
Balance between Pro- and Antifibrotic Proteins in Mesenchymal Stromal Cell Secretome Fractions Revealed by Proteome and Cell Subpopulation Analysis

 , , , ,
, , , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
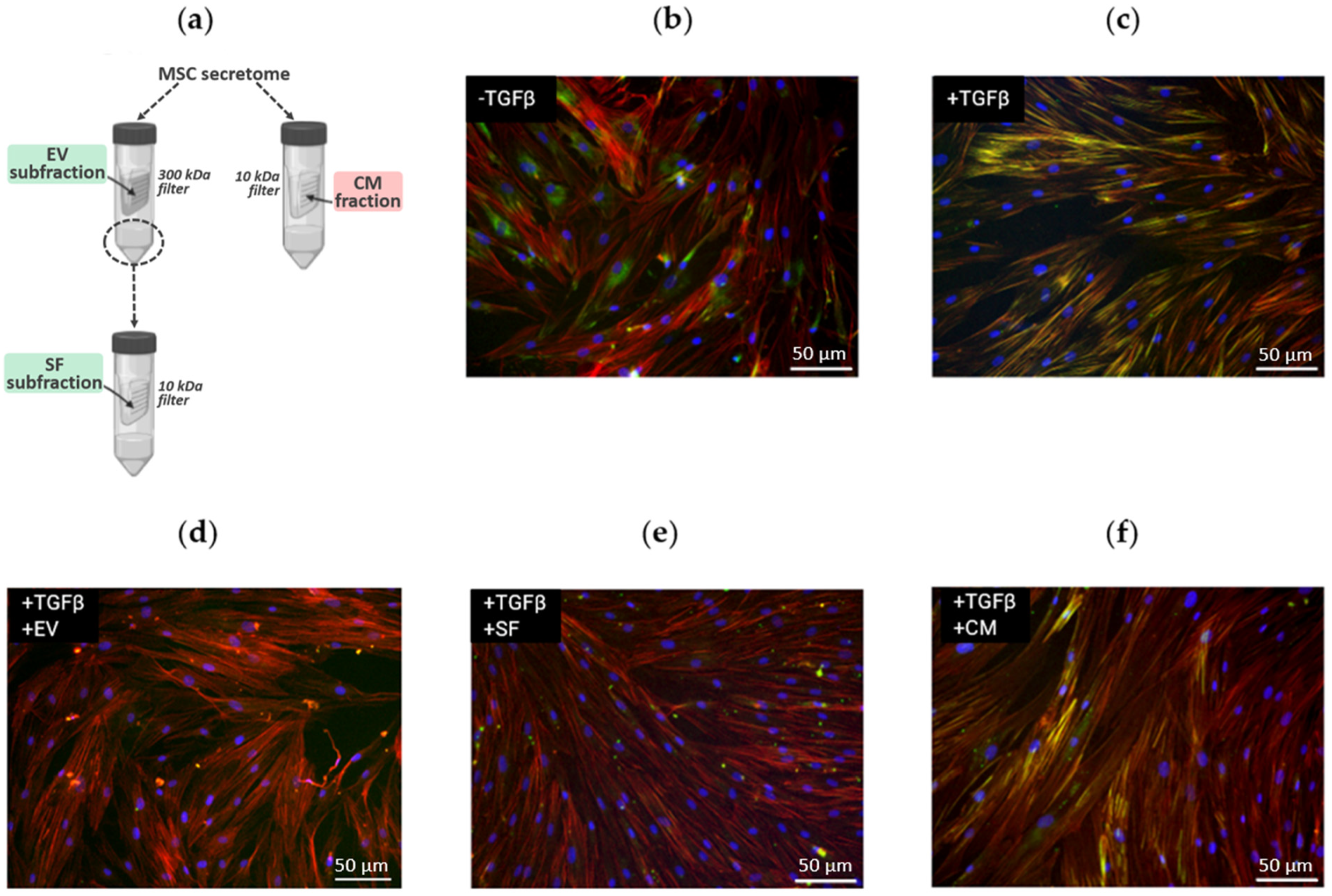
2.1. Distinct Effect of MSC Secretome Fractions on Fibroblast-to-Myofibroblast Differentiation Induced by TGF-β1
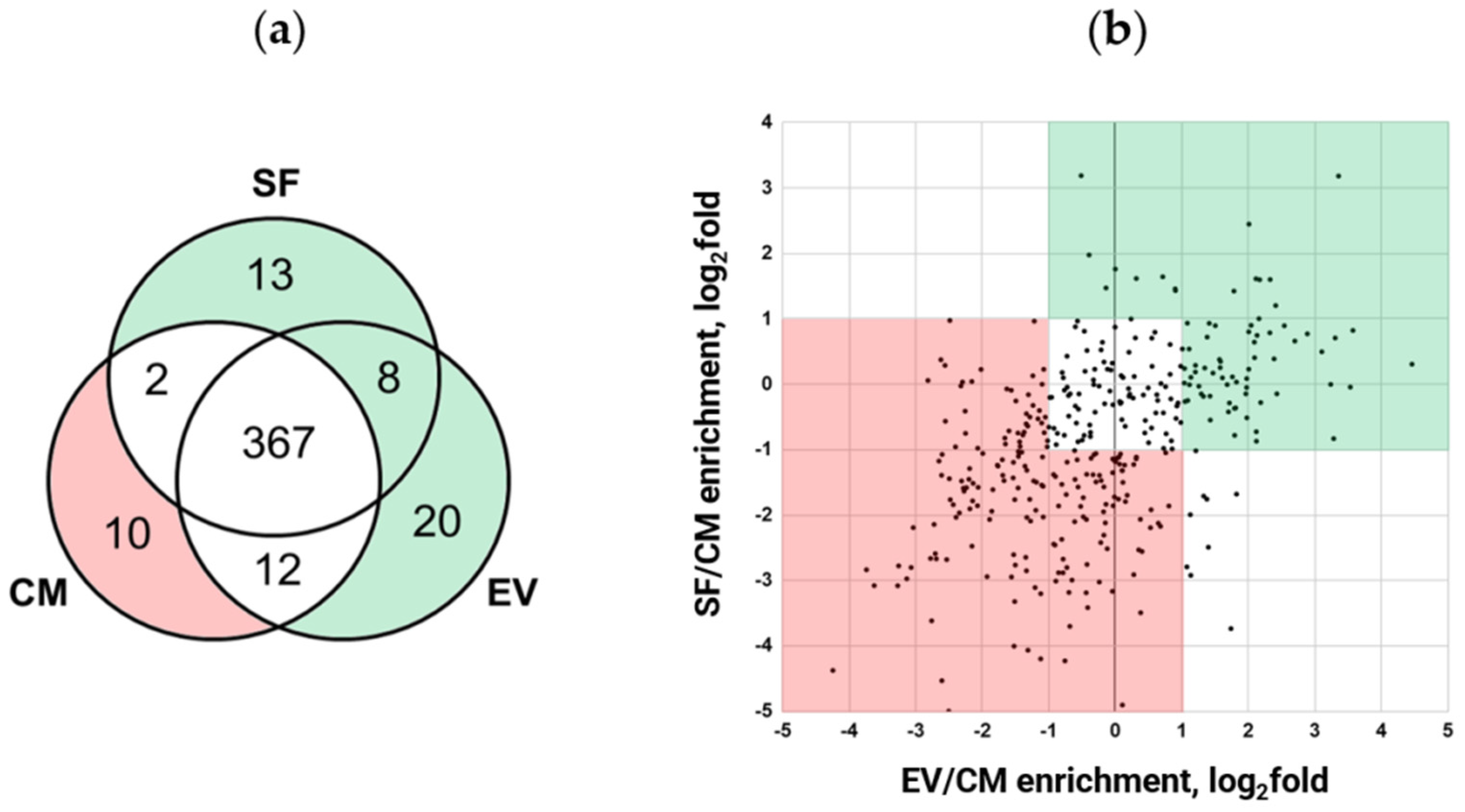
2.2. Fractionation Alters Quantitative Protein Content of MSC Secretome
2.3. Proteome Data Analysis of MSC Secretome Fractions Revealed Proteins Involved in NF-κB, TGF-β1, Wnt, Notch, and IGF Signaling Pathways
2.3.1. Total MSC Secretome Fraction, Not Able to Efficiently Prevent Myofibroblast Differentiation, Is Enriched in Proteins Involved in NF-κB Pathway Activation
2.3.2. Subfractions Preventing Myofibroblast Differentiation Are Enriched in Proteins Involved in the Regulation of TGF-β1, Wnt, Notch, and IGF Signaling Pathways
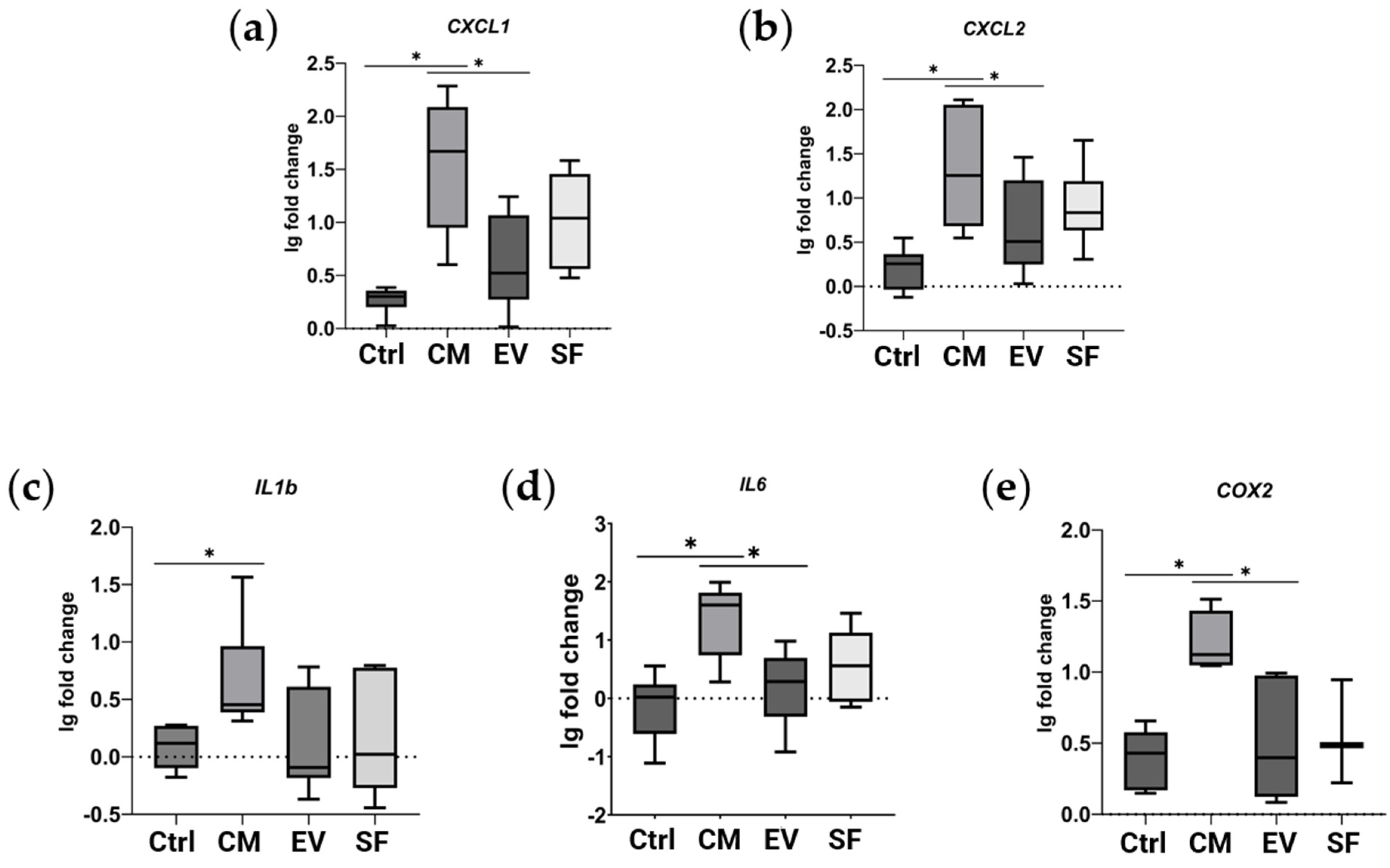
2.4. Gene Set Revealed through Proteomic Analysis Is Affected in a Subpopulation of MSCs under Profibrotic Conditions
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Conditioning and Isolation of Secretome Fractions
4.3. In Vitro Model of Fibroblast-to-Myofibroblast Differentiation
4.4. Immunocytochemistry
4.5. Sample Preparation for Proteomic Analysis
4.6. Bioinformatics Data Processing
4.7. Western Blotting
4.8. qPCR
4.9. Statistical Data Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darby, I.A.; Zakuan, N.; Billet, F.; Desmoulière, A. The myofibroblast, a key cell in normal and pathological tissue repair. Cell. Mol. Life Sci. 2016, 73, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.N.; Hardman, M.J. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10, 200223. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Fellabaum, C.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular mechanisms responsible for therapeutic potential of mesenchymal stem cell-derived secretome. Cells 2019, 8, 467. [Google Scholar] [CrossRef] [PubMed]
- Guillamat-Prats, R. The role of MSC in wound healing, scarring and regeneration. Cells 2021, 10, 1729. [Google Scholar] [CrossRef] [PubMed]
- Basalova, N.; Sagaradze, G.; Arbatskiy, M.; Evtushenko, E.; Kulebyakin, K.; Grigorieva, O.; Akopyan, Z.; Kalinina, N.; Efimenko, A. Secretome of mesenchymal stromal cells prevents myofibroblasts differentiation by transferring fibrosis-associated microRNAs within extracellular vesicles. Cells 2020, 9, 1272. [Google Scholar] [CrossRef] [PubMed]
- Basalova, N.; Arbatskiy, M.; Popov, V.; Grigorieva, O.; Vigovskiy, M.; Zaytsev, I.; Novoseletskaya, E.; Sagaradze, G.; Danilova, N.; Malkov, P.; et al. Mesenchymal stromal cells facilitate resolution of pulmonary fibrosis by miR-29c and miR-129 intercellular transfer. Exp. Mol. Med. 2023, 55, 1399–1412. [Google Scholar] [CrossRef]
- Aleksander, S.A.; Balhoff, J.; Carbon, S.; Cherry, J.M.; Drabkin, H.J.; Ebert, D.; Feuermann, M.; Gaudet, P.; Harris, N.L.; Hill, D.P.; et al. The Gene Ontology knowledgebase in 2023. Genetics 2023, 224, iyad031. [Google Scholar]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The reactome pathway knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef]
- Zhou, Q.; Tian, W.; Jiang, Z.; Huang, T.; Ge, C.; Liu, T.; Zhao, F.; Chen, T.; Cui, Y.; Li, H.; et al. A Positive Feedback Loop of AKR1C3-Mediated Activation of NF-κB and STAT3 Facilitates Proliferation and Metastasis in Hepatocellular Carcinoma. Cancer Res. 2021, 81, 1361–1374. [Google Scholar] [CrossRef] [PubMed]
- Kitasato, L.; Yamaoka-Tojo, M.; Hashikata, T.; Ishii, S.; Kameda, R.; Shimohama, T.; Tojo, T.; Ako, J. Factor Xa in Mouse Fibroblasts May Induce Fibrosis More Than Thrombin. Int. Heart J. 2014, 55, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Tanaka, T.; Narazaki, M.; Kishimoto, T. Targeting interleukin-6 signaling in clinic. Immunity 2019, 50, 1007–1023. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Yang, K.; Wang, H.; Xiao, Y.; Zhang, M.; Yu, X.; Xu, T.; Bai, T.; Zhu, H. MFAP4 deficiency alleviates renal fibrosis through inhibition of NF-κB and TGF-β/Smad signaling pathways. FASEB J. 2020, 34, 14250–14263. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Li, W.; Bai, T.; Xiao, Y.; Yu, W.; Luo, P.; Cheng, F. Mindin deficiency alleviates renal fibrosis through inhibiting NF-κB and TGF-β/Smad pathways. J. Cell. Mol. Med. 2020, 24, 5740–5750. [Google Scholar] [CrossRef] [PubMed]
- Kucia, M.; Jankowski, K.; Reca, R.; Wysoczynski, M.; Bandura, L.; Allendorf, D.J.; Zhang, J.; Ratajczak, J.; Ratajczak, M.Z. CXCR4–SDF-1 signalling, locomotion, chemotaxis and adhesion. J. Mol. Histol. 2004, 35, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Hwangbo, C.; Tae, N.; Lee, S.; Kim, O.; Park, O.K.; Kim, J.; Kwon, S.H.; Lee, J.H. Syntenin regulates TGF-β1-induced Smad activation and the epithelial-to-mesenchymal transition by inhibiting caveolin-mediated TGF-β type I receptor internalization. Oncogene 2016, 35, 389–401. [Google Scholar] [CrossRef]
- Onai, T.; Takai, A.; Setiamarga, D.H.E.; Holland, L.Z. Essential role of Dkk3 for head formation by inhibiting Wnt/β-catenin and Nodal/Vg1 signaling pathways in the basal chordate amphioxus. Evol. Dev. 2012, 14, 338–350. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Gu, L.; Lan, M.; Liu, C.; Wang, M.; Su, Y.; Ge, M.; Wang, T.; Yu, Y.; et al. Islr regulates canonical Wnt signaling-mediated skeletal muscle regeneration by stabilizing Dishevelled-2 and preventing autophagy. Nat. Commun. 2018, 9, 5129. [Google Scholar] [CrossRef]
- Robertson, I.B.; Horiguchi, M.; Zilberberg, L.; Dabovic, B.; Hadjiolova, K.; Rifkin, D.B. Latent TGF-β-binding proteins. Matrix Biol. 2015, 47, 44–53. [Google Scholar] [CrossRef]
- Moren, A.; Olofsson, A.; Stenman, G.; Sahlin, P.; Kanzaki, T.; Claesson-Welsh, L.; ten Dijke, P.; Miyazono, K.; Heldin, C.H. Identification and characterization of LTBP-2, a novel latent transforming growth factor-beta-binding protein. J. Biol. Chem. 1994, 269, 32469–32478. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Sanz, A.B.; Fernandez-Fernandez, B.; Carrasco, S.; Ruiz-Ortega, M.; Cannata-Ortiz, P.; Ortiz, A.; Sanchez-Niño, M.D. MXRA5 is a TGF-β1-regulated human protein with anti-inflammatory and anti-fibrotic properties. J. Cell. Mol. Med. 2017, 21, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, H.; Liu, X. Emerging role of CCN family proteins in fibrosis. J. Cell. Physiol. 2021, 236, 4195–4206. [Google Scholar] [CrossRef] [PubMed]
- Oxvig, C. The role of PAPP-A in the IGF system: Location, location, location. J. Cell Commun. Signal. 2015, 9, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Peradziryi, H.; Kaplan, N.A.; Podleschny, M.; Liu, X.; Wehner, P.; Borchers, A.; Tolwinski, N.S. PTK7/Otk interacts with Wnts and inhibits canonical Wnt signalling. EMBO J. 2011, 30, 3729–3740. [Google Scholar] [CrossRef] [PubMed]
- Vallet, S.D.; Davis, M.N.; Barqué, A.; Thahab, A.H.; Ricard-Blum, S.; Naba, A. Computational and experimental characterization of the novel ECM glycoprotein SNED1 and prediction of its interactome. Biochem. J. 2021, 478, 1413–1434. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, L.B.; Exposito, J.Y.; Chuvin, N.; Pommier, R.M.; Cluzel, C.; Martel, S.; Sentis, S.; Bartholin, L.; Lethias, C.; Valcourt, U. Tenascin-X promotes epithelial-to-mesenchymal transition by activating latent TGF-β. J. Cell Biol. 2014, 205, 409–428. [Google Scholar] [CrossRef] [PubMed]
- Grigorieva, O.; Basalova, N.; Vigovskiy, M.; Arbatskiy, M.; Dyachkova, U.; Kulebyakina, M.; Kulebyakin, K.; Tyurin-Kuzmin, P.; Kalinina, N.; Efimenko, A. Novel Potential Markers of Myofibroblast Differentiation Revealed by Single-Cell RNA Sequencing Analysis of Mesenchymal Stromal Cells in Profibrotic and Adipogenic Conditions. Biomedicines 2023, 11, 840. [Google Scholar] [CrossRef]
- Kalinina, N.; Kharlampieva, D.; Loguinova, M.; Butenko, I.; Pobeguts, O.; Efimenko, A.; Efimenko, A.; Ageeva, L.; Sharonov, G.; Ischenko, D.; et al. Characterization of secretomes provides evidence for adipose-derived mesenchymal stromal cells subtypes. Stem Cell Res. Ther. 2015, 6, 221. [Google Scholar] [CrossRef]
- Hou, J.; Ma, T.; Cao, H.; Chen, Y.; Wang, C.; Chen, X.; Xiang, Z.; Han, X. TNF-α-induced NF-κB activation promotes myofibroblast differentiation of LR-MSCs and exacerbates bleomycin-induced pulmonary fibrosis. J. Cell. Physiol. 2018, 233, 2409–2419. [Google Scholar] [CrossRef]
- Meng, X.; Nikolic-Paterson, D.; Lan, H. TGF-β: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, T.M.; Saldías, A.; Irigo, M.; Zamora, J.V.; Perone, M.J.; Dewey, R.A. Effect of TGF-β1 Stimulation on the Secretome of Human Adipose-Derived Mesenchymal Stromal Cells. Stem Cells Trans. Med. 2015, 4, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Dees, C.; Chakraborty, D.; Distler, J.H.W. Cellular and molecular mechanisms in fibrosis. Exp. Dermatol. 2021, 30, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Kis, K.; Liu, X.; Hagood, J. Myofibroblast differentiation and survival in fibrotic disease. Expert. Rev. Mol. Med. 2011, 13, E27. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, F.C.; Swain, M.T.; Klychnikov, O.I.; Niazi, U.; Ivens, A.; Quintana, J.F.; Hensbergen, P.J.; Hokke, C.H.; Buck, A.H.; Hoffmann, K.F. Protein and small non-coding RNA-enriched extracellular vesicles are released by the pathogenic blood fluke Schistosoma mansoni. J. Extracell. Vesicles 2015, 4, 28665. [Google Scholar] [CrossRef] [PubMed]
- Candiano, G.; Bruschi, M.; Musante, L.; Santucci, L.; Ghiggeri, G.M.; Carnemolla, B.; Orecchia, P.; Zardi, L.; Righetti, P.G. Blue silver: A very sensitive colloidal Coomassie G-250 staining for proteome analysis. Electrophoresis 2004, 25, 1327–1333. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef] [PubMed]
- Schaab, C.; Geiger, T.; Stoehr, G.; Cox, J.; Mann, M. Analysis of high accuracy, quantitative proteomics data in the MaxQB database. Mol. Cell. Proteom. 2012, 11, M111.014068. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote) omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Nakao, S.; Ogtata, Y.; Shimizu, E.; Yamazaki, M.; Furuyama, S.; Sugiya, H. Tumor necrosis factor α (TNF-α)-induced prostaglanding E2 release is mediated by the activation of cyclooxygenase-2 (COX-2) transcription via NFκB in human gingival fibroblasts. Mol. Cell. Biochem. 2002, 238, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Kogan-Sakin, I.; Cohen, M.; Paland, N.; Madar, S.; Solomon, H.; Molchadsky, A.; Brosh, R.; Buganim, Y.; Goldfinger, N.; Klocker, H.; et al. Prostate stromal cells produce CXCL-1, CXCL-2, CXCL-3 and IL-8 in response to epithelia-secreted IL-1. Carcinogenesis 2009, 30, 698–705. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Gene Name | CM Fraction Enrichment, Fold | Regulation of IGF Transport and Uptake by IGFBPs 1 | Positive Regulation of NF-κB Activation 2 | |
|---|---|---|---|---|---|
| Relative to EV Subfraction | Relative to SF Subfraction | ||||
| Aldo-keto reductase family 1 member C3 | AKR1C3 | 4.4 | 2.8 | +[11] | |
| Disintegrin and metalloproteinase domain-containing protein 10 | ADAM10 | 3.0 | 6.3 | + | |
| Cadherin-2 | CDH2 | 5.0 | 2.3 | + | |
| Coagulation factor X | F10 | 5.4 | 2.9 | +[12] | |
| Protein FAM20A | FAM20A | 8.8 | 6.4 | + | |
| Stress-70 protein, mitochondrial | HSPA9 | Detected in CM only | + | ||
| Insulin-like growth factor-binding protein 7 | IGFBP7 | 2.8 | 2.5 | + | |
| Interleukin-6 | IL6 | 4.9 | 2.5 | + | +[13] |
| Microfibril-associated glycoprotein 4 | MFAP4 | 2.9 | 13.2 | +[14] | |
| Macrophage migration inhibitory factor | MIF | 18.9 | 26.3 | + | |
| Matrix-remodeling-associated protein 8 | MXRA8 | 2.9 | 2.8 | + | |
| Plasminogen activator inhibitor 2 | SERPINB2 | 3.8 | 6.3 | + | |
| Superoxide dismutase [Cu-Zn] | SOD1 | 2.5 | 13.7 | + | |
| Superoxide dismutase [Mn], mitochondrial | SOD2 | 2.2 | Not detected in SF | + | |
| Spondin-2 | SPON2 | Detected in CM only | +[15] | ||
| Stromal cell-derived factor 1 | CXCL12 | 5.0 | 3.2 | +[16] | |
| T-complex protein 1 subunit alpha | TCP1 | Not detected in EV | 3049 | + | |
| Metalloproteinase inhibitor 1 | TIMP1 | 2416 | 3472 | + | |
| Protein Name | Gene Name | Enrichment Relative to CM Fraction, Fold | Expression Fold Change in Cluster 4 | Interacts with Signaling Pathways; Reference | |
|---|---|---|---|---|---|
| For EV Subfraction | For SF Subfraction | ||||
| Cytoskeleton-associated protein 4 | CKAP4 | Detected in EV only | 1.54 | Regulation of IGF transport and uptake by IGFBPs 1 | |
| Extracellular serine/threonine protein kinase FAM20C | FAM20C | 4.4 | 0.7 | - | |
| Growth arrest-specific protein 6 | GAS6 | Detected in SF only | (not detected) | ||
| Interstitial collagenase | MMP1 | 0.4 | 2.4 | (not detected) | |
| PRSS23 | PRSS23 | Detected in SF only | 1.63 | ||
| Stanniocalcin-2 | STC2 | Detected in EV only | 1.91 | ||
| Caveolin-1 | CAV1 | 2.1 | - | 1.45 | TGF-β [17] |
| Dickkopf-related protein 3 | DKK3 | 1.5 | 2.1 | 1.26 | Wnt [18] |
| Immunoglobulin superfamily containing leucine-rich repeat protein | ISLR | Not detected in CM | 1.84 | Wnt [19] | |
| Latent-transforming growth factor beta-binding protein 1 | LTBP1 | 5.0 | 3.7 | - | TGF-β [20]; Regulation of IGF transport and uptake by IGFBPs 1 |
| Latent-transforming growth factor beta-binding protein 2 | LTBP2 | 9.4 | 1.2 | 1.69 | TGF-β [21] |
| Matrix-remodeling-associated protein 5 | MXRA5 | 2.2 | 0.2 | 1.23 | TGF-β [22] |
| Protein NOV homolog | NOV | Detected in SF only | (not detected) | IGF, Notch [23] | |
| Pappalysin-1 | PAPPA | 3.1 | 1.3 | 1.48 | IGF [24]; Regulation of IGF transport and uptake by IGFBPs 1 |
| Inactive tyrosine-protein kinase 7 | PTK7 | Not detected in CM | 1.54 | Wnt [25] | |
| Sushi, nidogen, and EGF-like domain-containing protein 1 | SNED1 | 3.5 | 1.3 | (not detected) | Notch (predicted, [26]) |
| Tenascin | TNC | 11.9 | 2.2 | 1.4 | TGF-β [27]; Regulation of IGF transport and uptake by IGFBPs 1 |
| Gene Name | Primer Sequence (f—Forward, r—Reverse) | Source |
|---|---|---|
| 36B4 | f 5′-AACCGAAGTCATAGCCACAC-3′ | PrimerBlast [40] |
| r 5′-AACCGAAGTCATAGCCACAC-3′ | ||
| COX2 | f 5′-ATGAGATTGTGGAAAAATTGCT-3′ | [41] |
| r 5′-GATCATCTCTGCCTGAGTATC-3′ | ||
| CXCL1 | f 5′-AGTCATAGCCACACTCAAGAATGG-3′ | [42] |
| r 5′-GATGCAGGATTGAGGCAAGC-3′ | ||
| CXCL2 | f 5′-AACCGAAGTCATAGCCACAC-3′ | PrimerBlast [40] |
| r 5′-AACCGAAGTCATAGCCACAC-3′ | ||
| IL1B | f 5′-AACCGAAGTCATAGCCACAC-3′ | [42] |
| r 5′-AACCGAAGTCATAGCCACAC-3′ | ||
| IL6 | f 5′-AACCGAAGTCATAGCCACAC-3′ | PrimerBlast [40] |
| r 5′-AACCGAAGTCATAGCCACAC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulebyakina, M.; Basalova, N.; Butuzova, D.; Arbatsky, M.; Chechekhin, V.; Kalinina, N.; Tyurin-Kuzmin, P.; Kulebyakin, K.; Klychnikov, O.; Efimenko, A. Balance between Pro- and Antifibrotic Proteins in Mesenchymal Stromal Cell Secretome Fractions Revealed by Proteome and Cell Subpopulation Analysis. Int. J. Mol. Sci. 2024, 25, 290. https://doi.org/10.3390/ijms25010290
Kulebyakina M, Basalova N, Butuzova D, Arbatsky M, Chechekhin V, Kalinina N, Tyurin-Kuzmin P, Kulebyakin K, Klychnikov O, Efimenko A. Balance between Pro- and Antifibrotic Proteins in Mesenchymal Stromal Cell Secretome Fractions Revealed by Proteome and Cell Subpopulation Analysis. International Journal of Molecular Sciences. 2024; 25(1):290. https://doi.org/10.3390/ijms25010290
Chicago/Turabian StyleKulebyakina, Maria, Nataliya Basalova, Daria Butuzova, Mikhail Arbatsky, Vadim Chechekhin, Natalia Kalinina, Pyotr Tyurin-Kuzmin, Konstantin Kulebyakin, Oleg Klychnikov, and Anastasia Efimenko. 2024. "Balance between Pro- and Antifibrotic Proteins in Mesenchymal Stromal Cell Secretome Fractions Revealed by Proteome and Cell Subpopulation Analysis" International Journal of Molecular Sciences 25, no. 1: 290. https://doi.org/10.3390/ijms25010290
APA StyleKulebyakina, M., Basalova, N., Butuzova, D., Arbatsky, M., Chechekhin, V., Kalinina, N., Tyurin-Kuzmin, P., Kulebyakin, K., Klychnikov, O., & Efimenko, A. (2024). Balance between Pro- and Antifibrotic Proteins in Mesenchymal Stromal Cell Secretome Fractions Revealed by Proteome and Cell Subpopulation Analysis. International Journal of Molecular Sciences, 25(1), 290. https://doi.org/10.3390/ijms25010290