
AFB1 Toxicity in Human Food and Animal Feed Consumption: A Review of Experimental Treatments and Preventive Measures

 , ,
, ,
Abstract
1. Introduction
2. AFB1 Prevalence, Detection, and Treatment of the AFB1 Animal Model
3. AFB1 in Animal Injury Models: Dosing and the Use of the Model
4. Novel Development of the Application of Microbiota in AFB1 Toxicity Relief
5. Biomedical Development to Prevent AFB1 Poisoning
6. Hepatocarcinoma with Respect to AFB1 Toxicity
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Correction Statement
References
- Jallow, A.; Xie, H.; Tang, X.; Qi, Z.; Li, P. Worldwide aflatoxin contamination of agricultural products and foods: From occurrence to control. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2332–2381. [Google Scholar] [CrossRef] [PubMed]
- Bharose, A.A.; Hajare, S.T.; Narayanrao, D.R.; Gajera, H.G.; Prajapati, H.K.; Singh, S.C.; Upadhye, V. Whole genome sequencing and annotation of Aspergillus flavus JAM-JKB-B HA-GG20. Sci. Rep. 2024, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Manzanares, N.; Rodríguez-Estévez, V.; Arenas-Fernández, P.; García-Campaña, A.M.; Gámiz-Gracia, L. Occurrence of Mycotoxins in Swine Feeding from Spain. Toxins 2019, 11, 342. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, C.H.; Gonçalves, C.L.; da Silva Nascente, P. Aflatoxins B1 and M1: Risks related to milk produced in Brazil. Ann. Microbiol. 2018, 68, 793–802. [Google Scholar] [CrossRef]
- Sulaiman, S.H.; Jamaluddin, R.; Sabran, M.R. Association between Urinary Aflatoxin (AFM1) and Dietary Intake among Adults in Hulu Langat District, Selangor, Malaysia. Nutrients 2018, 10, 460. [Google Scholar] [CrossRef]
- Administration U.S. Food and Drug. Aflatoxin Poisoning in Pets. 2023. Available online: https://www.fda.gov/animal-veterinary/animal-health-literacy/aflatoxin-poisoning-pets (accessed on 9 May 2024).
- Umar, A.; Bhatti, H.S.; Honey, S.F. A call for aflatoxin control in Asia. CABI Agric. Biosci. 2023, 4, 27. [Google Scholar] [CrossRef]
- Hoeltz, M.; Einloft, T.C.; Oldoni, V.P.; Dottori, H.A.; Noll, I.B. The Occurrence of Aflatoxin B1 Contamination in Peanuts and Peanut Products Marketed in Southern Brazil. Braz. Arch. Biol. Technol. 2015, 75, 1698. [Google Scholar] [CrossRef]
- Bashiry, M.; Javanmardi, F.; Sadeghi, E.; Shokri, S.; Hossieni, H.; Oliveira, C.A.F.; Khaneghah, A.M. The prevalence of aflatoxins in commercial baby food products: A global systematic review, meta-analysis, and risk assessment study. Trends Food Sci. Technol. 2021, 114, 100–115. [Google Scholar] [CrossRef]
- Saad-Hussein, A.; Shahy, E.M.; Shaheen, W.; Ibrahim, K.S.; Mahdy-Abdallah, H.; Taha, M.M.; Hafez, S.F. Hepatotoxicity of aflatoxin B1 and its oxidative effects in wood dust Egyptian exposed workers. Arch. Environ. Occup. Health 2020, 8, 561–566. [Google Scholar] [CrossRef]
- Farag, R.M.M.A.; Al Ayobi, D.; Kwon, H.J.; Ansary, A.E. Relationship Between Aflatoxin B1 Exposure and Etiology of Liver Disease in Saudi Arabian Patients. J. Pure Appl. Microbiol. 2018, 12, 1147–1153. [Google Scholar] [CrossRef]
- Li, C.; Liu, X.; Wu, J.; Ji, X.; Xu, Q. Research progress in toxicological effects and mechanism of aflatoxin B1 toxin. PeerJ 2022, 10, e13850. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wu, Y.; Zhang, M.; Lan, H. Aflatoxin B1 exposure triggers inflammation and premature skin aging via ERMCS/Ca2+/ROS signaling cascade. Int. Immunopharmacol. 2023, 124, 110961. [Google Scholar] [CrossRef]
- Kumar, L.K.; Verma, S.K.; Chandel, R.; Thumar, M.; Singh, D.; Onteru, S.K. Aflatoxin M1 decreases the expression of genes encoding tight junction proteins and influences the intestinal epithelial integrity. Mycotoxin Res. 2023, 39, 453–467. [Google Scholar] [CrossRef]
- Smith, L.E.; Prendergast, A.J.; Turner, P.C.; Humphrey, J.H.; Stoltzfus, R.J. Aflatoxin Exposure During Pregnancy, Maternal Anemia, and Adverse Birth Outcomes. Am. J. Trop Med. Hyg. 2017, 96, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Popescu, R.G.; Radulescu, A.L.; Sergiu Emil Georgescu, S.E.; Dinischiotu, A. Aflatoxins in feed: Types, metabolism, health consequences in swine and mitigation strategies. Toxins 2022, 14, 853. [Google Scholar] [CrossRef] [PubMed]
- Al-Mudallal, N.H.A.L. The Expression of MMP1 and MMP7 in Mice Liver after Exposure to Aflatoxin B1 Using Immunohistochemistry Technique. Arch. Razi. Inst. 2023, 78, 63–72. [Google Scholar]
- Johnsson, P.; Lindblad, M.; Thim, A.M.; Jonsson, N.; Vargas, E.A.; Medeiros, N.L.; Brabet, C.; de Quaresma, M. Growth of aflatoxigenic moulds and aflatoxin formation in Brazil nuts. World Mycotoxin J. 2008, 1, 127–137. [Google Scholar] [CrossRef]
- Bragagnolo, L.M.M.N.; Calori, M.A.; Iamanaka, B.T.; Alves, M.C.; Silva, J.J.; Godoy, I.J.; Taniwaki, M.H. Assessment of early harvest in the prevention of aflatoxins in peanuts during drought stress conditions. Int. J. Food Microbiol. 2023, 405, 110336. [Google Scholar]
- Kortei, S.F.N.K.; Sackey, M.; Richard, S.A. Aflatoxin B1 induces fertility, fetal deformities, and potential therapies. Open Med. 2024, 19, 20240907. [Google Scholar]
- Zhang, Y.Z.; Zhao, Q.H.; Duan, H.W.; Zou, Y.J.; Sun, S.C.; Hu, L.L. Aflatoxin B1 exposure disrupts organalle distribution in mouse oocytes. PeerJ 2022, 10, e13497. [Google Scholar] [CrossRef]
- He, H.L.Y.; Gao, X.; Li, T.; Qiao, B.; Tang, L.; Lan, J.; Sun, Q.; Ruan, Z. Curcumin alleviates AFB1-induced nephrotoxicity in ducks: Regulating mitochondrial oxidative stress, ferritinophagy, and ferroptosis. Mycotoxin Res. 2023, 39, 437–451. [Google Scholar]
- Ren, X.-L.; Han, P.; Meng, Y. Aflatoxin B1-Induced COX-2 Expression Promotes Mitophagy and Contributes to Lipid Accumulation in Hepatocytes In Vitro and In Vivo. Int. J. Toxicol. 2020, 39, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Tan, Y.Q.; Leung, L.K. Aflatoxin B1 disrupts transient receptor potential channel activity and increases COX-2 expression in JEG-3 placental cells. Chem. Biol. Interact. 2016, 260, 84–90. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Zhan, D.L.; Chen, Y.Y.; Wang, W.H.; He, C.Y.; Lin, Y.; Lin, Y.C.; Lin, Z.N. Aflatoxin B1 enhances pyroptosis of hepatocytes and activation of Kupffer cells to promote liver inflammatory injury via dephosphorylation of cyclooxygenase-2: An in vitro, ex vivo and in vivo study. Arch. Toxicol. 2019, 93, 3305–3320. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Lin, Y.; Han, P.Y.; Jiang, S.; Che, L.; He, C.Y.; Lin, Y.C.; Lin, Z.N. HBx combined with AFB1 triggers hepatic steatosis via COX-2-mediated necrosome formation and mitochondrial dynamics disorder. J. Cell Mol. Med. 2019, 23, 5920–5933. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.H.; Chen, X.Y.; Hou, L.L.; Dong, J.H.; Liu, H.W.; Zhu, L.Q.; Chen, F. Limosilactobacillus reuteri peptidoglycan alleviates aflatoxin B1-induced toxicity through adsorbing toxins and improving growth, antioxidant status, immunity and liver pathological changes in chicks. Br. Poult. Sci. 2024, 11, 1–9. [Google Scholar] [CrossRef]
- Deng, J.; Zhao, L.; Zhang, N.Y.; Karrow, N.A.; Krumm, C.S.; Qi, D.S.; Sun, L.H. Aflatoxin B1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res. Rev. Mutat. Res. 2018, 778, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wen, T.; Cao, A.; Wang, J.; Pan, H.; Zhao, R. Bile Acids promote hepatic biotransformatin and excretion of aflatoxin B1 in broiler chicken. Toxins 2023, 15, 694. [Google Scholar] [CrossRef]
- Kovács, C.A.S.; Pócsi, I.; Pusztahelyi, T. Elimination of Deoxynivalenol, Aflatoxin B1, and Zearalenone by Gram-Positive Microbes (Firmicutes). Toxins 2022, 14, 591. [Google Scholar] [CrossRef]
- Wu, G.; San, J.; Pang, H.; Du, Y.; Li, W.; Zhou, X.; Yang, X.; Hu, J.; Yang, J. Taurine attenuates AFB1-induced liver injury by alleviating oxidative stress and regulating mitochondria-mediated apoptosis. Toxicon 2022, 215, 17–27. [Google Scholar] [CrossRef]
- Umarani, M.; Shanthi, P.; Sachdanandam, P. Protective effect of Kalpaamruthaa in combating the oxidative stress posed by aflatoxin B1-induced hepatocellular carcinoma with special reference to flavonoid structure-activity relationship. Liver Int. 2018, 28, 200–2013. [Google Scholar] [CrossRef]
- Altyar, A.E.; Kensara, O.A.; Sayed, A.A.; Aleya, L.; Almutairi, M.H.; Zaazouee, M.S.; Elshanbary, A.A.; El-Demerdash, F.M.; Abdel-Daim, M.M. Acute aflatoxin B1-induced hepatic and cardiac oxidative damage in rats: Ameliorative effects of morin. Heliyon 2023, 9, e21837. [Google Scholar] [CrossRef] [PubMed]
- Dobiesz, K.; Kozera, W.; Okorski, A.; Żak, G.; Krzysztoń, J.; Karpiesiuk, K. Effect of sex on the chemical composition and fatty acid profile of the longissimus dorsi muscle in danbred hybrid pigs. Rocz. Nauk. Zootech. 2023, 50, 311–320. [Google Scholar]
- Liu, H.; Xie, R.; Huang, W.; Yang, Y.; Zhou, M.; Lu, B.; Li, B.; Tan, B.; Dong, X. Effects of Dietary Aflatoxin B1 on Hybrid Grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂) Growth, Intestinal Health, and Muscle Quality. Aquac. Nutr. 2024, 2024, 3920254. [Google Scholar] [CrossRef]
- Karpiesiuk, K.; Polak-Śliwińska, M.; Dąbrowska, J.; Pszczółkowska, A.; Kozera, W.; Czarnik, U.; Okorski, A. Evaluation of fungal diversity and mycotoxin content in pig feed containing clusterbean (Cyamopsis tetragonoloba) meal. J. Elem. 2023, 4, 1203–1220. [Google Scholar] [CrossRef]
- Zhuo, Y.; Yang, P.; Hua, L.; Zhu, L.; Zhu, X.; Han, X.; Pang, X.; Xu, S.; Jiang, X.; Lin, Y.; et al. Effects of Chronic Exposure to Diets Containing Moldy Corn or Moldy Wheat Bran on Growth Performance, Ovarian Follicular Pool, and Oxidative Status of Gilts. Toxins 2022, 14, 413. [Google Scholar] [CrossRef] [PubMed]
- Popescu, R.G.; Bulgaru, C.; Untea, A.; Vlassa, M.; Filip, M.; Hermenean, A.; Marin, D.; Tăranu, I.; Georgescu, S.E.; Dinischiotu, A. The Effectiveness of Dietary Byproduct Antioxidants on Induced CYP Genes Expression and Histological Alteration in Piglets Liver and Kidney Fed with Aflatoxin B1 and Ochratoxin A. Toxins 2021, 13, 148. [Google Scholar] [CrossRef]
- Taranu, I.; Marin, D.E.; Palade, M.; Pistol, G.C.; Chedea, V.S.; Gras, M.A.; Rotar, C. Assessment of the efficacy of a grape seed waste in counteracting the changes induced by aflatoxin B1 contaminated diet on performance, plasma, liver and intestinal tissues of pigs after weaning. Toxicon 2019, 162, 24–31. [Google Scholar] [CrossRef]
- Tarasconi, L.; Dazuk, V.; Molosse, V.L.; Cécere, B.G.O.; Deolindo, G.L.; Mendes, R.E.; Gloria, E.M.; Ternus, E.M.; Galli, G.M.; Paiano, D.; et al. Nursery pigs fed with feed contaminated by aflatoxin B1 (Aspergillus flavus) and anti-mycotoxin blend: Pathogenesis and negative impact on animal health and weight gain. Microb. Pathog. 2023, 186, 106474. [Google Scholar] [CrossRef] [PubMed]
- Preetha, S.P.; Kanniappan, M.; Selvakumar, E.; Nagaraj, M.; Varalakshmi, P. Lupeol ameliorates aflatoxin B1-induced peroxidative hepatic damage in rats. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 333–339. [Google Scholar] [CrossRef]
- El-Agamy, D.S. Comparative effects of curcumin and resveratrol on aflatoxin B(1)-induced liver injury in rats. Arch. Toxicol. 2010, 84, 389–396. [Google Scholar] [CrossRef]
- Weaver, A.; See, T.; Hansen, J.; Kim, Y.; Souza, A.; Middleton, T.; Kim, S.W. The use of feed additives to reduce the effects of aflatoxin and deoxynivalenol on pig growth, organ health and immune status during chronic exposure. Toxins 2013, 5, 1261–1281. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, S.; Zhao, C.; Zhou, Y.; Zhang, L.; Liu, H.; Zhou, P.; Li, S.; Fu, L.; Zheng, Z.; et al. Genome-Scale CRISPR Knockout Screening Identifies BACH1 as a Key Regulator of Aflatoxin B1-Induced Oxidative Damage. Antioxidants 2022, 11, 1787. [Google Scholar] [CrossRef]
- Dazuk, V.; Tarasconi, L.; Molosse, V.L.; Cécere, B.G.O.; Deolindo, G.L.; Strapazzon, J.V.; Bottari, N.B.; Bissacotti, B.F.; Schetinger, M.R.C.; Sareta, L.; et al. Can the Inclusion of a Vegetable Biocholine Additive in Pig Feed Contaminated with Aflatoxin Reduce Toxicological Impacts on Animal Health and Performance? Animals 2023, 13, 3010. [Google Scholar] [CrossRef]
- Monmeesil, P.; Fungfuang, W.; Tulayakul, P.; Pongchairerk, U. The effects of astaxanthin on liver histopathology and expression of superoxide dismutase in rat aflatoxicosis. J. Vet. Med. Sci. 2019, 81, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Gross-Steinmeyer, K.; Stapleton, P.L.; Tracy, J.H.; Bammler, T.K.; Strom, S.C.; Eaton, D.L. Sulforaphane- and phenethyl isothiocyanate-induced inhibition of aflatoxin B1-mediated genotoxicity in human hepatocytes: Role of GSTM1 genotype and CYP3A4 gene expression. Toxicol. Sci. 2010, 116, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Qiu, L.; Deng, Q.; Fang, Z.; Sun, L.; Wang, Y.; Gooneratne, R.; Zhao, J. L-Cysteine hydrochloride inhibits Aspergillus flavus growth and AFB1 synthesis by disrupting cell structure and antioxidant system balance. J. Hazard Mater. 2023, 459, 132218. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, R.; Wei, G.; Guo, G.; Yu, H.; Zhang, Y.; Ishfaq, M.; Fazilani, S.A.; Zhang, X. Curcumin protects against Aflatoxin B1-induced liver injury in broilers via the modulation of long non-coding RNA expression. Ecotoxicol. Environ. Saf. 2020, 208, 111725. [Google Scholar] [CrossRef]
- Amminikutty, N.; Spalenza, V.; Jarriyawattanachaikul, W.; Badino, P.; Capucchio, M.T.; Colombino, E.; Schiavone, A.; Greco, D.; D’Ascanio, V.; Avantaggiato, G.; et al. Turmeric powder counteracts oxidative stress and reduces AFB1 content in the liver of broilers exposed to the EU maximum levels of the mycotoxin. Toxins 2023, 15, 687. [Google Scholar] [CrossRef]
- Lu, Q.; Hu, Y.; Nabi, F.; Li, Z.; Janyaro, H.; Zhu, W.; Liu, J. Effect of Penthorum Chinense Pursh Compound on AFB1-Induced Immune Imbalance via JAK/STAT Signaling Pathway in Spleen of Broiler Chicken. Vet. Sci. 2023, 10, 521. [Google Scholar] [CrossRef]
- Song, C.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Hesperetin protects hippocampal neurons from the neurotoxicity of Aflatoxin B1 in mice. Ecotoxicol Env. Saf. 2024, 269, 115782. [Google Scholar] [CrossRef]
- Oloruntola, O.D. Red chili powder dietary supplementation regularized the performance, hematobiochemical indices, oxidative status, and 8-hydroxy-2′-deoxyguanosine of aflatoxin B1 exposed broiler chickens. Transl. Anim. Sci. 2024, 8, txae006. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M.S.A.; Abadi, A.M.; El-Kott, A.F.; Mohamed, G.; Alrashdi, B.M.; Eid, R.A.; Salem, T.E. Protective efficacy of luteolin against aflatoxin-induced toxicity, oxidative damage, and apoptosis in the rat liver. Environ. Sci. Pollut. Res. Int. 2023, 30, 52358–52368. [Google Scholar] [CrossRef]
- Liew, W.P.P.; Mohd-Redzwan, S.; Than, L.T.L. Gut Microbiota Profiling of Aflatoxin B1-Induced Rats Treated with Lactobacillus casei Shirota. Toxins 2019, 11, 49. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Lin, L.; Wang, S.; Ding, W.; Zhang, C.; Shaukat, A.; Xu, B.; Yue, K.; Zhang, C.; Liu, F. Total Flavonoids of Rhizoma Drynariae Mitigates Aflatoxin B1-Induced Liver Toxicity in Chickens via Microbiota-Gut-Liver Axis Interaction Mechanisms. Antioxidants 2023, 12, 819. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Ford, E.; Hobbs, S.M.; Baird, S.M.; Lu, S.E. Occidiofungin Is the Key Metabolite for Antifungal Activity of the Endophytic Bacterium Burkholderia sp. MS455 Against Aspergillus flavus. Phytopathol. 2021, 112, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Fu, P.; Zhang, C.; Xu, T.; Cao, Q.; Shaukat, A.; Yue, K.; Liu, F.; Dong, H.; Huang, S.; et al. Evaluation of gut microbiota composition to screening for potential biomarker in AFB1-exposed sheep. 3 Biotech 2023, 13, 409. [Google Scholar] [CrossRef]
- Muñoz-Solano, B.; González-Peñas, E. Co-Occurrence of Mycotoxins in Feed for Cattle, Pigs, Poultry, and Sheep in Navarra, a Region of Northern Spain. Toxins 2023, 15, 172. [Google Scholar] [CrossRef] [PubMed]
- Hassanen, E.I.; Ahmed, L.I.; Fahim, K.M.; Shehata, M.G.; Badr, A.N. Chitosan nanoparticle encapsulation increased the prophylactic efficacy of Lactobacillus plantarum RM1 against AFM1-induced hepatorenal toxicity in rats. Environ. Sci. Pollut. Res. Int. 2023, 59, 123925–123938. [Google Scholar] [CrossRef] [PubMed]
- Marlida, Y.; Nurmiati, N.; Husmaini, H.; Huda, N.; Anggraini, L.; Ardani, L.R. The potential of lactic acid bacteria isolated from ikan budu (fermented fish) to inhibit the growth of pathogenic fungi and detoxify aflatoxin B1. Vet. World 2023, 16, 1373–1379. [Google Scholar] [CrossRef]
- Aoyanagi, M.M.C.C.; Budiño, F.E.L.; Raj, J.; Vasiljevic, M.; Ali, S.; Ramalho, L.N.Z.; Silva, F.R.; Corassin, C.H.; Ghantous, G.F.; de Oliveira, C.A.F. Efficacy of Two Commercially Available Adsorbents to Reduce the Combined Toxic Effects of Dietary Aflatoxins, Fumonisins and Zearalenone and Their Residues in the Tissues of Weaned Pigs. Toxins 2023, 15, 629. [Google Scholar] [CrossRef]
- Paiano, D.; Bottari, N.B.; Santurio, J.M.; Zampar, A.; Schetinger, M.R.C.; Zanette, R.A.; Mendes, R.E.; Gloria, E.M.; Baldissera, M.D.; Da Silva, A.S. Spray-dried porcine plasma added to diets contaminated with aflatoxins and fumonisins shows beneficial effects to piglet health. An. Acad. Bras. Cienc. 2018, 90, 3115–3128. [Google Scholar]
- Costamanga, D.; Gaggiotti, M.; Smulovitz, A.; Abdala, A.; Signorini, M. Mycotoxin sequestering agent: Impact on health and performance of dairy cows and efficacy in reducing AFM1 residues in milk. Environ. Toxicol. Pharmacol. 2023, 105, 104349. [Google Scholar] [CrossRef] [PubMed]
- Kalemba, M.R.K.; Makhuvele, R.; Njobeh, P.B. Phytochemical screening, antioxidant activity of selected methanolic plant extracts and their detoxification capabilities against AFB1 toxicity. Heliyon 2024, 10, e24435. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, Y.; Feng, T.; Yan, Z.; Zhu, D.; Lin, H.; Iqbal, M.; Deng, D.; Kulyar, M.F.; Shen, Y. Hedyotis diffusa alleviate aflatoxin B1-induced liver injury in ducks by mediating Nrf2 signaling pathway. Ecotoxicol. Environ. Saf. 2023, 249, 114339. [Google Scholar] [CrossRef] [PubMed]
- Shyamal, S.; Latha, P.G.; Suja, S.R.; Shine, V.J.; Anuja, G.I.; Sini, S.; Pradeep, S.; Shikha, P.; Rajasekharan, S. Hepatoprotective effect of three herbal extracts on aflatoxin B1-intoxicated rat liver. Singap. Med. J. 2010, 51, 326–331. [Google Scholar]
- Solcan, C.; Gogu, M.; Floristean, V.; Oprisan, B.; Solcan, G. The hepatoprotective effect of sea buckthorn (Hippophae rhamnoides) berries on induced aflatoxin B1 poisoning in chickens. Poult. Sci. 2013, 92, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, X.; Chai, X.; Jiao, Y.; Sun, J.; Wang, S.; Yu, H.; Feng, X. Curcumin Mitigates Oxidative Damage in Broiler Liver and Ileum Caused by Aflatoxin B1-Contaminated Feed through Nrf2 Signaling Pathway. Animals 2024, 14, 409. [Google Scholar] [CrossRef]
- Rastogi, R.; Srivastava, A.K.; Rastogi, A.K. Long term effect of aflatoxin B(1) on lipid peroxidation in rat liver and kidney: Effect of picroliv and silymarin. Phytother. Res. 2001, 15, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Vipin, A.V.; Raksha Rao, K.; Kurrey, N.K.; Anu Appaiah, K.A.; Venkateswaran, G. Protective effects of phenolics rich extract of ginger against Aflatoxin B1-induced oxidative stress and hepatotoxicity. Biomed. Pharmacother. 2017, 91, 415–424. [Google Scholar]
- Aborehab, N.M.; Waly, N.E. IL-6 and NFE2L2: A putative role for the hepatoprotective effect of N. Sativa, P. Ginseng and C. Sempervirens in AFB-1 induced hepatocellular carcinoma in rats. Toxicol. Rep. 2019, 6, 457–464. [Google Scholar] [CrossRef]
- Yaman, T.; Yener, Z.; Celik, I. Histopathological and biochemical investigations of protective role of honey in rats with experimental aflatoxicosis. BMC Complement. Altern. Med. 2016, 16, 232. [Google Scholar] [CrossRef] [PubMed]
- Okoro, N.; Alilonu, D.O.; Eze, M.C.; Ebokaiwe, A.P. Aflatoxin B1-induced redox imbalance in the hippocampus and cerebral cortex of male Wistar rats is accompanied by altered cholinergic, indoleaminergic, and purinergic pathways: Abatement by dietary rutin. Toxicon 2024, 239, 107595. [Google Scholar] [CrossRef] [PubMed]
- Kihal, A.; Rodríguez-Prado, M.; Calsamiglia, S. A network meta-analysis on the efficacy of different mycotoxin binders to reduce aflatoxin M1 in milk after aflatoxin B1 challenge in dairy cows. J. Dairy Sci. 2023, 106, 5379–5387. [Google Scholar] [CrossRef] [PubMed]
- Khedr, S.M.; Ghareeb, D.A.; Fathy, S.A.; Hamdy, G.M. Berberine-loaded albumin nanoparticles reverse aflatoxin B1-induced liver hyperplasia. BMC Pharmacol. Toxicol. 2023, 24, 24–42. [Google Scholar] [CrossRef]
- Li, S.F.; Zhang, S.B.; Zhai, H.C.; Lv, Y.Y.; Hu, Y.S.; Cai, J.P. Hexanal induces early apoptosis of Aspergillus flavus conidia by disrupting mitochondrial function and expression of key genes. Appl. Microbiol. Biotechnol. 2021, 105, 6871–6886. [Google Scholar] [CrossRef] [PubMed]
- Dallabona, C.; Pioli, M.; Spadola, G.; Orsoni, N.; Bisceglie, F.; Lodi, T.; Pelosi, G.; Restivo, F.M.; Degola, F. Sabotage at the Powerhouse? Unraveling the Molecular Target of 2-Isopropylbenzaldehyde Thiosemicarbazone, a Specific Inhibitor of Aflatoxin Biosynthesis and Sclerotia Development in Aspergillus flavus, Using Yeast as a Model System. Molecules 2019, 24, 2971. [Google Scholar] [CrossRef]
- Guo, H.; Wang, P.; Liu, C.; Chang, J.; Yin, Q.; Wang, L.; Jin, S.; Zhu, Q.; Lu, F. Compound mycotoxin detoxifier alleviating aflatoxin B1 toxic effects on broiler growth performance, organ damage and gut microbiota. Poult. Sci. 2022, 102, 102434. [Google Scholar] [CrossRef] [PubMed]
- Ghaemmaghami, S.S.; Rouhanipour, H.; Sharifi, S.D. Aflatoxin levels in poultry feed: A comparison of mash and pellet forms. Poult. Sci. 2024, 103, 103254. [Google Scholar] [CrossRef]
- Akinrinde, A.S.; Adebiyi, O.E.; Asekun, A. Amelioration of Aflatoxin B1-induced gastrointestinal injuries by Eucalyptus oil in rats. J. Complement. Integr. Med. 2020, 17, 20190002. [Google Scholar] [CrossRef]
- Accardi, R.; Gruffat, H.; Sirand, C.; Fusil, F.; Gheit, T.; Hernandez-Vargas, H.; Le Calvez-Kelm, F.; Traverse-Glehen, A.; Cosset, F.L.; Manet, E.; et al. The mycotoxin aflatoxin B1 stimulates Epstein-Barr virus-induced B-cell transformation in in vitro and in vivo experimental models. Carcinogenesis 2015, 36, 1440–1451. [Google Scholar] [CrossRef]
- Ahmadi, M.; Shahbahrami, R.; Khajeh, F.; Khodaeivandi, S.; Kakavandi, E.; Raziabad, R.H.; Ghanati, K. Aflatoxin B1 and viruses’ combined pathogenesis: A mini systematics review of invitro and invivo studies. Acta Histochem. 2023, 126, 152116. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Ma, Y.; Liang, J.; Wei, Z.; Li, M.; Zhang, Y.; Liu, M.; He, H.; Qu, C.; Cai, J.; et al. AHR mediates the aflatoxin B1 toxicity associated with hepatocellular carcinoma. Sig. Transduct. Target. Ther. 2021, 6, 299. [Google Scholar] [CrossRef] [PubMed]
- Kant, R.; Yen, C.H.; Lu, C.K.; Lin, Y.C.; Li, J.H.; Chen, Y.M. Identification of 1,2,3,4,6-Penta-O-galloyl-β-d-glucopyranoside as a Glycine N-Methyltransferase Enhancer by High-Throughput Screening of Natural Products Inhibits Hepatocellular Carcinoma. Int. J. Mol. Sci. 2016, 17, 669. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Robertson, I. The epidemiology of swine influenza. Anim. Dis. 2021, 1, 21. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Su, J.; Liu, Z.; Liu, D.; Gan, F.; Chen, X.; Huang, K. Aflatoxin B1 Promotes Influenza Replication and Increases Virus Related Lung Damage via Activation of TLR4 Signaling. Front. Immunol. 2018, 9, 2297. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Zhang, G.; Ma, Y.; Wu, D.; Jiang, S.; Zhou, S.; Jiang, X. Circulating Metabolic Markers Related to the Diagnosis of Hepatocellular Carcinoma. J. Oncol. 2022, 2022, 7840606. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Quintana, R.; Sánchez-Alarcón, J.; Tenorio-Arvide, M.G.; Deng, Y.; Montiel-González, J.M.; Gómez-Arroyo, S.; Villalobos-Pietrini, R.; Cortés-Eslava, J.; Flores-Márquez, A.R.; Arenas-Huertero, F. The microRNAs as potential biomarkers for predicting the onset of aflatoxin exposure in human beings: A review. Front. Microbiol. 2014, 5, 102. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.H.; Chan, C.; Yang, P.J.; Huang, Y.H.; Lee, M.H. Genetic Susceptibility to Hepatocellular Carcinoma in Patients with Chronic Hepatitis Virus Infection. Viruses 2023, 15, 559. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.I.; Murakami, A.E.; Fernandes, A.M.; Ospina-Rojas, I.C.; Nunes, K.C.; Hirata, A.K. Performance and serum biochemical profile of Japanese quail supplemented with silymarin and contaminated with aflatoxin B1. Poult. Sci. 2018, 97, 159–165. [Google Scholar] [CrossRef]
- Alshamrani, A.A.; Alwetaid, M.Y.; Al-Hamamah, M.A.; Attia, M.S.M.; Ahmad, S.F.; Algonaiah, M.A.; Nadeem, A.; Ansari, M.A.; Bakheet, S.A.; Attia, S.M. Aflatoxin B1 Exacerbates Genomic Instability and Apoptosis in the BTBR Autism Mouse Model via Dysregulating DNA Repair Pathway. Toxics 2023, 11, 636. [Google Scholar] [CrossRef]
- Huang, B.; Chen, Q.; Wang, L.; Gao, X.; Zhu, W.; Mu, P.; Deng, Y. Aflatoxin B1 Induces Neurotoxicity through Reactive Oxygen Species Generation, DNA Damage, Apoptosis, and S-Phase Cell Cycle Arrest. Int. J. Mol. Sci. 2020, 21, 6517. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Bai, H.; Chang, X.; Wu, Z.; Dong, W.; Ma, Q.; Yang, J. Aflatoxin B1-induced early developmental hepatotoxicity in larvae zebrafish. Chemosphere 2023, 340, 139940. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Zhang, H.; Wu, R.; Lu, Y.; Wang, J.; Ravikumar, A.; Tamil Selvan, G.; Zhu, N.; Zhang, Z.; Tang, J. A dual-signal aptasensor based on cascade amplification for ultrasensitive detection of aflatoxin B1. Biosens. Bioelectron. 2024, 250, 116057. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Shi, Y.; Zhang, X.; Hu, X.; Huang, X.; Liang, N.; Shen, T.; Zou, X.; Shi, J. A DNA tetrahedral scaffolds-based electrochemical biosensor for simultaneous detection of AFB1 and OTA. Food Chem. 2023, 442, 138312. [Google Scholar] [CrossRef]
- Gómez-Espinosa, D.; Cervantes-Aguilar, F.J.; Del Río-García, J.C.; Villarreal-Barajas, T.; Vázquez-Durán, A.; Méndez-Albores, A. Ameliorative Effects of Neutral Electrolyzed Water on Growth Performance, Biochemical Constituents, and Histopathological Changes in Turkey Poults during Aflatoxicosis. Toxins 2017, 9, 104. [Google Scholar] [CrossRef]
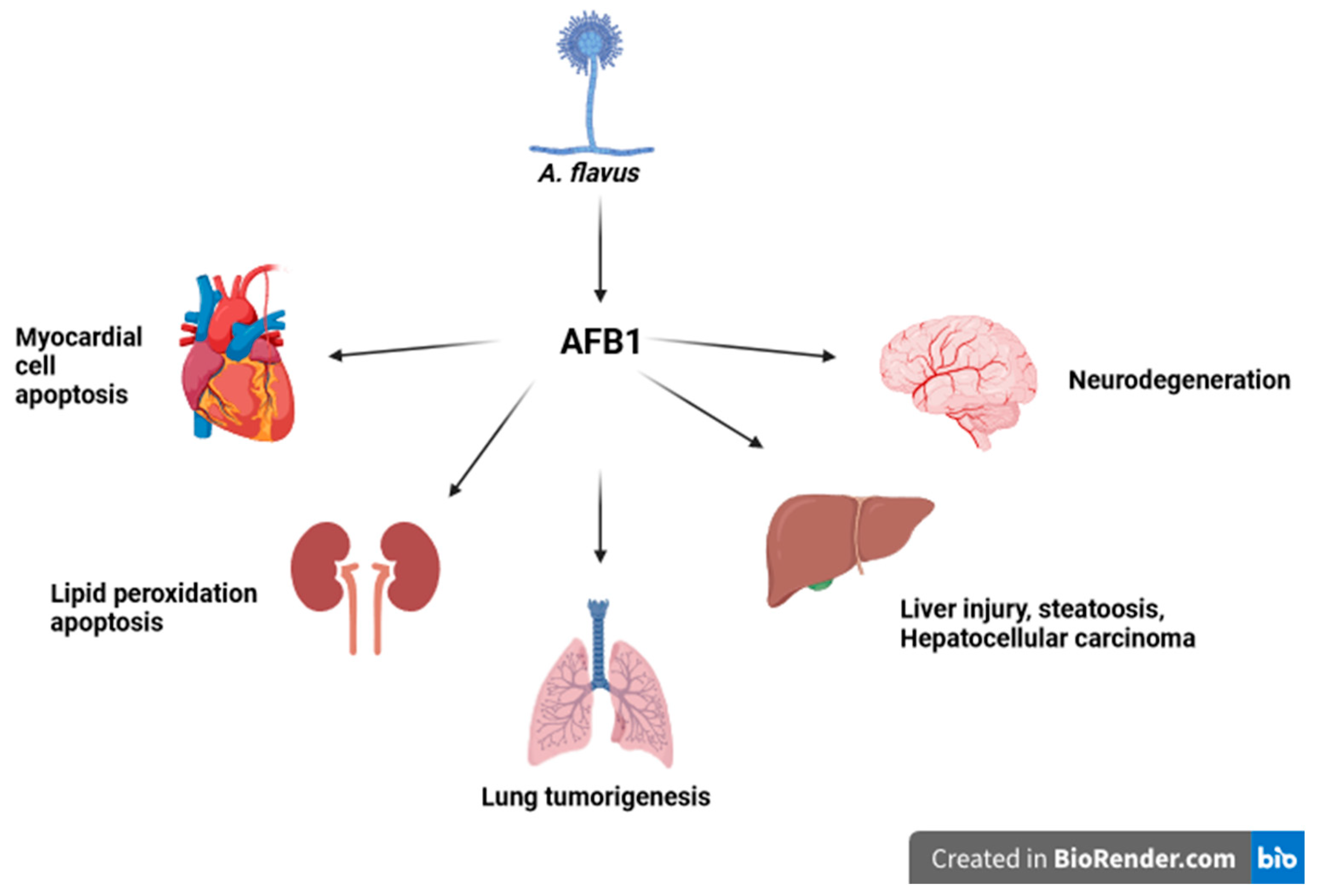
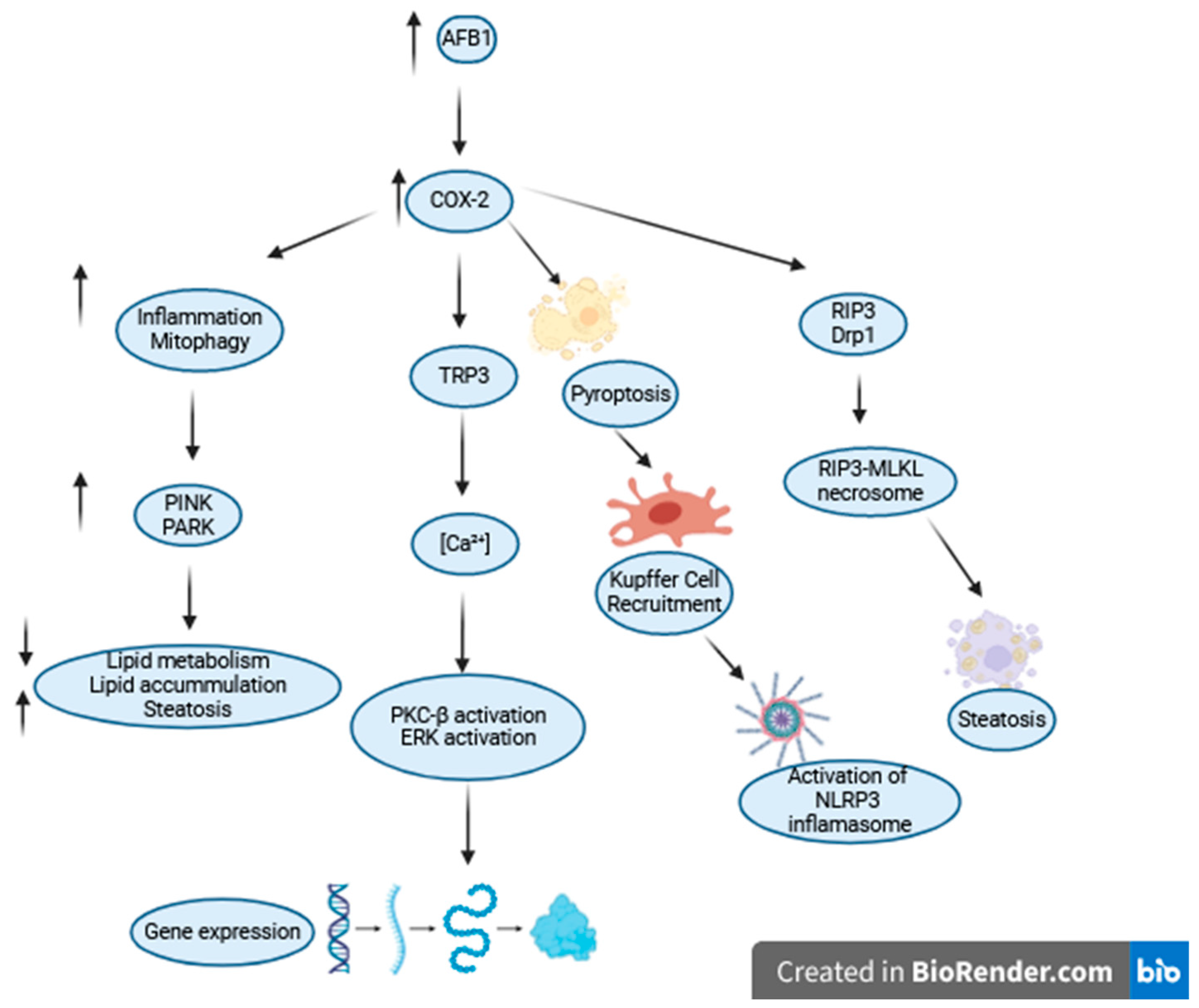


{kind=link}
{kind=link}
{kind=link}
| Treatment, Study Ref., Study Title | AFB1 Dose, Route Application, Species, Time of Treatment | Characteristics | Results |
|---|---|---|---|
| Curcumin 400 mg/kg, feed [22]; Curcumin alleviates AFB1-induced nephrotoxicity in ducks: regulating mitochondrial oxidative stress, ferritinophagy, and ferroptosis. | 0.1 mg/kg bw to ducks; gavage for 21 days | Increased mitochondrial oxidative stress, increase in MDA, 8-OHdG | Improves nephrotoxicity by inhibiting mitochondrial oxidative stress |
| Peptidoglycan L. reuteri 200 mg/kg/feed [27]; Limosilactobacillus reuteri peptidoglycan alleviates aflatoxin B1-induced toxicity through adsorbing toxins and improving growth, antioxidant status, immunity and liver pathological changes in chicks. | 71.43 µg/kg in Hy-Line brown chicks, feed for 42 days | Increased the plasma MDA content and decreased SOD and GSH-Px activity and T-AOC | Toxin adsorption, Improved immunotoxicity and hepatic status |
| Morin 30 mg/kg bw, oral [33]; Acute aflatoxin B1-induced hepatic and cardiac oxidative damage in rats: Ameliorative effects of morin. | 2.5 mg/kg bw to Wistar Albino rats, oral, twice on days 12 and 14, during a 15-day experiment | Increase in AST, ALP, LDH, GGT, CK, CK-MB, 8-OHdG, IL-1β, IL-6, TNF-a | Cardiac and hepatic levels improved of MDA, GSH, GSH-Px, antioxidant enzymes |
| Grape seed extract (320 ppm) [38]; Effectiveness of dietary byproduct antioxidants on induced CYP genes expression and histological alteration in piglets liver and kidney fed with aflatoxin B1 and ochratoxin A. | 62 ppb AFB1 to crossbred pigs (TOPIG)-40 hybrid piglets, feed for 30 days | Decrease in antioxidant activity, concentration of cytokines and TBARS | Improvement in structural morphology of liver and kidney |
| Grape waste feed (8% feed) [39]; Assessment of the efficacy of grape seed extract in counteracting the changes induced by aflatoxin B1 contaminated diet on performance, plasma, liver and intestinal tissues of pigs after weaning. | 320 ppb AFB to crossbred pig TOPIG, feed for 30 days | Increase in the pro-inflammatory levels in the liver and colon | Decrease in pro-inflammatory cytokines and TBARS |
| Lupeol isolate from Crataeva nurvala; 100 mg/kg, bw [41]; Lupeol ameliorates aflatoxin B1-induced peroxidative hepatic damage in rats. | 1 mg/kg body mass, orally to rats, for 7 days | Increase in the lipid peroxide levels decrease in the enzymatic and non-enzymatic antioxidants | Reversal of hepatic damage and improvement in lipid peroxide levels |
| Curcumin 200 mg/kg bw; Resveratrol 10 mg/kg bw [42]; Comparative effects of curcumin and resveratrol on aflatoxin B(1)-induced liver injury in rats. | 25 µg/kg bw, oral gavage, male Fischer rat for 90 days | Increased liver focal necrosis, liver enzymes, ALT, AST, γ-GT, lipid peroxidation, decrease in GSH, SOD, CAT, GSH-Px | Decrease in ALT, AST, γ-GT, GSH, SOD, CAT, GSH-Px, improved liver histology |
| Clay 2 mg/kg, dried yeast 1.5 mg/kg, yeast culture 1.1 mg/kg, [43]; The use of feed additives to reduce the effects of aflatoxin and deoxynivalenol on pig growth, organ health and immune status during chronic exposure. | 150 µg/kg to gilts, feed for 42 days | Alteration to the immune system through an increase in monocytes and immunoglobulins | Improvement in monocyte numbers, liver duct cellular hyperplasia |
| Vegetable biocholine 800 mg/kg [45]; Can the inclusion of a vegetable biocholine additive in pig feed contaminated with aflatoxin reduce toxicological impacts on animal health and performance? | 500 µg/kg daily to pig; feed for 20 days | Reduced feed consumption and weight gain Increase in the intestinal oxidative markers | Hepatoprotection |
| Curcumin 300 mg/kg; [49]; Curcumin protects against Aflatoxin B1-induced liver injury in broilers via the modulation of long non-coding RNA expression. | 1 mg/kg to broiler chicken; feed; for 28 days | Hepatic injury Increased production of ROS, antioxidant enzymes | Improved morphology, regulation of LncRNAs |
| Sea buckthorn berries oil 0.6 mL oil/kg of bw/day [68]; The hepatoprotective effect of sea buckthorn (Hippophae rhamnoides) berries on induced aflatoxin B1 poisoning in chickens. | 54 µg/kg/day to broiler chicken; oral gavage for 28 days | Reduced albumin, Increase in AST | Reduction in liver necrosis and fatty deposits |
| Turmeric Powder 400 mg/kg [50]; Turmeric powder counteracts oxidative stress and reduces AFB1 content in the liver of broilers exposed to the EU maximum levels of the mycotoxin. | 0.02 mg/kg in feed, broiler chicken for 10 days | Increase in lipid peroxidation | Increase in liver gene expression and counteracted lipid peroxidation |
| Curcumin 450 mg/kg [69]; Curcumin mitigates oxidative damage in broiler liver and ileum caused by aflatoxin B1-contaminated feed through Nrf2 Signaling pathway. | 5 mg/kg in broiler chicken; feed, for 28 days | Autophagy reduction, inflammation, mTOR increase, beclin-1, ATG, Nrf2, HO-1, dynein decrease | Inflammation, restored of Nrf2 and HO-1 expression, normalized hepatocytes morphology |
| Picroliv 25 mg/kg bw; Silymarin 20 mg/kg bw; [70]; Long-term effect of aflatoxin B(1) on lipid peroxidation in rat liver and kidney: effect of picroliv and silymarin. | 2 mg/kg bw, single i.p. injection, albino Wistar rat for 6 weeks | Increase in lipid peroxide level, decrease in enzymatic antioxidant levels | Reversal of liver peroxide enzyme pathology |
| Phenolics-rich ginger extract (GE) 100 and 250 mg/kg daily feed [71]; Protective effects of phenolics-rich extract of ginger against Aflatoxin B1-induced oxidative stress and hepatotoxicity. | 200 µg/kg in Wistar rat, i.p. alternate days for 28 days | Toxicity of liver damage, serum markers | Inhibition of the production of intracellular ROS, DNA strand break, lipid peroxidation, Increase in expression of Nrf2/HO-1 |
| N. Sativa 500 mg/kg/day, P. Ginseng 250 mg/kg/day, C. Sempervirens 300 mg/kg/day [72]; IL-6 and NFE2L2: A putative role for the hepatoprotective effect of N. Sativa, P. Ginseng and C. Sempervirens in AFB-1 induced hepatocellular carcinoma in rats. | 150 μg/kg/day to Albino Wistar rats; i.p. for 3 days | HCC | Reduction in IL-6, hs-CRP, MDA |
| Honey 1 mL/kg [73]; Histopathological and biochemical investigations of the protective role of honey in rats with experimental aflatoxicosis. | 25 µg/day; oral gavage, Sprague-Dawley rat for 90 days | Marked liver histopathological lesions, Increase in concentrations of AST, GGT, ALT, decrease in bw, decrease in CAT, GR, SOD | Decrease in lipid peroxidation, liver enzymes, increase in enzymatic and non-enzymatic antioxidants. Normalization of histology in liver and kidney |
| Rutin 50 mg/kg [74]; Aflatoxin B1-induced redox imbalance in the hippocampus and cerebral cortex of male Wistar rats is accompanied by altered cholinergic, indoleaminergic, and purinergic pathways: Abatement by dietary rutin. | 0.75 and 1.5 mg/kg bw, feed, male Wistar rat for 30 days | Neurotoxicity, decreases in acetylcholinesterase activity Decreases of SOD and CATin the hippocampus, Increase in IL-6, NO, MPO | Improved histological image of the cerebral cortex, correction of IL-6, NO, MPO levels, reduction in oxidative stress markers, increased hydrolysis of the purinergic molecules in brain areas |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pożarska, A.; Karpiesiuk, K.; Kozera, W.; Czarnik, U.; Dąbrowski, M.; Zielonka, Ł. AFB1 Toxicity in Human Food and Animal Feed Consumption: A Review of Experimental Treatments and Preventive Measures. Int. J. Mol. Sci. 2024, 25, 5305. https://doi.org/10.3390/ijms25105305
Pożarska A, Karpiesiuk K, Kozera W, Czarnik U, Dąbrowski M, Zielonka Ł. AFB1 Toxicity in Human Food and Animal Feed Consumption: A Review of Experimental Treatments and Preventive Measures. International Journal of Molecular Sciences. 2024; 25(10):5305. https://doi.org/10.3390/ijms25105305
Chicago/Turabian StylePożarska, Agnieszka, Krzysztof Karpiesiuk, Wojciech Kozera, Urszula Czarnik, Michał Dąbrowski, and Łukasz Zielonka. 2024. "AFB1 Toxicity in Human Food and Animal Feed Consumption: A Review of Experimental Treatments and Preventive Measures" International Journal of Molecular Sciences 25, no. 10: 5305. https://doi.org/10.3390/ijms25105305
APA StylePożarska, A., Karpiesiuk, K., Kozera, W., Czarnik, U., Dąbrowski, M., & Zielonka, Ł. (2024). AFB1 Toxicity in Human Food and Animal Feed Consumption: A Review of Experimental Treatments and Preventive Measures. International Journal of Molecular Sciences, 25(10), 5305. https://doi.org/10.3390/ijms25105305