
LncRNA and Protein Expression Profiles Reveal Heart Adaptation to High-Altitude Hypoxia in Tibetan Sheep

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Quality Analysis of Transcriptomic and Proteomics Analysis
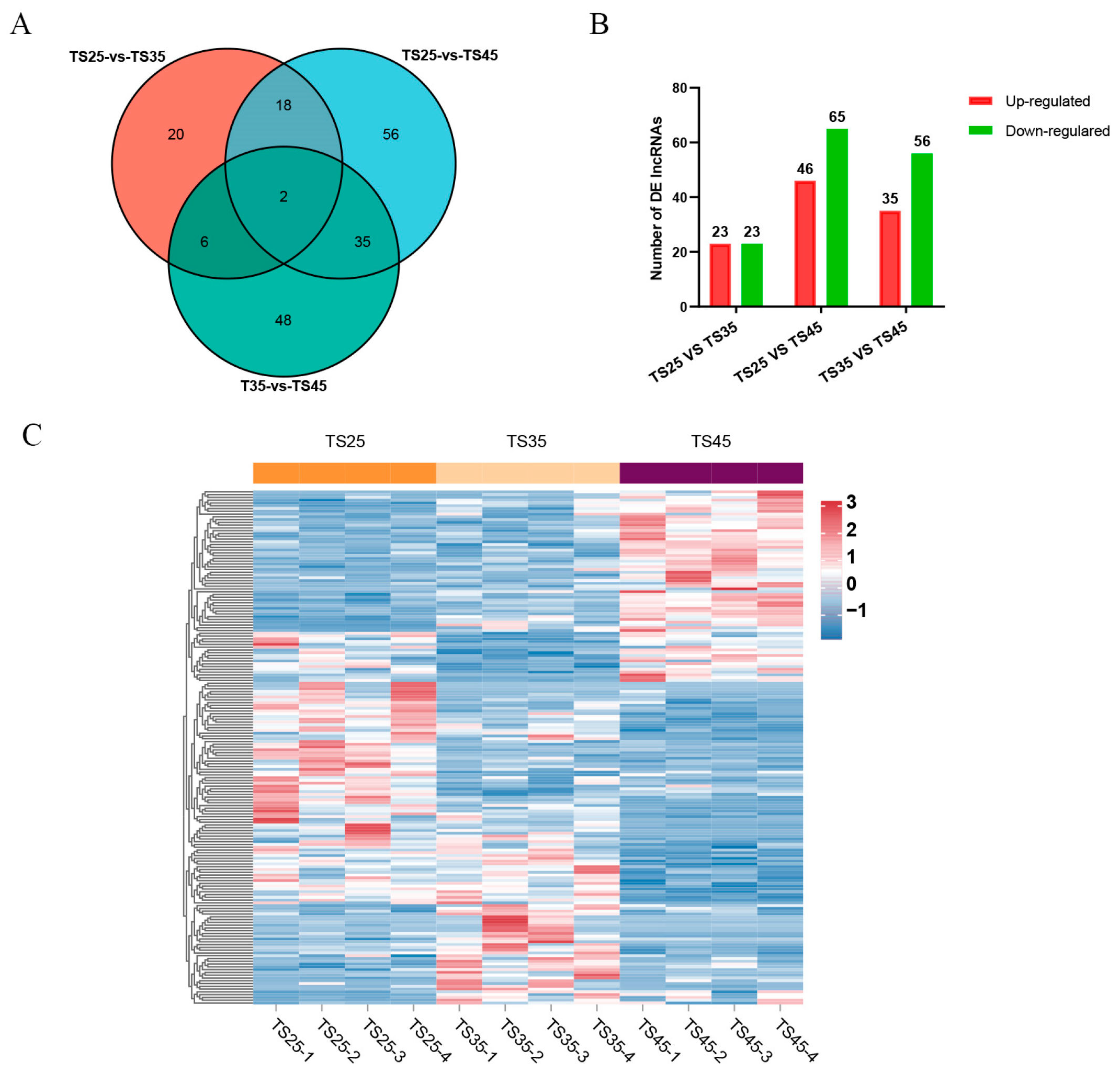
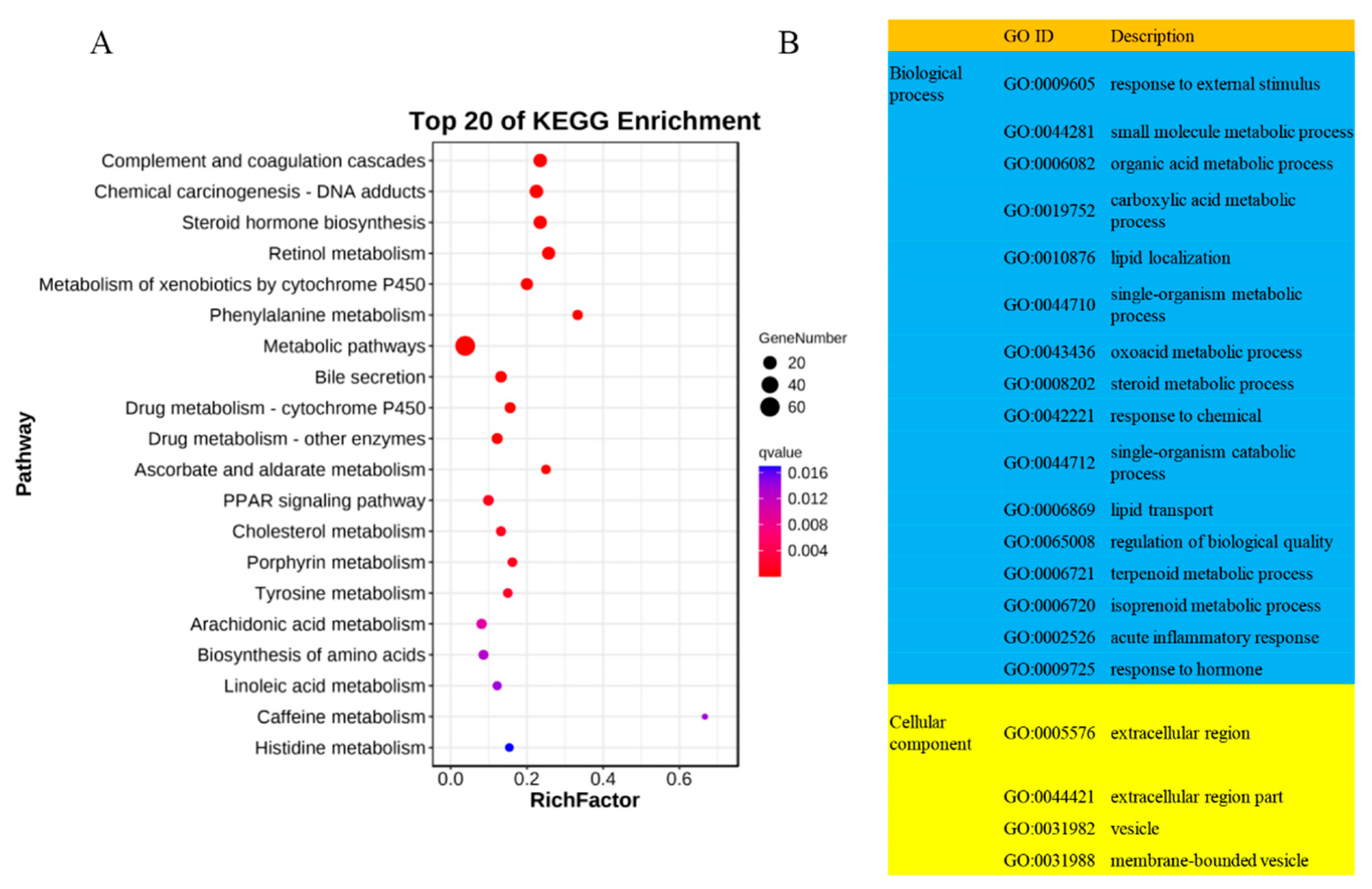
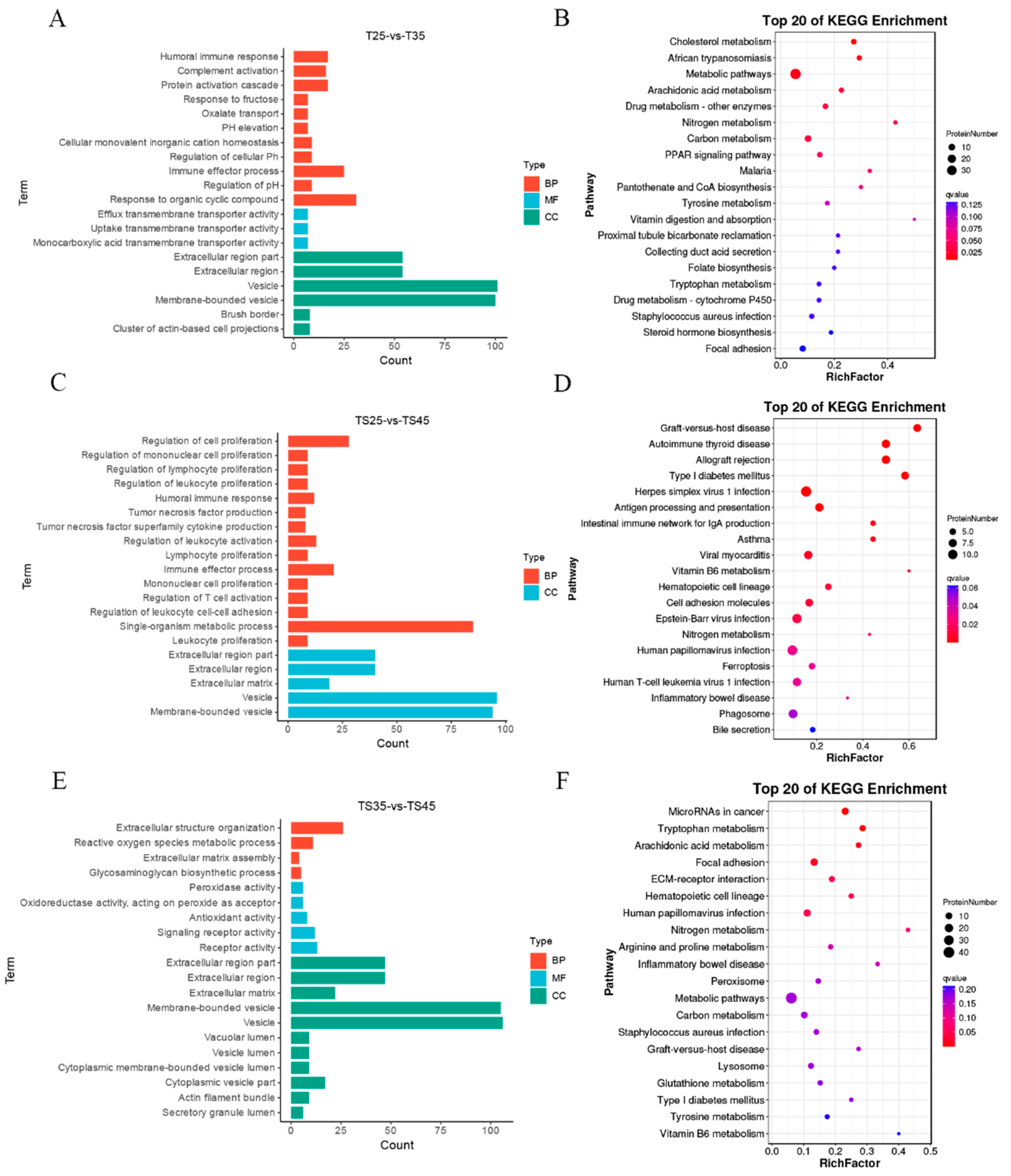
2.2. Functional Assessment of DE LncRNAs in Tibetan Sheep at Different Altitudes
2.3. Functional Assessment of DEPs in Tibetan Sheep at Different Altitudes
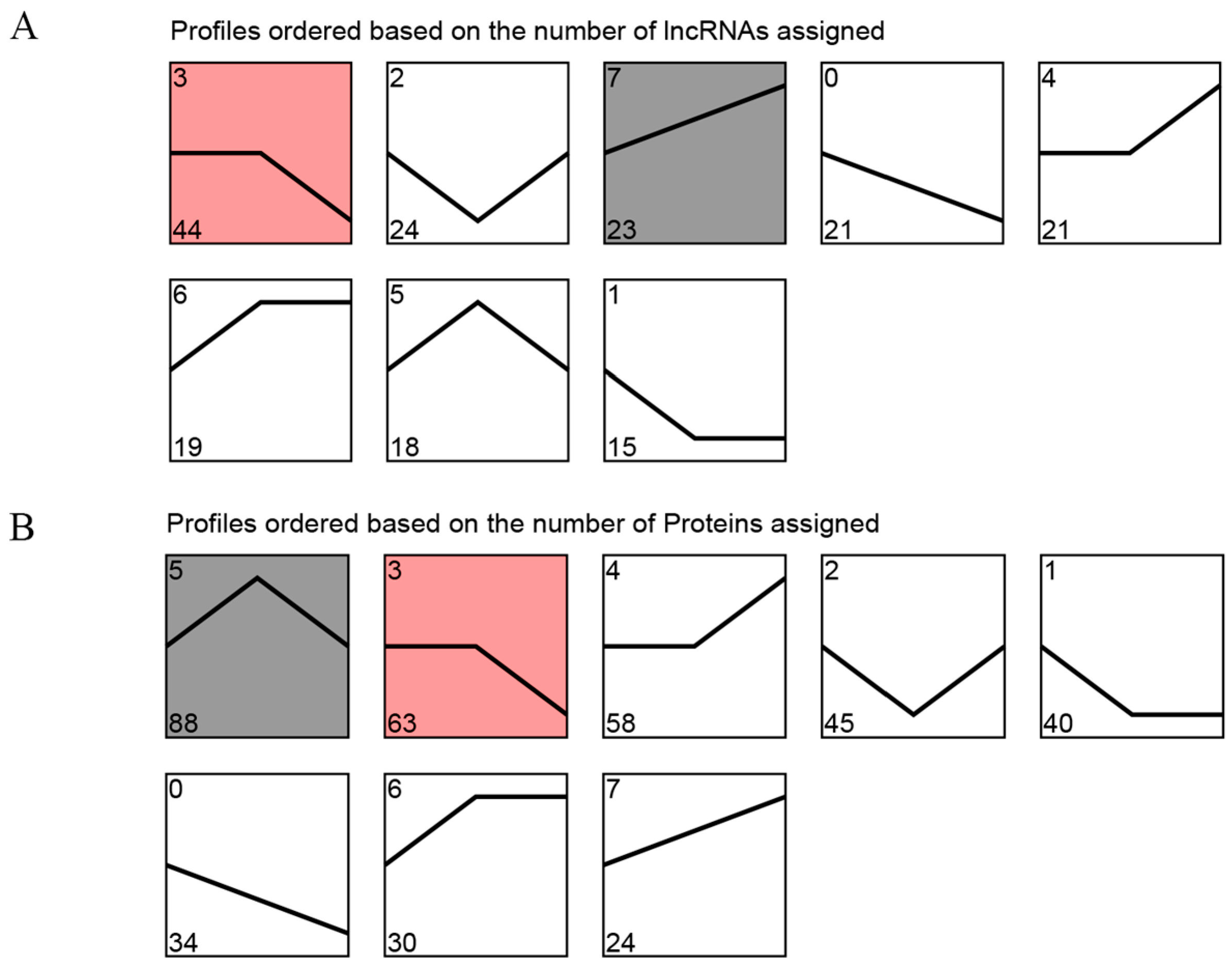
2.4. Trend Analysis of DE LncRNAs and DEPs
2.5. Protein–Protein Interaction Network Analysis of DEPs
2.6. Correlation between Transcriptome and Proteome
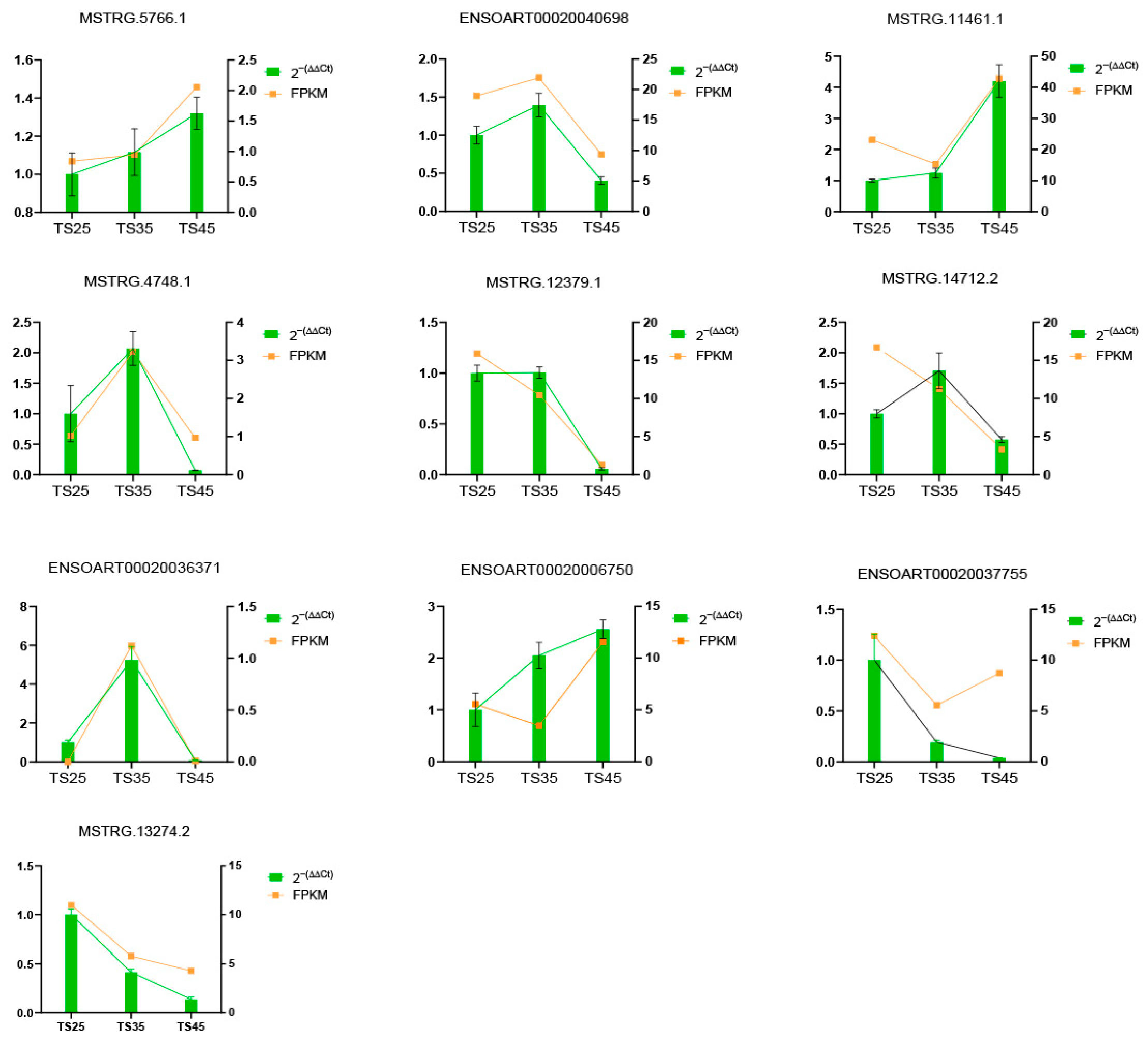
2.7. LncRNA Expression Validated by RT-qPCR
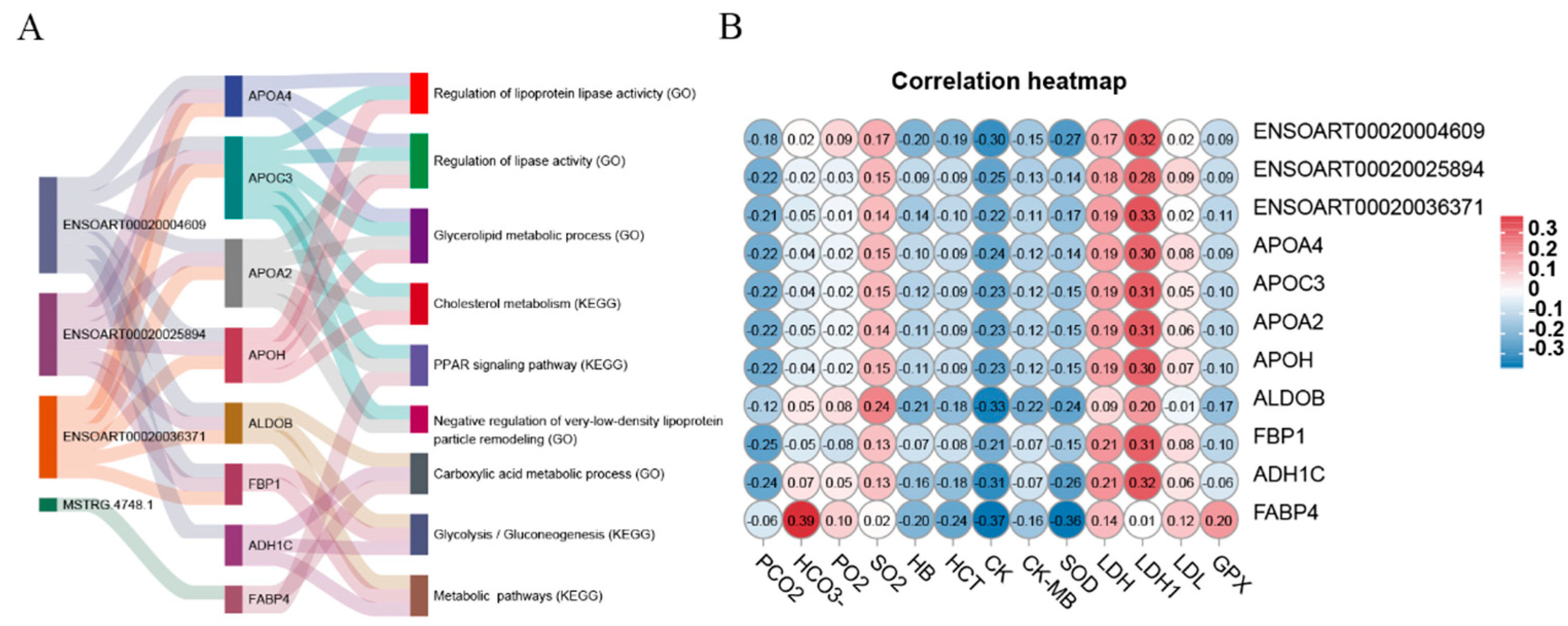
2.8. Correlation Analysis between Genes and Blood Physiological and Biochemical Indicators
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals and Sample Collection
4.3. Total RNA Extraction, cDNA Library Construction, and Sequencing
4.4. Quality Control, Reference Genome Alignment, and Differential Expression Analysis
4.5. Sample Preparation for Mass Spectrometry
4.6. Construction and Spectral Library of Qualitative Databases in Data-Dependent Acquisition
4.7. DIA Data Collection and Differential Expression Analysis
4.8. Functional Enrichment Analysis
4.9. RT-qPCR Analysis
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thompson, L.G.; Yao, T.; Mosley-Thompson, E.; Davis, M.E.; Henderson, K.A.; Lin, P.-N. A High-Resolution Millennial Record of the South Asian Monsoon from Himalayan Ice Cores. Science 2000, 289, 1916–1919. [Google Scholar] [CrossRef] [PubMed]
- He, J.K.; Lin, S.L.; Li, J.T.; Yu, J.H.; Jiang, H.S. Evolutionary history of zoogeographical regions surrounding the Tibetan Plateau. Commun. Biol. 2020, 3, 415. [Google Scholar] [CrossRef] [PubMed]
- West, J.B. Physiological Effects of Chronic Hypoxia. N. Engl. J. Med. 2017, 376, 1965–1971. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.J.; Li, G.Z.; Chen, S.M.; Han, J.L.; Hanif, Q. Rich maternal and paternal genetic diversity and divergent lineage composition in wild yak (Bos mutus). Anim. Biotechnol. 2022, 33, 1318–1321. [Google Scholar] [CrossRef]
- Ge, R.L.; Cai, Q.L.; Shen, Y.Y.; San, A.; Ma, L.; Zhang, Y.; Yi, X.; Chen, Y.; Yang, L.F.; Huang, Y.; et al. Draft genome sequence of the Tibetan antelope. Nat. Commun. 2013, 4, 1858. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.J.; Yang, J.; Xie, X.L.; Lv, F.H.; Cao, Y.H.; Li, W.R.; Liu, M.J.; Wang, Y.T.; Li, J.Q.; Liu, Y.G.; et al. The Genome Landscape of Tibetan Sheep Reveals Adaptive Introgression from Argali and the History of Early Human Settlements on the Qinghai-Tibetan Plateau. Mol. Biol. Evol. 2019, 36, 283–303. [Google Scholar] [CrossRef]
- Xin, G.S.; Long, R.J.; Guo, X.S.; Irvine, J.; Ding, L.M.; Ding, L.L.; Shang, Z.H. Blood mineral status of grazing Tibetan sheep in the Northeast of the Qinghai-Tibetan Plateau. Livest. Sci. 2011, 136, 102–107. [Google Scholar] [CrossRef]
- Persson, P.B.; Bondke Persson, A. Altitude sickness and altitude adaptation. Acta Physiol. 2017, 220, 303–306. [Google Scholar] [CrossRef]
- Beall, C.M. Adaptation to High Altitude: Phenotypes and Genotypes. Annu. Rev. Anthropol. 2014, 43, 251–272. [Google Scholar] [CrossRef]
- Ivy, C.M.; Scott, G.R. Control of breathing and the circulation in high-altitude mammals and birds. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 186, 66–74. [Google Scholar] [CrossRef]
- Stembridge, M.; Williams, A.M.; Gasho, C.; Dawkins, T.G.; Drane, A.; Villafuerte, F.C.; Levine, B.D.; Shave, R.; Ainslie, P.N. The overlooked significance of plasma volume for successful adaptation to high altitude in Sherpa and Andean natives. Proc. Natl. Acad. Sci. USA 2019, 116, 16177–16179. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.Y.; Li, J.B.; Qin, J.; Zhang, J.H.; Gao, X.B.; Yu, S.Y.; Yu, J.; Chen, G.Z.; Xu, B.D.; Li, H.J.; et al. Left Ventricular Function during Acute High-Altitude Exposure in a Large Group of Healthy Young Chinese Men. PLoS ONE 2015, 10, e0116936. [Google Scholar] [CrossRef]
- Naeije, R. Pulmonary hypertension at high altitude. Eur. Respir. J. 2019, 53, 1900985. [Google Scholar] [CrossRef] [PubMed]
- Teppema, L.J.; Dahan, A. The Ventilatory Response to Hypoxia in Mammals: Mechanisms, Measurement, and Analysis. Physiol. Rev. 2010, 90, 675–754. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; He, Y.Y.; Luo, Y.Z. Expression of OPA1 and Mic60 genes and their association with mitochondrial cristae morphology in Tibetan sheep. Cell Tissue Res. 2019, 376, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Penaloza, D.; Arias-Stella, J. The Heart and Pulmonary Circulation at High Altitudes. Circulation 2007, 115, 1132–1146. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.Q.; Long, K.R.; Ma, J.D.; Zhang, J.W.; He, D.F.; Jin, L.; Tang, Q.Z.; Jiang, A.A.; Wang, X.; Hu, Y.D.; et al. Comparative analysis of the microRNA transcriptome between yak and cattle provides insight into high-altitude adaptation. PeerJ 2017, 5, e3959. [Google Scholar] [CrossRef]
- Ayalew, W.; Chu, M.; Liang, C.; Wu, X.; Yan, P. Adaptation Mechanisms of Yak (Bos grunniens) to High-Altitude Environmental Stress. Animals 2021, 11, 2344. [Google Scholar] [CrossRef]
- Ai, H.S.; Yang, B.; Li, J.; Xie, X.H.; Chen, H.; Ren, J. Population history and genomic signatures for high-altitude adaptation in Tibetan pigs. BMC Genom. 2014, 15, 834. [Google Scholar] [CrossRef]
- Song, S.; Yao, N.; Yang, M.; Liu, X.X.; Dong, K.Z.; Zhao, Q.J.; Pu, Y.B.; He, X.H.; Guan, W.J.; Yang, N.; et al. Exome sequencing reveals genetic differentiation due to high-altitude adaptation in the Tibetan cashmere goat (Capra hircus). BMC Genom. 2016, 17, 122. [Google Scholar] [CrossRef]
- Choudhary, C.; Sharma, S.; Meghwanshi, K.K.; Patel, S.; Mehta, P.; Shukla, N.; Do, D.N.; Rajpurohit, S.; Suravajhala, P.; Shukla, J.N. Long Non-Coding RNAs in Insects. Animals 2021, 11, 1118. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.-H.; Guo, J.U. Coding functions of “noncoding” RNAs. Science 2020, 367, 1074–1075. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, N. LncRNAs on guard. Int. Immunopharmacol. 2018, 65, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ren, Q.L.; Hua, L.S.; Chen, J.F.; Zhang, J.Q.; Bai, H.J.; Li, H.L.; Xu, B.; Shi, Z.H.; Cao, H.; et al. Comprehensive Analysis of Differentially Expressed mRNA, lncRNA and circRNA and Their ceRNA Networks in the Longissimus Dorsi Muscle of Two Different Pig Breeds. Int. J. Mol. Sci. 2019, 20, 1107. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Z.X.; Yang, D.D.; Liu, P.Q.; Wang, Z.Q.; Zeng, Y.Q.; Chen, W. Diquat Determines a Deregulation of lncRNA and mRNA Expression in the Liver of Postweaned Piglets. Oxidative Med. Cell. Longev. 2019, 2019, 9148535. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, X.J.; Sun, X.L. The Functions of Long Non-Coding RNA (lncRNA) H19 in the Heart. Heart Lung Circ. 2022, 31, 341–349. [Google Scholar] [CrossRef]
- Zhang, T.; Cheng, G.W.; Sun, L.; Deng, L.; Wang, X.; Bi, N. Transcriptome alteration spectrum in rat lung induced by radiotherapy. Sci. Rep. 2019, 9, 19701. [Google Scholar] [CrossRef]
- Kong, Z.; Zhou, C.; Chen, L.; Ren, A.; Zhang, D.; Basang, Z.; Tan, Z.; Kang, J.; Li, B. Multi-Omics Analysis Reveals Up-Regulation of APR Signaling, LXR/RXR and FXR/RXR Activation Pathways in Holstein Dairy Cows Exposed to High-Altitude Hypoxia. Animals 2019, 9, 406. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, C.H.; Yan, X.H.; Jiang, G.C.; Dang, Q.; Wang, L.Y.; Liu, X.J. Physicochemical and functional properties of the muscle protein fraction of Hypomesus olidus. Food Chem. X 2022, 16, 100484. [Google Scholar] [CrossRef]
- Li, X.; Ren, Y.; Sorokin, V.; Poh, K.K.; Ho, H.H.; Lee, C.N.; Kleijn, D.D.; Lim, S.K.; Tam, J.P.; Sze, S.K. Quantitative profiling of the rat heart myoblast secretome reveals differential responses to hypoxia and re-oxygenation stress. J. Proteom. 2014, 98, 138–149. [Google Scholar] [CrossRef]
- Tang, N.; Liu, W.; Zhang, W.; Tang, D. Integrative analysis of transcriptomic and proteomic changes related to male sterility in Tagetes erecta. Physiol. Mol. Biol. Plants 2020, 26, 2061–2074. [Google Scholar] [CrossRef] [PubMed]
- Beall, C.M.; Cavalleri, G.L.; Deng, L.B.; Elston, R.C.; Gao, Y.; Knight, J.; Li, C.H.; Li, J.C.; Liang, Y.; McCormack, M.; et al. Natural selection on (α) associated with low hemoglobin concentration in Tibetan highlanders. Proc. Natl. Acad. Sci. USA 2010, 107, 11459–11464. [Google Scholar] [CrossRef] [PubMed]
- Bigham, A.; Bauchet, M.; Pinto, D.; Mao, X.Y.; Akey, J.M.; Mei, R.; Scherer, S.W.; Julian, C.G.; Wilson, M.J.; Herráez, D.L.; et al. Identifying Signatures of Natural Selection in Tibetan and Andean Populations Using Dense Genome Scan Data. PLoS Genet. 2010, 6, e1001116. [Google Scholar] [CrossRef]
- Brewster, L.M.; Haan, Y.C.; Zwinderman, A.H.; van den Born, B.J.; van Montfrans, G.A. CK (Creatine Kinase) Is Associated With Cardiovascular Hemodynamics The HELIUS Study. Hypertension 2020, 76, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Taufalele, P.V.; Cochran, J.D.; Robillard-Frayne, I.; Marx, J.M.; Soto, J.; Rauckhorst, A.J.; Tayyari, F.; Pewa, A.D.; Gray, L.R.; et al. Mitochondrial pyruvate carriers are required for myocardial stress adaptation. Nat. Metab. 2020, 2, 1248–1264. [Google Scholar] [CrossRef] [PubMed]
- Lother, A.; Bergemann, S.; Deng, L.; Moser, M.; Bode, C.; Hein, L. Cardiac Endothelial Cell Transcriptome. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 566–574. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.B.; Zhang, Q.; He, Y.X.; Yang, L.X.; Zhang, X.M.; Shi, P.; Yang, L.P.; Liu, Z.H.; Zhang, F.H.; Liu, F.Y.; et al. The Transcriptomic Landscape of Yaks Reveals Molecular Pathways for High Altitude Adaptation. Genome Biol. Evol. 2019, 11, 72–85. [Google Scholar] [CrossRef]
- Jin, M.Y.; Fuller, G.G.; Han, T.; Yao, Y.; Alessi, A.F.; Freeberg, M.A.; Roach, N.P.; Moresco, J.J.; Karnovsky, A.; Baba, M.; et al. Glycolytic Enzymes Coalesce in G Bodies under Hypoxic Stress. Cell Rep. 2017, 20, 895–908. [Google Scholar] [CrossRef]
- Zeng, L.; Zhou, J.; Zhang, Y.; Wang, X.; Li, Y.; Song, J.; Shao, J.; Su, P. Paternal cadmium exposure induces glucolipid metabolic reprogramming in offspring mice via PPAR signaling pathway. Chemosphere 2023, 339, 139592. [Google Scholar] [CrossRef]
- Xu, K.Q.; Xia, P.; Liu, P.; Zhang, X. A six lipid metabolism related gene signature for predicting the prognosis of hepatocellular carcinoma. Sci. Rep. 2022, 12, 20781. [Google Scholar] [CrossRef]
- Krone, K.G.; Ward, A.K.; Madder, K.M.; Hendrick, S.; McKinnon, J.J.; Buchanan, F.C. Interaction of vitamin A supplementation level with genotype on intramuscular fat in beef steers. Animal 2016, 10, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Arnold, P.K.; Finley, L.W.S. Regulation and function of the mammalian tricarboxylic acid cycle. J. Biol. Chem. 2023, 299, 102838. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, M.; Chen, H.Y.; Rong, J.; Dufresne, L.; Yao, J.; Guo, X.Q.; Tsai, M.Y.; Tsimikas, S.; Post, W.S.; Vasan, R.S.; et al. Genome-Wide Association Study Highlights APOH as a Novel Locus for Lipoprotein(a) Levels—Brief Report. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 458–464. [Google Scholar] [PubMed]
- Eide, R.P.; Asplund, C.A. Altitude Illness: Update on Prevention and Treatment. Curr. Sports Med. Rep. 2012, 11, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Manferdelli, G.; Marzorati, M.; Easton, C.; Porcelli, S. Changes in prefrontal cerebral oxygenation and microvascular blood volume in hypoxia and possible association with acute mountain sickness. Exp. Physiol. 2021, 106, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Narvaez-Guerra, O.; Herrera-Enriquez, K.; Medina-Lezama, J.; Chirinos, J.A. Systemic Hypertension at High Altitude. Hypertension 2018, 72, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chang, C.; Zhang, J.R.; Jiang, Y.; Ni, B.Y.; Wang, Y.L. Prevalence and risk factors associated with hypertension and prehypertension in a working population at high altitude in China: A cross-sectional study. Environ. Health Prev. Med. 2017, 22, 19. [Google Scholar] [CrossRef]
- Elmasri, H.; Karaaslan, C.; Teper, Y.; Ghelfi, E.; Weng, M.; Ince, T.A.; Kozakewich, H.; Bischoff, J.; Cataltepe, S. Fatty acid binding protein 4 is a target of VEGF and a regulator of cell proliferation in endothelial cells. FASEB J. 2009, 23, 3865–3873. [Google Scholar] [CrossRef]
- Furuhashi, M.; Tuncman, G.; Görgün, C.Z.; Makowski, L.; Atsumi, G.; Vaillancourt, E.; Kono, K.; Babaev, V.R.; Fazio, S.; Linton, M.F.; et al. Treatment of diabetes and atherosclerosis by inhibiting fatty-acid-binding protein aP2. Nature 2007, 447, 959–965. [Google Scholar] [CrossRef]
- Dahlström, E.H.; Saksi, J.; Forsblom, C.; Uglebjerg, N.; Mars, N.; Thorn, L.M.; Harjutsalo, V.; Rossing, P.; Ahluwalia, T.S.; Lindsberg, P.J.; et al. The Low-Expression Variant of FABP4 Is Associated With Cardiovascular Disease in Type 1 Diabetes. Diabetes 2021, 70, 2391–2401. [Google Scholar] [CrossRef]
- Tsai, H.Y.; Wu, Y.W.; Tseng, W.K.; Leu, H.B.; Yin, W.H.; Lin, T.H.; Chang, K.C.; Wang, J.H.; Yeh, H.I.; Wu, C.C.; et al. Circulating fatty-acid binding-protein 4 levels predict CV events in patients after coronary interventions. J. Formos. Med. Assoc. 2021, 120, 728–736. [Google Scholar] [CrossRef]
- Dai, W.; Zhang, Z.; Yao, C.; Zhao, S. Emerging evidences for the opposite role of apolipoprotein C3 and apolipoprotein A5 in lipid metabolism and coronary artery disease. Lipids Health Dis. 2019, 18, 220. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.L.; Tian, M.; Hu, S.L.; Wang, Y.; Wang, D.W. Identifying functional non-coding variants in APOA5/A4/C3/A1 gene cluster associated with coronary heart disease. J. Mol. Cell. Cardiol. 2020, 144, 54–62. [Google Scholar] [CrossRef]
- Andraski, A.B.; Singh, S.A.; Lee, L.; Higashi, H.; Smith, N.; Aikawa, M.; Sacks, F.M. Abstract MP41: A Diet High in Carbohydrate and Low in Fat Alters the HDL Proteome and Metabolism of 9 HDL Proteins in Humans. Circulation 2019, 139, AMP41. [Google Scholar] [CrossRef]
- Rader, D.J.; Hovingh, G.K. HDL and cardiovascular disease. Lancet 2014, 384, 618–625. [Google Scholar] [CrossRef]
- Davidson, W.S.; Cooke, A.L.; Swertfeger, D.K.; Shah, A.S. The difference between high density lipoprotein subfractions and subspecies: An evolving model in cardiovascular disease and diabetes. Curr. Atheroscler. Rep. 2021, 23, 23. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Chang, T.; Li, X.Q.; Wang, R.; Wu, L. Dual effects of fructose on ChREBP and FoxO1/3α are responsible for AldoB up-regulation and vascular remodelling. Clin. Sci. 2017, 131, 309–325. [Google Scholar] [CrossRef]
- Humphrey, J.D. Mechanisms of Vascular Remodeling in Hypertension. Am. J. Hypertens. 2021, 34, 432–441. [Google Scholar] [CrossRef]
- Tanaka, L.Y.; Laurindo, F.R.M. Vascular remodeling: A redox-modulated mechanism of vessel caliber regulation. Free Radic. Biol. Med. 2017, 109, 11–21. [Google Scholar] [CrossRef]
- Hunt, L.C.; Demontis, F. Age-Related Increase in Lactate Dehydrogenase Activity in Skeletal Muscle Reduces Life Span in Drosophila. J. Gerontol. A Biol. 2022, 77, 259–267. [Google Scholar] [CrossRef]
- Yao, X.L.; Gordon, E.M.; Figueroa, D.M.; Barochia, A.V.; Levine, S.J. Emerging Roles of Apolipoprotein E and Apolipoprotein A-I in the Pathogenesis and Treatment of Lung Disease. Am. J. Respir. Cell Mol. Biol. 2016, 55, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.J.; Zheng, J.Y.; Chen, L.J.; You, S.F.; Huang, H.B. Role of apolipoproteins in the pathogenesis of obesity. Clin. Chim. Acta 2023, 545, 117359. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Li, S.; Zhao, F.; Wang, J.; Liu, X.; Hu, J.; Bao, G.; Luo, Y. Changes in the Mitochondrial Dynamics and Functions Together with the mRNA/miRNA Network in the Heart Tissue Contribute to Hypoxia Adaptation in Tibetan Sheep. Animals 2022, 12, 583. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef]
- Sun, L.; Luo, H.T.; Bu, D.C.; Zhao, G.G.; Yu, K.T.; Zhang, C.H.; Liu, Y.N.; Chen, R.S.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Average Raw Reads | Average Clean Reads | Average Remaining Clean Reads | Average Mapped Reads (%) |
|---|---|---|---|---|
| TS25 | 73,744,200 | 73,433,514 (99.58%) | 73,324,286 | 63,119,866 (86.08%) |
| TS35 | 72,917,734 | 72,677,193 (99.67%) | 72,526,068 | 62,588,618 (86.30%) |
| TS45 | 89,686,072 | 89,332,025 (99.61%) | 89,184,858 | 78,171,553 (87.65%) |
| Blood Physiological Indexes | Tibetan Sheep | ||
| TS25 | TS35 | TS45 | |
| Oxygen pressure, PO2 (mmHg) | 37.75 ± 2.02 a | 34.00 ± 1.78 a | 28.50 ± 1.04 b |
| Oxygen saturation, SO2 (%) | 72.00 ± 0.82 a | 65.75 ± 0.91 b | 59.75 ± 0.83 c |
| Hemoglobin, HGB (g/dL) | 11.60 ± 0.44 c | 13.05 ± 0.44 b | 15.80 ± 0.37 a |
| Hematocrit, HCT (%PCV) | 34.75 ± 0.85 c | 39.50 ± 0.87 b | 50.50 ± 0.87 a |
| Potential of hydrogen, pH | 7.37 ± 0.03 a | 7.33 ± 0.02 a | 7.31 ± 0.01 a |
| Carbon dioxide pressure, PCO2 (mmHg) | 55.70 ± 1.42 a | 40.75 ± 1.19 b | 34.92 ± 1.64 c |
| Concentration of bicarbonate, HCO3− (mmol/L) | 27.35 ± 0.79 a | 25.38 ± 0.83 a | 25.03 ± 0.81 a |
| Base excess, BE (mmol/L) | 3.00 ± 0.41 a | 3.25 ± 0.48 a | 3.75 ± 0.48 a |
| Blood Biochemical Indexes | TS25 | TS35 | TS45 |
| Creatine kinase, CK (U/L) | 256.60 ± 4.43 c | 300.34 ± 4.31 b | 499.62 ± 6.38 a |
| Creatine kinase isoenzymes, CK-MB (U/L) | 42.12 ± 1.86 c | 49.70 ± 1.90 b | 58.91 ± 2.12 a |
| Lactate dehydrogenase, LDH (U/L) | 616.57 ± 7.68 c | 833.31 ± 6.37 b | 906.08 ± 5.09 a |
| Lactate dehydrogenase isoenzymes, LDH1 (U/L) | 131.34 ± 4.25 b | 156.68 ± 4.48 a | 164.78 ± 5.23 a |
| Superoxide dismutase, SOD (U/mL) | 198.03 ± 4.77 b | 206.28 ± 4.59 b | 244.20 ± 4.65 a |
| Glutathione peroxidase, GPX (U/mL) | 54.76 ± 3.03 b | 64.83 ± 2.46 ab | 69.06 ± 5.49 a |
| Low-density lipoprotein, LDL (mmol/L) | 1.26 ± 0.11 a | 1.23 ± 0.14 a | 1.23 ± 0.12 a |
| LncRNAs | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| MSTRG.5766.1 | TGAAGCCAGGTCCCTCCTAA | GCCCCAGACCTGGTGAATTA |
| ENSOART00020040698 | GGTCCTCTCCCTCTGTTGAC | AAATACCGCCCATCTCCACC |
| MSTRG.11461.1 | CGGTGTCTCACTGGTAGCTC | ACTCTCGCCTTCGTTCACAG |
| MSTRG.4748.1 | GAGCCCGGAACCCGAAATAG | CGTATCCAACAGTGCCTCGT |
| MSTRG.12379.1 | AGGAACCAACGTACCTGTCTC | TGCTTCCTGGTCCTATAGCAGT |
| MSTRG.14712.2 | AGGGGGAGTGTTAAGTGGGT | GATCCAACAACCCCACAGGA |
| ENSOART00020036371 | CACTGCTACCCGTTGAGGAA | TTCCCCGGTTGCATTCTGTC |
| ENSOART00020006750 | CTTTGTGGTTCTCCCCGTTTTC | GTCTCTGCTGTGGGTCTCCTG |
| ENSOART00020037755 | CCGACCTCAGTGAAGGGAAC | CCTCTGCTCCAAGACTGACG |
| MSTRG.13274.2 | GGGGCATCTGAGAACAACCA | AAACGAGTGCAGCTTTGCAG |
| β-actin | AGCCTTCCTTCCTGGGCATGGA | GGACAGCACCGTGTTGGCGTAA |
| GAPDH | GTCGGAGTGAACGGATTTGG | ACGATGTCCACTTTGCCAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Li, S.; Zhao, F.; Sun, H.; Hu, J.; Wang, J.; Liu, X.; Li, M.; Zhao, Z.; Luo, Y. LncRNA and Protein Expression Profiles Reveal Heart Adaptation to High-Altitude Hypoxia in Tibetan Sheep. Int. J. Mol. Sci. 2024, 25, 385. https://doi.org/10.3390/ijms25010385
He Z, Li S, Zhao F, Sun H, Hu J, Wang J, Liu X, Li M, Zhao Z, Luo Y. LncRNA and Protein Expression Profiles Reveal Heart Adaptation to High-Altitude Hypoxia in Tibetan Sheep. International Journal of Molecular Sciences. 2024; 25(1):385. https://doi.org/10.3390/ijms25010385
Chicago/Turabian StyleHe, Zhaohua, Shaobin Li, Fangfang Zhao, Hongxian Sun, Jiang Hu, Jiqing Wang, Xiu Liu, Mingna Li, Zhidong Zhao, and Yuzhu Luo. 2024. "LncRNA and Protein Expression Profiles Reveal Heart Adaptation to High-Altitude Hypoxia in Tibetan Sheep" International Journal of Molecular Sciences 25, no. 1: 385. https://doi.org/10.3390/ijms25010385
APA StyleHe, Z., Li, S., Zhao, F., Sun, H., Hu, J., Wang, J., Liu, X., Li, M., Zhao, Z., & Luo, Y. (2024). LncRNA and Protein Expression Profiles Reveal Heart Adaptation to High-Altitude Hypoxia in Tibetan Sheep. International Journal of Molecular Sciences, 25(1), 385. https://doi.org/10.3390/ijms25010385