

Loop-Mediated Isothermal Amplification of DNA (LAMP) as an Alternative Method for Determining Bacteria in Wound Infections
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Real-Time Polymerase Chain Reaction (Real-Time PCR, qPCR)
3. Genotyping of Strains from Wound Infections
- -
- Preparing the sample and isolating the genetic material;
- -
- Creation of libraries that will be compatible with the selected sequencer;
- -
- Amplification and the sequencing process;
- -
- Computer processing of the obtained data and its analysis [69].
4. Loop-Mediated Isothermal Amplification of DNA (LAMP)
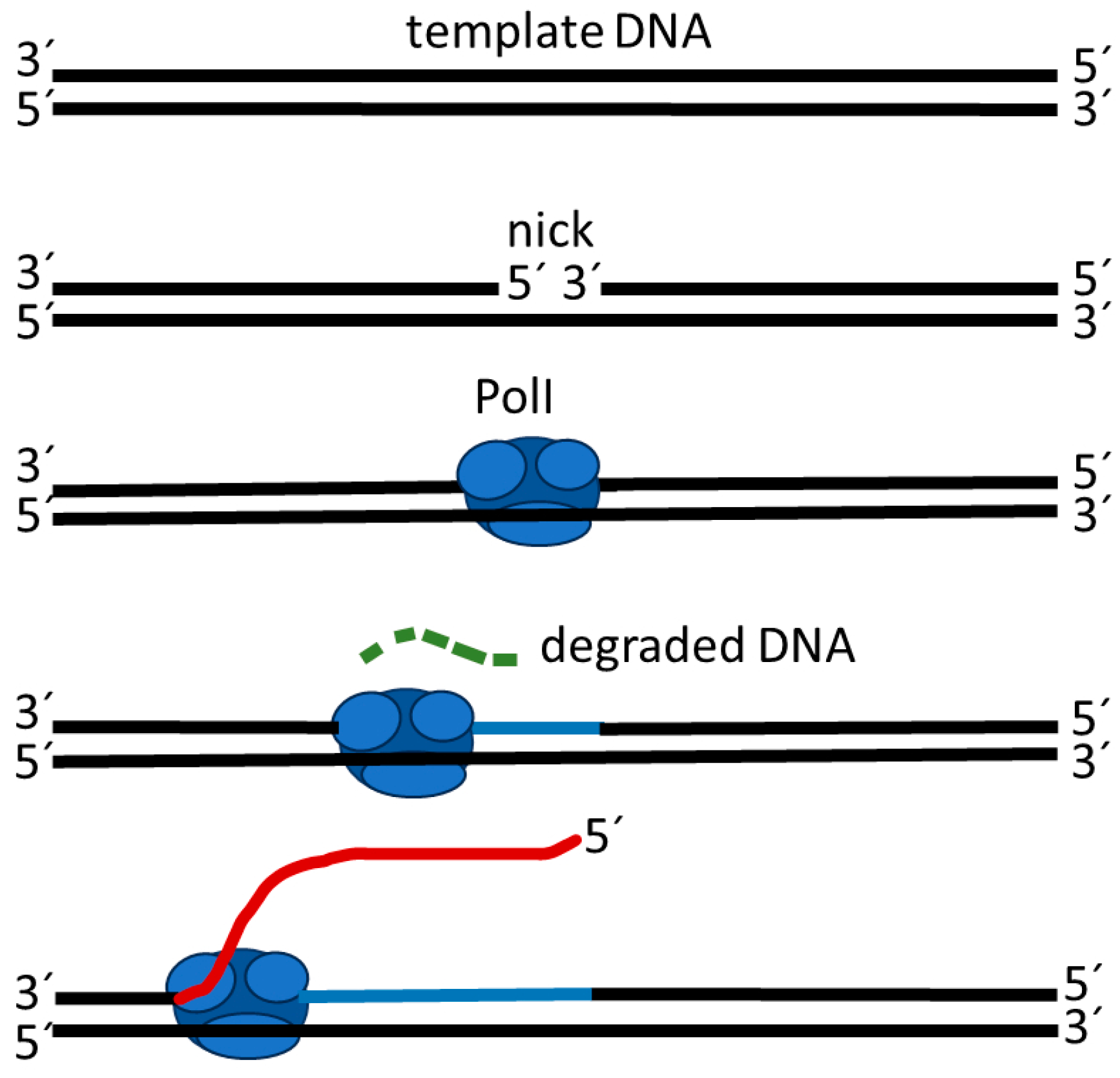
4.1. Strand Displacement Synthesis
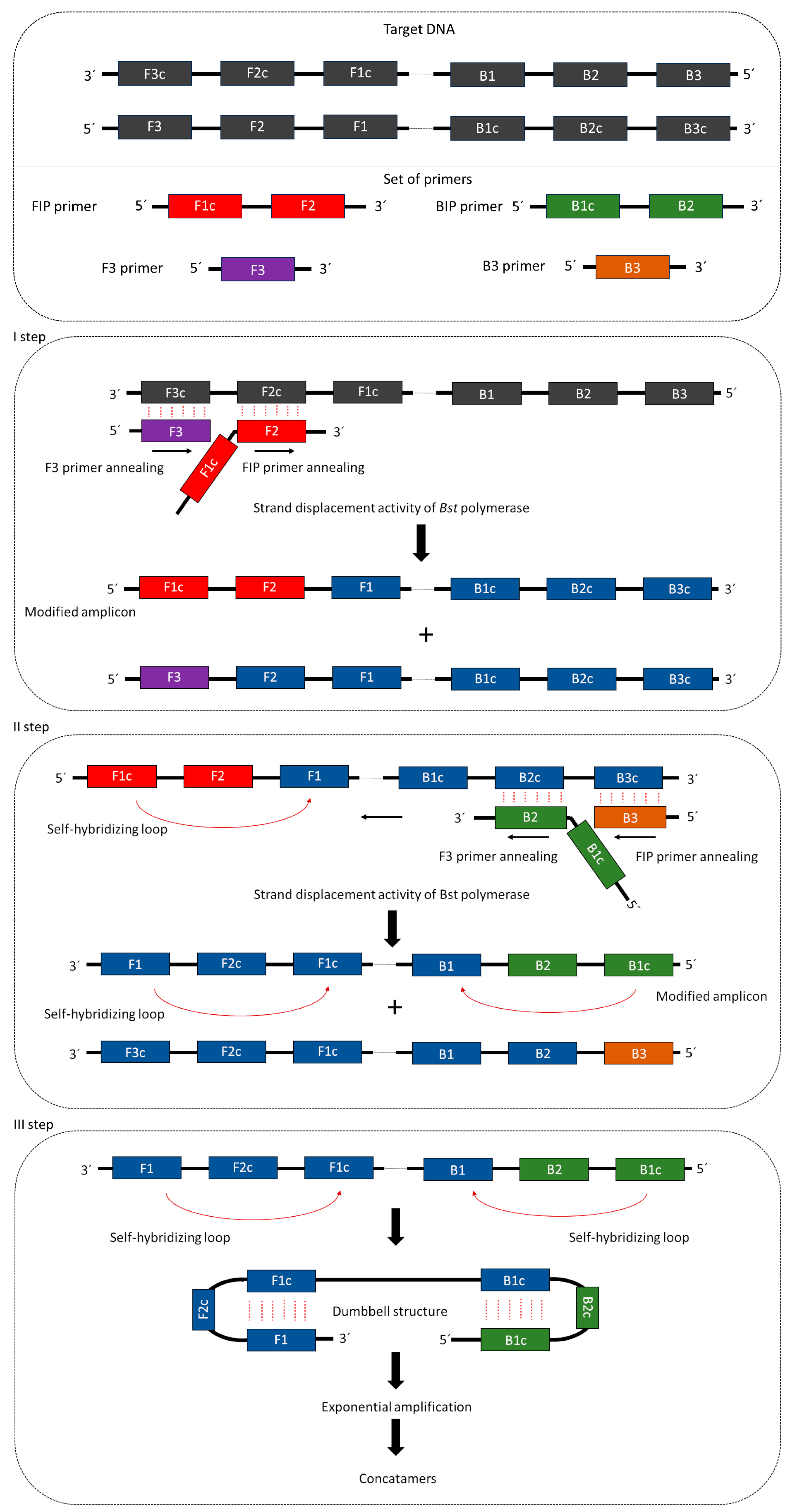
4.2. Mechanism of LAMP Reaction
4.3. Diagnostic Use of LAMP
Author Contributions
Funding
Conflicts of Interest
References
- Bekara, F.; Vitse, J.; Fluieraru, S.; Masson, R.; De Runz, A.; Georgescu, V.; Bressy, G.; Labbé, J.L.; Chaput, B.; Herlin, C. New Techniques for Wound Management: A Systematic Review of Their Role in the Management of Chronic Wounds. Arch. Plast. Surg. 2018, 45, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Krzystek-Korpacka, M.; Kȩdzior, K.; Masłowski, L.; Mierzchała, M.; Bednarz-Misa, I.; Bronowicka-Szydełko, A.; Kubiak, J.; Gacka, M.; Płaczkowska, S.; Gamian, A. Impact of Chronic Wounds of Various Etiology on Systemic Profiles of Key Inflammatory Cytokines, Chemokines and Growth Factors, and Their Interplay. Adv. Clin. Exp. Med. 2019, 28, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Oates, A.; Bowling, F.L.; Boulton, A.J.M.; Bowler, P.G.; Metcalf, D.G.; McBain, A.J. The Visualization of Biofilms in Chronic Diabetic Foot Wounds Using Routine Diagnostic Microscopy Methods. J. Diabetes Res. 2014, 2014, 153586. [Google Scholar] [CrossRef] [PubMed]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.D.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in Chronic Wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Omar, A.; Wright, J.B.; Schultz, G.; Burrell, R.; Nadworny, P. Microbial Biofilms and Chronic Wounds. Microorganisms 2017, 5, 9. [Google Scholar] [CrossRef]
- Berlanga, M.; Guerrero, R. Living Together in Biofilms: The Microbial Cell Factory and Its Biotechnological Implications. Microb. Cell Factories 2016, 15, 165. [Google Scholar] [CrossRef]
- Davis, S.C.; Ricotti, C.; Cazzaniga, A.; Welsh, E.; Eaglstein, W.H.; Mertz, P.M. Microscopic and Physiologic Evidence for Biofilm-Associated Wound Colonization in Vivo. Wound Repair Regen. 2008, 16, 23–29. [Google Scholar] [CrossRef]
- Dalton, T.; Dowd, S.E.; Wolcott, R.D.; Sun, Y.; Watters, C.; Griswold, J.A.; Rumbaugh, K.P. An in Vivo Polymicrobial Biofilm Wound Infection Model to Study Interspecies Interactions. PLoS ONE 2011, 6, e27317. [Google Scholar] [CrossRef]
- Schierle, C.F.; De La Garza, M.; Mustoe, T.A.; Galiano, R.D. Staphylococcal Biofilms Impair Wound Healing by Delaying Reepithelialization in a Murine Cutaneous Wound Model. Wound Repair Regen. 2009, 17, 354–359. [Google Scholar] [CrossRef]
- Phillips, P.L.; Schultz, G.S. Molecular Mechanisms of Biofilm Infection: Biofilm Virulence Factors. Adv. Wound Care 2012, 1, 109–114. [Google Scholar] [CrossRef]
- Stewart, P.S. Mechanisms of Antibiotic Resistance in Bacterial Biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Burmølle, M.; Webb, J.S.; Rao, D.; Hansen, L.H.; Sørensen, S.J.; Kjelleberg, S. Enhanced Biofilm Formation and Increased Resistance to Antimicrobial Agents and Bacterial Invasion Are Caused by Synergistic Interactions in Multispecies Biofilms. Appl. Environ. Microbiol. 2006, 72, 3916–3923. [Google Scholar] [CrossRef]
- DeLeon, S.; Clinton, A.; Fowler, H.; Everett, J.; Horswill, A.R.; Rumbaugh, K.P. Synergistic Interactions of Pseudomonas Aeruginosa and Staphylococcus Aureus in an In Vitro Wound Model. Infect. Immun. 2014, 82, 4718–4728. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.M.; Al-Badi, E.; Withycombe, C.; Jones, P.M.; Purdy, K.J.; Maddocks, S.E. Interaction between Staphylococcus Aureus and Pseudomonas Aeruginosa Is Beneficial for Colonisation and Pathogenicity in a Mixed Biofilm. Pathog. Dis. 2018, 76, fty003. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Hill, K.E.; Williams, D.W.; Hooper, S.J.; Thomas, D.W.; Costerton, J.W. A Review of the Scientific Evidence for Biofilms in Wounds. Wound Repair Regen. 2012, 20, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, M.D. Bacteria and Antibiotics in Wound Healing. Surg. Clin. N. Am. 2020, 100, 757–776. [Google Scholar] [CrossRef] [PubMed]
- Leaper, D.J.; Schultz, G.; Carville, K.; Fletcher, J.; Swanson, T.; Drake Leaper, R.D. Extending the TIME Concept: What Have We Learned in the Past 10 Years?*. Int. Wound J. 2012, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.E.; Frantz, R.A.; Park, H.; Scherubel, M. The Inter-Rater Reliability of the Clinical Signs and Symptoms Checklist in Diabetic Foot Ulcers. Ostomy Wound Manag. 2007, 53, 46–51. [Google Scholar]
- Gardner, S.E.; Hillis, S.L.; Frantz, R.A. Clinical Signs of Infection in Diabetic Foot Ulcers with High Microbial Load. Biol. Res. Nurs. 2009, 11, 119–128. [Google Scholar] [CrossRef]
- Cutting, K.F.; White, R.J. Criteria for Identifying Wound Infection--Revisited. Ostomy Wound Manag. 2005, 51, 28–34. [Google Scholar]
- Falcone, M.; De Angelis, B.; Pea, F.; Scalise, A.; Stefani, S.; Tasinato, R.; Zanetti, O.; Dalla Paola, L. Challenges in the Management of Chronic Wound Infections. J. Glob. Antimicrob. Resist. 2021, 26, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.E.; Frantz, R.A.; Doebbeling, B.N. The Validity of the Clinical Signs and Symptoms Used to Identify Localized Chronic Wound Infection. Wound Repair Regen. 2001, 9, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Lagier, J.C.; Edouard, S.; Pagnier, I.; Mediannikov, O.; Drancourt, M.; Raoult, D. Current and Past Strategies for Bacterial Culture in Clinical Microbiology. Clin. Microbiol. Rev. 2015, 28, 208–236. [Google Scholar] [CrossRef] [PubMed]
- Kalan, L.; Loesche, M.; Hodkinson, B.P.; Heilmann, K.; Ruthel, G.; Gardner, S.E.; Grice, E.A. Redefining the Chronic-Wound Microbiome: Fungal Communities Are Prevalent, Dynamic, and Associated with Delayed Healing. mBio 2016, 7, e01058-16. [Google Scholar] [CrossRef] [PubMed]
- Bowler, P.G.; Duerden, B.I.; Armstrong, D.G. Wound microbiology and associated approaches to wound management. Clin. Microbiol. Rev. 2001, 14, 244–269. [Google Scholar] [CrossRef]
- Brook, I.; Frazier, E.H. Aerobic and Anaerobic Microbiology of Chronic Venous Ulcers. Int. J. Dermatol. 1998, 37, 426–428. [Google Scholar] [CrossRef]
- Choi, Y.; Banerjee, A.; McNish, S.; Couch, K.S.; Torralba, M.G.; Lucas, S.; Tovchigrechko, A.; Madupu, R.; Yooseph, S.; Nelson, K.E.; et al. Co-Occurrence of Anaerobes in Human Chronic Wounds. Microb. Ecol. 2019, 77, 808–820. [Google Scholar] [CrossRef]
- Rhoads, D.D.; Wolcott, R.D.; Sun, Y.; Dowd, S.E. Comparison of Culture and Molecular Identification of Bacteria in Chronic Wounds. Int. J. Mol. Sci. 2012, 13, 2535–2550. [Google Scholar] [CrossRef]
- Rudkjøbing, V.B.; Thomsen, T.R.; Xu, Y.; Melton-Kreft, R.; Ahmed, A.; Eickhardt, S.; Bjarnsholt, T.; Poulsen, S.S.; Nielsen, P.H.; Earl, J.P.; et al. Comparing Culture and Molecular Methods for the Identification of Microorganisms Involved in Necrotizing Soft Tissue Infections. BMC Infect. Dis. 2016, 16, 652. [Google Scholar] [CrossRef]
- Rhoads, D.D.; Cox, S.B.; Rees, E.J.; Sun, Y.; Wolcott, R.D. Clinical Identification of Bacteria in Human Chronic Wound Infections: Culturing vs. 16S Ribosomal DNA Sequencing. BMC Infect. Dis. 2012, 12, 321. [Google Scholar] [CrossRef]
- Dowd, S.E.; Sun, Y.; Secor, P.R.; Rhoads, D.D.; Wolcott, B.M.; James, G.A.; Wolcott, R.D. Survey of Bacterial Diversity in Chronic Wounds Using Pyrosequencing, DGGE, and Full Ribosome Shotgun Sequencing. BMC Microbiol. 2008, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, M.S.; Mostow, E.; Mukherjee, P.; Hu, F.Z.; Melton-Kreft, R.; Ehrlich, G.D.; Dowd, S.E.; Ghannoum, M.A. Characterization of Bacterial Communities in Venous Insufficiency Wounds by Use of Conventional Culture and Molecular Diagnostic Methods. J. Clin. Microbiol. 2011, 49, 3812–3819. [Google Scholar] [CrossRef] [PubMed]
- Brook, I.; Frazier, E.H. Aerobic and Anaerobic Bacteriology of Wounds and Cutaneous Abscesses. Arch. Surg. 1990, 125, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Dowd, S.E.; Wolcott, R.R.D.; Kennedy, J.; Jones, C.; Cox, S.B. Molecular Diagnostics and Personalised Medicine in Wound Care: Assessment of Outcomes. J. Wound Care 2011, 20, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Wolcott, R.D.; Cox, S.B.; Dowd, S.E. Healing and Healing Rates of Chronic Wounds in the Age of Molecular Pathogen Diagnostics. J. Wound Care 2010, 19, 276–284. [Google Scholar] [CrossRef]
- Nestorov, J.; Matić, G.; Elaković, I.; Tanić, N. Gene expression studies: How to obtain accurate and reliable data by quantitative real-time RT PCR. J. Med. Biochem. 2013, 32, 325–338. [Google Scholar] [CrossRef]
- Joshi, M.; Deshpande, J.D. Polymerase chain reaction: Methods, principles and application. Int. J. BioMed. Res. 2011, 2, 81–97. [Google Scholar] [CrossRef]
- Kralik, P.; Ricchi, M. A Basic Guide to Real Time PCR in Microbial Diagnostics: Definitions, Parameters, and Everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef]
- Espy, M.J.; Uhl, J.R.; Sloan, L.M.; Buckwalter, S.P.; Jones, M.F.; Vetter, E.A.; Yao, J.D.C.; Wengenack, N.L.; Rosenblatt, J.E.; Iii, F.R.C.; et al. Real-Time PCR in Clinical Microbiology: Applications for Routine Laboratory Testing. Clin. Microbiol. Rev. 2006, 19, 165–256. [Google Scholar] [CrossRef]
- Trung, T.T.; Hetzer, A.; Topfstedt, E.; Wuthiekanun, V.; Limmathurotsakul, D.; Peacock, S.J.; Steinmetz, I. Highly Sensitive Direct Detection and Quantification of Burkholderia Pseudomallei Bacteria in Environmental Soil Samples by Using Real-Time PCR. Appl. Environ. Microbiol. 2011, 77, 6486–6494. [Google Scholar] [CrossRef]
- Ridley, A.M.; Allen, V.M.; Sharma, M.; Harris, J.A.; Newell, D.G. Real-Time PCR Approach for Detection of Environmental Sources of Campylobacter Strains Colonizing Broiler Flocks. Appl. Environ. Microbiol. 2008, 74, 2492–2504. [Google Scholar] [CrossRef]
- Chiba, N.; Murayama, S.Y.; Morozumi, M.; Nakayama, E.; Okada, T.; Iwata, S. Rapid Detection of Eight Causative Pathogens for the Diagnosis of Bacterial Meningitis by Real-Time PCR. J. Infect. Chemother. 2009, 15, 92–98. [Google Scholar] [CrossRef]
- Anbazhagan, D.; Mui, W.S.; Mansor, M.; Yan, G.O.S.; Yusof, M.Y.; Sekaran, S.D. Development of conventional and real-time multiplex PCR assays for the detection of nosocomial pathogens. Braz. J. Microbiol. 2011, 42, 448–458. [Google Scholar] [CrossRef]
- Curran, T.; Coyle, P.V.; Mcmanus, T.E.; Kidney, J.; Coulter, W.A. Evaluation of Real-Time PCR for the Detection and Quantification of Bacteria in Chronic Obstructive Pulmonary Disease. FEMS Immunol. Med. Microbiol. 2007, 50, 112–118. [Google Scholar] [CrossRef]
- Zemanick, E.T.; Wagner, B.D.; Sagel, S.D.; Stevens, M.J.; Accurso, F.J.; Kirk, J. Reliability of Quantitative Real-Time PCR for Bacterial Detection in Cystic Fibrosis Airway Specimens. PLoS ONE 2010, 5, e15101. [Google Scholar] [CrossRef]
- Gosiewski, T.; Jurkiewicz-badacz, D.; Sroka, A.; Brzychczy-Włoch, M.; Bulanda, M. A Novel, Nested, Multiplex, Real-Time PCR for Detection of Bacteria and Fungi in Blood. BMC Microbiol. 2014, 14, 144. [Google Scholar] [CrossRef]
- Gaibani, P.; Mariconti, M.; Bua, G.; Bonora, S.; Sassera, D.; Landini, M.P.; Mulatto, P.; Novati, S.; Bandi, C.; Sambri, V. Development of a Broad-Range 23S RDNA Real-Time PCR Assay for the Detection and Quantification of Pathogenic Bacteria in Human Whole Blood and Plasma Specimens. BioMed Res. Int. 2013, 2013, 13–17. [Google Scholar] [CrossRef]
- Oates, A.; Bowling, F.L.; Boulton, A.J.M.; Mcbain, A.J. Molecular and Culture-Based Assessment of the Microbial Diversity of Diabetic Chronic Foot Wounds and Contralateral Skin Sites. J. Clin. Microbiol. 2012, 50, 2263–2271. [Google Scholar] [CrossRef]
- Melendez, J.H.; Frankel, Y.M.; An, A.T.; Williams, L.; Price, L.B.; Wang, N.; Lazarus, G.S.; Zenilman, J.M. Real-Time PCR Assays Compared to Culture-Based Approaches for Identification of Aerobic Bacteria in Chronic Wounds. Clin. Microbiol. Infect. 2010, 16, 1762–1769. [Google Scholar] [CrossRef]
- Gentili, V.; Gianesini, S.; Balboni, P.G.; Menegatti, E.; Rotola, A.; Zuolo, M.; Caselli, E.; Zamboni, P.; Di Luca, D. Panbacterial Real-Time PCR to Evaluate Bacterial Burden in Chronic Wounds Treated with CutimedTM SorbactTM. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1523–1529. [Google Scholar] [CrossRef]
- Yan, W.; Xing, Y.; Yuan, L.C.; De Yang, R.; Tan, F.Y.; Zhang, Y.; Li, H. Application of RLEP Real-Time PCR for Detection of M. Leprae DNA in Paraffin-Embedded Skin Biopsy Specimens for Diagnosis of Paucibacillary Leprosy. Am. J. Trop. Med. Hyg. 2014, 90, 524–529. [Google Scholar] [CrossRef]
- Menezes, L.C.; Rocchetti, T.T.; Bauab, K.D.C.; Cappellano, P.; Quiles, M.G.; Carlesse, F.; Salvador, J.; De Oliveira, R.; Carlos, A.; Pignatari, C. Diagnosis by Real-Time Polymerase Chain Reaction of Pathogens and Antimicrobial Resistance Genes in Bone Marrow Transplant Patients with Bloodstream Infections. BMC Infect. Dis. 2013, 13, 166. [Google Scholar] [CrossRef]
- Ushida, K.; Segawa, T.; Kohshima, S.; Takeuchi, N.; Fukui, K. Application of Real-Time PCR Array to the Multiple Detection of Antibiotic Resistant Genes in Glacier Ice Samples Short Communication Application of Real-Time PCR Array to the Multiple Detection of Antibiotic Resistant Genes in Glacier Ice Samples. J. Gen. Appl. Microbiol. 2010, 56, 43–52. [Google Scholar] [CrossRef]
- Tsuru, A.; Setoguchi, T.; Kawabata, N.; Hirotsu, M.; Yamamoto, T.; Nagano, S. Enrichment of Bacteria Samples by Centrifugation Improves the Diagnosis of Orthopaedics—Related Infections via Real—Time PCR Amplification of the Bacterial Methicillin—Resistance Gene. BMC Res. Notes 2015, 8, 288. [Google Scholar] [CrossRef]
- Ogura, K.; Shimada, K.; Miyoshi-Akiyama, T. A Multilocus Sequence Typing Scheme of Pseudomonas Putida for Clinical and Environmental Isolates. Sci. Rep. 2019, 9, 13980. [Google Scholar] [CrossRef]
- Neoh, H.M.; Tan, X.E.; Sapri, H.F.; Tan, T.L. Pulsed-Field Gel Electrophoresis (PFGE): A Review of the “Gold Standard” for Bacteria Typing and Current Alternatives. Infect. Genet. Evol. 2019, 74, 103935. [Google Scholar] [CrossRef]
- Liou, C.H.; Wu, H.C.; Liao, Y.C.; Lauderdale, T.L.Y.; Huang, I.W.; Chen, F.J. Nanomlst: Accurate Multilocus Sequence Typing Using Oxford Nanopore Technologies Minion with a Dual-Barcode Approach to Multiplex Large Numbers of Samples. Microb. Genom. 2020, 6, e000336. [Google Scholar] [CrossRef]
- Joshi, C.J.; Ke, W.; Drangowska-Way, A.; O’Rourke, E.J.; Lewis, N.E. What Are Housekeeping Genes? PLoS Comput. Biol. 2022, 18, e1010295. [Google Scholar] [CrossRef]
- Mitchelmore, P.J.; Withers, N.J.; Sheldon, C.D.; Scotton, C.J.; Brown, A.R. Culture-Independent Multilocus Sequence Typing of Pseudomonas Aeruginosa for Cross-Infection Screening. Diagn. Microbiol. Infect. Dis. 2021, 100, 115315. [Google Scholar] [CrossRef]
- Drevinek, P.; Vosahlikova, S.; Dedeckova, K.; Cinek, O.; Mahenthiralingam, E. Direct Culture-Independent Strain Typing of Burkholderia Cepacia Complex in Sputum Samples from Patients with Cystic Fibrosis. J. Clin. Microbiol. 2010, 48, 1888–1891. [Google Scholar] [CrossRef]
- Abdel-Rhman, S.H.; Rizk, D.E. Comparative Assessment of Different PCR-Based Typing Methods of Pseudomonas Aeruginosa Isolates. Infect. Drug Resist. 2021, 14, 1019–1035. [Google Scholar] [CrossRef]
- Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski’, J.A.; Tingey1, S.V. DNA Polymorphisms Amplified by Arbitrary Primers Are Useful as Genetic Markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef]
- Hematzadeh, A.; Haghkhah, M. Biotyping of Isolates of Pseudomonas Aeruginosa Isolated from Human Infections by RAPD and ERIC-PCR. Heliyon 2021, 7, e07967. [Google Scholar] [CrossRef]
- Nanvazadeh, F.; Khosravi, A.D.; Zolfaghari, M.R.; Parhizgari, N. Genotyping of Pseudomonas Aeruginosa Strains Isolated from Burn Patients by RAPD-PCR. Burns 2013, 39, 1409–1413. [Google Scholar] [CrossRef]
- Hong, N.; Chen, M.; Xu, J. Molecular Markers Reveal Epidemiological Patterns and Evolutionary Histories of the Human Pathogenic Cryptococcus. Front. Cell. Infect. Microbiol. 2021, 11, 683670. [Google Scholar] [CrossRef]
- Bogiel, T.; Mikucka, A.; Kanarek, P. Agarose Gel Electrophoresis-Based RAPD-PCR—An Optimization of the Conditions to Rapidly Detect Similarity of the Alert Pathogens for the Purpose of Epidemiological Studies. Gels 2022, 8, 760. [Google Scholar] [CrossRef]
- Smith, C.J.; Osborn, A.M. Advantages and Limitations of Quantitative PCR (Q-PCR)-Based Approaches in Microbial Ecology. FEMS Microbiol. Ecol. 2009, 67, 6–20. [Google Scholar] [CrossRef]
- Didelot, X.; Bowden, R.; Wilson, D.J.; Peto, T.E.A.; Crook, D.W. Transforming Clinical Microbiology with Bacterial Genome Sequencing. Nat. Rev. Genet. 2012, 13, 601–612. [Google Scholar] [CrossRef]
- Vincent, A.T.; Derome, N.; Boyle, B.; Culley, A.I.; Charette, S.J. Next-Generation Sequencing (NGS) in the Microbiological World: How to Make the Most of Your Money. J. Microbiol. Methods 2017, 138, 60–71. [Google Scholar] [CrossRef]
- Hu, T.; Chitnis, N.; Monos, D.; Dinh, A. Next-Generation Sequencing Technologies: An Overview. Hum. Immunol. 2021, 82, 801–811. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Teng, J.L.L.; Tse, H.; Yuen, K.Y. Then and Now: Use of 16S RDNA Gene Sequencing for Bacterial Identification and Discovery of Novel Bacteria in Clinical Microbiology Laboratories. Clin. Microbiol. Infect. 2008, 14, 908–934. [Google Scholar] [CrossRef]
- Hodkinson, B.P.; Grice, E.A. Next-Generation Sequencing: A Review of Technologies and Tools for Wound Microbiome Research. Adv. Wound Care 2015, 4, 50–58. [Google Scholar] [CrossRef]
- Misic, A.M.; Gardner, S.E.; Grice, E.A. The Wound Microbiome: Modern Approaches to Examining the Role of Microorganisms in Impaired Chronic Wound Healing. Adv. Wound Care 2014, 3, 502–510. [Google Scholar] [CrossRef]
- Petersen, L.M.; Martin, I.W.; Moschetti, W.E.; Kershaw, C.M.; Tsongalis, G.J. Third-Generation Sequencing in the Clinical Laboratory: Exploring the Advantages and Challenges of Nanopore Sequencing. J. Clin. Microbiol. 2019, 58, e01315-19. [Google Scholar] [CrossRef]
- Zhang, J.; Tu, J.; Chen, Y.; Jin, X. Clinical Characteristics and Homology Analysis of Staphylococcus Aureus from Would Infection at a Tertiary Hospital in Southern Zhejiang, China. BMC Microbiol. 2023, 23, 217. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, F.; Zhang, Y.; Wang, X.; Gao, W.; Xu, B.; Li, Y.; Song, N. Virulence, Antimicrobial Resistance, and Molecular Characteristics of Carbapenem-Resistant Klebsiella Pneumoniae in a Hospital in Shijiazhuang City from China. Int. Microbiol. 2023, 26, 1073–1085. [Google Scholar] [CrossRef]
- Akinduti, A.P.; Osiyemi, J.A.; Banjo, T.T.; Ejilude, O.; El-Ashker, M.; Adeyemi, A.G.; Obafemi, Y.D.; Isibor, P.O. Clonal Diversity and Spatial Dissemination of Multi-Antibiotics Resistant Staphylococcus Aureus Pathotypes in Southwest Nigeria. PLoS ONE 2021, 16, e0247013. [Google Scholar] [CrossRef]
- Kwiatkowski, P.; Masiuk, H.; Pruss, A.; Łopusiewicz, Ł.; Sienkiewicz, M.; Wojciechowska-Koszko, I.; Roszkowska, P.; Bania, J.; Guenther, S.; Dołęgowska, B. Clonal Diversity, Antimicrobial Susceptibility and Presence of Genes Encoding Virulence Factors in Staphylococcus Aureus Strains Isolated from Cut Wound Infections. Curr. Microbiol. 2022, 79, 144. [Google Scholar] [CrossRef]
- Sharahi, J.Y.; Hashemi, A.; Ardebili, A.; Davoudabadi, S. Molecular Characteristics of Antibiotic-Resistant Escherichia Coli and Klebsiella Pneumoniae Strains Isolated from Hospitalized Patients in Tehran, Iran. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 32. [Google Scholar] [CrossRef]
- Schaumburg, F.; Vas Nunes, J.; Mönnink, G.; Falama, A.M.; Bangura, J.; Mathéron, H.; Conteh, A.; Sesay, M.; Sesay, A.; Grobusch, M.P. Chronic Wounds in Sierra Leone: Pathogen Spectrum and Antimicrobial Susceptibility. Infection 2022, 50, 907–914. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-Mediated Isothermal Amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Y. Development of Primer Sets for Loop-Mediated Isothermal Amplification That Enables Rapid and Specific Detection of Streptococcus Dysgalactiae, Streptococcus Uberis and Streptococcus Agalactiae. Int. J. Environ. Res. Public Health 2015, 12, 5735–5742. [Google Scholar] [CrossRef] [PubMed]
- Chantratita, N.; Meumann, E.; Thanwisai, A.; Limmathurotsakul, D.; Wuthiekanun, V.; Wannapasni, S.; Tumapa, S.; Day, N.P.J.; Peacock, S.J. Loop-Mediated Isothermal Amplification Method Targeting the TTS1 Gene Cluster for Detection of Burkholderia Pseudomallei and Diagnosis of Melioidosis. J. Clin. Microbiol. 2008, 46, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Etchebarne, B.E.; Li, Z.; Stedtfeld, R.D.; Nicholas, M.C.; Williams, M.R.; Johnson, T.A.; Stedtfeld, T.M.; Kostic, T.; Khalife, W.T.; Tiedje, J.M.; et al. Evaluation of Nucleic Acid Isothermal Amplification Methods for Human Clinical Microbial Infection Detection. Front. Microbiol. 2017, 8, 2211. [Google Scholar] [CrossRef] [PubMed]
- Klenow, H.; Henningsen, I. Selective Elimination of the Exonuclease Activity of the Deoxyribonucleic Acid Polymerase from Escherichia coli B by Limited Proteolysis. Proc. Natl. Acad. Sci. USA 1970, 65, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Srivastava, A.; Patel, S.S.; Modak, M.J. Participation of the Fingers Subdomain of Escherichia Coli DNA Polymerase I in the Strand Displacement Synthesis of DNA. J. Biol. Chem. 2007, 282, 10594–10604. [Google Scholar] [CrossRef]
- Su, H.; Xu, J.; Wang, Q.; Wang, F.; Zhou, X. High-Efficiency and Integrable DNA Arithmetic and Logic System Based on Strand Displacement Synthesis. Nat. Commun. 2019, 10, 5390. [Google Scholar] [CrossRef]
- Garafutdinov, R.R.; Gilvanov, A.R.; Sakhabutdinova, A.R. The Influence of Reaction Conditions on DNA Multimerization During Isothermal Amplification with Bst Exo− DNA Polymerase. Appl. Biochem. Biotechnol. 2020, 190, 758–771. [Google Scholar] [CrossRef]
- Qian, J.; Ferguson, T.M.; Shinde, D.N.; Ramírez-Borrero, A.J.; Hintze, A.; Adami, C.; Niemz, A. Sequence Dependence of Isothermal DNA Amplification via EXPAR. Nucleic Acids Res. 2012, 40, e87. [Google Scholar] [CrossRef]
- Giannattasio, M.; Branzei, D. DNA Replication through Strand Displacement during Lagging Strand DNA Synthesis in Saccharomyces Cerevisiae. Genes 2019, 10, 167. [Google Scholar] [CrossRef]
- Wong, Y.P.; Othman, S.; Lau, Y.L.; Radu, S.; Chee, H.Y. Loop-Mediated Isothermal Amplification (LAMP): A Versatile Technique for Detection of Micro-Organisms. J. Appl. Microbiol. 2018, 124, 626–643. [Google Scholar] [CrossRef] [PubMed]
- Soroka, M.; Wasowicz, B.; Rymaszewska, A. Loop-Mediated Isothermal Amplification (Lamp): The Better Sibling of PCR? Cells 2021, 10, 1931. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Azinheiro, S.; Roumani, F.; Prado, M.; Garrido-Maestu, A. Evaluation of the Effect of Outer Primer Structure, and Inner Primer Linker Sequences, in the Performance of Loop-Mediated Isothermal Amplification. Talanta 2023, 260, 124642. [Google Scholar] [CrossRef]
- Keikha, M. LAMP Method as One of the Best Candidates for Replacing with PCR Method. Malays. J. Med. Sci. 2018, 25, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.L.M. DNA Amplification in the Field: Move over PCR, Here Comes LAMP. Mol. Ecol. Resour. 2017, 17, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Mori, Y.; Tomita, N.; Kanda, H. Loop-Mediated Isothermal Amplification (LAMP): Principle, Features, and Future Prospects. J. Microbiol. 2015, 53, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Salamin, O.; Kuuranne, T.; Saugy, M.; Leuenberger, N. Loop-Mediated Isothermal Amplification (LAMP) as an Alternative to PCR: A Rapid on-Site Detection of Gene Doping. Drug Test. Anal. 2017, 9, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, P.; Zhou, S.; Zhang, L. Loop-Mediated Isothermal Amplification (LAMP): A Novel Rapid Detection Platform for Pathogens. Microb. Pathog. 2017, 107, 54–61. [Google Scholar] [CrossRef]
- Piotrowski, Y.; Gurung, M.K.; Larsen, A.N. Characterization and Engineering of a DNA Polymerase Reveals a Single Amino-Acid Substitution in the Fingers Subdomain to Increase Strand-Displacement Activity of A-Family Prokaryotic DNA Polymerases. BMC Mol. Cell Biol. 2019, 20, 31. [Google Scholar] [CrossRef]
- Chen, H.-W.; Ching, W.-M. Evaluation of the Stability of Lyophilized Loop-Mediated Isothermal Amplification Reagents for the Detection of Coxiella Burnetii. Heliyon 2017, 3, e00415. [Google Scholar] [CrossRef]
- Osawa, R.; Yoshida, A.; Masakiyo, Y.; Nagashima, S.; Ansai, T.; Watari, H.; Notomi, T.; Takehara, T. Rapid Detection of Actinobacillus Actinomycetemcomitans Using a Loop-Mediated Isothermal Amplification Method. Oral Microbiol. Immunol. 2007, 22, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Dugan, L.; Bearinger, J.; Hinckley, A.; Strout, C.; Souza, B. Detection of Bacillus Anthracis from Spores and Cells by Loop-Mediated Isothermal Amplification without Sample Preparation. J. Microbiol. Methods 2012, 90, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Park, B.H.; Jung, J.H.; Choi, G.; Lee, D.C.; Kim, D.H.; Seo, T.S. Centrifugal Loop-Mediated Isothermal Amplification Microdevice for Rapid, Multiplex and Colorimetric Foodborne Pathogen Detection. Biosens. Bioelectron. 2016, 75, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Achappa, B.; Madi, D.; Vidyalakshmi, K. Cutaneous Melioidosis. J. Clin. Diagn. Res. 2016, 10, WD01–WD02. [Google Scholar] [CrossRef]
- Lim, K.T.; Teh, C.S.J.; Thong, K.L. Loop-Mediated Isothermal Amplification Assay for the Rapid Detection of Staphylococcus Aureus. BioMed. Res. Int. 2013, 2013, 895816. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.J.; Zhou, S.; Zhang, X.Y.; Pu, J.H.; Ge, Q.L.; Tang, X.J.; Gao, Y.S. Rapid and Sensitive Detection of Listeria Monocytogenes by Loop-Mediated Isothermal Amplification. Curr. Microbiol. 2011, 63, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Yanagisawa, H.; Wachino, J. Rapid and Reliable Loop-Mediated Isothermal Amplification Method for Detecting Streptococcus Agalactiae. Jpn. J. Infect. Dis. 2013, 66, 546–548. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, R.; Kardoost Parizi, Z.; Ghorbanmehr, N.; Mirshafiee, H. Rapid and Simple Detection of Escherichia Coli by Loop-Mediated Isothermal Amplification Assay in Urine Specimens. Avicenna J. Med. Biotechnol. 2018, 10, 269. [Google Scholar]
- Li, C.; Fu, G.; Shi, Y.; Zhang, A.M.; Xia, X.; Fang, Y.; Mao, X.; Jiang, J.; Song, Y.; Yang, G. Rapid, Specific, and Sensitive Detection of the Urer_1 Gene in Klebsiella pneumoniae by Loop-Mediated Isothermal Amplification Method. Braz. J. Med. Biol. Res. 2019, 52, 1–9. [Google Scholar] [CrossRef]
- Qin, Y.; Duan, X.; Peng, Y.; Rui, Y. Rapid Detection of a Novel B1-β-Lactamase Gene, BlaAFM-1 Using a Loop-Mediated Isothermal Amplification (LAMP) Assay. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 80. [Google Scholar] [CrossRef]
- Li, C.; Shi, Y.; Yang, G.; Xia, X.; Mao, X.; Fang, Y.; Zhang, A.; Song, Y. Establishment of Loop-mediated Isothermal Amplification for Rapid Detection of Pseudomonas Aeruginosa. Exp. Ther. Med. 2018, 17, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gaind, R. Development of Loop-Mediated Isothermal Amplification Assay for Detection of Clinically Significant Members of Acinetobacter Calcoaceticus–Baumannii Complex and Associated Carbapenem Resistance. Front. Mol. Biosci. 2021, 8, 659256. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Feng, J.; Wang, Y. Direct Loop-Mediated Isothermal Amplification Assay for on-Site Detection of Staphylococcus Aureus. FEMS Microbiol. Lett. 2018, 365, fny092. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Liang, Y.; Chen, L.; Wang, W.; Wang, J.; Li, B.; Li, L.; Chen, D.; Xu, Z. Formation and Development of Staphylococcus Biofilm: With Focus on Food Safety. J. Food Saf. 2017, 37, e12358. [Google Scholar] [CrossRef]
- Rivoarilala, L.O.; Victor, J.; Crucitti, T.; Collard, J.M. LAMP Assays for the Simple and Rapid Detection of Clinically Important Urinary Pathogens Including the Detection of Resistance to 3rd Generation Cephalosporins. BMC Infect. Dis. 2021, 21, 1037. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.; Beriwal, S.; Chandra, I.; Paul, V.K.; Kapil, A.; Singh, T.; Wadowsky, R.M.; Singh, V.; Goyal, A.; Jahnukainen, T.; et al. Loop-Mediated Isothermal Amplification Assay for Rapid Detection of Common Strains of Escherichia Coli. J. Clin. Microbiol. 2008, 46, 2800–2804. [Google Scholar] [CrossRef]
- Dong, D.; Liu, W.; Li, H.; Wang, Y.; Li, X.; Zou, D.; Yang, Z.; Huang, S.; Zhou, D.; Huang, L.; et al. Survey and Rapid Detection of Klebsiella Pneumoniae in Clinical Samples Targeting the RcsA Gene in Beijing, China. Front. Microbiol. 2015, 6, 519. [Google Scholar] [CrossRef]
- Kato, H.; Yoshida, A.; Ansai, T.; Watari, H.; Notomi, T.; Takehara, T. Loop-Mediated Isothermal Amplification Method for the Rapid Detection of Enterococcus Faecalis in Infected Root Canals. Oral Microbiol. Immunol. 2007, 22, 131–135. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Species | Primers | Sequence (5′-3′) | References |
|---|---|---|---|
| Pseudomonas aeruginosa | F3 | CAAGCGCAAGATAGTCGCC | [111] |
| B3 | TCCGCTTGAACAGGCTGGTG | ||
| FIP | GAAGATATCCGGCTGGTTGCTTTTCAAGAGGGAATGCCGCAGT | ||
| BIP | AACGGATCATCGGCATCCTGGTTTTCATCGCCGTCCACAGGTAGA | ||
| Acinetobacter baumannii | F3 | CACAACAAGTTGTTCTTCATAGAT | [112] |
| B3 | CGAACTCCTGACCTCCTA | ||
| FIP | AGACTTGAACTTGTGACCCCACTGAGGGTCTGTAGCTCAG | ||
| BIP | ACCATGACTTTGACTGGTTRAAGTTCGCTCTACCAACTAAGCTAAG | ||
| Staphylococcus aureus | F3 | TCGCTTGCTATGATTGTGG | [113,114] |
| B3 | ACATACGCCAATGTTCTACC | ||
| FIP | GTACAGTTTCATGATTCGTCCCGCCATCATTATTGTAGGTGT | ||
| BIP | TGTTCAAAGAGTTGTGGATGGTGTACAGGCGTATTCGGTT | ||
| FLP | TTGAAAGGACCCGTATGATTCA | ||
| BLP | GATACGCCAGAAACGGTGA | ||
| Proteus mirabilis | F3 | AAAAAACGCGGWTCTGCA | [115] |
| B3 | AAGACAGATAGAGCCAACG | ||
| FIP | CTGTCGAGCTATGGGTATTAATCACTTTTATTGCGTAATTGGTTAAAARTC | ||
| BIP | GTTAGTTGCGCTATCTTGTGCTTCTTTTGAACGTGATACATCGGTAGA | ||
| LF | CCGCCATAGTACGTACTCGCCA | ||
| E. coli | F3 | GCC ATC TCC TGA TGACG | [116] |
| B3 | ATT TAC CGC AGC CAG ACG | ||
| FIP | CTG GGG CGA GGT CGT GGT ATT CCG ACA AAC ACC ACG AATT | ||
| BIP | CAT TTT GCA GCT GTA CGC TCG CAG CCC ATC ATG AAT GT | ||
| LF | CTT TGT AAC AAC CTG TCA TCG ACA | ||
| LB | ATC AAT CTC GAT ATC CAT GAA GGT G | ||
| K. pneumoniae | F3 | GGA TAT CTG ACC AGT CGG | [117] |
| B3 | GGG TTT TGC GTA ATG ATC TG | ||
| FIP | CGA CGT ACA GTG TTT CTG CAG TTT TAA AAA ACA GGA AAT CGT TGAGG | ||
| BIP | CGG CGG TGG TGT TTC TGA ATT TTG CGA ATA ATG CCA TTA CTT TC | ||
| LB | GAA GAC TGT TTC GTG CAT GATGA | ||
| E. faecalis | F3 | GCC GGA AAT CGA TGA AGA | [118] |
| B3 | TCC AGC AAC GTT GAT TGT | ||
| FIP | CAC TTT TTG TTG TTG GTT TTC GCT TTA TTA TCT GCT TGG GGT GC | ||
| BIP | ATC TGC AGA CAA AGT AGT AAT TGC TCC AAG CTT TTA AGC GTG TC | ||
| LF | AAA TGC TGC GCC AGC TCG | ||
| LB | TCC AAT GTG GAA CTT AAA CGT ACC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gieroń, M.; Żarnowiec, P.; Zegadło, K.; Gmiter, D.; Czerwonka, G.; Kaca, W.; Kręcisz, B. Loop-Mediated Isothermal Amplification of DNA (LAMP) as an Alternative Method for Determining Bacteria in Wound Infections. Int. J. Mol. Sci. 2024, 25, 411. https://doi.org/10.3390/ijms25010411
Gieroń M, Żarnowiec P, Zegadło K, Gmiter D, Czerwonka G, Kaca W, Kręcisz B. Loop-Mediated Isothermal Amplification of DNA (LAMP) as an Alternative Method for Determining Bacteria in Wound Infections. International Journal of Molecular Sciences. 2024; 25(1):411. https://doi.org/10.3390/ijms25010411
Chicago/Turabian StyleGieroń, Monika, Paulina Żarnowiec, Katarzyna Zegadło, Dawid Gmiter, Grzegorz Czerwonka, Wiesław Kaca, and Beata Kręcisz. 2024. "Loop-Mediated Isothermal Amplification of DNA (LAMP) as an Alternative Method for Determining Bacteria in Wound Infections" International Journal of Molecular Sciences 25, no. 1: 411. https://doi.org/10.3390/ijms25010411
APA StyleGieroń, M., Żarnowiec, P., Zegadło, K., Gmiter, D., Czerwonka, G., Kaca, W., & Kręcisz, B. (2024). Loop-Mediated Isothermal Amplification of DNA (LAMP) as an Alternative Method for Determining Bacteria in Wound Infections. International Journal of Molecular Sciences, 25(1), 411. https://doi.org/10.3390/ijms25010411