
Manganese and Vanadium Co-Exposure Induces Severe Neurotoxicity in the Olfactory System: Relevance to Metal-Induced Parkinsonism
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
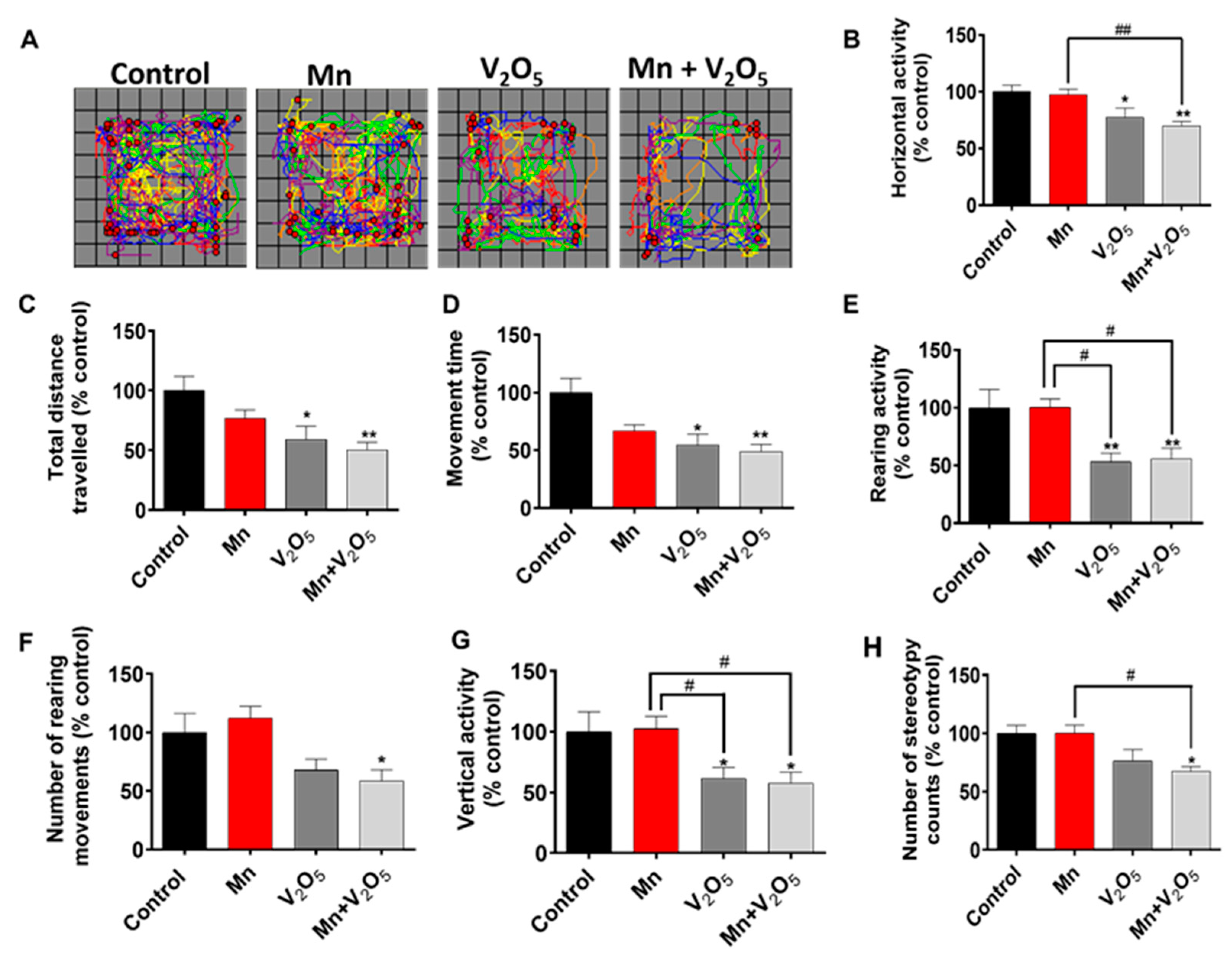
2.1. AP Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Induces Motor Deficits in Mice
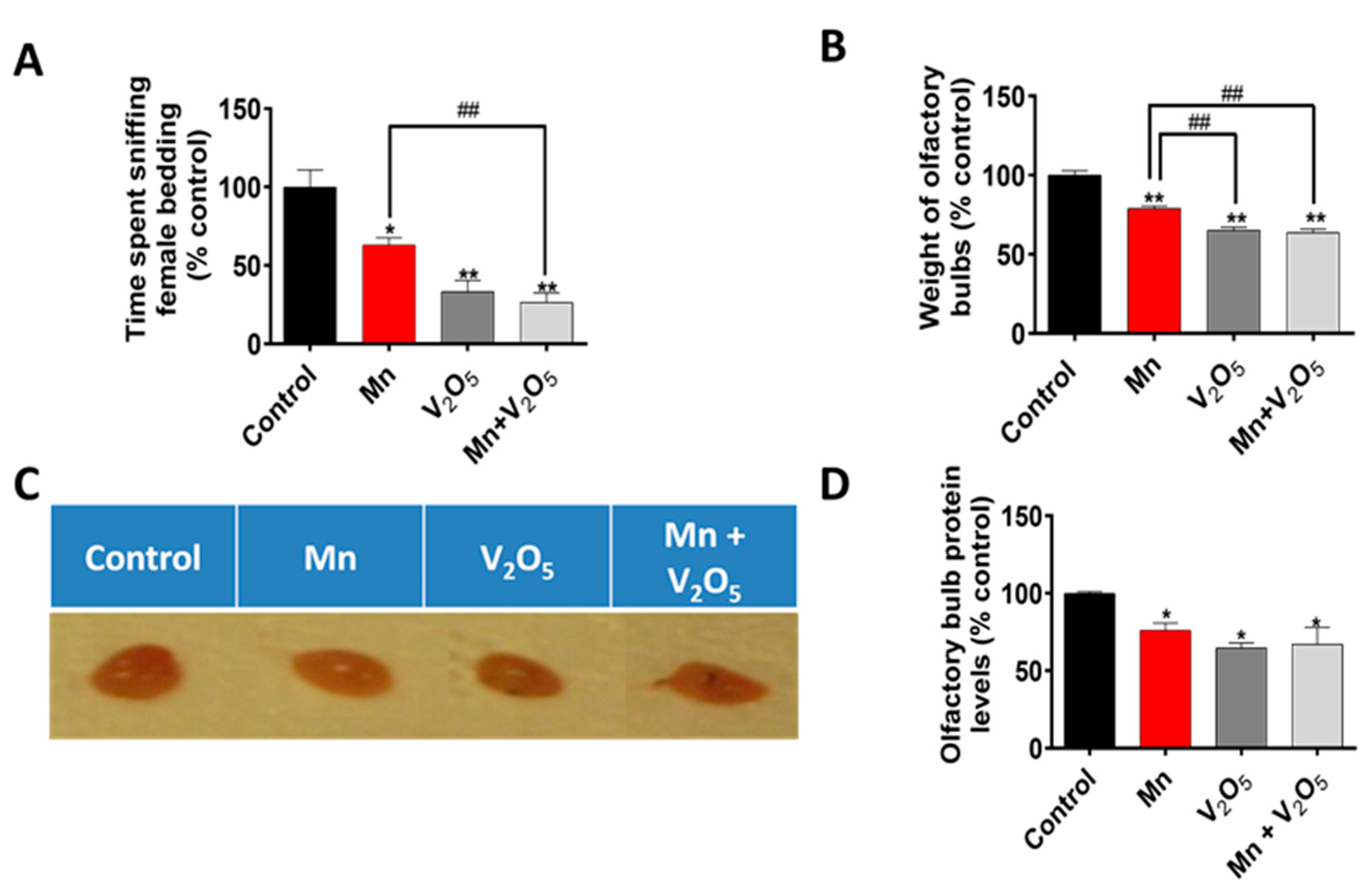
2.2. Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Induces Olfactory Dysfunction in Male Mice
2.3. Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Reduces the Weight of Olfactory Bulbs
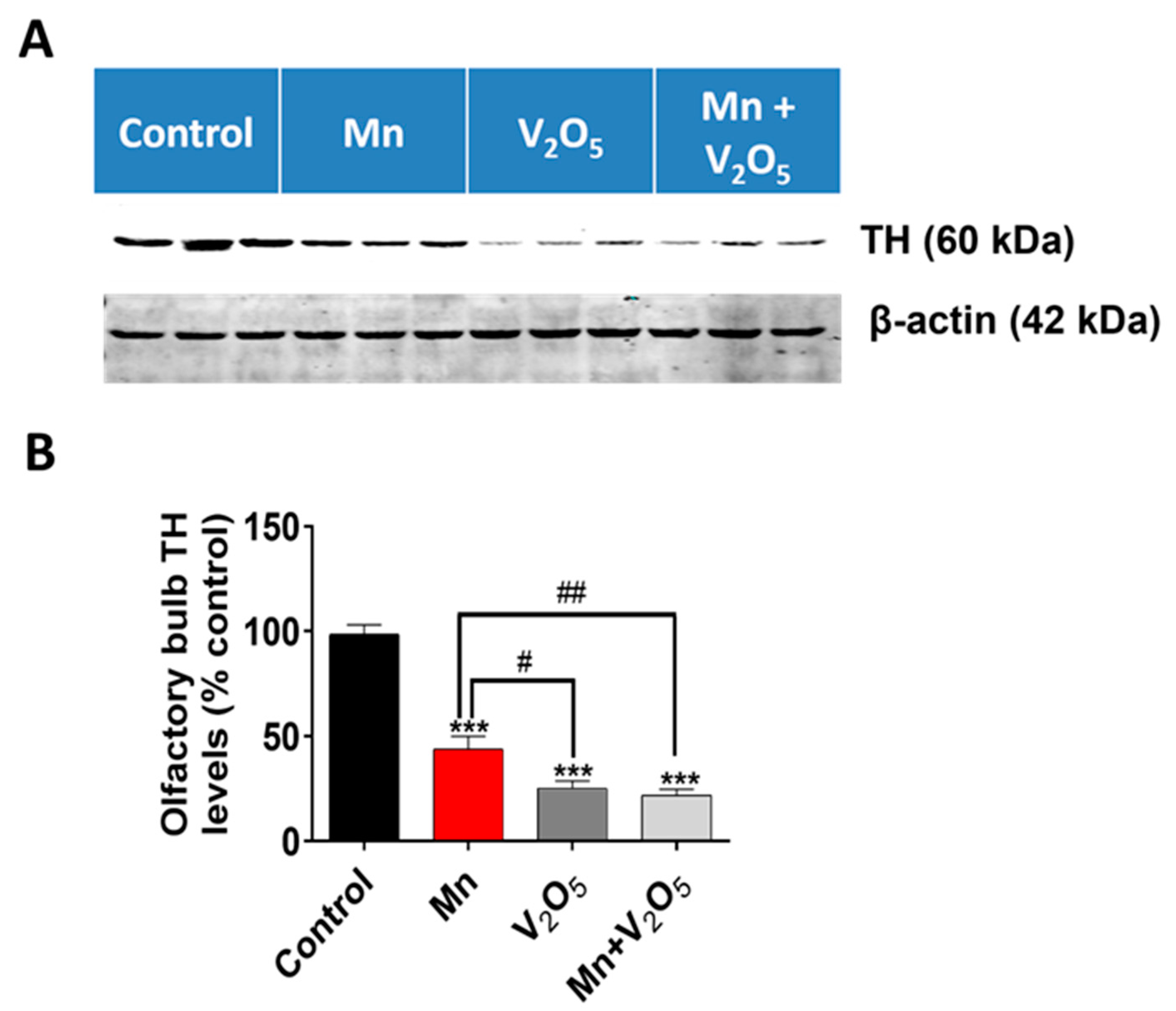
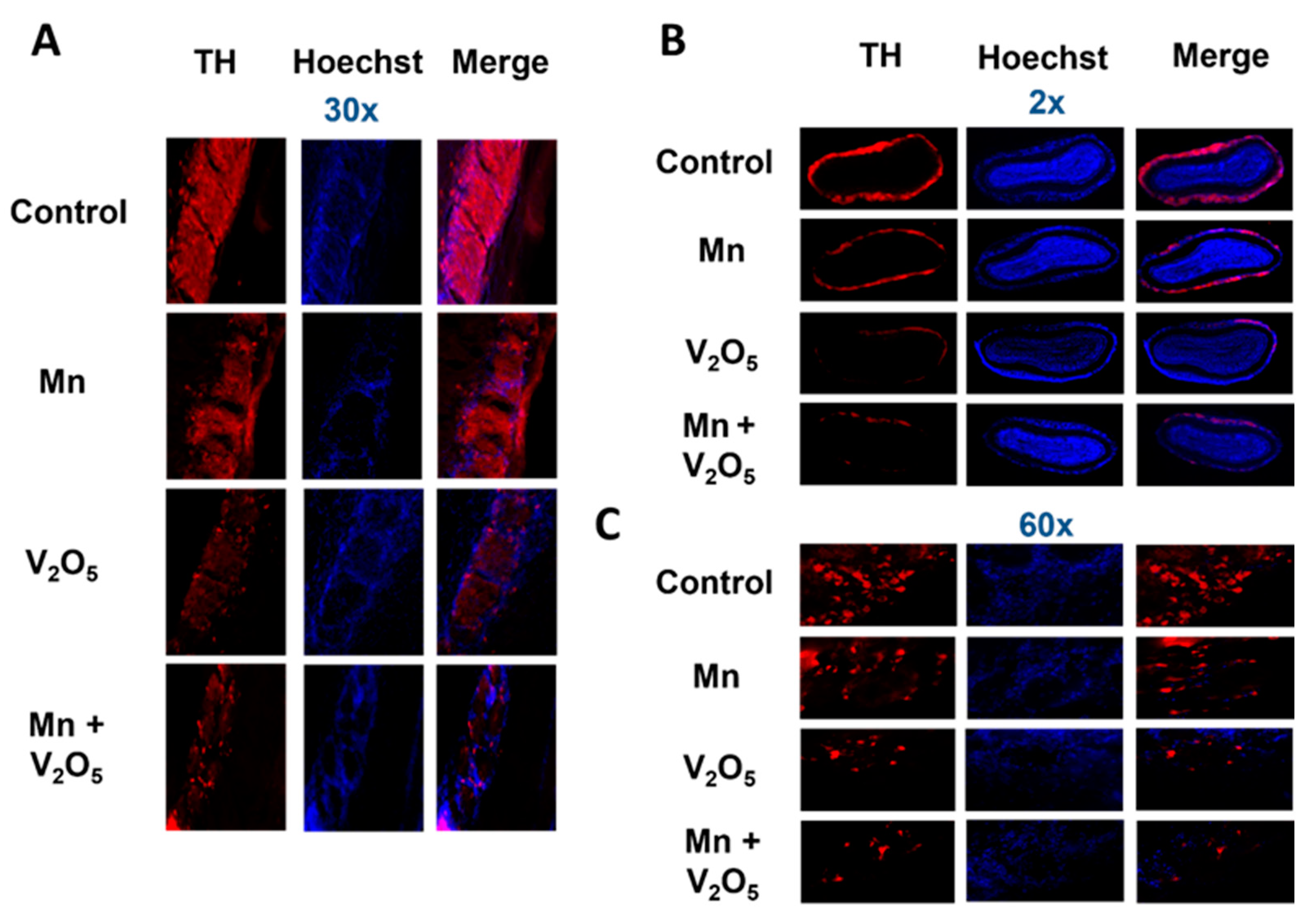
2.4. Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Causes the Loss of Tyrosine Hydroxylase (TH) and Dopaminergic Neurons in the Olfactory Bulb
2.5. Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Causes the Loss of the Neurotransmitter Dopamine and Its Metabolite 3,4-Dihydroxyphenylacetic Acid (DOPAC) in the Olfactory Bulb
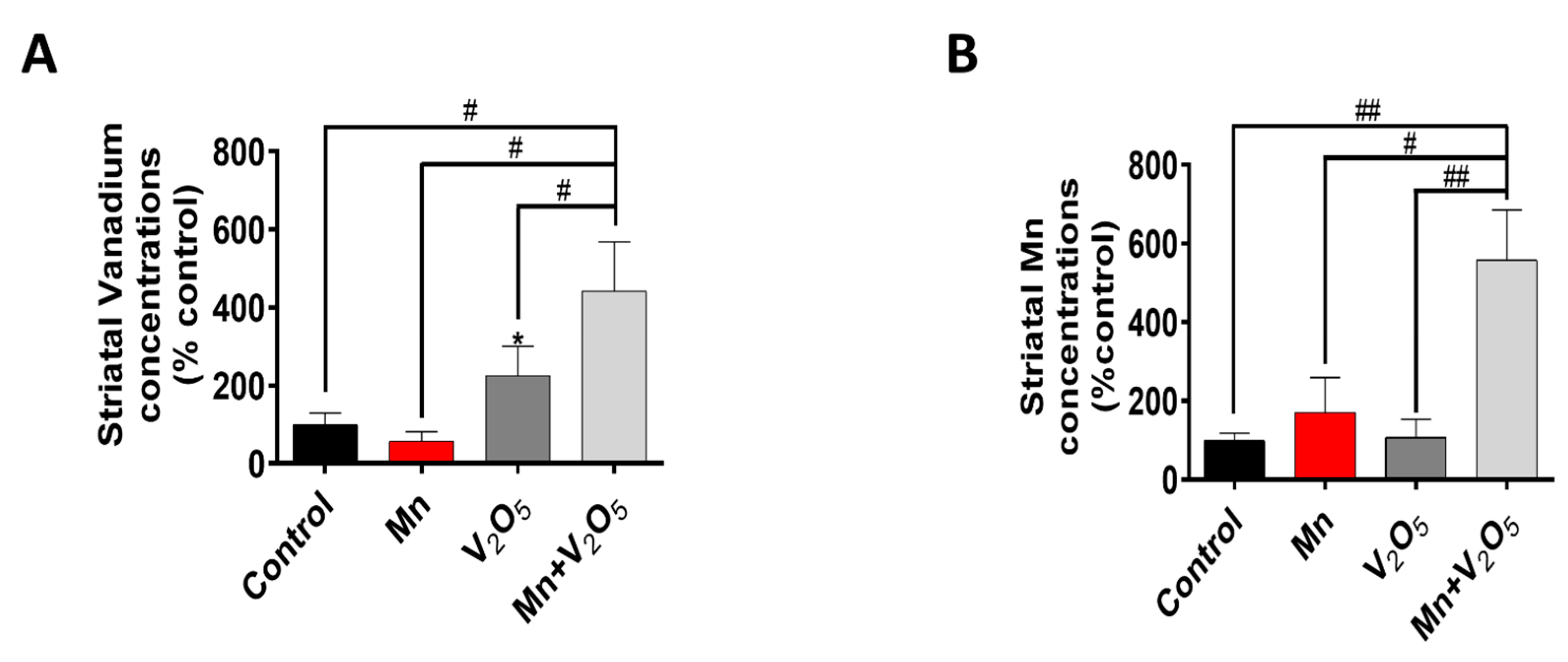
2.6. Exposure to Mn and V2O5 Mixture Results in a Higher Uptake of Both Metals in the Striatum
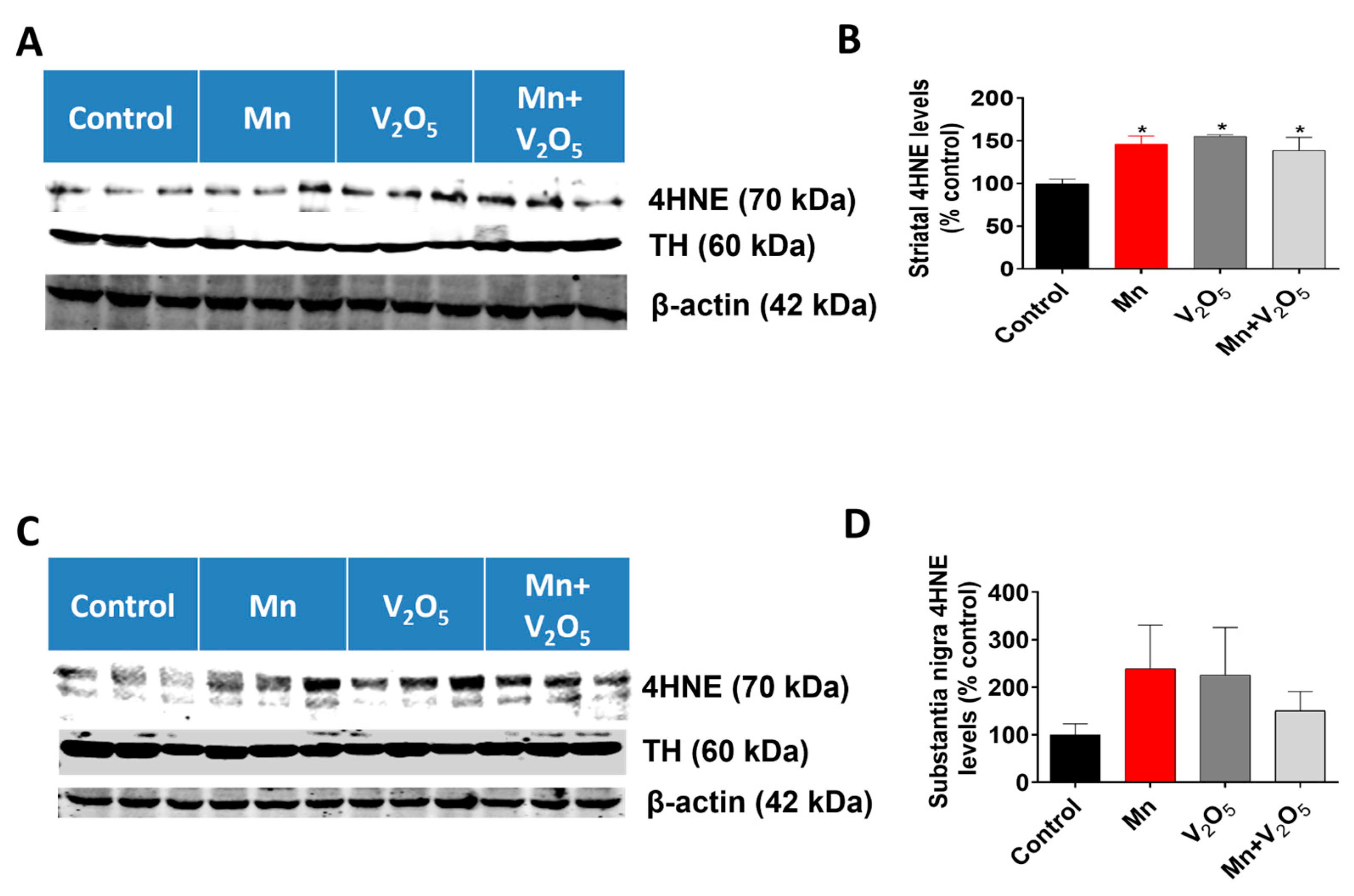
2.7. Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Induces Oxidative Stress in the Striatum and Sustantia Nigra
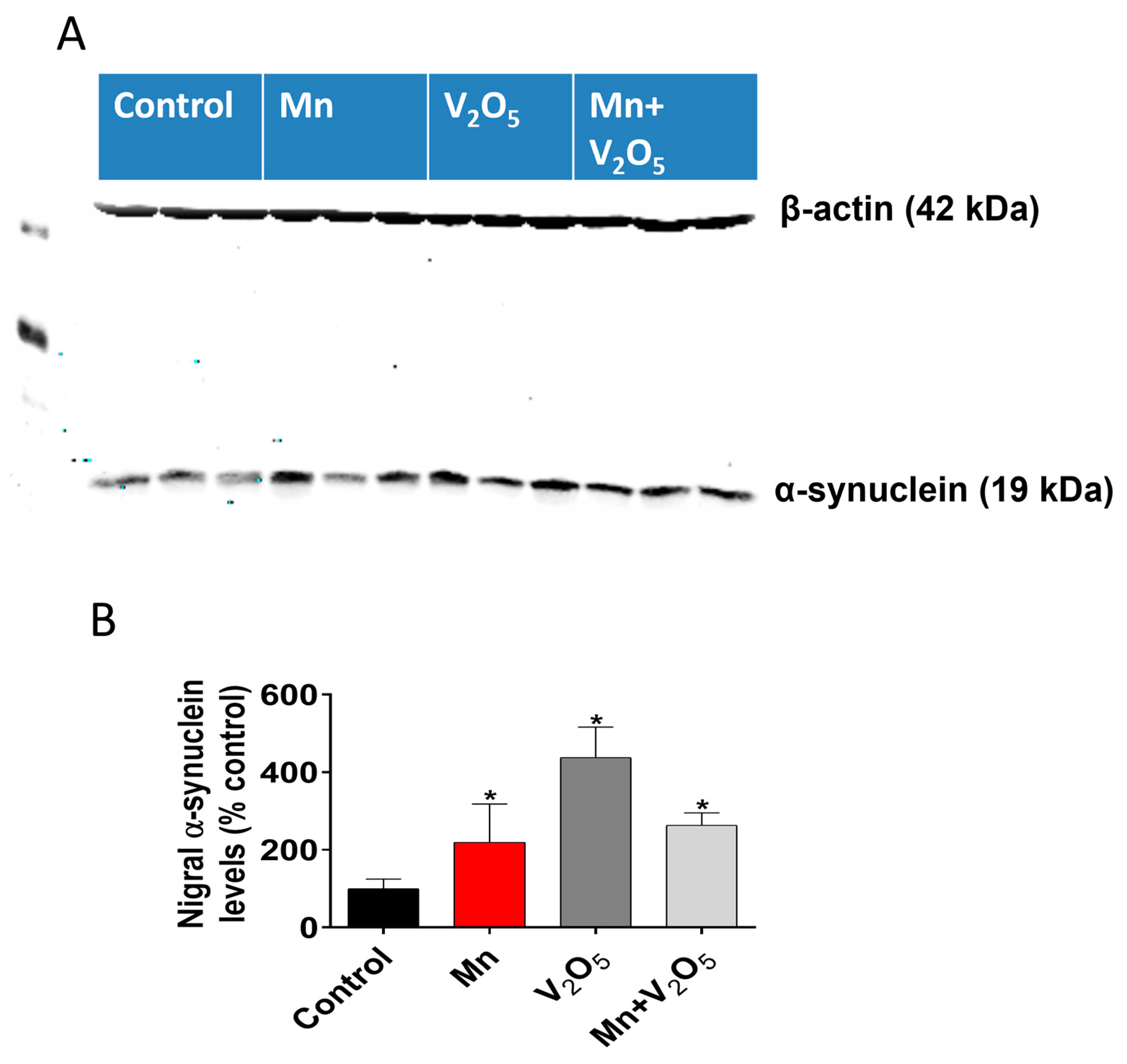
2.8. Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Induces the Accumulation of α-Synuclein in the Substantia Nigra
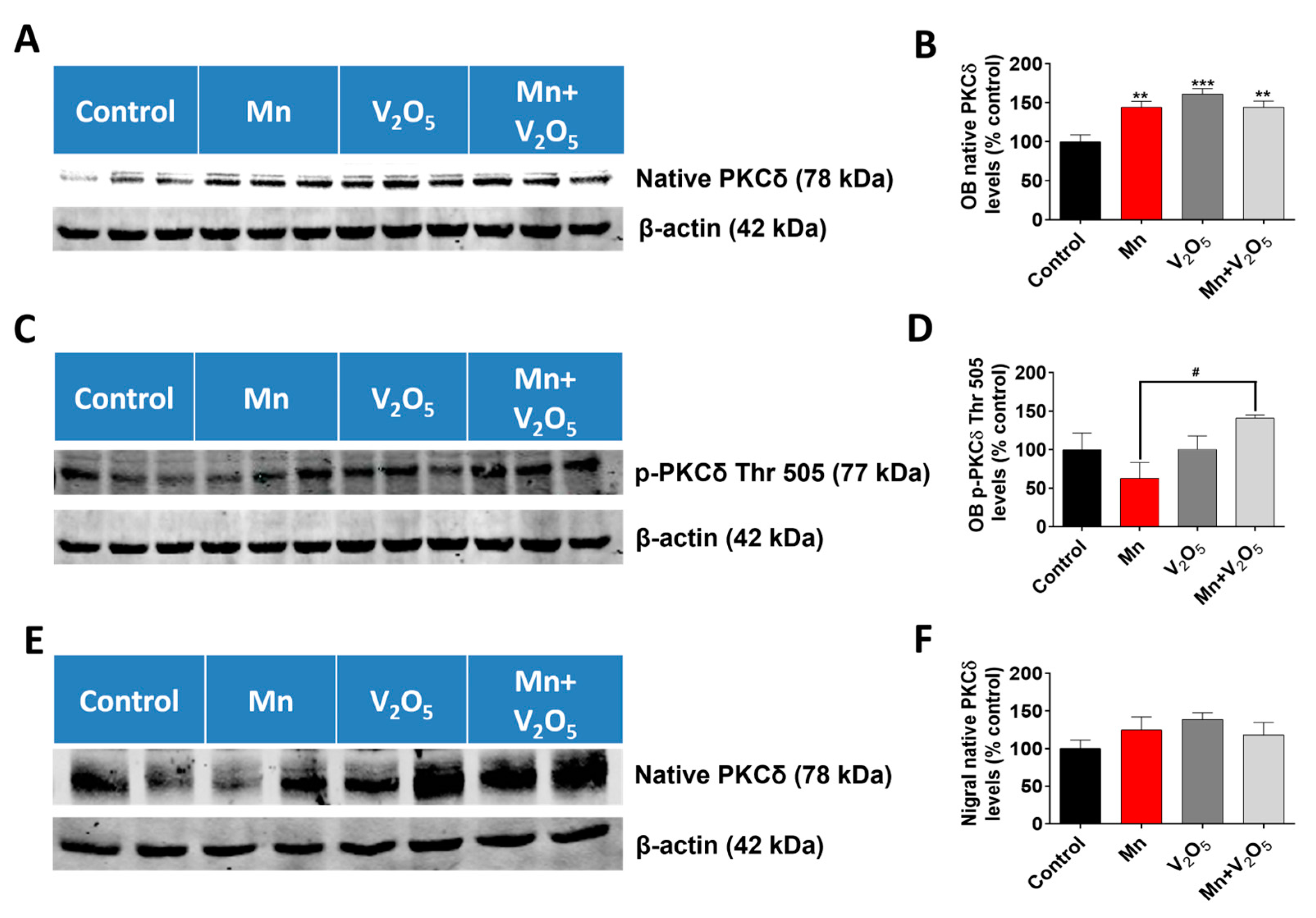
2.9. Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Induced Upregulation of the Pro-Apoptotic Kinase PKCδ in the Olfactory Bulb
2.10. Chronic Intranasal Exposure to Mn, V2O5, or Their Mixture (Mn/V2O5) Causes Astrocytes to Accumulate in the Glomerular Layer of the Olfactory Bulb
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Locomotor Activity
4.3. Animals
4.4. Weighing Olfactory Bulbs
4.5. Olfaction Test
4.6. HPLC Detection of Dopamine
4.7. Western Blot
4.8. Immunohistological Analysis of Olfactory Bulb Sections
4.9. Measurement of Metal Levels
4.10. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Zoroddu, M.A.; Aaseth, J.; Crisponi, G.; Medici, S.; Peana, M.; Nurchi, V.M. The essential metals for humans: A brief overview. J. Inorg. Biochem. 2019, 195, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Carmona, A.; Roudeau, S.; Ortega, R. Molecular Mechanisms of Environmental Metal Neurotoxicity: A Focus on the Interactions of Metals with Synapse Structure and Function. Toxics 2021, 9, 198. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L.; Hastings, L. Neurotoxic exposure and olfactory impairment. Clin. Occup. Environ. Med. 2001, 1, 547–575. [Google Scholar]
- Antunes, M.B.; Bowler, R.; Doty, R.L. San Francisco/Oakland Bay bridge welder study: Olfactory function. Neurology 2007, 69, 1278–1284. [Google Scholar] [CrossRef]
- Mascagni, P.; Consonni, D.; Bregante, G.; Chiappino, G.; Toffoletto, F. Olfactory Function in Workers Exposed to Moderate Airborne Cadmium Levels. NeuroToxicology 2003, 24, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Kothapalli, C.R. Differential impact of heavy metals on neurotoxicity during development and in aging central nervous system. Curr. Opin. Toxicol. 2021, 26, 33–38. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef]
- Jan, A.T.; Azam, M.; Siddiqui, K.; Ali, A.; Choi, I.; Haq, Q.M.R. Heavy metals and human health: Mechanistic insight into toxicity and counter defense system of antioxidants. Int. J. Mol. Sci. 2015, 16, 29592–29630. [Google Scholar] [CrossRef]
- Willis, A.W.; Evanoff, B.A.; Lian, M.; Galarza, A.; Wegrzyn, A.; Schootman, M.; Racette, B.A. Metal Emissions and Urban Incident Parkinson Disease: A Community Health Study of Medicare Beneficiaries by Using Geographic Information Systems. Am. J. Epidemiol. 2010, 172, 1357–1363. [Google Scholar] [CrossRef]
- Mortimer, J.A.; Borenstein, A.R.; Nelson, L.M. Associations of welding and manganese exposure with Parkinson disease: Review and meta-analysis. Neurology 2012, 79, 1174–1180. [Google Scholar] [CrossRef]
- Aschner, M.; Erikson, K.M.; Hernández, E.H.; Tjalkens, R. Manganese and its role in Parkinson’s disease: From transport to neuropathology. Neuromolecular Med. 2009, 11, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Benedetto, A.; Au, C.; Aschner, M. Manganese-Induced Dopaminergic Neurodegeneration: Insights into Mechanisms and Genetics Shared with Parkinson’s Disease. Chem. Rev. 2009, 109, 4862–4884. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, T.; Tan, X.; Luo, Y.; Kanda, H. Relationship between Blood Levels of Heavy Metals and Parkinson’s Disease in China. Neuroepidemiology 2009, 34, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Racette, B.A.; Criswell, S.R.; Lundin, J.I.; Hobson, A.; Seixas, N.; Kotzbauer, P.T.; Evanoff, B.A.; Perlmutter, J.S.; Zhang, J.; Sheppard, L.; et al. Increased risk of parkinsonism associated with welding exposure. NeuroToxicology 2012, 33, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Neal, A.P.; Guilarte, T.R. Mechanisms of lead and manganese neurotoxicity. Toxicol. Res. 2013, 2, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Hutchens, S.; Liu, C.; Jursa, T.; Shawlot, W.; Aschner, M.; Smith, D.R.; Mukhopadhyay, S. SLC30A10 transporter in the digestive system regulates brain manganese under basal conditions while brain SLC30A10 protects against neurotoxicity. J. Biol. Chem. 2019, 294, 1860–1876. [Google Scholar] [CrossRef]
- O’neal, S.L.; Zheng, W. Manganese Toxicity Upon Overexposure: A Decade in Review. Curr. Environ. Health Rep. 2015, 2, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Mergler, D. Neurotoxic Effects of Low Level Exposure to Manganese in Human Populations. Environ. Res. 1999, 80, 99–102. [Google Scholar] [CrossRef]
- Pal, P.K.; Samii, A.; Calne, D.B. Manganese neurotoxicity: A review of clinical features, imaging and pathology. Neurotoxicology 1999, 20, 227–238. [Google Scholar] [PubMed]
- Chillrud, S.N.; Epstein, D.; Ross, J.M.; Sax, S.N.; Pederson, D.; Spengler, J.D.; Kinney, P.L. Elevated airborne exposures of teenagers to manganese, chromium, and iron from steel dust and New York City’s subway system. Environ. Sci. Technol. 2004, 38, 732–737. [Google Scholar] [CrossRef]
- Mattison, D.R.; Milton, B.; Krewski, D.; Levy, L.; Dorman, D.C.; Aggett, P.J.; Roels, H.A.; Andersen, M.E.; Karyakina, N.A.; Shilnikova, N.; et al. Severity scoring of manganese health effects for categorical regression. NeuroToxicology 2016, 58, 203–216. [Google Scholar] [CrossRef]
- Milton, B.; Krewski, D.; Mattison, D.R.; Karyakina, N.A.; Ramoju, S.; Shilnikova, N.; Birkett, N.; Farrell, P.J.; McGough, D. Modeling U-shaped dose-response curves for manganese using categorical regression. NeuroToxicology 2017, 58, 217–225. [Google Scholar] [CrossRef]
- Peres, T.V.; Parmalee, N.L.; Martinez-Finley, E.J.; Aschner, M. Untangling the Manganese-α-Synuclein Web. Front. Neurosci. 2016, 10, 364. [Google Scholar] [CrossRef]
- Chen, P.; Culbreth, M.; Aschner, M. Exposure, epidemiology, and mechanism of the environmental toxicant manganese. Environ. Sci. Pollut. Res. 2016, 23, 13802–13810. [Google Scholar] [CrossRef]
- Caito, S.; Aschner, M. Neurotoxicity of metals. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 131, pp. 169–189. [Google Scholar] [CrossRef]
- Brenneman, K.A.; Cattley, R.C.; Ali, S.F.; Dorman, D.C. Manganese-induced developmental neurotoxicity in the CD rat: Is oxidative damage a mechanism of action? Neurotoxicology 1999, 20, 477–487. [Google Scholar] [PubMed]
- Calne, D.B.; Chu, N.S.; Huang, C.C.; Lu, C.S.; Olanow, W. Manganism and idiopathic parkinsonism: Similarities and differences. Neurology 1994, 44, 1583. [Google Scholar] [CrossRef]
- Eriksson, H.; Tedroff, J.; Thuomas, K.; Aquilonius, S.-M.; Hartvig, P.; Fasth, K.-J.; Bjurling, P.; Långström, B.; Hedström, K.-G.; Heilbronn, E. Manganese induced brain lesions inMacaca fascicularis as revealed by positron emission tomography and magnetic resonance imaging. Arch. Toxicol. 1992, 66, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, S.; Umehara, F.; Hanada, K.; Nobuhara, Y.; Takenaga, S.; Arimura, K.; Osame, M. Manganese intoxication during total parenteral nutrition: Report of two cases and review of the literature. J. Neurol. Sci. 1999, 162, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Bowman, A.B.; Kwakye, G.F.; Hernández, E.H.; Aschner, M. Role of manganese in neurodegenerative diseases. J. Trace Elem. Med. Biol. 2011, 25, 191–203. [Google Scholar] [CrossRef]
- Verity, M.A. Manganese neurotoxicity: A mechanistic hypothesis. Neurotoxicology 1999, 20, 489–497. [Google Scholar]
- Parenti, M.; Flauto, C.; Parati, E.; Vescovi, A.; Groppetti, A. Manganese neurotoxicity: Effects ofl-DOPA and pargyline treatments. Brain Res. 1986, 367, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Vescovi, A.; Facheris, L.; Zaffaroni, A.; Malanca, G.; Parati, E. Dopamine metabolism alterations in a manganese-treated pheochromocytoma cell line (PC12). Toxicology 1991, 67, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Dorman, D.C.; Struve, M.F.; James, R.; Marshall, M.W.; Parkinson, C.U.; Wong, B.A. Influence of Particle Solubility on the Delivery of Inhaled Manganese to the Rat Brain: Manganese Sulfate and Manganese Tetroxide Pharmacokinetics Following Repeated (14-Day) Exposure. Toxicol. Appl. Pharmacol. 2001, 170, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Lyon, F. Cobalt in Hard Metals and Cobalt Sulfate, Gallium Arsenide, Indium Phosphide and Vanadium Pentoxide. IARC Monogr. Eval. Carcinog. Risks Hum. 2006, 86, 227–292. [Google Scholar]
- Kiviluoto, M.; Pyy, L.; Pakarinen, A. Serum and urinary vanadium of vanadium-exposed workers. Scand. J. Work. Environ. Health 1979, 5, 362–367. [Google Scholar] [CrossRef]
- Ngwa, H.A.; Kanthasamy, A.; Anantharam, V.; Song, C.; Witte, T.; Houk, R.; Kanthasamy, A.G. Vanadium induces dopaminergic neurotoxicity via protein kinase Cdelta dependent oxidative signaling mechanisms: Relevance to etiopathogenesis of Parkinson’s disease. Toxicol. Appl. Pharmacol. 2009, 240, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Ngwa, H.A.; Kanthasamy, A.; Jin, H.; Anantharam, V.; Kanthasamy, A.G. Vanadium exposure induces olfactory dysfunction in an animal model of metal neurotoxicity. NeuroToxicology 2013, 43, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Avila-Costa, M.R.; Colín-Barenque, L.; Zepeda-Rodríguez, A.; Antuna, S.B.; Saldivar, L.; Espejel-Maya, G.; Mussali-Galante, P.; del Carmen Avila-Casado, M.; Reyes-Olivera, A.; Anaya-Martinez, V.; et al. Ependymal epithelium disruption after vanadium pentoxide inhalation: A mice experimental model. Neurosci. Lett. 2005, 381, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Avila-Costa, M.R.; Fortoul, T.I.; Niño-Cabrera, G.; Colín-Barenque, L.; Bizarro-Nevares, P.; Gutiérrez-Valdez, A.L.; Ordóñez-Librado, J.L.; Rodríguez-Lara, V.; Mussali-Galante, P.; Díaz-Bech, P.; et al. Hippocampal cell alterations induced by the inhalation of vanadium pentoxide (V2O5) promote memory deterioration. NeuroToxicology 2006, 27, 1007–1012. [Google Scholar] [CrossRef]
- Avila-Costa, M.R.; Flores, E.M.; Colin-Barenque, L.; Ordoñez, J.L.; Gutiérrez, A.L.; Niño-Cabrera, H.G.; Mussali-Galante, P.; Fortoul, T.I. Nigrostriatal Modifications After Vanadium Inhalation: An Immunocytochemical and Cytological Approach. Neurochem. Res. 2004, 29, 1365–1369. [Google Scholar] [CrossRef]
- Fatola, O.I.; Olaolorun, F.A.; Olopade, F.E.; Olopade, J.O. Trends in vanadium neurotoxicity. Brain Res. Bull. 2019, 145, 75–80. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, J.D.; Heal, M.R.; Beverland, I.J.; Howe, A.; Gibson, M.D.; Hibbs, L.R.; MacNee, W.; Donaldson, K. Soluble transition metals cause the pro-inflammatory effects of welding fumes in vitro. Toxicol. Appl. Pharmacol. 2004, 196, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Nemery, B. Metal toxicity and the respiratory tract. Eur. Respir. J. 1990, 3, 202–219. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, J.A.; Du, H. Re: Vanadium exposure-induced neurobehavioral alterations among Chinese workers Li et al. (2013). NeuroToxicology 2014, 44, 369–370. [Google Scholar] [CrossRef] [PubMed]
- Adrian, H. A mechanism for effect of vanadium on hardenability of medium carbon manganese steel. Mater. Sci. Technol. 1999, 15, 366–378. [Google Scholar] [CrossRef]
- Latchoumycandane, C.; Anantharam, V.; Kitazawa, M.; Yang, Y.; Kanthasamy, A.; Kanthasamy, A.G. Protein kinase Cδ is a key downstream mediator of manganese-induced apoptosis in dopaminergic neuronal cells. J. Pharmacol. Exp. Ther. 2005, 313, 46–55. [Google Scholar] [CrossRef]
- Davila, N.G.; Blakemore, L.J.; Trombley, P.Q. Dopamine Modulates Synaptic Transmission Between Rat Olfactory Bulb Neurons in Culture. J. Neurophysiol. 2003, 90, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Johansson, O.; Goldstein, M.; Halász, N.; Hökfelt, T.; Ljungdahl, Å. Immunohistochemical identification of two types of dopamine neuron in the rat olfactory bulb as seen by serial sectioning. J. Neurocytol. 1981, 10, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Adam, Y.; Mizrahi, A. Long-Term Imaging Reveals Dynamic Changes in the Neuronal Composition of the Glomerular Layer. J. Neurosci. 2011, 31, 7967–7973. [Google Scholar] [CrossRef]
- Hsia, A.Y.; Vincent, J.-D.; Lledo, P.-M.; Wang, Z.-J.; Hu, S.S.-J.; Bradshaw, H.B.; Sun, L.; Mackie, K.; Straiker, A.; Heinbockel, T.; et al. Dopamine Depresses Synaptic Inputs Into the Olfactory Bulb. J. Neurophysiol. 1999, 82, 1082–1085. [Google Scholar] [CrossRef]
- Tillerson, J.L.; Caudle, W.M.; Parent, J.M.; Gong, C.; Schallert, T.; Miller, G.W. Olfactory discrimination deficits in mice lacking the dopamine transporter or the D2 dopamine receptor. Behav. Brain Res. 2006, 172, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Liu, S. Dopaminergic Modulation of Glomerular Circuits in the Mouse Olfactory Bulb. Front. Cell. Neurosci. 2020, 14, 172. [Google Scholar] [CrossRef] [PubMed]
- Kanthasamy, A.G.; Kitazawa, M.; Kanthasamy, A.; Anantharam, V. Role of proteolytic activation of protein kinase Cδ in oxidative stress-induced apoptosis. Antioxid. Redox Signal. 2003, 5, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Tansey, M.G.; McCoy, M.K.; Frank-Cannon, T.C. Neuroinflammatory mechanisms in Parkinson’s disease: Potential environmental triggers, pathways, and targets for early therapeutic intervention. Exp. Neurol. 2007, 208, 1–25. [Google Scholar] [CrossRef]
- Whitton, P.S. Inflammation as a causative factor in the aetiology of Parkinson’s disease. Br. J. Pharmacol. 2007, 150, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H.V.; Morris, H.R. Pathogenetic insights into young-onset Parkinson disease. Nat. Rev. Neurol. 2020, 16, 245–246. [Google Scholar] [CrossRef]
- Schneider, S.A.; Hizli, B.; Alcalay, R.N. Emerging Targeted Therapeutics for Genetic Subtypes of Parkinsonism. Neurotherapeutics 2020, 17, 1378–1392. [Google Scholar] [CrossRef]
- Racette, B.A.; McGee-Minnich, L.; Moerlein, S.M.; Mink, J.W.; Videen, T.O.; Perlmutter, J.S. Welding-related parkinsonism: Clinical features, treatment, and pathophysiology. Neurology 2001, 56, 8–13. [Google Scholar] [CrossRef]
- Park, R.M.; Schulte, P.A.; Bowman, J.D.; Walker, J.T.; Bondy, S.C.; Yost, M.G.; Touchstone, J.A.; Dosemeci, M. Potential occupational risks for neurodegenerative diseases. Am. J. Ind. Med. 2005, 48, 63–77. [Google Scholar] [CrossRef]
- Guilarte, T.R. Manganese and Parkinson’s disease: A critical review and new findings. Environ. Health Perspect. 2010, 118, 1071–1080. [Google Scholar] [CrossRef]
- Živančević, K.; Baralić, K.; Jorgovanović, D.; Djordjević, A.B.; Ćurčić, M.; Miljaković, E.A.; Antonijević, B.; Bulat, Z.; Đukić-Ćosić, D. Elucidating the influence of environmentally relevant toxic metal mixture on molecular mechanisms involved in the development of neurodegenerative diseases: In silico toxicogenomic data-mining. Environ. Res. 2021, 194, 110727. [Google Scholar] [CrossRef] [PubMed]
- Bunting, R.M. Vanadium: How market developments affect the titanium industry. Strategic minerals corporation. In Proceedings of the Titanium 2006, International Titanium Association Conference, San Diego, CA, USA, 3 October 2006. [Google Scholar]
- Imtiaz, M.; Rizwan, M.S.; Xiong, S.; Li, H.; Ashraf, M.; Shahzad, S.M.; Shahzad, M.; Rizwan, M.; Tu, S. Vanadium, recent advancements and research prospects: A review. Environ. Int. 2015, 80, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Han, M.J.; Ozaki, T.; Yu, J. Electronic structure and magnetic properties of small manganese oxide clusters. J. Chem. Phys. 2005, 123, 34306. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lee, C.Y.; Marschilok, A.C.; Takeuchi, K.J.; Takeuchi, E.S. AgxVOPO4: A demonstration of the dependence of battery-related electrochemical properties of silver vanadium phosphorous oxides on Ag/V ratios. J. Power Sources 2011, 196, 3325–3330. [Google Scholar] [CrossRef] [PubMed]
- Marschilok, A.C.; Kozarsky, E.S.; Tanzil, K.; Zhu, S.; Takeuchi, K.J.; Takeuchi, E.S. Electrochemical reduction of silver vanadium phosphorous oxide, Ag2VO2PO4: Silver metal deposition and associated increase in electrical conductivity. J. Power Source 2010, 195, 6839–6846. [Google Scholar] [CrossRef] [PubMed]
- Hossain, H.; Abdullah, N.; Tan, K.H.; Saidur, R.; Radzi, M.A.M.; Shafie, S. Evolution of Vanadium Redox Flow Battery in Electrode. Chem. Rec. 2023, 24, e202300092. [Google Scholar] [CrossRef] [PubMed]
- Mousavihashemi, S.; Murcia-López, S.; Rodriguez-Olguin, M.A.; Gardeniers, H.; Andreu, T.; Morante, J.R.; Arce, A.S.; Flox, C. Overcoming Voltage Losses in Vanadium Redox Flow Batteries Using WO3 as a Positive Electrode. ChemCatChem 2022, 14, e202201106. [Google Scholar] [CrossRef]
- Ye, Z.; Chen, N.; Zheng, Z.; Xiong, L.; Chen, D. Preparation of Sulfonated Poly (arylene ether)/SiO2 Composite Membranes with Enhanced Proton Selectivity for Vanadium Redox Flow Batteries. Molecules 2023, 28, 3130. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yan, J.; Hong, Y.; Yu, Z.; Chen, J.; Zheng, J. Ultrahigh-rate and ultralong-life aqueous batteries enabled by special pair-dancing proton transfer. Sci. Adv. 2023, 9, eadf4589. [Google Scholar] [CrossRef]
- Dobson, A.W.; Erikson, K.M.; Aschner, M. Manganese Neurotoxicity. Ann. N. Y. Acad. Sci. 2004, 1012, 115–128. [Google Scholar] [CrossRef]
- Lucchini, R.; Tieu, K. Manganese-Induced Parkinsonism: Evidence from Epidemiological and Experimental Studies. Biomolecules 2023, 13, 1190. [Google Scholar] [CrossRef] [PubMed]
- Avila, D.S.; Puntel, R.L.; Aschner, M. Manganese in health and disease. In Interrelations between Essential Metal Ions and Human Diseases; Sigel, A., Sigel, H., Sigel, R., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 199–227. [Google Scholar] [CrossRef]
- Pajarillo, E.; Nyarko-Danquah, I.; Digman, A.; Multani, H.K.; Kim, S.; Gaspard, P.; Aschner, M.; Lee, E. Mechanisms of manganese-induced neurotoxicity and the pursuit of neurotherapeutic strategies. Front. Pharmacol. 2022, 13, 1011947. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.C.; Gubert, P.; Boas, G.R.V.; Paes, M.M.; Santamaría, A.; Lee, E.; Tinkov, A.A.; Bowman, A.B.; Aschner, M. Manganese-induced neurodegenerative diseases and possible therapeutic approaches. Expert Rev. Neurother. 2020, 20, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, A.M.; Santos, M.L.; Batoréu, M.C.; Aschner, M. Prolactin is a peripheral marker of manganese neurotoxicity. Brain Res. 2011, 1382, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Dlamini, W.W.; Nelson, G.; Nielsen, S.S.; Racette, B.A. Manganese exposure, parkinsonian signs, and quality of life in South African mine workers. Am. J. Ind. Med. 2019, 63, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Criswell, S.R.M.; Nielsen, S.S.; Warden, M.N.; Flores, H.P.; Lenox-Krug, J.; Racette, S.; Sheppard, L.; Checkoway, H.; Racette, B.A. MRI Signal Intensity and Parkinsonism in Manganese-Exposed Workers. J. Occup. Environ. Med. 2019, 61, 641–645. [Google Scholar] [CrossRef]
- Criswell, S.R.; Perlmutter, J.S.; Huang, J.L.; Golchin, N.; Flores, H.P.; Hobson, A.; Aschner, M.; Erikson, K.M.; Checkoway, H.; Racette, B.A. Basal ganglia intensity indices and diffusion weighted imaging in manganese-exposed welders. Occup. Environ. Med. 2012, 69, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Powers, K.M.; Smith-Weller, T.; Franklin, G.M.; Longstreth, W.T., Jr.; Swanson, P.D.; Checkoway, H. Parkinson’s disease risks associated with dietary iron, manganese, and other nutrient intakes. Neurology 2003, 60, 1761–1766. [Google Scholar] [CrossRef] [PubMed]
- Gorell, J.M.; Johnson, C.C.; Rybicki, B.A.; Peterson, E.L.; Richardson, R.J. The risk of Parkinson’s disease with exposure to pesticides, farming, well water, and rural living. Neurology 1998, 50, 1346–1350. [Google Scholar] [CrossRef]
- Gorell, J.M.; Johnson, C.C.; Rybicki, B.A.; Peterson, E.L.; Kortsha, G.X.; Brown, G.G.; Richardson, R.J. Occupational exposure to manganese, copper, lead, iron, mercury and zinc and the risk of Parkinson’s disease. Neurotoxicology 1999, 20, 239–247. [Google Scholar]
- Gorell, J.M.; Johnson, C.C.; Rybicki, B.A.; Peterson, E.L.; Kortsha, G.X.; Brown, G.G.; Richardson, R.J. Occupational exposures to metals as risk factors for Parkinson’s disease. Neurology 1997, 48, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Dexter, D.T.; Jenner, P.; Schapira, A.H.V.; Marsden, C.D. The Royal Kings and Queens Parkinson’s Disease Research Group Alterations in levels of iron, ferritin, and other trace metals in neurodegenerative diseases affecting the basal ganglia. Ann. Neurol. 1992, 32, S94–S100. [Google Scholar] [CrossRef] [PubMed]
- Dexter, D.T.; Wells, F.R.; Lee, A.J.; Agid, F.; Agid, Y.; Jenner, P.; Marsden, C.D. Increased Nigral Iron Content and Alterations in Other Metal Ions Occurring in Brain in Parkinson’s Disease. J. Neurochem. 1989, 52, 1830–1836. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.V.; Ali, M.M.; Saxena, D.K.; Murthy, R.C. Behavioral and neurochemical changes in rats simultaneously exposed to manganese and lead. Arch. Toxicol. 1981, 49, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.V.; Murthy, R.; Saxena, D.; Lal, B. Effects of pre- and postnatal combined exposure to Pb and Mn on brain development in rats. Ind. Health 1983, 21, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Mejía, J.; Díaz-Barriga, F.; Calderón, J.; Ríos, C.; Jiménez-Capdeville, M. Effects of Lead–Arsenic Combined Exposure on Central Monoaminergic Systems. Neurotoxicol. Teratol. 1997, 19, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Kalia, K.; Murthy, R.; Chandra, S.V. Tissue disposition of 54Mn in lead pretreated rats. Ind. Health 1984, 22, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.O.; Baccarelli, A. Metals and Neurotoxicology. J. Nutr. 2007, 137, 2809–2813. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, V.M.; Dufour, L.; Carrizales, L.; Díaz-Barriga, F.; Jiménez-Capdeville, M.E. Effects of oral exposure to mining waste on in vivo dopamine release from rat striatum. Environ. Health Perspect. 1998, 106, 487–491. [Google Scholar]
- Kanthasamy, A.; Jin, H.; Mehrotra, S.; Mishra, R.; Kanthasamy, A.; Rana, A. Novel cell death signaling pathways in neurotoxicity models of dopaminergic degeneration: Relevance to oxidative stress and neuroinflammation in Parkinson’s disease. NeuroToxicology 2010, 31, 555–561. [Google Scholar] [CrossRef]
- Kaul, S.; Ananrharam, V.; Kanthasamy, A. Low dose oxidative insult promotes apoptosis in dopaminergic cells via caspase-3 dependent proteolytic activation of PKC delta: Relevance to environmental factors and Parkinson’s disease. Toxicol. Sci. 2003, 72, 79. [Google Scholar]
- Kitazawa, M.; Anantharam, V.; Kanthasamy, A.G. Translocation of protein kinase CS to mitochondria promotes proteolytic inactivation of Bcl-2 during MMT-induced apoptosis in dopaminergic cells. Neurotoxicology 2003, 24, 305–306. [Google Scholar]
- Yoritaka, A.; Hattori, N.; Uchida, K.; Tanaka, M.; Stadtman, E.R.; Mizuno, Y. Immunohistochemical detection of 4-hydroxynonenal protein adducts in Parkinson disease. Proc. Natl. Acad. Sci. USA 1996, 93, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.K. Oxidative stress in neurodegeneration: Cause or consequence? Nat. Med. 2004, 10 (Suppl. 7), S18–S25. [Google Scholar] [CrossRef] [PubMed]
- Kanthasamy, A.; Kitazawa, M.; Kaul, S.; Anantharam, V.; Kanthasamy, A.G. Methylcyclopentadienyl manganese tricarbonyl (MMT) induces apoptosis by PKC delta dependent activation of NF-kappa B in mesencephalic dopaminergic neuronal cells. Toxicol. Sci. 2003, 72, 353. [Google Scholar]
- Reyland, M.E.; Jones, D.N. Multifunctional roles of PKCδ: Opportunities for targeted therapy in human disease. Pharmacol. Ther. 2016, 165, 1–13. [Google Scholar] [CrossRef]
- Brodie, C.; Blumberg, P.M. Regulation of cell apoptosis by protein kinase c δ. Apoptosis 2003, 8, 19–27. [Google Scholar] [CrossRef]
- Steinberg, S.F. Distinctive activation mechanisms and functions for protein kinase Cδ. Biochem. J. 2004, 384, 449–459. [Google Scholar] [CrossRef]
- Gordon, R.; Singh, N.; Lawana, V.; Ghosh, A.; Harischandra, D.S.; Jin, H.; Hogan, C.; Sarkar, S.; Rokad, D.; Panicker, N.; et al. Protein kinase Cδ upregulation in microglia drives neuroinflammatory responses and dopaminergic neurodegeneration in experimental models of Parkinson’s disease. Neurobiol. Dis. 2016, 93, 96–114. [Google Scholar] [CrossRef]
- Anantharam, V.; Kitazawa, M.; Wagner, J.; Kaul, S.; Kanthasamy, A.G. Caspase-3-dependent proteolytic cleavage of protein kinase Cδ is essential for oxidative stress-mediated dopaminergic cell death after exposure to methylcyclopentadienyl manganese tricarbonyl. J. Neurosci. 2002, 22, 1738–1751. [Google Scholar] [CrossRef]
- Kanthasamy, A.G.; Kitazawa, M.; Kaul, S.; Yang, Y.; Lahiri, D.K.; Anantharam, V.; Kanthasamy, A. Proteolytic Activation of Proapoptotic Kinase PKCδ Is Regulated by Overexpression of Bcl-2: Implications for Oxidative Stress and Environmental Factors in Parkinson’s Disease. Ann. N. Y. Acad. Sci. 2003, 1010, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Mesholam, R.I.; Moberg, P.J.; Mahr, R.N.; Doty, R.L. Olfaction in neurodegenerative disease: A meta-analysis of olfactory functioning in Alzheimer’s and Parkinson’s diseases. Arch. Neurol. 1998, 55, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L.; Stern, M.B.; Pfeiffer, C.; Gollomp, S.M.; Hurtig, H.I. Bilateral olfactory dysfunction in early stage treated and untreated idiopathic Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 1992, 55, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Tissingh, G.; Booij, J.; Bergmans, P.; Winogrodzka, A.; Janssen, A.G.; van Royen, E.A.; Wolters, E.C. Iodine-123-N-ω-fluoropropyl-2β-carbomethoxy-3β-(4-iodophenyl) tropane SPECT in healthy controls and early-stage, drug-naive Parkinson’s disease. J. Nucl. Med. 1998, 39, 1143–1148. [Google Scholar] [PubMed]
- Ward, C.D.; Hess, W.A.; Calne, D.B. Olfactory impairment in Parkinson’s disease. Neurology 1983, 33, 943. [Google Scholar] [CrossRef] [PubMed]
- Rombaux, P.; Mouraux, A.; Bertrand, B.; Nicolas, G.; Duprez, T.; Hummel, T. Olfactory Function and Olfactory Bulb Volume in Patients with Postinfectious Olfactory Loss. Laryngoscope 2006, 116, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Yousem, D.M.; Geckle, R.J.; Bilker, W.B.; Kroger, H.; Doty, R.L. Posttraumatic smell loss: Relationship of psychophysica tests and volumes of the olfactory bulbs and tracts and the temporal lobes. Acad. Radiol. 1999, 6, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Choi, W.; Jeong, H.-Y.; Lee, J.; Kim, J. MR Imaging–Based Evaluations of Olfactory Bulb Atrophy in Patients with Olfactory Dysfunction. Am. J. Neuroradiol. 2017, 39, 532–537. [Google Scholar] [CrossRef]
- Wang, J.; You, H.; Liu, J.-F.; Ni, D.-F.; Zhang, Z.-X.; Guan, J. Association of Olfactory Bulb Volume and Olfactory Sulcus Depth with Olfactory Function in Patients with Parkinson Disease. Am. J. Neuroradiol. 2011, 32, 677–681. [Google Scholar] [CrossRef]
- Rombaux, P.; Huart, C.; Deggouj, N.; Duprez, T.; Hummel, T. Prognostic Value of Olfactory Bulb Volume Measurement for Recovery in Postinfectious and Posttraumatic Olfactory Loss. Otolaryngol. Neck Surg. 2012, 147, 1136–1141. [Google Scholar] [CrossRef]
- Huisman, E.; Uylings, H.B.; Hoogland, P.V. Gender-related changes in increase of dopaminergic neurons in the olfactory bulb of Parkinson’s disease patients. Mov. Disord. 2008, 23, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.; Steur, E.N.J.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Harding, A.J.; Stimson, E.; Henderson, J.M.; Halliday, G.M. Clinical correlates of selective pathology in the amygdala of patients with Parkinson’s disease. Brain 2002, 125, 2431–2445. [Google Scholar] [CrossRef]
- Pearce, R.K.B.; Hawkes, C.H.; Daniel, S.E. The anterior olfactory nucleus in Parkinson’s disease. Mov. Disord. 1995, 10, 283–287. [Google Scholar] [CrossRef]
- Silveira-Moriyama, L.; Holton, J.L.; Kingsbury, A.; Ayling, H.; Petrie, A.; Sterlacci, W.; Poewe, W.; Maier, H.; Lees, A.J.; Revesz, T. Regional differences in the severity of Lewy body pathology across the olfactory cortex. Neurosci. Lett. 2009, 453, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Ross, G.W.; Petrovitch, H.; Abbott, R.D.; Tanner, C.M.; Popper, J.; Masaki, K.; Launer, L.; White, L.R. Association of olfactory dysfunction with risk for future Parkinson’s disease. Ann. Neurol. 2008, 63, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Atianjoh, F.E.; Ladenheim, B.; Krasnova, I.N.; Cadet, J.L. Amphetamine causes dopamine depletion and cell death in the mouse olfactory bulb. Eur. J. Pharmacol. 2008, 589, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Ladenheim, B.; Jayanthi, S.; Cadet, J.L. Methamphetamine Administration Causes Death of Dopaminergic Neurons in the Mouse Olfactory Bulb. Biol. Psychiatry 2007, 61, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.; Kawano, T.; Margolis, F.; Joh, T. Transneuronal regulation of tyrosine hydroxylase expression in olfactory bulb of mouse and rat. J. Neurosci. 1983, 3, 69–78. [Google Scholar] [CrossRef]
- Lledo, P.-M.; Alonso, M.; Grubb, M.S. Adult neurogenesis and functional plasticity in neuronal circuits. Nat. Rev. Neurosci. 2006, 7, 179–193. [Google Scholar] [CrossRef]
- Schröder, H.; Moser, N.; Huggenberger, S. The Mouse Olfactory System. In Neuroanatomy of the Mouse: An Introduction; Schröder, H., Moser, N., Huggenberger, S., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 319–331. [Google Scholar]
- Doty, R.L.; Risser, J.M. Influence of the D-2 dopamine receptor agonist quinpirole on the odor detection performance of rats before and after spiperone administration. Psychopharmacology 1989, 98, 310–315. [Google Scholar] [CrossRef]
- Duchamp-Viret, P.; Coronas, V.; Delaleu, J.-C.; Moyse, E.; Duchamp, A. Dopaminergic modulation of mitral cell activity in the frog olfactory bulb: A combined radioligand binding–electrophysiological study. Neuroscience 1997, 79, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Koster, N.; Norman, A.; Richtand, N.; Nickell, W.; Puche, A.; Pixley, S.; Shipley, M. Olfactory receptor neurons express D2 dopamine receptors. J. Comp. Neurol. 1999, 411, 666–673. [Google Scholar] [CrossRef]
- Wilson, D.; Sullivan, R. The D2 antagonist spiperone mimics the effects of olfactory deprivation on mitral/tufted cell odor response patterns. J. Neurosci. 1995, 15, 5574–5581. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C. Olfaction in neurodegenerative disorder. Mov. Disord. 2003, 18, 364–372. [Google Scholar] [CrossRef]
- Salehi, F.; Krewski, D.; Mergler, D.; Normandin, L.; Kennedy, G.; Philippe, S.; Zayed, J. Bioaccumulation and locomotor effects of manganese phosphate/sulfate mixture in Sprague-Dawley rats following subchronic (90 days) inhalation exposure. Toxicol. Appl. Pharmacol. 2003, 191, 264–271. [Google Scholar] [CrossRef]
- Tapin, D.; Kennedy, G.; Lambert, J.; Zayed, J. Bioaccumulation and locomotor effects of manganese sulfate in Sprague–Dawley rats following subchronic (90 days) inhalation exposure. Toxicol. Appl. Pharmacol. 2006, 211, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Normandin, L.; Beaupré, L.A.; Salehi, F.; Pierre, A.S.; Kennedy, G.; Mergler, D.; Butterworth, R.F.; Philippe, S.; Zayed, J. Manganese distribution in the brain and neurobehavioral changes following inhalation exposure of rats to three chemical forms of manganese. Neurotoxicology 2004, 25, 433–441. [Google Scholar] [CrossRef]
- Normandin, L.; Carrier, G.; Gardiner, P.F.; Kennedy, G.; Hazell, A.S.; Mergler, D.; Butterworth, R.F.; Philippe, S.; Zayed, J. Assessment of bioaccumulation, neuropathology, and neurobehavior following subchronic (90 days) inhalation in Sprague–Dawley rats exposed to manganese phosphate. Toxicol. Appl. Pharmacol. 2002, 183, 135–145. [Google Scholar] [CrossRef]
- Vezér, T.; Kurunczi, A.; Náray, M.; Papp, A.; Nagymajtényi, L. Behavioral effects of subchronic inorganic manganese exposure in rats. Am. J. Ind. Med. 2007, 50, 841–852. [Google Scholar] [CrossRef]
- Dorman, D.C.; Brenneman, K.A.; McElveen, A.M.; Lynch, S.E.; Roberts, K.C.; Wong, B.A. Olfactory Transport: A Direct Route Of Delivery Of Inhaled Manganese Phosphate To The Rat Brain. J. Toxicol. Environ. Health Part A 2002, 65, 1493–1511. [Google Scholar] [CrossRef] [PubMed]
- Dorman, D.C.; Struve, M.F.; Vitarella, D.; Byerly, F.L.; Goetz, J.; Miller, R. Neurotoxicity of manganese chloride in neonatal and adult CD rats following subchronic (21-day) high-dose oral exposure. J. Appl. Toxicol. 2000, 20, 179–187. [Google Scholar]
- Done, A.K. Of metals and chelation. Emer. Med. 1979, 11, 186–218. [Google Scholar]
- WHO. Vanadium. In Chapter 6.12; WHO Regional Office for Europe: Copenhagen, Denmark, 2000; p. 9. [Google Scholar]
- Hashimoto, M.; Masliah, E. Alpha-synuclein in Lewy Body Disease and Alzheimer’s Disease. Brain Pathol. 1999, 9, 707–720. [Google Scholar] [CrossRef]
- Takeda, A.; Hashimoto, M.; Mallory, M.; Sundsumo, M.; Hansen, L.; Sisk, A.; Masliah, E. Abnormal distribution of the non-Abeta component of Alzheimer’s disease amyloid precursor/alpha-synuclein in Lewy body disease as revealed by proteinase K and formic acid pretreatment. Lab. Investig. J. Tech. Methods Pathol. 1998, 78, 1169–1177. [Google Scholar]
- Soto, C. Unfolding the role of protein misfolding in neurodegenerative diseases. Nat. Rev. Neurosci. 2003, 4, 49–60. [Google Scholar] [CrossRef]
- Miraglia, F.; Betti, L.; Palego, L.; Giannaccini, G. Parkinson’s Disease and Alpha-Synucleinopathies: From Arising Pathways to Therapeutic Challenge. Cent. Nerv. Syst. Agents Med. Chem. 2015, 15, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Spires-Jones, T.L.; Attems, J.; Thal, D.R. Interactions of pathological proteins in neurodegenerative diseases. Acta Neuropathol. 2017, 134, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Chesselet, M.F. In vivo alpha-synuclein overexpression in rodents: A useful model of Parkinson’s disease? Exp. Neurol. 2008, 209, 22–27. [Google Scholar] [CrossRef]
- Breydo, L.; Wu, J.W.; Uversky, V.N. α-Synuclein misfolding and Parkinson’s disease. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2012, 1822, 261–285. [Google Scholar] [CrossRef]
- Roberts, R.F.; Wade-Martins, R.; Alegre-Abarrategui, J. Direct visualization of alpha-synuclein oligomers reveals previously undetected pathology in Parkinson’s disease brain. Brain 2015, 138, 1642–1657. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Chandran, K.; Kalivendi, S.V.; Joseph, J.; Antholine, W.E.; Hillard, C.J.; Kanthasamy, A.; Kanthasamy, A.; Kalyanaraman, B. Neuroprotection by a mitochondria-targeted drug in a Parkinson’s disease model. Free. Radic. Biol. Med. 2010, 49, 1674–1684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A.G. Neuroprotective effect of protein kinase Cδ inhibitor rottlerin in cell culture and animal models of Parkinson’s disease. J. Pharmacol. Exp. Ther. 2007, 322, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Langley, M.R.; Ghaisas, S.; Ay, M.; Luo, J.; Palanisamy, B.N.; Jin, H.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A.G. Manganese exposure exacerbates progressive motor deficits and neurodegeneration in the MitoPark mouse model of Parkinson’s disease: Relevance to gene and environment interactions in metal neurotoxicity. NeuroToxicology 2018, 64, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Saminathan, H.; Kanthasamy, A.; Anantharam, V.; Jin, H.; Sondarva, G.; Harischandra, D.S.; Qian, Z.; Rana, A.; Kanthasamy, A.G. The peptidyl-prolyl isomerase Pin1 up-regulation and proapoptotic function in dopaminergic neurons: Relevance to the pathogenesis of Parkinson disease. J. Biol. Chem. 2013, 288, 21955–21971. [Google Scholar] [CrossRef] [PubMed]
- Graff, C.L.; Pollack, G.M. Nasal Drug Administration: Potential for Targeted Central Nervous System Delivery. J. Pharm. Sci. 2005, 94, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Fleming, S.M.; Tetreault, N.A.; Mulligan, C.K.; Hutson, C.B.; Masliah, E.; Chesselet, M. Olfactory deficits in mice overexpressing human wildtype α-synuclein. Eur. J. Neurosci. 2008, 28, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lussier, S.; Rane, A.; Choi, S.; Andersen, J. Inducible dopaminergic glutathione depletion in an alpha-synuclein transgenic mouse model results in age-related olfactory dysfunction. Neuroscience 2011, 172, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Lucas, P.; Donohoe, S.; Thody, A. The role of estrogen and progesterone in the control of preputial gland sex attractant odors in the female rat. Physiol. Behav. 1982, 28, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.G.; Clancy, A.N.; Macrides, F.; Agosta, W.C.; Bronson, F.H. Chemical Properties of a Female Mouse Pheromone that Stimulates Gonadotropin Secretion in Males1. Biol. Reprod. 1988, 38, 193–199. [Google Scholar] [CrossRef]
- Kitazawa, M.; Anantharam, V.; Kanthasamy, A.G. Dieldrin-induced oxidative stress and neurochemical changes contribute to apoptopic cell death in dopaminergic cells. Free. Radic. Biol. Med. 2001, 31, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Kaul, S.; Kanthasamy, A.; Kitazawa, M.; Anantharam, V.; Kanthasamy, A.G. Caspase-3 dependent proteolytic activation of protein kinase Cδ mediates and regulates 1-methyl-4-phenylpyridinium (MPP+)-induced apoptotic cell death in dopaminergic cells: Relevance to oxidative stress in dopaminergic degeneration. Eur. J. Neurosci. 2003, 18, 1387–1401. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Anantharam, V.; Sun, F.; Kanthasamy, A.G. Environmental Neurotoxic Pesticide Increases Histone Acetylation to Promote Apoptosis in Dopaminergic Neuronal Cells: Relevance to Epigenetic Mechanisms of Neurodegeneration. Mol. Pharmacol. 2010, 77, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Shum, S.C.K.; Neddersen, R.; Houk, R.S. Elemental speciation by liquid chromatography–inductively coupled plasma mass spectrometry with direct injection nebulization. Analyst 1992, 117, 577–582. [Google Scholar] [CrossRef]
- Choi, C.J.; Anantharam, V.; Saetveit, N.J.; Houk, R.S.; Kanthasamy, A.G. Normal Cellular Prion Protein Protects against Manganese-Induced Oxidative Stress and Apoptotic Cell Death. Toxicol. Sci. 2007, 98, 495–509. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngwa, H.A.; Bargues-Carot, A.; Jin, H.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A.G. Manganese and Vanadium Co-Exposure Induces Severe Neurotoxicity in the Olfactory System: Relevance to Metal-Induced Parkinsonism. Int. J. Mol. Sci. 2024, 25, 5285. https://doi.org/10.3390/ijms25105285
Ngwa HA, Bargues-Carot A, Jin H, Anantharam V, Kanthasamy A, Kanthasamy AG. Manganese and Vanadium Co-Exposure Induces Severe Neurotoxicity in the Olfactory System: Relevance to Metal-Induced Parkinsonism. International Journal of Molecular Sciences. 2024; 25(10):5285. https://doi.org/10.3390/ijms25105285
Chicago/Turabian StyleNgwa, Hilary Afeseh, Alejandra Bargues-Carot, Huajun Jin, Vellareddy Anantharam, Arthi Kanthasamy, and Anumantha G. Kanthasamy. 2024. "Manganese and Vanadium Co-Exposure Induces Severe Neurotoxicity in the Olfactory System: Relevance to Metal-Induced Parkinsonism" International Journal of Molecular Sciences 25, no. 10: 5285. https://doi.org/10.3390/ijms25105285
APA StyleNgwa, H. A., Bargues-Carot, A., Jin, H., Anantharam, V., Kanthasamy, A., & Kanthasamy, A. G. (2024). Manganese and Vanadium Co-Exposure Induces Severe Neurotoxicity in the Olfactory System: Relevance to Metal-Induced Parkinsonism. International Journal of Molecular Sciences, 25(10), 5285. https://doi.org/10.3390/ijms25105285