
A Dual Strategy—In Vitro and In Silico—To Evaluate Human Antitetanus mAbs Addressing Their Potential Protective Action on TeNT Endocytosis in Primary Rat Neuronal Cells

 , , , and
, , , and 
Abstract
1. Introduction
2. Results
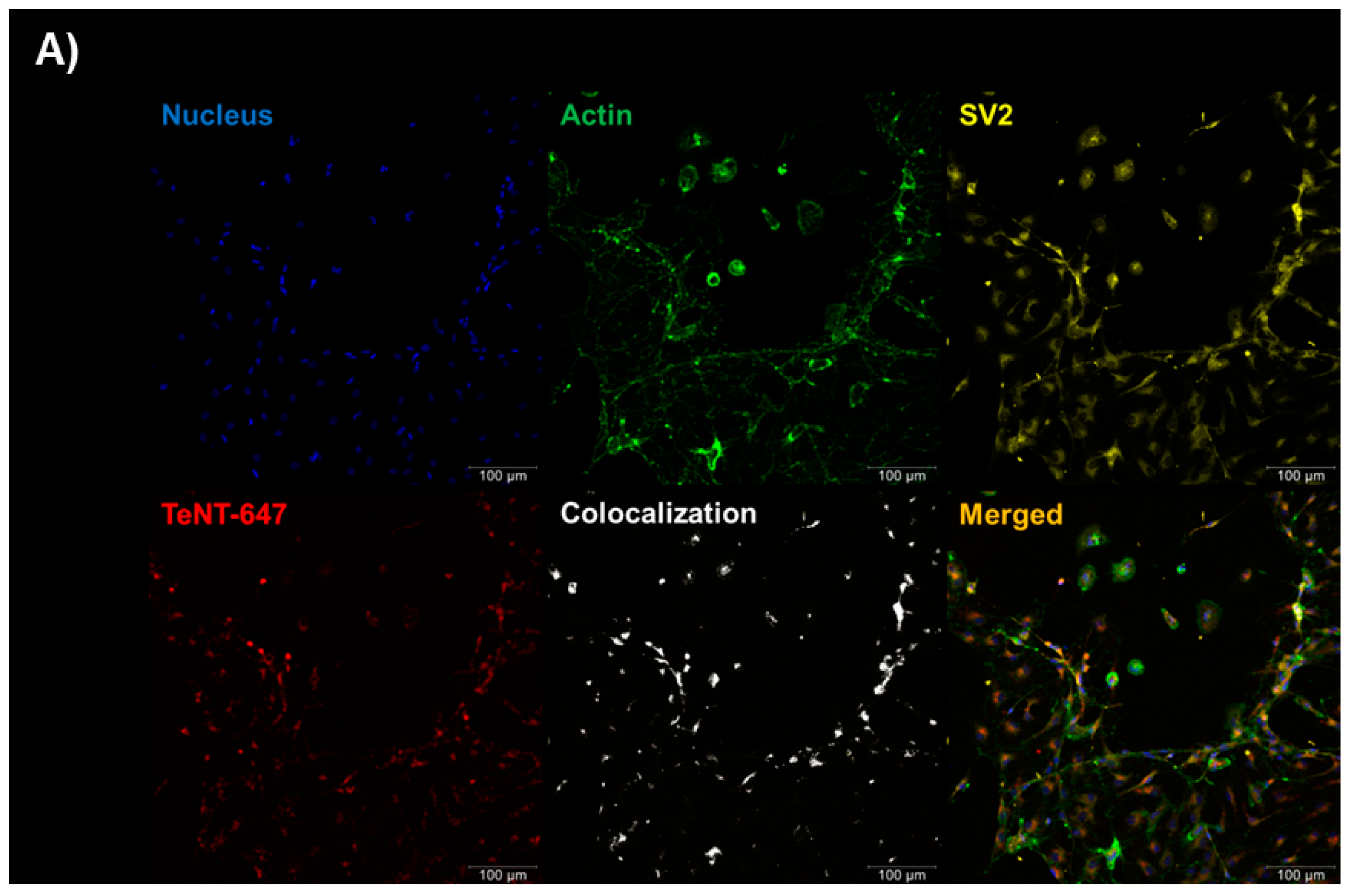
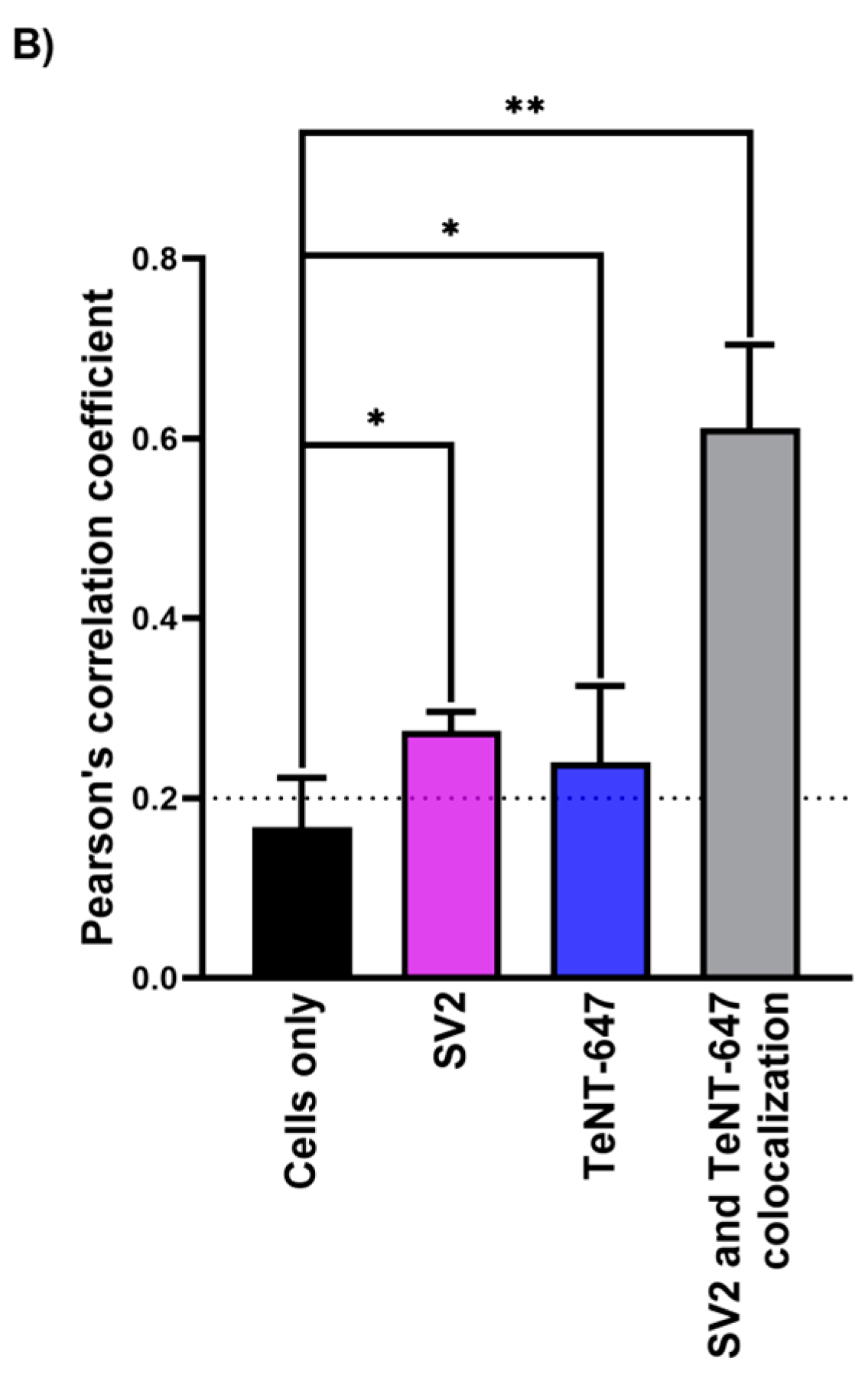
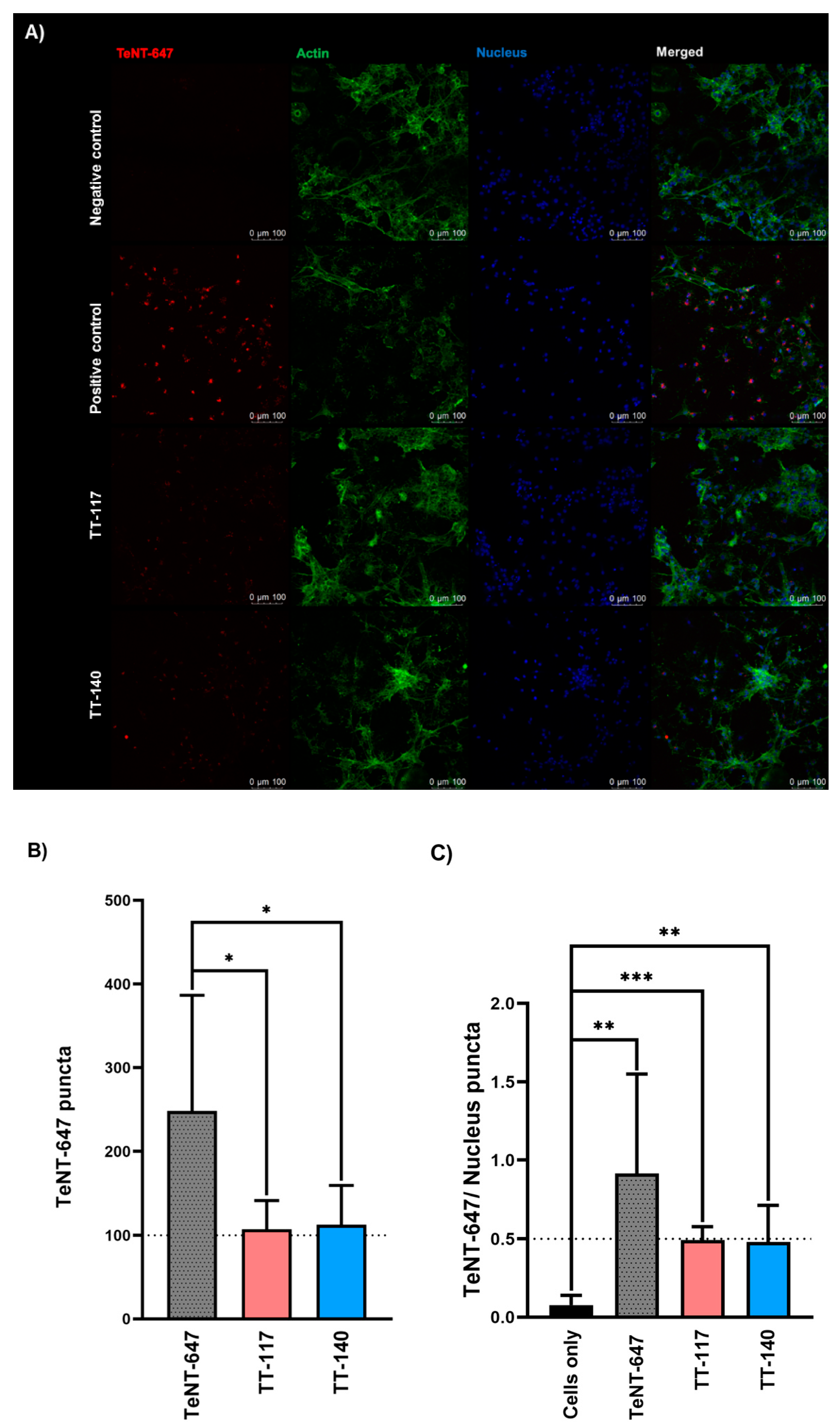
2.1. Tetanus Toxin Internalization Mediated by SV2 in RSpN Culture
2.2. Tetanus Toxin Internalization Inhibition by mAbs
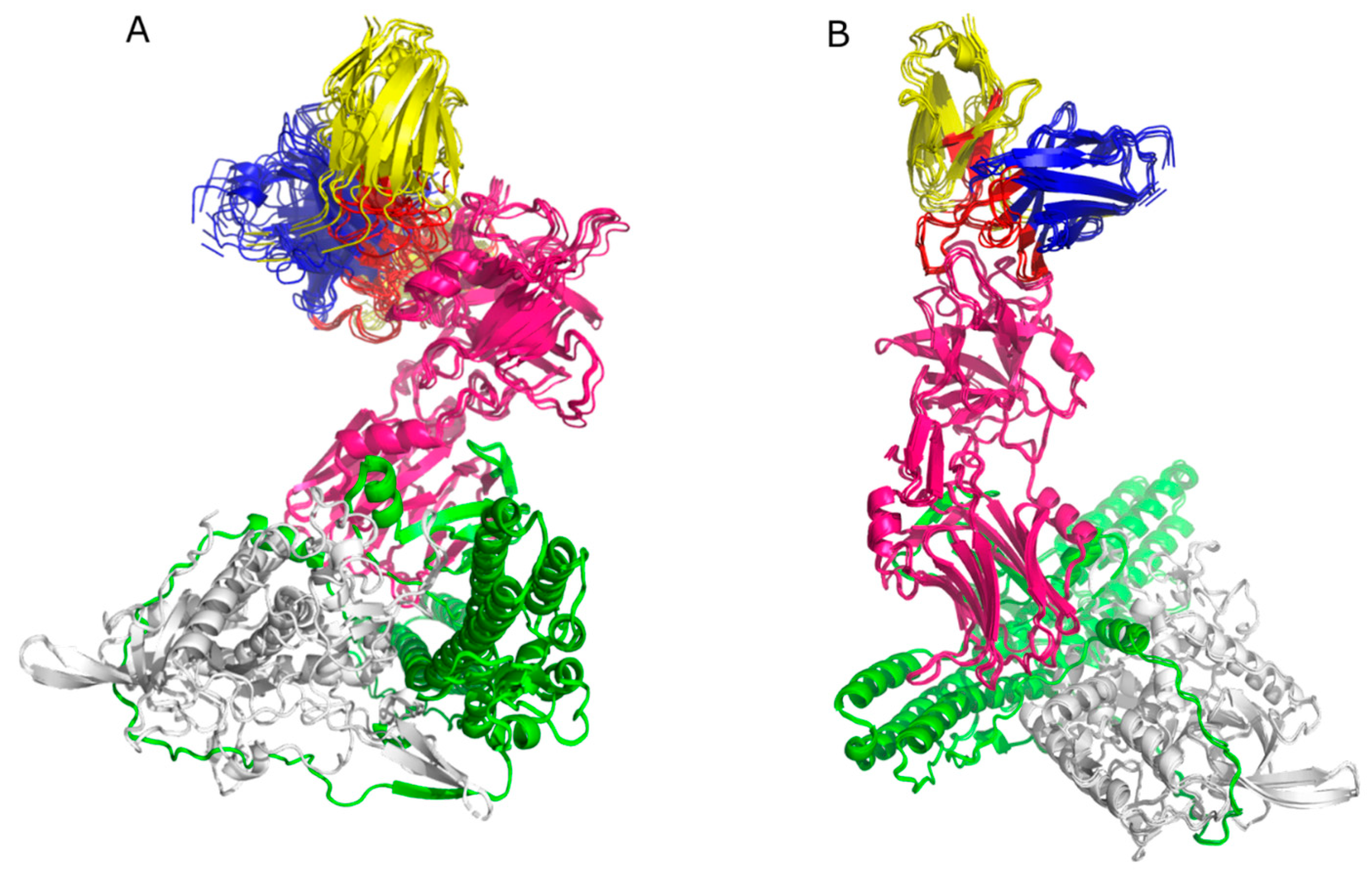
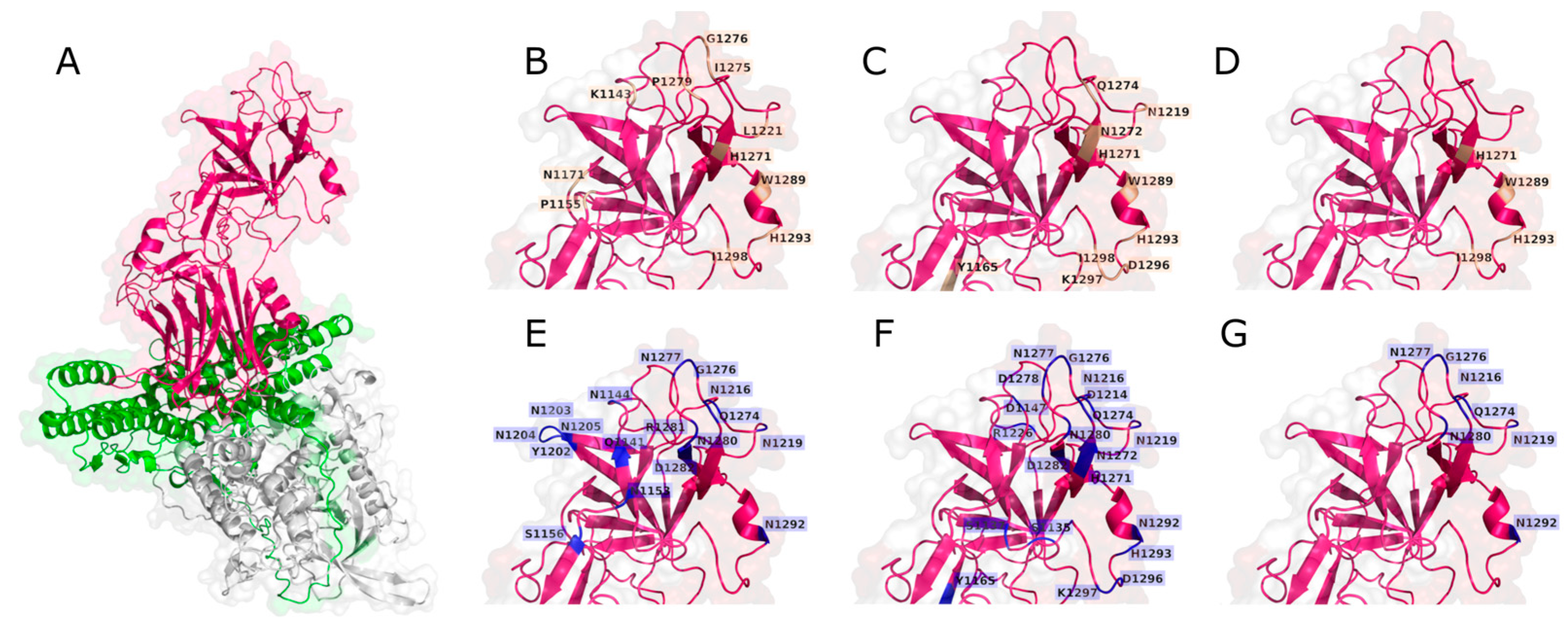
2.3. Molecular Docking
2.4. Epitopes Prediction
3. Discussion
4. Materials and Methods
4.1. Tetanus Toxin Labeling
4.2. Isolation and Cell Culture of Rat Spinal Cord Neurons (RSpN)
4.3. Tetanus Toxin Internalization Inhibition Assay
4.4. Immunofluorescence
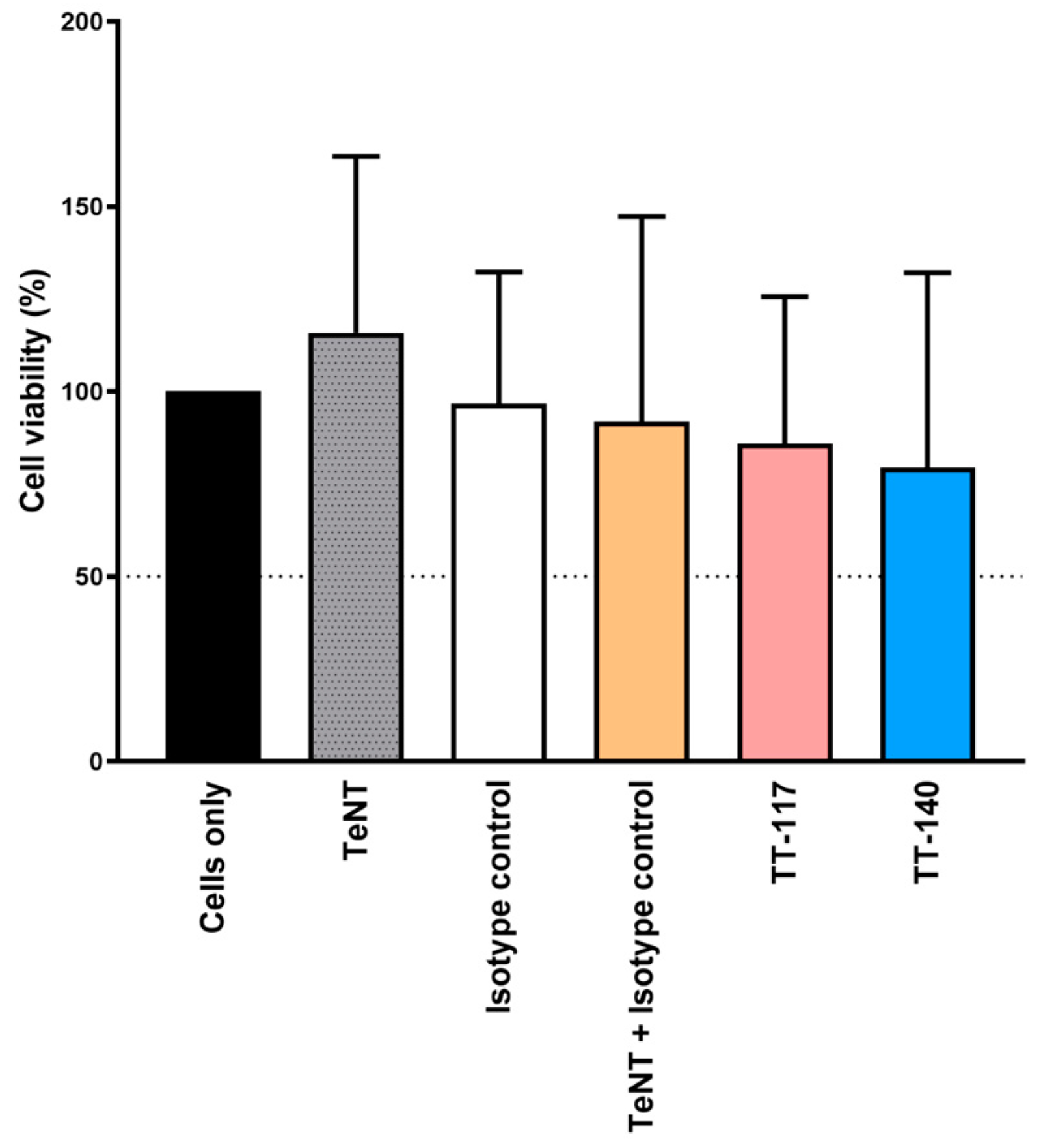
4.5. Statistical Analysis
4.6. Analysis of Antibody Sequences
4.7. Obtaining the 3D Structure of Antibodies
4.8. Molecular Docking by HADDOCK 2.4
4.9. Molecular Docking by SnugDock
4.10. Determination of Epitopes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- dos Santos, M.L.; Quintilio, W.; Manieri, T.M.; Tsuruta, L.R.; Moro, A.M. Advances and challenges in therapeutic monoclonal antibodies drug development. Braz. J. Pharm. Sci. 2018, 54, e01007. [Google Scholar] [CrossRef]
- Collingridge, G.L.; Davies, J. The in vitro inhibition of GABA release by tetanus toxin. Neuropharmacology 1982, 21, 851–855. [Google Scholar] [CrossRef]
- Yen, L.M.; Thwaites, C.L. Tetanus. Lancet 2019, 393, 1657–1668. [Google Scholar] [CrossRef]
- Chen, C.; Fu, Z.J.; Kim, J.J.P.; Barbieri, J.T.; Baldwin, M.R. Gangliosides as High Affinity Receptors for Tetanus Neurotoxin. J. Biol. Chem. 2009, 284, 26569–26577. [Google Scholar] [CrossRef] [PubMed]
- Minamitani, T.; Kiyose, K.; Otsubo, R.; Ito, T.; Akiba, H.; Furuta, R.A.; Inoue, T.; Tsumoto, K.; Satake, M.; Yasui, T. Novel neutralizing human monoclonal antibodies against tetanus neurotoxin. Sci. Rep. 2021, 11, 12134. [Google Scholar] [CrossRef]
- Restani, L.; Giribaldi, F.; Manich, M.; Bercsenyi, K.; Menendez, G.; Rossetto, O.; Caleo, M.; Schiavo, G. Botulinum Neurotoxins A and E Undergo Retrograde Axonal Transport in Primary Motor Neurons. PLoS Pathog. 2012, 8, e1003087. [Google Scholar] [CrossRef]
- Cook, T.M.; Protheroe, R.T.; Handel, J.M. Tetanus: A review of the literature. Br. J. Anaesth. 2001, 87, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, R.; Fang, T.; Yu, T.; Chi, X.Y.; Zhang, X.P.; Liu, S.L.; Fu, L.; Yu, C.M.; Chen, W. Tetanus Neurotoxin Neutralizing Antibodies Screened from a Human Immune scFv Antibody Phage Display Library. Toxins 2016, 8, 266. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, C.; Yu, J.; Lin, S.; Liu, T.; Zan, L.; Li, N.; Hong, P.; Wang, X.; Jia, Z.; et al. Structural basis of tetanus toxin neutralization by native human monoclonal antibodies. Cell Rep. 2021, 35, 109070. [Google Scholar] [CrossRef]
- WHO. Total Tetanus: Reported Cases by WHO Region. Available online: https://apps.who.int/gho/data/view.main.1520_46?lang=en (accessed on 15 January 2024).
- BRAZIL. Situação Epidemiológica. Available online: https://www.gov.br/saude/pt-br/assuntos/saude-de-a-a-z/t/tetano-acidental/situacao-epidemiologica-do-tetano-acidental (accessed on 13 February 2024).
- Aliprandini, E.; Takata, D.Y.; Lepique, A.; Kalil, J.; Boscardin, S.B.; Moro, A.M. An oligoclonal combination of human monoclonal antibodies able to neutralize tetanus toxin in vivo. Toxicon X 2019, 2, 100006. [Google Scholar] [CrossRef]
- Manieri, T.M.; Takata, D.Y.; Targino, R.C.; Quintilio, W.; Batalha-Carvalho, J.V.; da Silva, C.M.L.; Moro, A.M. Characterization of Neutralizing Human Anti-Tetanus Monoclonal Antibodies Produced by Stable Cell Lines. Pharmaceutics 2022, 14, 1985. [Google Scholar] [CrossRef] [PubMed]
- Wodak, S.J.; Mendez, R. Prediction of protein-protein interactions: The CAPRI experiment, its evaluation and implications. Curr. Opin. Struct. Biol. 2004, 14, 242–249. [Google Scholar] [CrossRef]
- Bercsenyi, K.; Schmieg, N.; Bryson, J.B.; Wallace, M.; Caccin, P.; Golding, M.; Zanotti, G.; Greensmith, L.; Nischt, R.; Schiavo, G. Tetanus toxin entry. Nidogens are therapeutic targets for the prevention of tetanus. Science 2014, 346, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Cubi, R.; Candalija, A.; Ortega, A.; Gil, C.; Aguilera, J. Tetanus Toxin Hc Fragment Induces the Formation of Ceramide Platforms and Protects Neuronal Cells against Oxidative Stress. PLoS ONE 2013, 8, e68055. [Google Scholar] [CrossRef] [PubMed]
- Purkiss, J.R.; Friis, L.M.; Doward, S.; Quinn, C.P. Clostridium botulinum neurotoxins act with a wide range of potencies on SH-SY5Y human neuroblastoma cells. Neurotoxicology 2001, 22, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Gorio, A. Ganglioside enhancement of neuronal differentiation, plasticity, and repair. CRC Crit. Rev. Clin. Neurobiol. 1986, 2, 241–296. [Google Scholar] [PubMed]
- Adekar, S.P.; Takahashi, T.; Jones, R.M.; Al-Saleem, F.H.; Ancharski, D.M.; Root, M.J.; Kapadnis, B.P.; Simpson, L.L.; Dessain, S.K. Neutralization of botulinum neurotoxin by a human monoclonal antibody specific for the catalytic light chain. PLoS ONE 2008, 3, e3023. [Google Scholar] [CrossRef] [PubMed]
- Ghotloo, S.; Golsaz-Shirazi, F.; Amiri, M.M.; Jeddi-Tehrani, M.; Shokri, F. Neutralization of tetanus toxin by a novel chimeric monoclonal antibody. Toxicon Off. J. Int. Soc. Toxinol. 2021, 201, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.L.; Dong, M.; Yao, J.; Tepp, W.H.; Lin, G.; Johnson, E.A.; Chapman, E.R. SV2 mediates entry of tetanus neurotoxin into central neurons. PLoS Pathog. 2010, 6, e1001207. [Google Scholar] [CrossRef]
- Tehran, D.A.; Pirazzini, M. Preparation of Cerebellum Granule Neurons from Mouse or Rat Pups and Evaluation of Clostridial Neurotoxin Activity and Their Inhibitors by Western Blot and Immunohistochemistry. Bio-Protocol 2018, 8, e2918. [Google Scholar] [CrossRef]
- Pirazzini, M.; Grinzato, A.; Corti, D.; Barbieri, S.; Leka, O.; Vallese, F.; Tonellato, M.; Silacci-Fregni, C.; Piccoli, L.; Kandiah, E.; et al. Exceptionally potent human monoclonal antibodies are effective for prophylaxis and treatment of tetanus in mice. J. Clin. Investig. 2021, 131, e151676. [Google Scholar] [CrossRef] [PubMed]
- Lalli, G.; Herreros, J.; Osborne, S.L.; Montecucco, C.; Rossetto, O.; Schiavo, G. Functional characterisation of tetanus and botulinum neurotoxins binding domains. J. Cell Sci. 1999, 112 Pt. 16, 2715–2724. [Google Scholar] [CrossRef]
- Williamson, L.C.; Bateman, K.E.; Clifford, J.C.; Neale, E.A. Neuronal sensitivity to tetanus toxin requires gangliosides. J. Biol. Chem. 1999, 274, 25173–25180. [Google Scholar] [CrossRef] [PubMed]
- Bahney, J.; von Bartheld, C.S. The Cellular Composition and Glia-Neuron Ratio in the Spinal Cord of a Human and a Nonhuman Primate: Comparison With Other Species and Brain Regions. Anat. Rec. 2018, 301, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Seybold, V.S.; Abrahams, L.G. Pain Research: Methods and Protocols; Luo, Z.D., Ed.; Humana Press: Totowa, NJ, USA, 2004; pp. 203–213. [Google Scholar]
- Fotinou, C.; Emsley, P.; Black, I.; Ando, H.; Ishida, H.; Kiso, M.; Sinha, K.A.; Fairweather, N.F.; Isaacs, N.W. The crystal structure of tetanus toxin Hc fragment complexed with a synthetic GT1b analogue suggests cross-linking between ganglioside receptors and the toxin. J. Biol. Chem. 2001, 276, 32274–32281. [Google Scholar] [CrossRef]
- Lukic, I.; Marinkovic, E.; Filipovic, A.; Krnjaja, O.; Kosanovic, D.; Inic-Kanada, A.; Stojanovic, M. Key protection factors against tetanus: Anti-tetanus toxin antibody affinity and its ability to prevent tetanus toxin—Ganglioside interaction. Toxicon Off. J. Int. Soc. Toxinol. 2015, 103, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Tahmasebi, F.; Younesi, V.; Razavi, A.; Khoshnoodi, J.; Bayat, A.A.; Abbasi, E.; Rabbani, H.; Jeddi-Tehrani, M.; Shokri, F. Characterization of neutralizing monoclonal antibodies directed against tetanus toxin fragment C. J. Immunotoxicol. 2014, 11, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yu, R.; Chi, X.; Chen, Z.; Hao, M.; Du, P.; Fan, P.; Liu, Y.; Dong, Y.; Fang, T.; et al. Tetanus vaccine-induced human neutralizing antibodies provide full protection against neurotoxin challenge in mice. Int. Immunopharmacol. 2021, 91, 107297. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Y.; Cui, J.; Liu, D.; Zhang, W.; Xue, C.; Xiong, X.; Liu, G.; Chen, H. Preparation and characterization of a neutralizing murine monoclonal antibody against tetanus toxin. J. Immunol. Methods 2023, 513, 113427. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. In Assay Guidance Manual [Internet]; Grossman, A., Arkin, M., Auld, D., Austin, C., Baell, J., Brimacombe, K., Chung, T.D.Y., Coussens, N.P., Dahlin, J.L., Devanarayan, V., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2013. [Google Scholar]
- Swindells, M.B.; Porter, C.T.; Couch, M.; Hurst, J.; Abhinandan, K.R.; Nielsen, J.H.; Macindoe, G.; Hetherington, J.; Martin, A.C. abYsis: Integrated Antibody Sequence and Structure-Management, Analysis, and Prediction. J. Mol. Biol. 2017, 429, 356–364. [Google Scholar] [CrossRef]
- Lefranc, M.P.; Pommie, C.; Ruiz, M.; Giudicelli, V.; Foulquier, E.; Truong, L.; Thouvenin-Contet, V.; Lefranc, G. IMGT unique numbering for immunoglobulin and T cell receptor variable domains and Ig superfamily V-like domains. Dev. Comp. Immunol. 2003, 27, 55–77. [Google Scholar] [CrossRef] [PubMed]
- Leem, J.; Dunbar, J.; Georges, G.; Shi, J.; Deane, C.M. ABodyBuilder: Automated antibody structure prediction with data-driven accuracy estimation. mAbs 2016, 8, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- van Zundert, G.C.P.; Rodrigues, J.; Trellet, M.; Schmitz, C.; Kastritis, P.L.; Karaca, E.; Melquiond, A.S.J.; van Dijk, M.; de Vries, S.J.; Bonvin, A. The HADDOCK2.2 Web Server: User-Friendly Integrative Modeling of Biomolecular Complexes. J. Mol. Biol. 2016, 428, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Sircar, A.; Gray, J.J. SnugDock: Paratope structural optimization during antibody-antigen docking compensates for errors in antibody homology models. PLoS Comput. Biol. 2010, 6, e1000644. [Google Scholar] [CrossRef] [PubMed]
- Pierce, B.G.; Wiehe, K.; Hwang, H.; Kim, B.H.; Vreven, T.; Weng, Z. ZDOCK server: Interactive docking prediction of protein-protein complexes and symmetric multimers. Bioinformatics 2014, 30, 1771–1773. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand-Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mAb | Program | Input | Output | Clusters | Total Models | Models Binding HCR/T |
|---|---|---|---|---|---|---|
| TT-117 | HADDOCK 2.4 | 1 * | 400 | 6 | 24 | 8 |
| TT-140 | HADDOCK 2.4 | 1 * | 400 | 10 | 40 | 4 |
| TT-117 | Snug Dock | 10 ** | 10,000 *** | - | 100 | 93 |
| TT-140 | Snug Dock | 10 ** | 10,000 *** | - | 100 | 63 |
| TT-117 | TT-140 | ||
|---|---|---|---|
| Residue | Incidence | Residue | Incidence |
| HIS 1293 | 2.56% | TRP 1289 | 3.06% |
| HIS 1271 | 2.41% | HIS 1271 | 3.06% |
| ASN 1171 | 2.34% | ASN 1219 | 2.64% |
| ILE 1275 | 2.11% | HIS 1293 | 2.64% |
| ILE 1298 | 2.11% | LYS 1297 | 2.53% |
| LEU 1221 | 1.96% | ASN 1272 | 2.53% |
| GLY 1276 | 1.96% | ASP 1296 | 2.53% |
| PRO 1279 | 1.89% | ILE 1298 | 2.53% |
| PRO 1155 | 1.89% | TYR 1165 | 2.11% |
| LYS 1143 | 1.89% | GLN 1274 | 1.90% |
| TRP 1289 | 1.81% | ||
| TT-117 | TT-140 | ||
|---|---|---|---|
| Residue | Incidence | Residue | Incidence |
| ASN 1219 | 3.92% | HIS 1271 | 4.27% |
| ASN 1277 | 3.92% | ASN 1219 | 3.96% |
| ASN 1153 | 3.23% | ASN 1280 | 3.66% |
| ASN 1216 | 3.23% | LYS 1297 | 3.66% |
| GLN 1274 | 3.23% | ASN 1277 | 3.35% |
| ASN 1203 | 3.00% | ASP 1296 | 3.35% |
| ASN 1205 | 3.00% | ASN 1272 | 3.05% |
| ASN 1280 | 2.76% | ASN 1216 | 3.05% |
| SER 1156 | 2.30% | GLN 1274 | 3.05% |
| ASN 1204 | 2.07% | ASP 1278 | 2.44% |
| ASN 1292 | 2.07% | ASP 1282 | 2.44% |
| ASN 1144 | 1.84% | HIS 1293 | 2.44% |
| ARG 1281 | 1.84% | SER 1135 | 2.44% |
| GLN 1141 | 1.84% | ASP 1214 | 2.13% |
| GLY 1276 | 1.84% | ARG 1226 | 2.13% |
| TYR 1202 | 1.84% | ASN 1163 | 2.13% |
| ASP 1282 | 1.84% | ASN 1292 | 2.13% |
| ASP 1147 | 1.83% | ||
| SER 1137 | 1.83% | ||
| GLY 1276 | 1.83% | ||
| TYR 1165 | 1.83% | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, C.P.; Barreiros, G.M.; Oliveira, A.S.A.; de Souza, M.M.; Manieri, T.M.; Moro, A.M. A Dual Strategy—In Vitro and In Silico—To Evaluate Human Antitetanus mAbs Addressing Their Potential Protective Action on TeNT Endocytosis in Primary Rat Neuronal Cells. Int. J. Mol. Sci. 2024, 25, 5788. https://doi.org/10.3390/ijms25115788
Lima CP, Barreiros GM, Oliveira ASA, de Souza MM, Manieri TM, Moro AM. A Dual Strategy—In Vitro and In Silico—To Evaluate Human Antitetanus mAbs Addressing Their Potential Protective Action on TeNT Endocytosis in Primary Rat Neuronal Cells. International Journal of Molecular Sciences. 2024; 25(11):5788. https://doi.org/10.3390/ijms25115788
Chicago/Turabian StyleLima, Cauã Pacheco, Gabriela Massaro Barreiros, Adriele Silva Alves Oliveira, Marcelo Medina de Souza, Tania Maria Manieri, and Ana Maria Moro. 2024. "A Dual Strategy—In Vitro and In Silico—To Evaluate Human Antitetanus mAbs Addressing Their Potential Protective Action on TeNT Endocytosis in Primary Rat Neuronal Cells" International Journal of Molecular Sciences 25, no. 11: 5788. https://doi.org/10.3390/ijms25115788
APA StyleLima, C. P., Barreiros, G. M., Oliveira, A. S. A., de Souza, M. M., Manieri, T. M., & Moro, A. M. (2024). A Dual Strategy—In Vitro and In Silico—To Evaluate Human Antitetanus mAbs Addressing Their Potential Protective Action on TeNT Endocytosis in Primary Rat Neuronal Cells. International Journal of Molecular Sciences, 25(11), 5788. https://doi.org/10.3390/ijms25115788