Extracellular Vesicles Attenuate Immune Responses in Two Autoimmune Murine Models: Type 1 Diabetes and Uveoretinitis. Stem Cell Rep. 2017, 8, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Montanucci, P.; Pescara, T.; Alunno, A.; Bistoni, O.; Basta, G.; Calafiore, R. Remission of hyperglycemia in spontaneously diabetic NOD mice upon transplant of microencapsulated human umbilical cord Wharton jelly-derived mesenchymal stem cells (hUCMS). Xenotransplantation 2019, 26, e12476. [Google Scholar] [CrossRef]
- Savkovic, V.; Li, H.; Simon, J.C.; Etz Schneider, M. Method of Culturing Mesenchymal Stem Cells. U.S. Patent No. PCT/EP2020/070027, 13 July 2019. [Google Scholar]
- Li, H.; Masieri, F.F.; Schneider, M.; Kottek, T.; Hahnel, S.; Yamauchi, K.; Obradović, D.; Seon, J.K.; Yun, S.J.; Ferrer, R.A.; et al. Autologous, Non-Invasively Available Mesenchymal Stem Cells from the Outer Root Sheath of Hair Follicle Are Obtainable by Migration from Plucked Hair Follicles and Expandable in Scalable Amounts. Cells 2020, 9, 2069. [Google Scholar] [CrossRef]
- Li, C.; Zhao, H.; Cheng, L.; Wang, B. Allogeneic vs. autologous mesenchymal stem/stromal cells in their medication practice. Cell Biosci. 2021, 11, 187. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ziemer, M.; Stojanovic, I.; Saksida, T.; Maksimovic-Ivanic, D.; Mijatovic, S.; Djmura, G.; Gajic, D.; Koprivica, I.; Krajnovic, T.; et al. Mesenchymal Stem Cells From Mouse Hair Follicles Reduce Hypertrophic Scarring in a Murine Wound Healing Model. Stem Cell Rev. Rep. 2022, 18, 2028–2044. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xiong, S.; Masieri, F.F.; Monika, S.; Lethaus, B.; Savkovic, V. Mesenchymal Stem Cells Isolated from Equine Hair Follicles Using a Method of Air-Liquid Interface. Stem Cell Rev Rep. 2023, 19, 2943–2956. [Google Scholar] [CrossRef] [PubMed]
- Kulus, M.; Sibiak, R.; Stefańska, K.; Zdun, M.; Wieczorkiewicz, M.; Piotrowska-Kempisty, H.; Jaśkowski, J.M.; Bukowska, D.; Ratajczak, K.; Zabel, M.; et al. Mesenchymal Stem/Stromal Cells Derived from Human and Animal Perinatal Tissues-Origins, Characteristics, Signaling Pathways, and Clinical Trials. Cells 2021, 10, 3278. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Tammik, C.; Rosendahl, K.; Zetterberg, E.; Ringdén, O. HLA expression and immunologic properties of differentiated and undifferentiated mesenchymal stem cells. Exp. Hematol. 2003, 31, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Djouad, F.; Charbonnier, L.M.; Bouffi, C.; Louis-Plence, P.; Bony, C.; Apparailly, F.; Cantos, C.; Jorgensen, C.; Noël, D. Mesenchymal stem cells inhibit the differentiation of dendritic cells through an interleukin-6-dependent mechanism. Stem Cells 2007, 25, 2025–2032. [Google Scholar] [CrossRef] [PubMed]
- Tatara, R.; Ozaki, K.; Kikuchi, Y.; Hatanaka, K.; Oh, I.; Meguro, A.; Matsu, H.; Sato, K.; Ozawa, K. Mesenchymal stromal cells inhibit Th17 but not regulatory T-cell differentiation. Cytotherapy 2011, 13, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Shin, J.S.; Kim, Y.H.; Hong, S.H.; Yang, S.H.; Shin, J.Y.; Kim, S.Y.; Kim, B.; Kim, J.S.; Park, C.G. Murine mesenchymal stem cells suppress T lymphocyte activation through IL-2 receptor α (CD25) cleavage by producing matrix metalloproteinases. Stem Cell Rev. Rep. 2011, 7, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.S.; Lee, K.; Na, K.; Zhang, Y.X.; Lim, H.J.; Yi, T.; Song, S.U.; Jeon, M.S. Mesenchymal stromal cells inhibit CD25 expression via the mTOR pathway to potentiate T-cell suppression. Cell Death Dis. 2017, 8, e2632. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.; Wang, F.; Wang, L.; Yu, X.; Sun, R.; Wang, Z.; Wang, L.; Gao, H.; Fu, Z.; et al. Effect and mechanisms of human Wharton’s jelly-derived mesenchymal stem cells on type 1 diabetes in NOD model. Endocrine 2015, 48, 124–134. [Google Scholar] [CrossRef]
- Jurewicz, M.; Yang, S.; Augello, A.; Godwin, J.G.; Moore, R.F.; Azzi, J.; Fiorina, P.; Atkinson, M.; Sayegh, M.H.; Abdi, R. Congenic mesenchymal stem cell therapy reverses hyperglycemia in experimental type 1 diabetes. Diabetes 2010, 59, 3139–3147. [Google Scholar] [CrossRef] [PubMed]
- Yaochite, J.N.; Caliari-Oliveira, C.; de Souza, L.E.; Neto, L.S.; Palma, P.V.; Covas, D.T.; Malmegrim, K.C.; Voltarelli, J.C.; Donadi, E.A. Therapeutic efficacy and biodistribution of allogeneic mesenchymal stem cells delivered by intrasplenic and intrapancreatic routes in streptozotocin-induced diabetic mice. Stem Cell Res. Ther. 2015, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Gou, W.; Hua, W.; Swaby, L.; Cui, W.; Green, E.; Morgan, K.A.; Strange, C.; Wang, H. Stem Cell Therapy Improves Human Islet Graft Survival in Mice via Regulation of Macrophages. Diabetes 2022, 71, 2642–2655. [Google Scholar] [CrossRef] [PubMed]
- Ghannam, S.; Pène, J.; Moquet-Torcy, G.; Jorgensen, C.; Yssel, H. Mesenchymal stem cells inhibit human Th17 cell differentiation and function and induce a T regulatory cell phenotype. J. Immunol. 2010, 185, 302–312. [Google Scholar] [CrossRef]
- Luz-Crawford, P.; Noël, D.; Fernandez, X.; Khoury, M.; Figueroa, F.; Carrión, F.; Jorgensen, C.; Djouad, F. Mesenchymal stem cells repress Th17 molecular program through the PD-1 pathway. PLoS ONE 2012, 7, e45272. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Sun, X.; Kuang, X.; Liao, Y.; Li, H.; Luo, D. Mesenchymal stem cells suppress CD8+ T cell-mediated activation by suppressing natural killer group 2, member D protein receptor expression and secretion of prostaglandin E2, indoleamine 2, 3-dioxygenase and transforming growth factor-β. Clin. Exp. Immunol. 2014, 178, 516–524. [Google Scholar] [CrossRef]
- Zhang, W.; Ling, Q.; Wang, B.; Wang, K.; Pang, J.; Lu, J.; Bi, Y.; Zhu, D. Comparison of therapeutic effects of mesenchymal stem cells from umbilical cord and bone marrow in the treatment of type 1 diabetes. Stem Cell Res. Ther. 2022, 13, 406. [Google Scholar] [CrossRef]
- Chen, L.B.; Jiang, X.B.; Yang, L. Differentiation of rat marrow mesenchymal stem cells into pancreatic islet beta-cells. World J. Gastroenterol. 2004, 10, 3016–3020. [Google Scholar] [CrossRef]
- Kassem, D.H.; Kamal, M.M.; El-Kholy, A.E.L.G.; El-Mesallamy, H.O. Exendin-4 enhances the differentiation of Wharton’s jelly mesenchymal stem cells into insulin-producing cells through activation of various β-cell markers. Stem Cell Res. Ther. 2016, 7, 108. [Google Scholar] [CrossRef]
- Thorel, F.; Népote, V.; Avril, I.; Kohno, K.; Desgraz, R.; Chera, S.; Herrera, P.L. Conversion of adult pancreatic alpha-cells to beta-cells after extreme beta-cell loss. Nature 2010, 464, 1149–1154. [Google Scholar] [CrossRef]
- Chera, S.; Baronnier, D.; Ghila, L.; Cigliola, V.; Jensen, J.N.; Gu, G.; Furuyama, K.; Thorel, F.; Gribble, F.M.; Reimann, F.; et al. Diabetes recovery by age-dependent conversion of pancreatic δ-cells into insulin producers. Nature 2014, 514, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Chakravarthy, H.; Gu, X.; Enge, M.; Dai, X.; Wang, Y.; Damond, N.; Downie, C.; Liu, K.; Wang, J.; Xing, Y.; et al. Converting Adult Pancreatic Islet α Cells into β Cells by Targeting Both Dnmt1 and Arx. Cell Metab. 2017, 25, 622–634. [Google Scholar] [CrossRef] [PubMed]
- Aali, E.; Mirzamohammadi, S.; Ghaznavi, H.; Madjd, Z.; Larijani, B.; Rayegan, S.; Sharifi, A.M. A comparative study of mesenchymal stem cell transplantation with its paracrine effect on control of hyperglycemia in type 1 diabetic rats. J. Diabetes Metab. Disord. 2014, 13, 76. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, L.; Byrne, R.N.; van Haaften, T.; Vadivel, A.; Alphonse, R.S.; Rey-Parra, G.J.; Weissmann, G.; Hall, A.; Eaton, F.; Thébaud, B. Stem cell conditioned medium improves acute lung injury in mice: In vivo evidence for stem cell paracrine action. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, L967–L977. [Google Scholar] [CrossRef] [PubMed]
- Casellas, A.; Salavert, A.; Agudo, J.; Ayuso, E.; Jimenez, V.; Moya, M.; Muñoz, S.; Franckhauser, S.; Bosch, F. Expression of IGF-I in pancreatic islets prevents lymphocytic infiltration and protects mice from type 1 diabetes. Diabetes 2006, 55, 3246–3255. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghe, M.; Prathiraja, O.; Perera, P.B.; Jena, R.; Silva, M.S.; Weerawarna, P.S.H.; Singhal, M.; Kayani, A.M.A.; Karnakoti, S.; Jain, S. The Role of Mesenchymal Stem Cells in the Treatment of Type 1 Diabetes. Cureus 2022, 14, e27337. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Mahato, R.I. Mesenchymal stem cell-based therapy for type 1 diabetes. Discov. Med. 2014, 17, 139–143. [Google Scholar]
- Galipeau, J. The mesenchymal stromal cells dilemma--does a negative phase III trial of random donor mesenchymal stromal cells in steroid-resistant graft-versus-host disease represent a death knell or a bump in the road? Cytotherapy 2013, 15, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Isildar, B.; Ozkan, S.; Ercin, M.; Gezginci-Oktayoglu, S.; Oncul, M.; Koyuturk, M. 2D and 3D cultured human umbilical cord-derived mesenchymal stem cell-conditioned medium has a dual effect in type 1 diabetes model in rats: Immunomodulation and beta-cell regeneration. Inflamm. Regen. 2022, 42, 55. [Google Scholar] [CrossRef]
- Koprivica, I.; Gajic, D.; Saksida, T.; Cavalli, E.; Auci, D.; Despotovic, S.; Pejnovic, N.; Stosic-Grujicic, S.; Nicoletti, F.; Stojanovic, I. Orally delivered all-trans-retinoic acid- and transforming growth factor-β-loaded microparticles ameliorate type 1 diabetes in mice. Eur. J. Pharmacol. 2019, 864, 172721. [Google Scholar] [CrossRef]
- Koprivica, I.; Jonić, N.; Diamantis, D.; Gajić, D.; Saksida, T.; Pejnović, N.; Tzakos, A.G.; Stojanović, I. Phenethyl ester of rosmarinic acid attenuates autoimmune responses during type 1 diabetes development in mice. Life Sci. 2022, 288, 120184. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium for cell culture | RPMI 1640 1% Pen/Strep 2 mM L-glutamine 25 mM HEPES 5% FCS |
| Magnetic bead buffer | PBS 0.5% BSA 2.5 mM EDTA |
| T lymphocyte medium | RPMI 1640 1% Pen/Strep 0.02 mM Na-pyruvate 5 μM β-mercaptoethanol 2 mM L-glutamine 25 mM HEPES 10% FCS |
| Antibody | Fluorophore | Host | Manufacturer |
|---|---|---|---|
| CD45 | eFluor™506 | rat IgG2b, κ | Invitrogen, Waltham, MA, USA |
| CD4 | eFluor™450 | rat IgG2a, κ | |
| CD8 | APC-eFluor™780 | rat IgG2a, κ | |
| CD25 | PE | rat IgG1, λ | |
| CD25 | PE-Cy5.5 | rat IgG1, λ | |
| B220 | Alexa Fluor™700 | rat IgG2a, κ | |
| F4/80 | APC-eFluor™780 | rat IgG2a, κ | |
| CD40 | eFluor™450 | Armenian hamster IgM, κ | |
| CD206 | eFluor™660 | rat IgG2b, κ | |
| CD11b | PE | rat IgG2b, κ | |
| CD11b | APC | rat IgG2b, κ | |
| CD11c | PE-Cy5 | Armenian hamster IgG | |
| CD90.2 | FITC | rat IgG2a, κ | |
| CD73 | PerCP-eFluor™710 | rat IgG1 | |
| CD44 | APC-eFluor™780 | rat IgG2b, κ | |
| CD103 | PE/Dazzle™594 | Armenian hamster IgG | BioLegend, San Diego, CA, USA |
| CD105 | PE-Cy7 | rat IgG2a, κ | |
| Sca-1 | PE | rat IgG2a, κ |
| Antibody | Fluorophore | Host | Manufacturer |
|---|---|---|---|
| IFN-γ | APC | rat IgG1, κ | Invitrogen, Waltham, MA, USA |
| IL-17 | eFluor™610 | rat IgG1, κ | |
| IL-10 | FITC | rat IgG2b, κ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mićanović, D.; Stanisavljević, S.; Li, H.; Koprivica, I.; Jonić, N.; Stojanović, I.; Savković, V.; Saksida, T. Mesenchymal Stem Cells from Mouse Hair Follicles Inhibit the Development of Type 1 Diabetes. Int. J. Mol. Sci. 2024, 25, 5974. https://doi.org/10.3390/ijms25115974
Mićanović D, Stanisavljević S, Li H, Koprivica I, Jonić N, Stojanović I, Savković V, Saksida T. Mesenchymal Stem Cells from Mouse Hair Follicles Inhibit the Development of Type 1 Diabetes. International Journal of Molecular Sciences. 2024; 25(11):5974. https://doi.org/10.3390/ijms25115974
Chicago/Turabian StyleMićanović, Dragica, Suzana Stanisavljević, Hanluo Li, Ivan Koprivica, Natalija Jonić, Ivana Stojanović, Vuk Savković, and Tamara Saksida. 2024. "Mesenchymal Stem Cells from Mouse Hair Follicles Inhibit the Development of Type 1 Diabetes" International Journal of Molecular Sciences 25, no. 11: 5974. https://doi.org/10.3390/ijms25115974
APA StyleMićanović, D., Stanisavljević, S., Li, H., Koprivica, I., Jonić, N., Stojanović, I., Savković, V., & Saksida, T. (2024). Mesenchymal Stem Cells from Mouse Hair Follicles Inhibit the Development of Type 1 Diabetes. International Journal of Molecular Sciences, 25(11), 5974. https://doi.org/10.3390/ijms25115974