

Zonulin as a Potential Therapeutic Target in Microbiota-Gut-Brain Axis Disorders: Encouraging Results and Emerging Questions
 ,
,
Abstract
1. Introduction
2. Zonulin
2.1. Zonulin as Pre-Haptoglobin 2
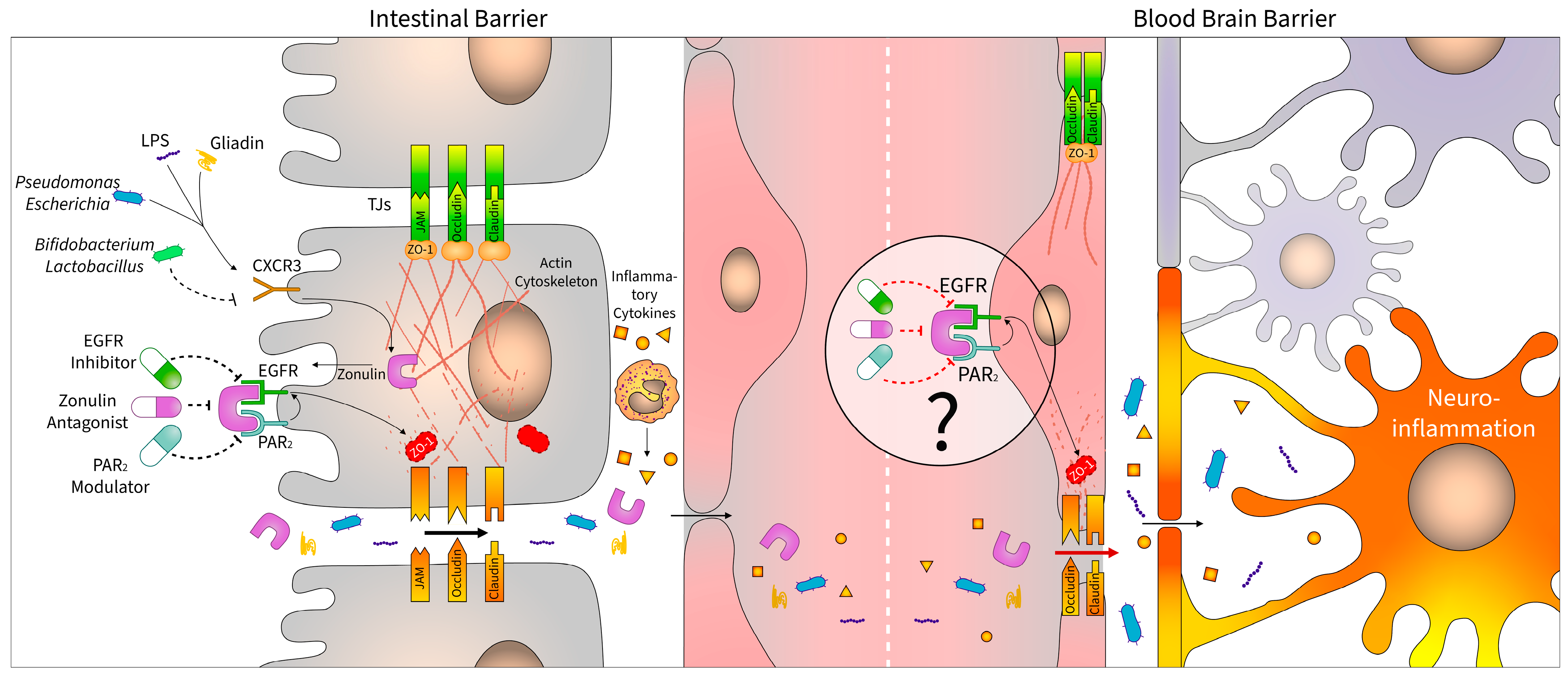
2.2. Regulation of Zonulin
2.3. Biological Activity of Zonulin
3. Diseases Associated with Altered Zonulin Levels
3.1. Central Nervous System Diseases
3.2. Viral Infections
4. Zonulin Pathway as a Therapeutic Target
4.1. Human Studies with Larazotide Acetate
4.2. Preclinical Studies with Larazotide Acetate
4.3. Future Perspectives of Zonulin Antagonists
4.4. Other Receptor Modulators

{kind=link}
| Target | Type | Compound | Cell Line | Effect on TJs and/or Transcellular Permeability | Ref. |
|---|---|---|---|---|---|
| PAR2 | peptidic antagonist | FSLLRY-NH2 | pHNECs | harmful | [121] |
| SLIGRL-NH2 | |||||
| non-peptidic full agonist | AC-55541 | hBMECs | protective | [237] | |
| small molecule antagonist | GB88 | A549 | [235] | ||
| hECs | [236] | ||||
| GB83 | Caco2 | harmful | [234] | ||
| EGFR | tyrosine kinase inhibitor | AG1478 | hCMEC/D3 | protective | [238] |
| Calu-3 | [239] | ||||
| HSC-3 | [240] | ||||
| erlotinib | IEC-6 | harmful | [241] | ||
| gefitinib | [242] | ||||
| icotinib | |||||
| dacomitinib | T84 | [244] | |||
| lapatinib | HBCCs | [245] | |||
| vandetanib | Calu-6 | [246] |
5. Considerations
5.1. Nomenclature
5.2. Technical Issues: Zonulin as a Biomarker and Therapeutic Target
6. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V. The microbiota-gut-brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Oriach, C.S.; Robertson, R.C.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Food for thought: The role of nutrition in the microbiota-gut–brain axis. Clin. Nutr. Exp. 2016, 6, 25–38. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L., IV; Mazmanian, S.K. The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Pap, D.; Veres-Székely, A.; Szebeni, B.; Vannay, Á. PARK7/DJ-1 as a Therapeutic Target in Gut-Brain Axis Diseases. Int. J. Mol. Sci. 2022, 23, 6626. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.-D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef]
- Camilleri, M. Leaky gut: Mechanisms, measurement and clinical implications in humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Tripathi, A.; Lammers, K.M.; Goldblum, S.; Shea-Donohue, T.; Netzel-Arnett, S.; Buzza, M.S.; Antalis, T.M.; Vogel, S.N.; Zhao, A.; Yang, S. Identification of human zonulin, a physiological modulator of tight junctions, as prehaptoglobin-2. Proc. Natl. Acad. Sci. USA 2009, 106, 16799–16804. [Google Scholar] [CrossRef]
- Sturgeon, C.; Fasano, A. Zonulin, a regulator of epithelial and endothelial barrier functions, and its involvement in chronic inflammatory diseases. Tissue Barriers 2016, 4, e1251384. [Google Scholar] [CrossRef]
- Fasano, A.; Not, T.; Wang, W.; Uzzau, S.; Berti, I.; Tommasini, A.; Goldblum, S.E. Zonulin, a newly discovered modulator of intestinal permeability, and its expression in coeliac disease. Lancet 2000, 355, 1518–1519. [Google Scholar] [CrossRef]
- Hoilat, G.J.; Altowairqi, A.K.; Ayas, M.F.; Alhaddab, N.T.; Alnujaidi, R.A.; Alharbi, H.A.; Alyahyawi, N.; Kamal, A.; Alhabeeb, H.; Albazee, E.; et al. Larazotide acetate for treatment of celiac disease: A systematic review and meta-analysis of randomized controlled trials. Clin. Res. Hepatol. Gastroenterol. 2022, 46, 101782. [Google Scholar] [CrossRef]
- Wang, W.; Uzzau, S.; Goldblum, S.E.; Fasano, A. Human zonulin, a potential modulator of intestinal tight junctions. J. Cell Sci. 2000, 113, 4435–4440. [Google Scholar] [CrossRef] [PubMed]
- Rittirsch, D.; Flierl, M.A.; Nadeau, B.A.; Day, D.E.; Huber-Lang, M.S.; Grailer, J.J.; Zetoune, F.S.; Andjelkovic, A.V.; Fasano, A.; Ward, P.A. Zonulin as prehaptoglobin2 regulates lung permeability and activates the complement system. Am. J. Physiol. Cell Mol. Physiol. 2013, 304, L863–L872. [Google Scholar] [CrossRef] [PubMed]
- Rittirsch, D.; Flierl, M.A.; Day, D.E.; Nadeau, B.A.; Werner, C.M.L.; Wanner, G.A.; Simmen, H.; Fasano, A.; Ward, P.A. Role of zonulin as prehaptoglobin2 in acute lung injury. FASEB J. 2011, 25, 114.1. [Google Scholar] [CrossRef]
- Ghanadi, K.; Naghdi, N. The Role of Zonulin as a Prognostic Biomarker in Liver Diseases: A Systematic Review. Adv. Life Sci. 2022, 9, 277–283. [Google Scholar]
- Yu, J.; Shen, Y.; Zhou, N. Advances in the role and mechanism of zonulin pathway in kidney diseases. Int. Urol. Nephrol. 2021, 53, 2081–2088. [Google Scholar] [CrossRef] [PubMed]
- Smecuol, E.; Sugai, E.; Niveloni, S.; Vázquez, H.; Pedreira, S.; Mazure, R.; Moreno, M.L.; Label, M.; Mauriño, E.; Fasano, A. Permeability, zonulin production, and enteropathy in dermatitis herpetiformis. Clin. Gastroenterol. Hepatol. 2005, 3, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Sheen, Y.; Jee, H.; Kim, D.; Ha, E.; Jeong, I.; Lee, S.; Baek, H.; Lee, S.; Lee, K.; Lee, K. Serum zonulin is associated with presence and severity of atopic dermatitis in children, independent of total IgE and eosinophil. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2018, 48, 1059–1062. [Google Scholar] [CrossRef]
- Miranda-Ribera, A.; Serena, G.; Liu, J.; Fasano, A.; Kingsbury, M.A.; Fiorentino, M.R. The Zonulin-transgenic mouse displays behavioral alterations ameliorated via depletion of the gut microbiota. Tissue Barriers 2022, 10, 2000299. [Google Scholar] [CrossRef]
- Uzzau, S.; Lu, R.; Wang, W.; Fiore, C.; Fasano, A. Purification and preliminary characterization of the zonula occludens toxin receptor from human (CaCo2) and murine (IEC6) intestinal cell lines. FEMS Microbiol. Lett. 2001, 194, 1–5. [Google Scholar] [CrossRef]
- Lu, R.; Wang, W.; Uzzau, S.; Vigorito, R.; Zielke, H.R.; Fasano, A. Affinity Purification and Partial Characterization of the Zonulin/Zonula Occludens Toxin (Zot) Receptor from Human Brain. J. Neurochem. 2001, 74, 320–326. [Google Scholar] [CrossRef]
- Fasano, A.; Baudry, B.; Pumplin, D.W.; Wasserman, S.S.; Tall, B.D.; Ketley, J.M.; Kaper, J. Vibrio cholerae produces a second enterotoxin, which affects intestinal tight junctions. Proc. Natl. Acad. Sci. USA 1991, 88, 5242–5246. [Google Scholar] [CrossRef] [PubMed]
- Baudry, B.; Fasano, A.; Ketley, J.; Kaper, J.B. Cloning of a gene (zot) encoding a new toxin produced by Vibrio cholerae. Infect. Immun. 1992, 60, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Fiorentini, C.; Donelli, G.; Uzzau, S.; Kaper, J.; Margaretten, K.; Ding, X.; Guandalini, S.; Comstock, L.; Goldblum, S.E. Zonula occludens toxin modulates tight junctions through protein kinase C-dependent actin reorganization, in vitro. J. Clin. Investig. 1995, 96, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Uzzau, S.; Fiore, C.; Margaretten, K. The enterotoxic effect of zonula occludens toxin on rabbit small intestine involves the paracellular pathway. Gastroenterology 1997, 112, 839–846. [Google Scholar] [CrossRef]
- Pérez-Reytor, D.; Pavón, A.; Lopez-Joven, C.; Ramírez-Araya, S.; Peña-Varas, C.; Plaza, N.; Alegría-Arcos, M.; Corsini, G.; Jaña, V.; Pavez, L.; et al. Analysis of the Zonula occludens Toxin Found in the Genome of the Chilean Non-toxigenic Vibrio parahaemolyticus Strain PMC53.7. Front. Cell Infect. Microbiol. 2020, 10, 482. [Google Scholar] [CrossRef]
- Mahendran, V.; Liu, F.; Riordan, S.M.; Grimm, M.C.; Tanaka, M.M.; Zhang, L. Examination of the effects of Campylobacter concisus zonula occludens toxin on intestinal epithelial cells and macrophages. Gut Pathog. 2016, 8, 18. [Google Scholar] [CrossRef]
- Zhang, L.; Lee, H.; Grimm, M.C.; Riordan, S.M.; Day, A.S.; Lemberg, D.A. Campylobacter concisus and inflammatory bowel disease. World J.Gastroenterol. WJG 2014, 20, 1259. [Google Scholar] [CrossRef]
- Liu, F.; Lee, H.; Lan, R.; Zhang, L. Zonula occludens toxins and their prophages in Campylobacter species. Gut Pathog. 2016, 8, 1–11. [Google Scholar] [CrossRef]
- di Masi, A.; De Simone, G.; Ciaccio, C.; D’Orso, S.; Coletta, M.; Ascenzi, P. Haptoglobin: From hemoglobin scavenging to human health. Mol. Asp. Med. 2020, 73, 100851. [Google Scholar] [CrossRef]
- Andersen, C.B.F.; Stødkilde, K.; Saederup, K.L.; Kuhlee, A.; Raunser, S.; Graversen, J.H.; Moestrup, S.K. Haptoglobin. Antioxid. Redox Signal. 2017, 26, 814–831. [Google Scholar] [CrossRef]
- Sadrzadeh, S.H.; Bozorgmehr, J. Haptoglobin Phenotypes in Health and Disorders. Pathol. Patterns Rev. 2004, 121, S97–S104. [Google Scholar] [CrossRef] [PubMed]
- Jelena, A.; Mirjana, M.; Desanka, B.; Svetlana, I.-M.; Aleksandra, U.; Goran, P.; Ilijana, G. Haptoglobin and the inflammatory and oxidative status in experimental diabetic rats: Antioxidant role of haptoglobin. J. Physiol. Biochem. 2013, 69, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Naryzny, S.; Legina, O. Haptoglobin as a Biomarker. Biochem. Suppl. Ser. B Biomed. Chem. 2021 15, 184–198. [CrossRef]
- Vanuytsel, T.; Vermeire, S.; Cleynen, I. The role of Haptoglobin and its related protein, Zonulin, in inflammatory bowel disease. Tissue Barriers 2013, 1, e27321. [Google Scholar] [CrossRef] [PubMed]
- Stuart, C.M.; Varatharaj, A.; Winberg, M.E.; Galea, P.; Larsson, H.B.; Cramer, S.P.; Fasano, A.; Maherally, Z.; Pilkington, G.J.; Keita, Å.V. Zonulin and blood-brain barrier permeability are dissociated in humans. Clin. Transl. Med. 2022, 12, e965. [Google Scholar] [CrossRef] [PubMed]
- Schaer, C.A.; Owczarek, C.; Deuel, J.W.; Schauer, S.; Baek, J.H.; Yalamanoglu, A.; Hardy, M.P.; Scotney, P.D.; Schmidt, P.M.; Pelzing, M.; et al. Phenotype-specific recombinant haptoglobin polymers co-expressed with C1r-like protein as optimized hemoglobin-binding therapeutics. BMC Biotechnol. 2018, 18, 15. [Google Scholar] [CrossRef]
- Wassler, M.; Fries, E. Proteolytic cleavage of haptoglobin occurs in a subcompartment of the endoplasmic reticulum: Evidence from membrane fusion in vitro. J. Cell Biol. 1993, 123, 285–291. [Google Scholar] [CrossRef]
- Wicher, K.B.; Fries, E. Prohaptoglobin is proteolytically cleaved in the endoplasmic reticulum by the complement C1r-like protein. Proc. Natl. Acad. Sci. USA 2004, 101, 14390–14395. [Google Scholar] [CrossRef]
- Fasano, A. Zonulin and Its Regulation of Intestinal Barrier Function: The Biological Door to Inflammation, Autoimmunity, and Cancer. Physiol. Rev. 2011, 91, 151–175. [Google Scholar] [CrossRef]
- Buzzi, R.M.; Owczarek, C.M.; Akeret, K.; Tester, A.; Pereira, N.; Butcher, R.; Brügger-Verdon, V.; Hardy, M.P.; Illi, M.; Wassmer, A.; et al. Modular Platform for the Development of Recombinant Hemoglobin Scavenger Biotherapeutics. Mol. Pharm. 2021, 18, 3158–3170. [Google Scholar] [CrossRef]
- Ligoudistianou, C.; Xu, Y.; Garnier, G.; Circolo, A.; Volanakis, J.E. A novel human complement-related protein, C1r-like protease (C1r-LP), specifically cleaves pro-C1s. Biochem. J. 2005, 387, 165–173. [Google Scholar] [CrossRef] [PubMed]
- El Asmar, R.; Panigrahi, P.; Bamford, P.; Berti, I.; Not, T.; Coppa, G.V.; Catassi, C.; Fasano, A. Host-dependent zonulin secretion causes the impairment of the small intestine barrier function after bacterial exposure. Gastroenterology 2002, 123, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Lammers, K.M.; Lu, R.; Brownley, J.; Lu, B.; Gerard, C.; Thomas, K.; Rallabhandi, P.; Shea-Donohue, T.; Tamiz, A.; Alkan, S.; et al. Gliadin Induces an Increase in Intestinal Permeability and Zonulin Release by Binding to the Chemokine Receptor CXCR3. Gastroenterology 2008, 135, 194–204.e3. [Google Scholar] [CrossRef] [PubMed]
- Groom, J.R.; Luster, A.D. CXCR3 ligands: Redundant, collaborative and antagonistic functions. Immunol. Cell Biol. 2011, 89, 207–215. [Google Scholar] [CrossRef]
- Van Raemdonck, K.; Van den Steen, P.E.; Liekens, S.; Van Damme, J.; Struyf, S. CXCR3 ligands in disease and therapy. Cytokine Growth Factor Rev. 2015, 26, 311–327. [Google Scholar] [CrossRef]
- Singh, U.P.; Venkataraman, C.; Singh, R.; Lillard, J.W. CXCR3 axis: Role in inflammatory bowel disease and its therapeutic implication. Endocr. Metab. Immune Disord.-Drug Targets (Former. Curr. Drug Targets-Immune Endocr. Metab. Disord.) 2007, 7, 111–123. [Google Scholar] [CrossRef]
- Haghbin, M.; Rostami-Nejad, M.; Forouzesh, F.; Sadeghi, A.; Rostami, K.; Aghamohammadi, E.; Asadzadeh-Aghdaei, H.; Masotti, A.; Zali, M.R. The role of CXCR3 and its ligands CXCL10 and CXCL11 in the pathogenesis of celiac disease. Medicine 2019, 98, e15949. [Google Scholar] [CrossRef]
- Heickman, L.K.W.; DeBoer, M.D.; Fasano, A. Zonulin as a potential putative biomarker of risk for shared type 1 diabetes and celiac disease autoimmunity. Diabetes/Metabolism Res. Rev. 2020, 36, e3309. [Google Scholar] [CrossRef]
- Thomas, K.E.; Sapone, A.; Fasano, A.; Vogel, S.N. Gliadin Stimulation of Murine Macrophage Inflammatory Gene Expression and Intestinal Permeability Are MyD88-Dependent: Role of the Innate Immune Response in Celiac Disease. J. Immunol. 2006, 176, 2512–2521. [Google Scholar] [CrossRef]
- Saikh, K.U. MyD88 and beyond: A perspective on MyD88-targeted therapeutic approach for modulation of host immunity. Immunol. Res. 2021, 69, 117–128. [Google Scholar] [CrossRef]
- Uhde, M.; Ajamian, M.; Caio, G.; De Giorgio, R.; Indart, A.; Green, P.H.; Verna, E.C.; Volta, U.; Alaedini, A. Intestinal cell damage and systemic immune activation in individuals reporting sensitivity to wheat in the absence of coeliac disease. Gut 2016, 65, 1930–1937. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.; Linglong, P.; Weixia, D.; Hong, W. Protective effects of bifidobacterium on intestinal barrier function in LPS-induced enterocyte barrier injury of caco-2 monolayers and in a rat NEC model. PLoS ONE 2016, 11, e0161635. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, S.; Jiang, Z.; Wang, X.; Liu, Y. Chemokine receptor CXCR3 in turbot (Scophthalmus maximus): Cloning, characterization and its responses to lipopolysaccharide. Fish Physiol. Biochem. 2016, 42, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, Z.; Zhu, K.; Bao, R.; Meng, Y.; Bian, J.; Wan, X.; Yang, T. Effects of CXCL4/CXCR3 on the lipopolysaccharide-induced injury in human umbilical vein endothelial cells. J. Cell Physiol. 2019, 234, 22378–22385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Deng, Y.; Zhang, Y.; Ba, T.; Niu, S.; Chen, Y.; Gao, Y.; Dai, H. CXCR3 Inhibition Blocks the NF-κB Signaling Pathway by Elevating Autophagy to Ameliorate Lipopolysaccharide-Induced Intestinal Dysfunction in Mice. Cells 2023, 12, 182. [Google Scholar] [CrossRef]
- Lauxmann, M.A.; Vazquez, D.S.; Schilbert, H.M.; Neubauer, P.R.; Lammers, K.M.; Dodero, V.I. From celiac disease to coccidia infection and vice-versa: The polyQ peptide CXCR3-interaction axis. Bioessays 2021, 43, e2100101. [Google Scholar] [CrossRef]
- Quaye, I.K. Haptoglobin, inflammation and disease. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 735–742. [Google Scholar] [CrossRef]
- El-Doueik, H.; El-Doueik, A. Assessment of the Correlation between Intestinal Permeability, Inflammation and Dysbiosis in Patients with Inflammatory Conditions within a Clinical Setting. 2019; Preprint. [Google Scholar]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Alessandro, R.; Luchetti, M.M.; Milling, S.; Saieva, L.; Cypers, H.; Stampone, T.; Di Benedetto, P.; et al. Dysbiosis and zonulin upregulation alter gut epithelial and vascular barriers in patients with ankylosing spondylitis. Ann. Rheum. Dis. 2017, 76, 1123–1132. [Google Scholar] [CrossRef]
- Gargari, G.; Mantegazza, G.; Taverniti, V.; Del Bo, C.; Rsquo, C.; Bernardi, S.; Andres-Lacueva, C.; González-Domínguez, R.; Kroon, P.A.; Winterbone, M.S.; et al. Bacterial DNAemia is associated with serum zonulin levels in older subjects. Sci. Rep. 2021, 11, 11054. [Google Scholar] [CrossRef]
- Jendraszak, M.; Gałęcka, M.; Kotwicka, M.; Schwiertz, A.; Regdos, A.; Pazgrat-Patan, M.; Andrusiewicz, M.J.B. Impact of Biometric Patient Data, Probiotic Supplementation, and Selected Gut Microorganisms on Calprotectin, Zonulin, and sIgA Concentrations in the Stool of Adults Aged 18–74 Years. Biomolecules 2022, 12, 1781. [Google Scholar] [CrossRef]
- Żak-Gołąb, A.; Kocełak, P.; Aptekorz, M.; Zientara, M.; Juszczyk, Ł.; Martirosian, G.; Chudek, J.; Olszanecka-Glinianowicz, M. Gut Microbiota, Microinflammation, Metabolic Profile, and Zonulin Concentration in Obese and Normal Weight Subjects. Int. J. Endocrinol. 2013, 2013, 674106. [Google Scholar] [CrossRef] [PubMed]
- Cayres, L.C.d.F.; de Salis, L.V.V.; Rodrigues, G.S.P.; Lengert, A.v.H.; Biondi, A.P.C.; Sargentini, L.D.B.; Brisotti, J.L.; Gomes, E.; de Oliveira, G.L.V.J.F.i.I. Detection of alterations in the gut microbiota and intestinal permeability in patients with Hashimoto thyroiditis. Front. Immunol. 2021, 12, 579140. [Google Scholar] [CrossRef]
- Sánchez-Alcoholado, L.; Ordóñez, R.; Otero, A.; Plaza-Andrade, I.; Laborda-Illanes, A.; Medina, J.A.; Ramos-Molina, B.; Gómez-Millán, J.; Queipo-Ortuño, M.I. Gut Microbiota-Mediated Inflammation and Gut Permeability in Patients with Obesity and Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 6782. [Google Scholar] [CrossRef] [PubMed]
- Mörkl, S.; Lackner, S.; Meinitzer, A.; Mangge, H.; Lehofer, M.; Halwachs, B.; Gorkiewicz, G.; Kashofer, K.; Painold, A.; Holl, A.K.; et al. Gut microbiota, dietary intakes and intestinal permeability reflected by serum zonulin in women. Eur. J. Nutr. 2018, 57, 2985–2997. [Google Scholar] [CrossRef] [PubMed]
- Vorobjova, T.; Raikkerus, H.; Kadaja, L.; Talja, I.; Uibo, O.; Heilman, K.; Uibo, R. Circulating Zonulin Correlates with Density of Enteroviruses and Tolerogenic Dendritic Cells in the Small Bowel Mucosa of Celiac Disease Patients. Dig. Dis. Sci. 2017, 62, 358–371. [Google Scholar] [CrossRef]
- Cangemi, R.; Pignatelli, P.; Carnevale, R.; Bartimoccia, S.; Nocella, C.; Falcone, M.; Taliani, G.; Violi, F.; Battaglia, S.; Bertazzoni, G.; et al. Low-grade endotoxemia, gut permeability and platelet activation in community-acquired pneumonia. J. Infect. 2016, 73, 107–114. [Google Scholar] [CrossRef]
- Carrera-Bastos, P.; Picazo, Ó.; Fontes-Villalba, M.; Pareja-Galeano, H.; Lindeberg, S.; Martínez-Selles, M.; Lucia, A.; Emanuele, E. Serum Zonulin and Endotoxin Levels in Exceptional Longevity versus Precocious Myocardial Infarction. Aging Dis. 2018, 9, 317–321. [Google Scholar] [CrossRef]
- Jayashree, B.; Bibin, Y.S.; Prabhu, D.; Shanthirani, C.S.; Gokulakrishnan, K.; Lakshmi, B.S.; Mohan, V.; Balasubramanyam, M. Increased circulatory levels of lipopolysaccharide (LPS) and zonulin signify novel biomarkers of proinflammation in patients with type 2 diabetes. Mol. Cell Biochem. 2014, 388, 203–210. [Google Scholar] [CrossRef]
- Zheng, D.; Liao, H.; Chen, S.; Liu, X.; Mao, C.; Zhang, C.; Meng, M.; Wang, Z.; Wang, Y.; Jiang, Q.; et al. Elevated Levels of Circulating Biomarkers Related to Leaky Gut Syndrome and Bacterial Translocation Are Associated With Graves’ Disease. Front. Endocrinol. 2021, 12, 796212. [Google Scholar] [CrossRef]
- Niewiem, M.; Grzybowska-Chlebowczyk, U.J.N. Assessment of Selected Intestinal Permeability Markers in Children with Food Allergy Depending on the Type and Severity of Clinical Symptoms. Nutrients 2022, 14, 4385. [Google Scholar] [CrossRef]
- Arslan, S.; Altunisik, N.; Turkmen, D.; Uremis, M.M.; Sener, S.; Turkoz, Y. Evaluation of plasma zonulin level and its relationship with inflammatory cytokines in patients with vitiligo. J. Cosmet. Dermatol. 2022, 22, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, J.; Teng, T.; Yin, B.; He, Y.; Jiang, Y.; Liu, X.; Yu, Y.; Li, X.; Zhou, X. Biomarkers of intestinal permeability and blood-brain barrier permeability in adolescents with major depressive disorder. J. Affect. Disord. 2023, 323, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Klaus, D.A.; Motal, M.C.; Burger-Klepp, U.; Marschalek, C.; Schmidt, E.M.; Lebherz-Eichinger, D.; Krenn, C.G.; Roth, G.A. Increased plasma zonulin in patients with sepsis. Biochem. Medica 2013, 23, 107–111. [Google Scholar] [CrossRef]
- Kaczmarczyk, M.; Löber, U.; Adamek, K.; Węgrzyn, D.; Skonieczna-Żydecka, K.; Malinowski, D.; Łoniewski, I.; Markó, L.; Ulas, T.; Forslund, S.K.; et al. The gut microbiota is associated with the small intestinal paracellular permeability and the development of the immune system in healthy children during the first two years of life. J. Transl. Med. 2021, 19, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-H.; Huang, M.-J.; Zhang, X.-W.; Wang, L.; Huang, N.-Q.; Peng, H.; Lan, P.; Peng, J.S.; Yang, Z.; Xia, Y.; et al. The effects of perioperative probiotic treatment on serum zonulin concentration and subsequent postoperative infectious complications after colorectal cancer surgery: A double-center and double-blind randomized clinical trial. Am. J. Clin. Nutr. 2013, 97, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, C.; Huang, M.; Tong, C.; Zhang, X.; Wang, L.; Peng, H.; Lan, P.; Zhang, P.; Huang, N.; et al. Positive regulatory effects of perioperative probiotic treatment on postoperative liver complications after colorectal liver metastases surgery: A double-center and double-blind randomized clinical trial. BMC Gastroenterol. 2015, 15, 34. [Google Scholar] [CrossRef] [PubMed]
- Stenman, L.K.; Lehtinen, M.J.; Meland, N.; Christensen, J.E.; Yeung, N.; Saarinen, M.T.; Courtney, M.; Burcelin, R.; Lähdeaho, M.-L.; Linros, J.J.E. Probiotic with or without fiber controls body fat mass, associated with serum zonulin, in overweight and obese adults—Randomized controlled trial. EBioMedicine 2016, 13, 190–200. [Google Scholar] [CrossRef]
- Kantah, M.; Catanzaro, R.; Kumar, M.; Jeong, W.; Marcellino, M.J.J.G.D.S. Beneficial Gut Effect of a Symbiotic-Probiotic Regimen in Healthy Stressed Individuals: Effectiveness on Permeability, Microbiota and Detoxification Parameters. J. Gastrointest. Dig. Syst. 2018, 8, 560. [Google Scholar]
- Janczy, A.; Aleksandrowicz-Wrona, E.; Kochan, Z.; Małgorzewicz, S.J.A.B.P. Impact of diet and synbiotics on selected gut bacteria and intestinal permeability in individuals with excess body weight–a prospective, randomized study. Acta Biochim. Pol. 2020, 67, 571–578. [Google Scholar] [CrossRef]
- Lamprecht, M.; Bogner, S.; Schippinger, G.; Steinbauer, K.; Fankhauser, F.; Hallstroem, S.; Schuetz, B.; Greilberger, J.F. Probiotic supplementation affects markers of intestinal barrier, oxidation, and inflammation in trained men; a randomized, double-blinded, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2012, 9, 45. [Google Scholar] [CrossRef]
- Wilms, E.; Gerritsen, J.; Smidt, H.; Besseling-van der Vaart, I.; Rijkers, G.T.; Garcia Fuentes, A.; Masclee, A.A.; Troost, F.J.J.P.O. Effects of supplementation of the synbiotic Ecologic® 825/FOS P6 on intestinal barrier function in healthy humans: A randomized controlled trial. PLoS ONE 2016, 11, e0167775. [Google Scholar] [CrossRef] [PubMed]
- Cakir, M.; Isbilen, A.A.; Eyupoglu, I.; Sag, E.; Orem, A.; Sen, T.M.; Kaklikkaya, N.; Kaya, G. Effects of long-term synbiotic supplementation in addition to lifestyle changes in children with obesity-related non-alcoholic fatty liver disease. Turk. J. Gastroenterol. 2017, 28, 377–383. [Google Scholar] [CrossRef] [PubMed]
- de Roos, N.M.; van Hemert, S.; Rovers, J.M.P.; Smits, M.G.; Witteman, B.J.M. The effects of a multispecies probiotic on migraine and markers of intestinal permeability–results of a randomized placebo-controlled study. Eur. J. Clin. Nutr. 2017, 71, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Wegh, C.A.M.; De Roos, N.M.; Hovenier, R.; Meijerink, J.; Der Vaart, I.B.-V.; Van Hemert, S.; Witteman, B.J.M. Intestinal Permeability Measured by Urinary Sucrose Excretion Correlates with Serum Zonulin and Faecal Calprotectin Concentrations in UC Patients in Remission. J. Nutr. Metab. 2019, 2019, 2472754. [Google Scholar] [CrossRef] [PubMed]
- Townsend, J.R.; Bender, D.; Vantrease, W.C.; Sapp, P.A.; Toy, A.M.; Woods, C.A.; Johnson, K.D. Effects of Probiotic (Bacillus subtilis DE111) Supplementation on Immune Function, Hormonal Status, and Physical Performance in Division I Baseball Players. Sports 2018, 6, 70. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.; Sanz, Y. Bifidobacteria inhibit the inflammatory response induced by gliadins in intestinal epithelial cells via modifications of toxic peptide generation during digestion. J. Cell Biochem. 2010, 109, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. Sourdough lactobacilli and celiac disease. Food Microbiol. 2007, 24, 187–196. [Google Scholar] [CrossRef]
- De Angelis, M.; Rizzello, C.G.; Fasano, A.; Clemente, M.G.; De Simone, C.; Silano, M.; De Vincenzi, M.; Losito, I.; Gobbetti, M. VSL#3 probiotic preparation has the capacity to hydrolyze gliadin polypeptides responsible for Celiac Sprue probiotics and gluten intolerance. Biochim. et Biophys. Acta (BBA) Mol. Basis Dis. 2006, 1762, 80–93. [Google Scholar] [CrossRef]
- Martorell, P.; Alvarez, B.; Llopis, S.; Navarro, V.; Ortiz, P.; Gonzalez, N.; Balaguer, F.; Rojas, A.; Chenoll, E.; Ramón, D. Heat-treated Bifidobacterium longum CECT-7347: A whole-cell postbiotic with antioxidant, anti-inflammatory, and gut-barrier protection properties. Antioxidants 2021, 10, 536. [Google Scholar] [CrossRef]
- Orlando, A.; Linsalata, M.; Notarnicola, M.; Tutino, V.; Russo, F. Lactobacillus GG restoration of the gliadin induced epithelial barrier disruption: The role of cellular polyamines. BMC Microbiol. 2014, 14, 19. [Google Scholar] [CrossRef]
- Xiong, W.; Huang, J.; Li, X.; Zhang, Z.; Jin, M.; Wang, J.; Xu, Y.; Wang, Z. Icariin and its phosphorylated derivatives alleviate intestinal epithelial barrier disruption caused by enterotoxigenic Escherichia coli through modulate p38 MAPK in vivo and in vitro. FASEB J. 2020, 34, 1783–1801. [Google Scholar] [CrossRef] [PubMed]
- Jian, C.; Kanerva, S.; Qadri, S.; Yki-Järvinen, H.; Salonen, A. In vitro Effects of Bacterial Exposure on Secretion of Zonulin Family Peptides and Their Detection in Human Tissue Samples. Front. Microbiol. 2022, 13, 848128. [Google Scholar] [CrossRef] [PubMed]
- Strauman, M.C.; Harper, J.M.; Harrington, S.M.; Boll, E.J.; Nataro, J.P. Enteroaggregative Escherichia coli Disrupts Epithelial Cell Tight Junctions. Infect. Immun. 2010, 78, 4958–4964. [Google Scholar] [CrossRef] [PubMed]
- Mukiza, C.N.; Dubreuil, J.D. Escherichia coli Heat-Stable Toxin b Impairs Intestinal Epithelial Barrier Function by Altering Tight Junction Proteins. Infect. Immun. 2013, 81, 2819–2827. [Google Scholar] [CrossRef]
- Brown, E.M.; Wlodarska, M.; Willing, B.P.; Vonaesch, P.; Han, J.; Reynolds, L.A.; Arrieta, M.-C.; Uhrig, M.; Scholz, R.; Partida, O.; et al. Diet and specific microbial exposure trigger features of environmental enteropathy in a novel murine model. Nat. Commun. 2015, 6, 7806. [Google Scholar] [CrossRef]
- Li, C.; Gao, M.; Zhang, W.; Chen, C.; Zhou, F.; Hu, Z.; Zeng, C. Zonulin Regulates Intestinal Permeability and Facilitates Enteric Bacteria Permeation in Coronary Artery Disease. Sci. Rep. 2016, 6, 29142. [Google Scholar] [CrossRef]
- Doguer, C.; Akalan, H.; Demirok, N.T.; Erdal, B.; Mete, R.; Bilgen, T. Protective effects of Acetobacter ghanensis against gliadin toxicity in intestinal epithelial cells with immunoregulatory and gluten-digestive properties. Eur. J. Nutr. 2022, 62, 605–614. [Google Scholar] [CrossRef]
- Nakajima, M.; Arimatsu, K.; Kato, T.; Matsuda, Y.; Minagawa, T.; Takahashi, N.; Ohno, H.; Yamazaki, K. Oral Administration of P. gingivalis Induces Dysbiosis of Gut Microbiota and Impaired Barrier Function Leading to Dissemination of Enterobacteria to the Liver. PLoS ONE 2015, 10, e0134234. [Google Scholar] [CrossRef]
- Yoseph, B.P.; Klingensmith, N.J.; Liang, Z.; Breed, E.; Burd, E.M.; Mittal, R.; Dominguez, J.A.; Petrie, B.; Ford, M.L.; Coopersmith, C.M. Mechanisms of Intestinal Barrier Dysfunction in Sepsis. Shock 2016, 46, 52–59. [Google Scholar] [CrossRef]
- Liu, H.; Hong, X.L.; Sun, T.T.; Huang, X.W.; Wang, J.L.; Xiong, H. Fusobacterium nucleatum exacerbates colitis by damaging epithelial barriers and inducing aberrant inflammation. J. Dig. Dis. 2020, 21, 385–398. [Google Scholar] [CrossRef]
- Deng, J.; Azzouz, D.F.; Ferstler, N.; Silverman, G.J.J.b. Sex-dependent Lupus Ruminococcus blautia gnavus strain induction of zonulin-mediated intestinal permeability and autoimmunity. Front. Immunol. 2022, 13, 897971. [Google Scholar]
- Nusrat, A.; von Eichel-Streiber, C.; Turner, J.R.; Verkade, P.; Madara, J.L.; Parkos, C.A. Clostridium difficile Toxins Disrupt Epithelial Barrier Function by Altering Membrane Microdomain Localization of Tight Junction Proteins. Infect. Immun. 2001, 69, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liang, R.; Zhang, W.; Tian, K.; Li, J.; Chen, X.; Yu, T.; Chen, Q.; Tao, Y. Faecalibacterium prausnitzii-derived microbial anti-inflammatory molecule regulates intestinal integrity in diabetes mellitus mice via modulating tight junction protein expression. J. Diabetes 2020, 12, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Li, X.; Wang, E.; He, Y.; Yin, B.; Fang, D.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. A cellular model for screening of lactobacilli that can enhance tight junctions. RSC Adv. 2016, 6, 111812–111821. [Google Scholar] [CrossRef]
- Ewaschuk, J.B.; Diaz, H.; Meddings, L.; Diederichs, B.; Dmytrash, A.; Backer, J.; Looijer-van Langen, M.; Madsen, K.L. Secreted bioactive factors from Bifidobacterium infantis enhance epithelial cell barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1025–G1034. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Wang, S.; Nagpal, R.; Wang, B.; Jain, S.; Razazan, A.; Mishra, S.P.; Zhu, X.; Wang, Z.; Kavanagh, K.; et al. A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis. J. Clin. Investig. 2020, 5, e132055. [Google Scholar] [CrossRef]
- Giorgi, A.; Cerrone, R.; Capobianco, D.; Filardo, S.; Mancini, P.; Zanni, F.; Fanelli, S.; Mastromarino, P.; Mosca, L. A Probiotic Preparation Hydrolyzes Gliadin and Protects Intestinal Cells from the Toxicity of Pro-Inflammatory Peptides. Nutrients 2020, 12, 495. [Google Scholar] [CrossRef]
- Bandara, M.; MacNaughton, W.K. Protease-activated receptor-2 activation enhances epithelial wound healing via epidermal growth factor receptor. Tissue Barriers 2022, 10, 1968763. [Google Scholar] [CrossRef]
- van der Merwe, J.Q.; Hollenberg, M.D.; MacNaughton, W.K. EGF receptor transactivation and MAP kinase mediate proteinase-activated receptor-2-induced chloride secretion in intestinal epithelial cells. Am. J. Physiol. Liver Physiol. 2008, 294, G441–G451. [Google Scholar] [CrossRef]
- Heijink, I.H.; van Oosterhout, A.; Kapus, A. Epidermal growth factor receptor signalling contributes to house dust mite-induced epithelial barrier dysfunction. Eur. Respir. J. 2010, 36, 1016–1026. [Google Scholar] [CrossRef]
- Yarden, Y.; Shilo, B.-Z. SnapShot: EGFR Signaling Pathway. Cell 2007, 131, 1018.e1–1018.e2. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Intestinal Permeability and Its Regulation by Zonulin: Diagnostic and Therapeutic Implications. Clin. Gastroenterol. Hepatol. 2012, 10, 1096–1100. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Fu, L. Mechanisms of resistance to EGFR tyrosine kinase inhibitors. Acta Pharm. Sin. B 2015, 5, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Goldblum, S.E.; Rai, U.; Tripathi, A.; Thakar, M.; De Leo, L.; Di Toro, N.; Not, T.; Ramachandran, R.; Puche, A.C.; Hollenberg, M.D. The active Zot domain (aa 288–293) increases ZO-1 and myosin 1C serine/threonine phosphorylation, alters interaction between ZO-1 and its binding partners, and induces tight junction disassembly through proteinase activated receptor 2 activation. FASEB J. 2011, 25, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Larsson, C. Protein kinase C and the regulation of the actin cytoskeleton. Cell Signal. 2006, 18, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhang, X.-F.; Van Goor, D.; Dunn, A.P.; Hyland, C.; Medeiros, N.; Forscher, P. Protein kinase C activation decreases peripheral actin network density and increases central nonmuscle myosin II contractility in neuronal growth cones. Mol. Biol. Cell 2013, 24, 3097–3114. [Google Scholar] [CrossRef] [PubMed]
- Veres-Székely, A.; Bernáth, M.; Pap, D.; Rokonay, R.; Szebeni, B.; Takács, I.M.; Lippai, R.; Cseh, Á.; Szabó, A.J.; Vannay, Á. PARK7 Diminishes Oxidative Stress-Induced Mucosal Damage in Celiac Disease. Oxid. Med. Cell Longev. 2020, 2020, 4787202. [Google Scholar] [CrossRef]
- Clemente, M.G.; De Virgiliis, S.; Kang, J.S.; Macatagney, R.; Musu, M.P.; Di Pierro, M.R.; Drago, S.; Congia, M.; Fasano, A. Early effects of gliadin on enterocyte intracellular signalling involved in intestinal barrier function. Gut 2003, 52, 218–223. [Google Scholar] [CrossRef]
- Enjoji, S.; Ohama, T.; Sato, K. Regulation of Epithelial Cell Tight Junctions by Protease-Activated Receptor 2. J. Veter Med. Sci. 2014, 76, 1225–1229. [Google Scholar] [CrossRef]
- Wang, J.; Kang, X.; Zhi-Qun, H.; Shen, L.; Qing, L.; Meng-Yue, L.; Li-Ping, L.; Jun-Hao, Y.; Mei, H.; Ye, J. Protease-activated receptor-2 decreased zonula occlidens-1 and claudin-1 expression and induced epithelial barrier dysfunction in allergic rhinitis. Am. J. Rhinol. Allergy 2021, 35, 26–35. [Google Scholar] [CrossRef]
- Vesey, D.; Suen, J.; Seow, V.; Lohman, R.-J.; Liu, L.; Gobe, G.C.; Johnson, D.W.; Fairlie, D. PAR2-induced inflammatory responses in human kidney tubular epithelial cells. Am. J. Physiol. Physiol. 2013, 304, F737–F750. [Google Scholar] [CrossRef] [PubMed]
- Flores-Benitez, D.; Rincon-Heredia, R.; Razgado, L.F.; Larre, I.; Cereijido, M.; Contreras, R.G. Control of tight junctional sealing: Roles of epidermal growth factor and prostaglandin E2. Am. J. Physiol. Physiol. 2009, 297, C611–C620. [Google Scholar] [CrossRef] [PubMed]
- Basuroy, S.; Seth, A.; Elias, B.; Naren, A.P.; Rao, R. MAPK interacts with occludin and mediates EGF-induced prevention of tight junction disruption by hydrogen peroxide. Biochem. J. 2006, 393, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Jauregi-Miguel, A. The tight junction and the epithelial barrier in coeliac disease. Int. Rev. Cell Mol. Biol. 2020, 358, 105–132. [Google Scholar] [CrossRef]
- Barbaro, M.R.; Cremon, C.; Morselli-Labate, A.M.; Di Sabatino, A.; Giuffrida, P.; Corazza, G.R.; Di Stefano, M.; Caio, G.; Latella, G.; Ciacci, C.; et al. Serum zonulin and its diagnostic performance in non-coeliac gluten sensitivity. Gut 2021, 69, 1966–1974. [Google Scholar] [CrossRef]
- Singh, P.; Silvester, J.; Chen, X.; Xu, H.; Sawhney, V.; Rangan, V.; Iturrino, J.; Nee, J.; Duerksen, D.R.; Lembo, A. Serum zonulin is elevated in IBS and correlates with stool frequency in IBS-D. United Eur. Gastroenterol. J. 2019, 7, 709–715. [Google Scholar] [CrossRef]
- Rezazadegan, M.; Soheilipour, M.; Tarrahi, M.J.; Amani, R. Correlation Between Zinc Nutritional Status with Serum Zonulin and Gastrointestinal Symptoms in Diarrhea-Predominant Irritable Bowel Syndrome: A Case–Control Study. Dig. Dis. Sci. 2022, 67, 3632–3638. [Google Scholar] [CrossRef]
- Caviglia, G.P.; Dughera, F.; Ribaldone, D.G.; Rosso, C.; Abate, M.L.; Pellicano, R.; Bresso, F.; Smedile, A.; Saracco, G.M.; Astegiano, M. Serum zonulin in patients with inflammatory bowel disease: A pilot study. Minerva Med. 2019, 110, 95–100. [Google Scholar] [CrossRef]
- Tarko, A.; Suchojad, A.; Michalec, M.; Majcherczyk, M.; Brzozowska, A.; Maruniak-Chudek, I. Zonulin: A Potential Marker of Intestine Injury in Newborns. Dis. Markers 2017, 2017, 2413437. [Google Scholar] [CrossRef]
- Łoniewska, B.; Węgrzyn, D.; Adamek, K.; Kaczmarczyk, M.; Skonieczna-Żydecka, K.; Adler, G.; Jankowska, A.; Uzar, I.; Kordek, A.; Celewicz, M.; et al. The Influence of Maternal-Foetal Parameters on Concentrations of Zonulin and Calprotectin in the Blood and Stool of Healthy Newborns during the First Seven Days of Life. An Observational Prospective Cohort Study. J. Clin. Med. 2019, 8, 473. [Google Scholar] [CrossRef]
- El Wakeel, M.A.; El-Kassas, G.M.; Hashem, S.A.; Hasanin, H.M.; Ali, W.H.; Elkhatib, A.A.; Sibaii, H.; Fadl, N.N. Serum Biomarkers of Environmental Enteric Dysfunction and Growth Perspective in Egyptian Children. Open Access Maced. J. Med. Sci. 2021, 9, 1625–1632. [Google Scholar] [CrossRef]
- Mwape, I.; Bosomprah, S.; Mwaba, J.; Mwila-Kazimbaya, K.; Laban, N.M.; Chisenga, C.C.; Sijumbila, G.; Simuyandi, M.; Chilengi, R. Immunogenicity of rotavirus vaccine (RotarixTM) in infants with environmental enteric dysfunction. PLoS ONE 2017, 12, e0187761. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, M.-M.; Niu, Y.; Zhang, X.; Yin, J.-B.; Zhao, C.-J.; Wang, R.-T. Serum Zonulin in HBV-Associated Chronic Hepatitis, Liver Cirrhosis, and Hepatocellular Carcinoma. Dis. Markers 2019, 2019, 5945721. [Google Scholar] [CrossRef] [PubMed]
- A Voulgaris, T.; Karagiannakis, D.; Hadziyannis, E.; Manolakopoulos, S.; Karamanolis, G.P.; Papatheodoridis, G.; Vlachogiannakos, J. Serum zonulin levels in patients with liver cirrhosis: Prognostic implications. World J. Hepatol. 2021, 13, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- De Munck, T.J.I.; Xu, P.; Verwijs, H.J.A.; Masclee, A.A.M.; Jonkers, D.; Verbeek, J.; Koek, G.H. Intestinal permeability in human nonalcoholic fatty liver disease: A systematic review and meta-analysis. Liver Int. 2020, 40, 2906–2916. [Google Scholar] [CrossRef]
- Sapone, A.; de Magistris, L.; Pietzak, M.; Clemente, M.G.; Tripathi, A.; Cucca, F.; Lampis, R.; Kryszak, D.; Cartenì, M.; Generoso, M.; et al. Zonulin Upregulation Is Associated With Increased Gut Permeability in Subjects With Type 1 Diabetes and Their Relatives. Diabetes 2006, 55, 1443–1449. [Google Scholar] [CrossRef]
- Watts, T.; Berti, I.; Sapone, A.; Gerarduzzi, T.; Not, T.; Zielke, R.; Fasano, A. Role of the intestinal tight junction modulator zonulin in the pathogenesis of type I diabetes in BB diabetic-prone rats. Proc. Natl. Acad. Sci. USA 2005, 102, 2916–2921. [Google Scholar] [CrossRef]
- Mønsted, M.; Falck, N.D.; Pedersen, K.; Buschard, K.; Holm, L.J.; Haupt-Jorgensen,, M. Intestinal permeability in type 1 diabetes: An updated comprehensive overview. J. Autoimmun. 2021, 122, 102674. [Google Scholar] [CrossRef]
- Yuan, J.-H.; Xie, Q.-S.; Chen, G.-C.; Huang, C.-L.; Yu, T.; Chen, Q.-K.; Li, J.-Y. Impaired intestinal barrier function in type 2 diabetic patients measured by serum LPS, Zonulin, and IFABP. J. Diabetes Complicat. 2021, 35, 107766. [Google Scholar] [CrossRef]
- Olivieri, F.; Maguolo, A.; Corradi, M.; Zusi, C.; Huber, V.; Fornari, E.; Morandi, A.; Maffeis, C. Serum zonulin as an index of glucose dysregulation in children and adolescents with overweight and obesity. Pediatr. Obes. 2022, 17, e12946. [Google Scholar] [CrossRef]
- Mokkala, K.; Pellonperä, O.; Röytiö, H.; Pussinen, P.; Rönnemaa, T.; Laitinen, K. Increased intestinal permeability, measured by serum zonulin, is associated with metabolic risk markers in overweight pregnant women. Metabolism 2017, 69, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Sabater, M.; Ortega, F.; Ricart, W.; Fernández-Real, J.M. Circulating Zonulin, a Marker of Intestinal Permeability, Is Increased in Association with Obesity-Associated Insulin Resistance. PLoS ONE 2012, 7, e37160. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, B.; Orho-Melander, M.; Nilsson, P.M. Higher Levels of Serum Zonulin May Rather Be Associated with Increased Risk of Obesity and Hyperlipidemia, Than with Gastrointestinal Symptoms or Disease Manifestations. Int. J. Mol. Sci. 2017, 18, 582. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, L.; Yue, F.; Zheng, Y.; Russell, R. Serum zonulin is elevated in women with polycystic ovary syndrome and correlates with insulin resistance and severity of anovulation. Eur. J. Endocrinol. 2015, 172, 29–36. [Google Scholar] [CrossRef]
- Parker, J.; O’Brien, C.; Hawrelak, J. A narrative review of the role of gastrointestinal dysbiosis in the pathogenesis of polycystic ovary syndrome. Obstet. Gynecol. Sci. 2022, 65, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Mokkala, K.; Tertti, K.; Rönnemaa, T.; Vahlberg, T.; Laitinen, K. Evaluation of serum zonulin for use as an early predictor for gestational diabetes. Nutr. Diabetes 2017, 7, e253. [Google Scholar] [CrossRef]
- Güvey, H.; Çelik, S.; Çalışkan, C.S.; Yılmaz, Z.; Yılmaz, M.; Erten, Ö.; Tinelli, A. How Do Serum Zonulin Levels Change in Gestational Diabetes Mellitus, Pregnancy Cholestasis, and the Coexistence of Both Diseases? Int. J. Environ. Res. Public Health 2021, 18, 12555. [Google Scholar] [CrossRef]
- Demir, E.; Ozkan, H.; Seckin, K.D.; Sahtiyancı, B.; Demir, B.; Tabak, O.; Kumbasar, A.; Uzun, H. Plasma Zonulin Levels as a Non-Invasive Biomarker of Intestinal Permeability in Women with Gestational Diabetes Mellitus. Biomolecules 2019, 9, 24. [Google Scholar] [CrossRef]
- Oral, S.; Celik, S.; Akpak, Y.K.; Golbasi, H.; Bayraktar, B.; Unver, G.; Sahin, S.; Yurtcu, N.; Caliskan, C.S. Prediction of gestational diabetes mellitus and perinatal outcomes by plasma zonulin levels. Arch. Gynecol. Obstet. 2022, 1–8. [Google Scholar] [CrossRef]
- Daneshvar, M.; Yadegari, A.; Ribaldone, D.G.; Hasanzadeh, M.; Djafarian, K. Zonulin levels in complicated pregnancy: A systematic review and meta-analysis. J. Obstet. Gynaecol. 2022, 42, 2621–2628. [Google Scholar] [CrossRef]
- Yilmaz, Z.; Oral, S.; Yurtcu, N.; Akpak, Y.K.; Celik, S.; Caliskan, C. Predictive and Prognostic Value of Plasma Zonulin for Gestational Diabetes Mellitus in Women at 24–28 Weeks of Gestation. Z. Geburtshilfe Neonatol. 2022, 226, 358–390. [Google Scholar] [CrossRef] [PubMed]
- Tajik, N.; Frech, M.; Schulz, O.; Schälter, F.; Lucas, S.; Azizov, V.; Dürholz, K.; Steffen, F.; Omata, Y.; Rings, A.; et al. Targeting zonulin and intestinal epithelial barrier function to prevent onset of arthritis. Nat. Commun. 2020, 11, 1995. [Google Scholar] [CrossRef] [PubMed]
- Chmielińska, M.; Olesińska, M.; Romanowska-Próchnicka, K.; Szukiewicz, D. Haptoglobin and Its Related Protein, Zonulin—What Is Their Role in Spondyloarthropathy? J. Clin. Med. 2021, 10, 1131. [Google Scholar] [CrossRef] [PubMed]
- Audo, R.; Sanchez, P.; Rivière, B.; Mielle, J.; Tan, J.; Lukas, C.; Macia, L.; Morel, J.; Daien, C.I. Rheumatoid arthritis is associated with increased gut permeability and bacterial translocation that are reversed by inflammation control. Rheumatology 2022, 62, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Karim, A.; Khan, J.; Qaisar, R. Plasma zonulin correlates with cardiac dysfunction and poor physical performance in patients with chronic heart failure. Life Sci. 2022, 311, 121150. [Google Scholar] [CrossRef]
- Baioumy, S.A.; Elgendy, A.; Ibrahim, S.M.; Taha, S.I.; Fouad, S.H. Association between serum zonulin level and severity of house dust mite allergic asthma. Allergy Asthma Clin. Immunol. 2021, 17, 86. [Google Scholar] [CrossRef]
- Karim, A.; Muhammad, T.; Ustrana, S.; Qaisar, R. Intestinal permeability marker zonulin as a predictor of sarcopenia in chronic obstructive pulmonary disease. Respir. Med. 2021, 189, 106662. [Google Scholar] [CrossRef]
- Carpes, L.S.; Nicoletto, B.B.; Canani, L.H.; Rheinhemer, J.; Crispim, D.; Souza, G.C. Could serum zonulin be an intestinal permeability marker in diabetes kidney disease? PLoS ONE 2021, 16, e0253501. [Google Scholar] [CrossRef]
- Trachtman, H.; Gipson, D.S.; Lemley, K.V.; Troost, J.P.; Faul, C.; Morrison, D.J.; Vento, S.M.; Ahn, D.-H.; Goldberg, J.D. Plasma Zonulin Levels in Childhood Nephrotic Syndrome. Front. Pediatr. 2019, 7, 197. [Google Scholar] [CrossRef]
- Hasslacher, C.; Kulozik, F.; Platten, I.; Kraft, M.; Siegel, E. Serum zonulin as parameter of intestinal permeability in longstanding type 2 diabetes: Correlations with metabolism parameter and renal function. J. Diabetes, Metab. Disord. Control 2018, 5, 58–62. [Google Scholar] [CrossRef]
- Asbjornsdottir, B.; Snorradottir, H.; Andresdottir, E.; Fasano, A.; Lauth, B.; Gudmundsson, L.S.; Gottfredsson, M.; Halldorsson, T.I.; Birgisdottir, B.E. Zonulin-Dependent Intestinal Permeability in Children Diagnosed with Mental Disorders: A Systematic Review and Meta-Analysis. Nutrients 2020, 12, 1982. [Google Scholar] [CrossRef] [PubMed]
- Özyurt, G.; Öztürk, Y.; Appak, Y.; Arslan, F.D.; Baran, M.; Karakoyun, I.; Tufan, A.E.; Pekcanlar, A.A. Increased zonulin is associated with hyperactivity and social dysfunctions in children with attention deficit hyperactivity disorder. Compr. Psychiatry 2018, 87, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Esnafoglu, E.; Cırrık, S.; Ayyıldız, S.N.; Erdil, A.; Ertürk, E.Y.; Daglı, A.; Noyan, T. Increased serum zonulin levels as an intestinal permeability marker in autistic subjects. J. Pediatr. 2017, 188, 240–244. [Google Scholar] [CrossRef]
- Karagözlü, S.; Dalgıç, B.; Işeri, E. The Relationship of Severity of Autism with Gastrointestinal Symptoms and Serum Zonulin Levels in Autistic Children. J. Autism Dev. Disord. 2021, 52, 623–629. [Google Scholar] [CrossRef]
- Kılıç, F.; Işık, Ü.; Demirdaş, A.; Doğuç, D.K.; Bozkurt, M. Serum zonulin and claudin-5 levels in patients with bipolar disorder. J. Affect. Disord. 2020, 266, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Usta, A.; Kılıç, F.; Demirdaş, A.; Işık, Ü.; Doğuç, D.K.; Bozkurt, M. Serum zonulin and claudin-5 levels in patients with schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 2021, 271, 767–773. [Google Scholar] [CrossRef]
- Stevens, B.R.; Goel, R.; Seungbum, K.; Richards, E.M.; Holbert, R.C.; Pepine, C.J.; Raizada, M.K. Increased human intestinal barrier permeability plasma biomarkers zonulin and FABP2 correlated with plasma LPS and altered gut microbiome in anxiety or depression. Gut 2018, 67, 1555–1557. [Google Scholar] [CrossRef]
- Karim, A.; Iqbal, M.S.; Muhammad, T.; Ahmad, F.; Qaisar, R. Elevated plasma zonulin and CAF22 are correlated with sarcopenia and functional dependency at various stages of Alzheimer’s diseases. Neurosci. Res. 2022, 184, 47–53. [Google Scholar] [CrossRef]
- Dumitrescu, L.; Marta, D.; Dănău, A.; Lefter, A.; Tulbă, D.; Cozma, L.; Manole, E.; Gherghiceanu, M.; Ceafalan, L.C.; Popescu, B.O. Serum and Fecal Markers of Intestinal Inflammation and Intestinal Barrier Permeability Are Elevated in Parkinson’s Disease. Front. Neurosci. 2021, 15, 689723. [Google Scholar] [CrossRef]
- Camara-Lemarroy, C.R.; Silva, C.; Greenfield, J.; Liu, W.-Q.; Metz, L.M.; Yong, V.W. Biomarkers of intestinal barrier function in multiple sclerosis are associated with disease activity. Mult. Scler. J. 2020, 26, 1340–1350. [Google Scholar] [CrossRef]
- Galea, I. The blood–brain barrier in systemic infection and inflammation. Cell Mol. Immunol. 2021, 18, 2489–2501. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.T.; Ghosh, C.; Hossain, M.; Linfield, D.; Rezaee, F.; Janigro, D.; Marchi, N.; van Boxel-Dezaire, A.H. IFN-γ, IL-17A, or zonulin rapidly increase the permeability of the blood-brain and small intestinal epithelial barriers: Relevance for neuro-inflammatory diseases. Biochem. Biophys. Res. Commun. 2018, 507, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Greene, C.; Hanley, N.; Campbell, M. Claudin-5: Gatekeeper of neurological function. Fluids Barriers CNS 2019, 16, 3. [Google Scholar] [CrossRef]
- Kılıç, A.O.; Akın, F.; Yazar, A.; Metin Akcan, Ö.; Topcu, C.; Aydın, O. Zonulin and claudin-5 levels in multisystem inflammatory syndrome and SARS-CoV-2 infection in children. J. Paediatr. Child Health 2022, 58, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Lasek-Bal, A.; Kokot, A.; de Carrillo, D.G.; Student, S.; Pawletko, K.; Krzan, A.; Puz, P.; Bal, W.; Jędrzejowska-Szypułka, H. Plasma Levels of Occludin and Claudin-5 in Acute Stroke Are Correlated with the Type and Location of Stroke but Not with the Neurological State of Patients—Preliminary Data. Brain Sci. 2020, 10, 831. [Google Scholar] [CrossRef] [PubMed]
- Okuyucu, M.; Kehribar, D.Y.; Çapraz, M.; Çapraz, A.; Arslan, M.; Çelik, Z.B.; Usta, B.; Birinci, A.; Ozgen, M. The Relationship between COVID-19 Disease Severity and Zonulin Levels. Cureus 2022, 14, e28255. [Google Scholar] [CrossRef] [PubMed]
- Yonker, L.M.; Gilboa, T.; Ogata, A.F.; Senussi, Y.; Lazarovits, R.; Boribong, B.P.; Bartsch, Y.C.; Loiselle, M.; Rivas, M.N.; Porritt, R.A.; et al. Multisystem inflammatory syndrome in children is driven by zonulin-dependent loss of gut mucosal barrier. J. Clin. Investig. 2021, 131, e149633. [Google Scholar] [CrossRef]
- Hensley-McBain, T.R.; Manuzak, J.A. Zonulin as a biomarker and potential therapeutic target in multisystem inflammatory syndrome in children. J. Clin. Investig. 2021, 131, e151467. [Google Scholar] [CrossRef]
- Llorens, S.; Nava, E.; Muñoz-López, M.; Sánchez-Larsen, Á.; Segura, T. Neurological Symptoms of COVID-19: The Zonulin Hypothesis. Front. Immunol. 2021, 12, 665300. [Google Scholar] [CrossRef]
- Palomino-Kobayashi, L.A.; Ymaña, B.; Ruiz, J.; Mayanga-Herrera, A.; Ugarte-Gil, M.F.; Pons, M.J. Zonulin, a marker of gut permeability, is associated with mortality in a cohort of hospitalised peruvian COVID-19 patients. Front. Cell. Infect. Microbiol. 2022, 12, 1310. [Google Scholar]
- Bawah, A.; Yakubu, Y.; Nanga, S. The relationship between zonulin and liver function test in patients with human immune deficiency virus infection. J. Med. Lab. Sci. Technol. S. Afr. 2021, 3, 71–76. [Google Scholar]
- Pastor, L.; Langhorst, J.; Schröder, D.; Casellas, A.; Ruffer, A.; Carrillo, J.; Urrea, V.; Massora, S.; Mandomando, I.; Blanco, J.; et al. Different pattern of stool and plasma gastrointestinal damage biomarkers during primary and chronic HIV infection. PLoS ONE 2019, 14, e0218000. [Google Scholar] [CrossRef] [PubMed]
- A Koay, W.L.; Lindsey, J.C.; Uprety, P.; Bwakura-Dangarembizi, M.; Weinberg, A.; Levin, M.J.; Persaud, D. Intestinal Integrity Biomarkers in Early Antiretroviral-Treated Perinatally HIV-1–Infected Infants. J. Infect. Dis. 2018, 218, 1085–1089. [Google Scholar] [CrossRef]
- Akao, T.; Morita, A.; Onji, M.; Miyake, T.; Watanabe, R.; Uehara, T.; Kawasaki, K.; Miyaike, J.; Oomoto, M. Low serum levels of zonulin in patients with HCV-infected chronic liver diseases. Euroasian J. Hepato-Gastroenterol. 2018, 8, 112. [Google Scholar]
- Calgin, M.K.; Cetinkol, Y. Decreased levels of serum zonulin and copeptin in chronic Hepatitis-B patients. Pak. J. Med. Sci. 2019, 35, 847–851. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, C.R.; Tamaki, R.; Gamerdinger, M.; Lessmann, V.; Behl, C.; Kempski, O.S.; Luhmann, H.J. Inhibition of the myosin light chain kinase prevents hypoxia-induced blood-brain barrier disruption. J. Neurochem. 2007, 102, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Thompson, J.F.; Taheri, S.; Salayandia, V.M.; McAvoy, T.A.; Hill, J.W.; Yang, Y.; Estrada, E.Y.; A Rosenberg, G. Early Inhibition of MMP Activity in Ischemic Rat Brain Promotes Expression of Tight Junction Proteins and Angiogenesis During Recovery. J. Cereb. Blood Flow Metab. 2013, 33, 1104–1114. [Google Scholar] [CrossRef]
- Cui, J.; Chen, S.; Zhang, C.; Meng, F.; Wu, W.; Hu, R.; Hadass, O.; Lehmidi, T.; Blair, G.J.; Lee, M.; et al. Inhibition of MMP-9 by a selective gelatinase inhibitor protects neurovasculature from embolic focal cerebral ischemia. Mol. Neurodegener. 2012, 7, 21. [Google Scholar] [CrossRef]
- Candelario-Jalil, E.; Taheri, S.; Yang, Y.; Sood, R.; Grossetete, M.; Estrada, E.Y.; Fiebich, B.L.; Rosenberg, G.A. Cyclooxygenase Inhibition Limits Blood-Brain Barrier Disruption following Intracerebral Injection of Tumor Necrosis Factor-α in the Rat. Experiment 2007, 323, 488–498. [Google Scholar] [CrossRef]
- Wu, G.; Jiao, Y.; Wu, J.; Ren, S.; Wang, L.; Tang, Z.; Zhou, H. Rosiglitazone Infusion Therapy Following Minimally Invasive Surgery for Intracranial Hemorrhage Evacuation Decreased Perihematomal Glutamate Content and Blood-Brain Barrier Permeability in Rabbits. World Neurosurg. 2018, 111, e40–e46. [Google Scholar] [CrossRef]
- Wu, G.; Wu, J.; Jiao, Y.; Wang, L.; Wang, F.; Zhang, Y. Rosiglitazone infusion therapy following minimally invasive surgery for intracerebral hemorrhage evacuation decreases matrix metalloproteinase-9 and blood-brain barrier disruption in rabbits. BMC Neurol. 2015, 15, 37. [Google Scholar] [CrossRef] [PubMed]
- Culman, J.; Nguyen-Ngoc, M.; Glatz, T.; Gohlke, P.; Herdegen, T.; Zhao, Y. Treatment of rats with pioglitazone in the reperfusion phase of focal cerebral ischemia: A preclinical stroke trial. Exp. Neurol. 2012, 238, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Gao, D.; Hei, Y.; Zhang, X.; Chen, X.; Fei, Z. D-allose protects the blood brain barrier through PPARγ-mediated anti-inflammatory pathway in the mice model of ischemia reperfusion injury. Brain Res. 2016, 1642, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Slifer, Z.M.; Krishnan, B.R.; Madan, J.; Blikslager, A.T. Larazotide acetate: A pharmacological peptide approach to tight junction regulation. Am. J. Physiol. Liver Physiol. 2021, 320, G983–G989. [Google Scholar] [CrossRef] [PubMed]
- Safety of Larazotide Acetate in Healthy Volunteers. Available online: https://clinicaltrials.gov/ct2/show/NCT00386490 (accessed on 17 March 2023).
- Safety Study of Larazotide Acetate to Treat Celiac Disease. Available online: https://clinicaltrials.gov/ct2/show/NCT00386165f (accessed on 17 March 2023).
- A Leffler, D.; Kelly, C.P.; Abdallah, H.Z.; Colatrella, A.M.; A Harris, L.; Leon, F.; A Arterburn, L.; Paterson, B.M.; Lan, Z.H.; Murray, J. A Randomized, Double-Blind Study of Larazotide Acetate to Prevent the Activation of Celiac Disease During Gluten Challenge. Am. J. Gastroenterol. 2012, 107, 1554–1562. [Google Scholar] [CrossRef]
- Paterson, B.M.; Lammers, K.M.; Arrieta, M.C.; Fasano, A.; Meddings, J.B. The safety, tolerance, pharmacokinetic and pharmacodynamic effects of single doses of AT-1001 in coeliac disease subjects: A proof of concept study. Aliment. Pharmacol. Ther. 2007, 26, 757–766. [Google Scholar] [CrossRef]
- Fasano, A.; Paterson, B. Materials and Methods for the Treatment of Celiac Disease. U.S. Patent 8034776B2, 11 November 2011. [Google Scholar]
- Safety and Tolerability Study of Larazotide Acetate in Celiac Disease Subjects. Available online: https://clinicaltrials.gov/ct2/show/NCT00362856 (accessed on 17 March 2023).
- Pérez, L.C.; León, F. Clinical trial data provides hope for attenuation of mucosal injury in coeliac disease. Eur. J. Intern. Med. 2012, 23, e77. [Google Scholar] [CrossRef]
- Randomized, Double-Blind, Placebo-Controlled Study of Larazotide Acetate in Subjects with Active Celiac Disease. Available online: https://clinicaltrials.gov/ct2/show/NCT00620451 (accessed on 17 March 2023).
- Kelly, C.P.; Green, P.H.R.; Murray, J.A.; Dimarino, A.; Colatrella, A.; Leffler, D.A.; Alexander, T.; Arsenescu, R.; Leon, F.; Jiang, J.G.; et al. Larazotide acetate in patients with coeliac disease undergoing a gluten challenge: A randomised placebo-controlled study. Aliment. Pharmacol. Ther. 2013, 37, 252–262. [Google Scholar] [CrossRef]
- Study to Assess the Efficacy of Larazotide Acetate for the Treatment of Celiac Disease. Available online: https://www.clinicaltrials.gov/ct2/show/NCT00492960 (accessed on 17 March 2023).
- Leffler, D.A.; Kelly, C.P.; Green, P.H.; Fedorak, R.; DiMarino, A.; Perrow, W.; Rasmussen, H.; Wang, C.; Bercik, P.; Bachir, N.M.; et al. Larazotide Acetate for Persistent Symptoms of Celiac Disease Despite a Gluten-Free Diet: A Randomized Controlled Trial. Gastroenterology 2015, 148, 1311–1319.e6. [Google Scholar] [CrossRef]
- A Double-blind Placebo-controlled Study to Evaluate Larazotide Acetate for the Treatment of Celiac Disease. Available online: https://www.clinicaltrials.gov/ct2/show/NCT01396213 (accessed on 17 March 2023).
- Study to Evaluate the Efficacy and Safety of Larazotide Acetate for the Relief of CeD Symptoms. Available online: https://clinicaltrials.gov/ct2/show/NCT03569007 (accessed on 17 March 2023).
- Machado, M.V. New Developments in Celiac Disease Treatment. Int. J. Mol. Sci. 2023, 24, 945. [Google Scholar] [CrossRef]
- Yonker, L.M.; Swank, Z.; Gilboa, T.; Senussi, Y.M.; Kenyon, V.B.; Papadakis, L.B.; Boribong, B.P.; Carroll, R.W.M.; Walt, D.R.; Fasano, A. Zonulin Antagonist, Larazotide (AT1001), As an Adjuvant Treatment for Multisystem Inflammatory Syndrome in Children: A Case Series. Crit. Care Explor. 2022, 10, e0641. [Google Scholar] [CrossRef] [PubMed]
- AT1001 for the Treatment of COVID-19 Related MIS-C. Available online: https://clinicaltrials.gov/ct2/show/NCT05022303 (accessed on 17 March 2023).
- Al Refaei, A. Larazotide acetate as a preventive and therapeutic pharmacotherapy in obesity and metabolic syndrome. Med. Hypotheses 2022, 167, 110940. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Durai, M.; Kitchens, K.; Tamiz, A.P.; Somerville, R.; Ginski, M.; Paterson, B.M.; Murray, J.A.; Verdu, E.F.; Alkan, S.S.; et al. Larazotide acetate regulates epithelial tight junctions in vitro and in vivo. Peptides 2012, 35, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.; Jury, J.; Sanz, Y.; Wiepjes, M.; Huang, X.; Murray, J.; David, C.S.; Fasano, A.; Verdú, E.F. Increased Bacterial Translocation in Gluten-Sensitive Mice Is Independent of Small Intestinal Paracellular Permeability Defect. Dig. Dis. Sci. 2012, 57, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Shen, T.; Chen, H.; Zhou, Y.; Zhang, P.; Ma, Y.; Moyer, M.P.; Zhang, M.; Chu, Z.; Qin, H. Functional characterization of MIMP for its adhesion to the intestinal epithelium. Front. Biosci. 2011, 16, 2106–2127. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, M.C.; Madsen, K.; Doyle, J.; Meddings, J. Reducing small intestinal permeability attenuates colitis in the IL10 gene-deficient mouse. Gut 2008, 58, 41–48. [Google Scholar] [CrossRef]
- Sturgeon, C.; Lan, J.; Fasano, A. Zonulin transgenic mice show altered gut permeability and increased morbidity/mortality in the DSS colitis model. Ann. N. Y. Acad. Sci. 2017, 1397, 130–142. [Google Scholar] [CrossRef]
- Kwak, S.Y.; Jang, W.I.; Park, S.; Cho, S.S.; Lee, S.B.; Kim, M.-J.; Park, S.; Shim, S.; Jang, H. Metallothionein 2 activation by pravastatin reinforces epithelial integrity and ameliorates radiation-induced enteropathy. Ebiomedicine 2021, 73, 103641. [Google Scholar] [CrossRef]
- Enomoto, H.; Yeatts, J.; Carbajal, L.; Krishnan, B.R.; Madan, J.P.; Laumas, S.; Blikslager, A.T.; Messenger, K.M. In vivo assessment of a delayed release formulation of larazotide acetate indicated for celiac disease using a porcine model. PLoS ONE 2021, 16, e0249179. [Google Scholar] [CrossRef]
- Matei, D.E.; Menon, M.; Alber, D.G.; Smith, A.M.; Nedjat-Shokouhi, B.; Fasano, A.; Magill, L.; Duhlin, A.; Bitoun, S.; Gleizes, A.; et al. Intestinal barrier dysfunction plays an integral role in arthritis pathology and can be targeted to ameliorate disease. Med 2021, 2, 864–883.e9. [Google Scholar] [CrossRef]
- Rivas, M.N.; Wakita, D.; Franklin, M.K.; Carvalho, T.T.; Abolhesn, A.; Gomez, A.C.; Fishbein, M.C.; Chen, S.; Lehman, T.J.; Sato, K.; et al. Intestinal Permeability and IgA Provoke Immune Vasculitis Linked to Cardiovascular Inflammation. Immunity 2019, 51, 508–521.e6. [Google Scholar] [CrossRef] [PubMed]
- Shirey, K.; Lai, W.; Patel, M.; Pletneva, L.; Pang, C.; Kurt-Jones, E.; Lipsky, M.; Roger, T.; Calandra, T.; Tracey, K.; et al. Novel strategies for targeting innate immune responses to influenza. Mucosal Immunol. 2016, 9, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Min, S.; Zhu, M.; He, L.; Zhang, Y.; Li, J.; Tian, Y.; Yu, G.; Wu, L.; Cong, X. The Role of Endothelial Barrier Function in the Fibrosis of Salivary Gland. J. Dent. Res. 2022, 102, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, R.; Marioneaux, J.; Bhat, M.; Prior, C.; Madan, J.; Laumas, S.; Sanyal, A. FRI-267-Serial measurement of serum dextran absorption by novel competition ELISA demonstrates larazotide acetate significantly improves “leaky gut” in a Western diet murine model of metabolic liver disease. J. Hepatol. 2019, 70, e511–e512. [Google Scholar] [CrossRef]
- Caliskan, A.R.; Gul, M.; Yılmaz, I.; Otlu, B.; Uremis, N.; Uremis, M.M.; Kilicaslan, I.; Gul, S.; Tikici, D.; Saglam, O.; et al. Effects of larazotide acetate, a tight junction regulator, on the liver and intestinal damage in acute liver failure in rats. Hum. Exp. Toxicol. 2021, 40, S693–S701. [Google Scholar] [CrossRef] [PubMed]
- Haddadzadegan, S.; Dorkoosh, F.; Bernkop-Schnürch, A. Oral delivery of therapeutic peptides and proteins: Technology landscape of lipid-based nanocarriers. Adv. Drug Deliv. Rev. 2022, 182, 114097. [Google Scholar] [CrossRef] [PubMed]
- Di Micco, S.; Musella, S.; Sala, M.; Scala, M.C.; Andrei, G.; Snoeck, R.; Bifulco, G.; Campiglia, P.; Fasano, A. Peptide derivatives of the zonulin inhibitor larazotide (AT1001) as potential anti SARS-CoV-2: Molecular modelling, synthesis and bioactivity evaluation. Int. J. Mol. Sci. 2021, 22, 9427. [Google Scholar] [CrossRef] [PubMed]
- Di Micco, S.; Musella, S.; Scala, M.C.; Sala, M.; Campiglia, P.; Bifulco, G.; Fasano, A. In silico analysis revealed potential anti-SARS-CoV-2 main protease activity by the zonulin inhibitor larazotide acetate. Front. Chem. 2021, 8, 628609. [Google Scholar] [CrossRef]
- Di Micco, S.; Rahimova, R.; Sala, M.; Scala, M.C.; Vivenzio, G.; Musella, S.; Andrei, G.; Remans, K.; Mammri, L.; Snoeck, R. Rational design of the zonulin inhibitor AT1001 derivatives as potential anti SARS-CoV-2. Eur. J. Med. Chem. 2022, 244, 114857. [Google Scholar] [CrossRef]
- Motlekar, N.A.; Fasano, A.; Wachtel, M.S.; Youan, B.-B.C. Zonula occludens toxin synthetic peptide derivative AT1002 enhances in vitro and in vivo intestinal absorption of low molecular weight heparin. J. Drug Target. 2006, 14, 321. [Google Scholar] [CrossRef]
- Li, M.; Oliver, E.; Kitchens, K.M.; Vere, J.; Alkan, S.S.; Tamiz, A.P. Structure–activity relationship studies of permeability modulating peptide AT-1002. Bioorganic Med. Chem. Lett. 2008, 18, 4584–4586. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.; Zhao, Z.; He, J.; Tao, Y.; Zhang, H.; Yuan, R.; Sun, K.; Shi, Y. Preparation, Drug Distribution, and In Vivo Evaluation of the Safety of Protein Corona Liposomes for Liraglutide Delivery. Nanomaterials 2023, 13, 540. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.; Ragupathy, S.; Borchard, G. Target specific tight junction modulators. Adv. Drug Deliv. Rev. 2021, 171, 266–288. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, Y.; Heo, G.; Jeong, S.; Park, S.; Yoo, J.-W.; Jung, Y.; Im, E. Modulation of Intestinal Epithelial Permeability via Protease-Activated Receptor-2-Induced Autophagy. Cells 2022, 11, 878. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Yu, S.-J.; Tsai, J.-J.; Yu, C.-H.; Liao, E.-C. Antagonism of Protease Activated Receptor-2 by GB88 Reduces Inflammation Triggered by Protease Allergen Tyr-p3. Front. Immunol. 2021, 12, 557433. [Google Scholar] [CrossRef] [PubMed]
- Ushakumari, C.J.; Zhou, Q.L.; Wang, Y.-H.; Na, S.; Rigor, M.C.; Zhou, C.Y.; Kroll, M.K.; Lin, B.D.; Jiang, Z.Y. Neutrophil Elastase Increases Vascular Permeability and Leukocyte Transmigration in Cultured Endothelial Cells and Obese Mice. Cells 2022, 11, 2288. [Google Scholar] [CrossRef]
- Xu, B.; Chen, J.; Fu, J.; Yang, R.; Yang, B.; Huo, D.; Tan, C.; Chen, H.; Wang, X. Meningitic Escherichia coli-Induced Interleukin-17A Facilitates Blood–Brain Barrier Disruption via Inhibiting Proteinase 3/Protease-Activated Receptor 2 Axis. Front. Cell Neurosci. 2022, 16, 814867. [Google Scholar] [CrossRef]
- Liu, W.; Wang, P.; Shang, C.; Chen, L.; Cai, H.; Ma, J.; Yao, Y.; Shang, X.; Xue, Y. Endophilin-1 regulates blood–brain barrier permeability by controlling ZO-1 and occludin expression via the EGFR–ERK1/2 pathway. Brain Res. 2014, 1573, 17–26. [Google Scholar] [CrossRef]
- Petecchia, L.; Sabatini, F.; Usai, C.; Caci, E.; Varesio, L.; Rossi, G.A. Cytokines induce tight junction disassembly in airway cells via an EGFR-dependent MAPK/ERK1/2-pathway. Lab. Investig. 2012, 92, 1140–1148. [Google Scholar] [CrossRef]
- Kakei, Y.; Teraoka, S.; Akashi, M.; Hasegawa, T.; Komori, T. Changes in cell junctions induced by inhibition of epidermal growth factor receptor in oral squamous cell carcinoma cells. Exp. Ther. Med. 2017, 14, 953–960. [Google Scholar] [CrossRef]
- Fan, L.; Hu, L.; Yang, B.; Fang, X.; Gao, Z.; Li, W.; Sun, Y.; Shen, Y.; Wu, X.; Shu, Y.; et al. Erlotinib promotes endoplasmic reticulum stress-mediated injury in the intestinal epithelium. Toxicol. Appl. Pharmacol. 2014, 278, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Gu, Y.; Gao, Z.; Guo, L.; Guo, W.; Wu, X.; Shen, Y.; Sun, Y.; Wu, X.; Xu, Q. EGFR inhibitor-driven endoplasmic reticulum stress-mediated injury on intestinal epithelial cells. Life Sci. 2014, 119, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Van Sebille, Y.Z.; Gibson, R.J.; Wardill, H.R.; Ball, I.A.; Keefe, D.M.; Bowen, J.M. Dacomitinib-induced diarrhea: Targeting chloride secretion with crofelemer. Int. J. Cancer 2018, 142, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Van Sebille, Y.Z.; Gibson, R.J.; Wardill, H.R.; Secombe, K.R.; Ball, I.A.; Keefe, D.M.; Finnie, J.W.; Bowen, J.M. Dacomitinib-induced diarrhoea is associated with altered gastrointestinal permeability and disruption in ileal histology in rats. Int. J. Cancer 2017, 140, 2820–2829. [Google Scholar] [CrossRef]
- Leech, A.O.; Vellanki, S.H.; Rutherford, E.J.; Keogh, A.; Jahns, H.; Hudson, L.; O’donovan, N.; Sabri, S.; Abdulkarim, B.; Sheehan, K.M.; et al. Cleavage of the extracellular domain of junctional adhesion molecule-A is associated with resistance to anti-HER2 therapies in breast cancer settings. Breast Cancer Res. 2018, 20, 140. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.; Zou, H.; Cai, N.; Chen, X.; Xu, L.; Kong, X.; Liu, P. The multi-targeted tyrosine kinase inhibitor vandetanib plays a bifunctional role in non-small cell lung cancer cells. Sci. Rep. 2015, 5, 8629. [Google Scholar] [CrossRef]
- Hirsh, V.; Blais, N.; Burkes, R.; Verma, S.; Croitoru, K. Management of Diarrhea Induced by Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors. Curr. Oncol. 2014, 21, 329–336. [Google Scholar] [CrossRef]
- Oda, K.; Matsuoka, Y.; Funahashi, A.; Kitano, H. A comprehensive pathway map of epidermal growth factor receptor signaling. Mol. Syst. Biol. 2005, 1, 2005-0010. [Google Scholar] [CrossRef]
- Pellegrini, C.; D’antongiovanni, V.; Miraglia, F.; Rota, L.; Benvenuti, L.; Di Salvo, C.; Testa, G.; Capsoni, S.; Carta, G.; Antonioli, L.; et al. Enteric α-synuclein impairs intestinal epithelial barrier through caspase-1-inflammasome signaling in Parkinson’s disease before brain pathology. NPJ Park. Dis. 2022, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Study to Compare the Efficacy and Safety of Oral AT1001 and Enzyme Replacement Therapy in Patients with Fabry Disease. Available online: https://clinicaltrials.gov/ct2/show/NCT01218659 (accessed on 17 March 2023).
- Study of the Effects of Oral AT1001 (Migalastat Hydrochloride) in Patients with Fabry Disease. Available online: https://clinicaltrials.gov/ct2/show/NCT00925301 (accessed on 17 March 2023).
- Toll, L.; Zaveri, N.T.; E Polgar, W.; Jiang, F.; Khroyan, T.V.; Zhou, W.; Xie, X.S.; Stauber, G.B.; Costello, M.R.; Leslie, F.M. AT-1001: A High Affinity and Selective α3β4 Nicotinic Acetylcholine Receptor Antagonist Blocks Nicotine Self-Administration in Rats. Neuropsychopharmacology 2012, 37, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Talley, N.J.; Holtmann, G.J.; Jones, M.; A Koloski, N.; Walker, M.M.; Burns, G.; E Potter, M.D.; Shah, A.; Keely, S. Zonulin in serum as a biomarker fails to identify the IBS, functional dyspepsia and non-coeliac wheat sensitivity. Gut 2020, 69, 1719–1722. [Google Scholar] [CrossRef] [PubMed]
- Hałasa, M.; Maciejewska, D.; Ryterska, K.; Baśkiewicz-Hałasa, M.; Safranow, K.; Stachowska, E. Assessing the Association of Elevated Zonulin Concentration in Stool with Increased Intestinal Permeability in Active Professional Athletes. Medicina 2019, 55, 710. [Google Scholar] [CrossRef] [PubMed]
- Kuzma, J.N.; Hagman, D.K.; Cromer, G.; Breymeyer, K.L.; Roth, C.L.; Foster-Schubert, K.E.; Holte, S.E.; Weigle, D.S.; Kratz, M. Intraindividual Variation in Markers of Intestinal Permeability and Adipose Tissue Inflammation in Healthy Normal-Weight to Obese AdultsBiomarker Reliability: Adipose Inflammation Gut Permeability. Cancer Epidemiol. Biomark. Prev. 2019, 28, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Linsalata, M.; Riezzo, G.; D’attoma, B.; Clemente, C.; Orlando, A.; Russo, F. Noninvasive biomarkers of gut barrier function identify two subtypes of patients suffering from diarrhoea predominant-IBS: A case-control study. BMC Gastroenterol. 2018, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Meira de-Faria, F.; Bednarska, O.; Ström, M.; Söderholm, J.D.; Walter, S.A.; Keita, Å.V. Colonic paracellular permeability and circulating zonulin-related proteins. Scand. J. Gastroenterol. 2021, 56, 424–431. [Google Scholar] [CrossRef]
- Fasano, A. All disease begins in the (leaky) gut: Role of zonulin-mediated gut permeability in the pathogenesis of some chronic inflammatory diseases. F1000Research 2020, 9, 69. [Google Scholar] [CrossRef]
- Scheffler, L.; Crane, A.; Heyne, H.; Tönjes, A.; Schleinitz, D.; Ihling, C.H.; Stumvoll, M.; Freire, R.; Fiorentino, M.; Fasano, A.; et al. Widely Used Commercial ELISA Does Not Detect Precursor of Haptoglobin2, but Recognizes Properdin as a Potential Second Member of the Zonulin Family. Front. Endocrinol. 2018, 9, 22. [Google Scholar] [CrossRef]
- Ajamian, M.; Steer, D.; Rosella, G.; Gibson, P.R. Serum zonulin as a marker of intestinal mucosal barrier function: May not be what it seems. PLoS ONE 2019, 14, e0210728. [Google Scholar] [CrossRef]
- Massier, L.; Chakaroun, R.; Kovacs, P.; Heiker, J.T. Blurring the picture in leaky gut research: How shortcomings of zonulin as a biomarker mislead the field of intestinal permeability. Gut 2021, 70, 1801–1802. [Google Scholar] [CrossRef]
- Sollid, L.M.; Koning, F. Lack of relationship of AT1001 to zonulin and prehaptoglobin-2: Clinical implications. Gut 2020, 70, 2211–2212. [Google Scholar] [CrossRef]
- Fasano, A. Zonulin measurement conundrum: Add confusion to confusion does not lead to clarity. Gut 2021, 70, 2007–2008. [Google Scholar] [CrossRef] [PubMed]
- Konno, T.; Martinez, E.E.; Ji, J.; Miranda-Ribera, A.; Fiorentino, M.R.; Fasano, A. Human coagulation factor X and CD5 antigen-like are potential new members of the zonulin family proteins. Biochem. Biophys. Res. Commun. 2023, 638, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Memon, A.A.; Palmér, K.; Hedelius, A.; Sundquist, J.; Sundquist, K. The association of zonulin-related proteins with prevalent and incident inflammatory bowel disease. BMC Gastroenterol. 2022, 22, 3. [Google Scholar] [CrossRef] [PubMed]

| Gut Microbiome Member | Population | Zonulin Levels in Relation with Microbial Abundance | Ref. |
|---|---|---|---|
| Escherichia coli | ankylosing spondylitis patients | ↑ | [59] |
| relatively healthy elderly volunteers | ↑ | [60] | |
| healthy adult volunteers | ↑ | [61] | |
| Bacteroides | normal weight and obese volunteers | ↑ | [62] |
| Hashimoto-thyroiditis patients | ↑ | [63] | |
| Prevotella | ankylosing spondylitis patients | ↑ | [59] |
| obese colorectal carcinoma patients | ↑ | [64] | |
| Pseudomonas | relatively healthy elderly volunteers | ↑ | [60] |
| Shigella | ↑ | ||
| γ-Proteobacteria | ↑ | ||
| Rhizobiales | ↑ | ||
| Firmicutes | normal weight and obese volunteers | ↑ | [62] |
| Erysipelotrichales | healthy women | ↑ | [65] |
| Actinobacteria | relatively healthy elderly volunteers | ↑ | [60] |
| Clostridium | healthy adult volunteers | ↑ | [61] |
| Enteroviridae | celiac disease with or without T1D | ↑ | [66] |
| LPS (in serum) | community-acquired pneumonia patients | ↑ | [67] |
| precocious acute myocardial infarction patients | ↑ | [68] | |
| T1D | ↑ | [69] | |
| Graves’ disease patients | ↑ | [70] | |
| children with IgE mediated and non-IgE-mediated food allergy | ↑ | [71] | |
| vitiligo patients | ↑ | [72] | |
| adolescents with major depressive disorder | ↑ | [73] | |
| septic patients | ↑ | [74] | |
| Lachnoclostridium | healthy newborns | ↑ | [75] |
| Ruminococcus gnavus | ↑ | ||
| Ruminococcus torques | ↑ | ||
| Erysipelotrichales | ↑ | ||
| Coriobacteriales | ↑ | ||
| Alphaproteobacteria | ↓ | ||
| Corynebacterium | ↓ | ||
| Pdeudomonadales | ↓ | ||
| Moraxellaceae | ↓ | ||
| Staphylococcus | ↓ | ||
| Bifidobacterium | Hashimoto-thyroiditis patients | ↓ | [63] |
| Lactobacillus spp. | healthy adult volunteers | ↓ | [61] |
| Ruminococcaceae | healthy women | ↓ | [65] |
| Faecalibacterium | ↓ | ||
| Odoribacter | ↓ | ||
| Rikenellaceae | ↓ |
| Species | Strain | Treatment and Population | Findings | Ref. | |
|---|---|---|---|---|---|
| Blood Zonulin | Fecal Zonulin | ||||
| Lactobacillus plantarum | CGMCC no.1258 | pre- and postoperative probiotic treatment of patients operated on for colorectal carcinoma | ↓ | NE | [76] |
| Lactobacillus acidophilus | 11 | ||||
| Bifidobacterium longum | 88 | ||||
| Lactobacillus plantarum | CGMCC no.1258 | pre- and postoperative probiotic treatment of patients operated on for colorectal carcinoma and liver metastasis | ↓ | NE | [77] |
| Lactobacillus acidophilus | 11 | ||||
| Bifidobacterium longum | 88 | ||||
| Bifidobacterium animalis | lactis 420 | probiotic and fiber treatment of healthy overweight volunteers | ↓ | NE | [78] |
| SCM-III synbiotic mixture: | synbiotic treatment of healthy stressed individuals | ↓ | ↓ | [79] | |
| Lactobacillus acidophilus | 145 | ||||
| Lactobacillus helveticus | ATC15009 | ||||
| Bifidobacterium | 420 | probiotic treatment of healthy stressed individuals | - | ↓ | |
| P3T/J probiotic mixture: | |||||
| Bifidobacterium animalis | lactis Bi1 | ||||
| Bifidobacterium breve | Bbr8 | synbiotic and probiotic treatment of healthy stressed individuals | ↓ | ↓ | |
| Lactobacillus acidophilus | LA1 | ||||
| Lactobacillus paracasei | 101/37 | ||||
| Bifidobacterium lactis | W51 | dietary changes and probiotic treatment in obese patients | NE | ↓ | [80] |
| W52 | |||||
| Lactobacillus acidophilus | W22 | ||||
| Lactobacillus paracasei | W20 | ||||
| Lactobacillus plantarum | W21 | ||||
| Lactobacillus salivarius | W24 | ||||
| Lactococcus lactis | W19 | ||||
| Bifidobacterium bifidum | W23 | impact of exercise in trained men treated with probiotics | NE | ↓ | [81] |
| Bifidobacterium lactis | W51 | ||||
| Enterococcus faecium | W54 | ||||
| Lactobacillus acidophilus | W22 | ||||
| Lactobacillus brevis | W63 | ||||
| Lactococcus lactis | W58 | ||||
| Bifidobacterium bifidum | W23 | synbiotic treatment of healthy volunteers | - | NE | [82] |
| Bifidobacterium lactis | W51 | ||||
| W52 | |||||
| Lactobacillus acidophilus | W22 | ||||
| Lactobacillus casei | W56 | ||||
| Lactobacillus paracasei | W20 | ||||
| Lactobacillus plantarum | W62 | ||||
| Lactobacillus salivarius | W24 | ||||
| Lactococcus lactis | W19 | ||||
| Bifidobacterium lactis | synbiotic treatment of children with NAFLD | - | NE | [83] | |
| Lactobacillus acidophilus | |||||
| Lactobacillus casei | |||||
| Bifidobacterium bifidum | W23 | probiotic treatment of migraine patients | - | - | [84] |
| Bifidobacterium lactis | W52 | ||||
| Lactobacillus acidophilus | W37 | ||||
| Lactobacillus brevis | W63 | ||||
| Lactobacillus casei | W56 | probiotic treatment of ulcerative colitis patients | ↓ | - | [85] |
| Lactobacillus salivarius | W24 | ||||
| Lactococcus lactis | W19 | ||||
| W58 | |||||
| Bacillus subtilis | DE111 | probiotic treatment of professional baseball players | - | NE | [86] |
| Species | Strain | Cell Line/ Experimental Model | Findings | Ref. | |
|---|---|---|---|---|---|
| Zonulin | ZO-1 | ||||
| Escherichia coli | 6-1 | CaCo2 | ↑ | ↓, disruption | [42] |
| rat, rabbit, and monkey small intestinal organoids | ↑ | NE | |||
| K-12 DH5α | rabbit and monkey small intestinal organoids | ↑ | NE | ||
| 21-1 | rabbit small intestinal organoids | ↑ | NE | ||
| K88 | 4-day-old piglets | ↑ | ↓ | [92] | |
| K88 | IPEC-J2 | - | ↓ | ||
| RY13 | HT-29 | - | NE | [93] | |
| K12 DH5α | - | NE | |||
| 042, JM221 | T84 | NE | disruption | [94] | |
| 055:B5 (LPS) | CaCo2 | ↑ | ↓ | [52] | |
| CaCo2 | ↑ | NE | [59] | ||
| HB101 | T84 | NE | disruption | [95] | |
| Bacteroidales and Escherichia coli | malnourished mice | ↑ | ↓ | [96] | |
| Salmonella typhimurium | SO1344 | rabbit small intestinal organoids | ↑ | NE | [42] |
| Pseudomonas fluorescens | CaCo2 | ↑ | disruption | [97] | |
| Prevotella | CaCo2 | ↑ | NE | [59] | |
| Acetobacter ghanensis | CaCo2 treated with PT-gliadin | ↓ | - | [98] | |
| Porphyromonas gingivalis | healthy mice | NE | ↓ | [99] | |
| Pseudomonas aeruginosa | pneumonia induced in mice | NE | ↑ | [100] | |
| Fusobacterium nucleatum | CaCo2 | NE | ↓ | [101] | |
| DSS-induced colitis in mice | NE | ↓, disruption | |||
| Ruminococcus blautia gnavus | VPI C7-9 | germ-free mice | - | NE | [102] |
| CC55_001C | - | NE | |||
| S107-48 | ↑ | NE | |||
| S47-18 | - | NE | |||
| Clostridium difficile toxin A and B | T84 | NE | disruption | [103] | |
| Faecalibacterium prausnitzii MAM | NCM460 transfection | NE | ↑ | [104] | |
| Caco2 transfection | NE | ↑ | |||
| HT-29 transfection | NE | ↑ | |||
| diabetes mellitus induced in mice | NE | ↑ | |||
| Lactobacillus rhamnosus | GG | CaCo2 treated with gliadin | ↓ | ↑ | [91] |
| HT-29 | ↑ | NE | [93] | ||
| P1 | HT-29 treated with PT-gliadin | ↓ | ↑ | [105] | |
| P2 | ↓ | ↑ | |||
| F1 | ↓ | ↑ | |||
| P3 | ↓ | - | |||
| GG | ↓ | ↑ | |||
| Lactobacillus casei | C1 | HT-29 treated with PT-gliadin | ↓ | ↑ | |
| Bifidobacterium longum | CECT-7347 | HT-29 treated with TNF-α | NE | ↑ | [90] |
| T84 | NE | ↑ | [106] | ||
| HT-29 | - | NE | [93] | ||
| Bifidobacterium (not specified) | CaCo2 treated with LPS | ↓ | ↑ | [52] | |
| LPS-induced NEC in rats | ↓ | ↑ | |||
| VSL#3 | IEC-6 treated with hydrolyzed gliadin | ↓ | NE | [89] | |
| mouse small intestinal organoid treated with hydrolyzed gliadin | ↓ | NE | |||
| Lactobacillus paracasei | D3-5 | high-fat diet in old mice | NE | ↑ | [107] |
| Lactobacillus rhamnosus | D4-4 | ||||
| D7-5 | |||||
| Lactobacillus plantarum | D6-2 | ||||
| D13-4 | |||||
| Enterococcus rafnosus | D24-1 | ||||
| Enterococcus INBio | D24-2 | ||||
| Enterococcus Avium | D25-1 | ||||
| D25-2 | |||||
| D26-1 | |||||
| Lactobacillus paracasei | 101/37 LMG P-17504 | CaCo2 treated with PT-gliadin | NE | ↑ | [108] |
| Lactobacillus plantarum | 14 D CECT 4528 | ||||
| Bifidobacterium animalis | lactis Bi1 LMG P-17502 | ||||
| Bifidobacterium breve | Bbr8 LMG P-17501 | ||||
| BL10 LMG P-17500 | |||||
| Condition | Results | Study (Enrollment) | Clinical Trials Identifier | Ref. |
|---|---|---|---|---|
| Healthy | good tolerability | Phase I (24) | NCT00386490 | [196] |
| Celiac disease, gluten-free diet | good tolerability | Phase Ib (21) | NCT00386165 | [197] |
| Celiac disease, gluten challenge | improvement in GI symptoms, good tolerability | Phase IIa (80) | NCT00362856 | [198,199,200,201] |
| Celiac disease, gluten challenge | improvement in histological scores, good tolerability | Phase IIb (105) | NCT00620451 | [202,203] |
| Celiac disease, gluten challenge | improvement in GI symptoms, decreased level of anti-tTG IgA | Phase IIb (171) | NCT00492960 | [204,205] |
| Celiac disease, persistent symptoms with gluten-free diet | improvement in GI and extra-GI symptoms, good tolerability | Phase IIb (342) | NCT01396213 | [206,207] |
| Celiac disease, gluten-free diet | (terminated based on interim analysis) | Phase III (307) | NCT03569007 | [208,209] |
| COVID19—MIS-C | improvement in clinical symptoms, decreased level of inflammatory markers and SARS-CoV-2 nucleocapsid (N) protein | case report (1) | [178] | |
| COVID19—MIS-C | improvement in GI symptoms, decreased level of SARS-CoV-2 Spike (S) protein | case series (4) | [210] | |
| COVID19—MIS-C | (not completed) | Phase IIa (20) | NCT05022303 | [211] |
| Model | Species | Administration | Daily Dose | Results | Ref. |
|---|---|---|---|---|---|
| celiac disease | gliadin-sensitized HLA-HCD4/DQ8 transgenic mouse | p.o. gavage | 0.25 mg | reduced intestinal permeability and macrophage infiltration | [213] |
| p.o. gavage | 0.3 mg | reduced intestinal permeability | [214] | ||
| intestinal permeability | Il10−/− mouse | p.o. gavage | 5 mg | reduced intestinal permeability and inflammation | [215] |
| spontaneous colitis | p.o. in drinking water | 0.1 or 1 mg/mL | reduced intestinal permeability and inflammation | [216] | |
| DSS induced colitis | zonulin transgenic mouse | p.o. in drinking water | 1 mg/mL | reduced intestinal permeability | [217] |
| radiation-induced enteropathy | mouse | i.p. | 0.25 mg | improved clinical state and histological scores, inhibited bacterial translocation, elevated TJ protein levels | [218] |
| healthy (pharmacokinetics) | pig | p.o. capsule | 0.05 mg/kg | determining pharmacokinetics of larazotide acetate in the small intestine | [219] |
| Ruminococcus blautia gnavus colonization | germ-free mouse | p.o. in drinking water | 0.15 mg/mL | reduced intestinal permeability | [102] |
| spontaneous T1D | BB diabetic-prone rat | p.o. in drinking water | 0.01 mg/mL | inhibited development of diabetes | [138] |
| rheumatoid arthritis | mouse | p.o. in drinking water | 0.15 mg/mL | attenuated arthritis | [153] |
| Il10ra−/− mouse, Cldn8−/− mouse | p.o. gavage | 2 × 0.05 mg | reduced intestinal permeability, inflammation, and joint swelling | [220] | |
| vasculitis | mouse | i.p. | 0.5 mg | reduced intestinal permeability and LPS translocation, prevented cardiovascular lesions | [221] |
| LPS-induced acute lung injury | i.t. | 0.05 mg | reduced severity, decreased inflammatory markers | [12] | |
| i.v. | 0.01 or 0.025 or 0.05 mg | ||||
| influenza | i.v. | 0.15 mg | reduced severity of acute lung injury | [222] | |
| salivary gland fibrosis | i.p. | 5 mg/kg | improved epithelial barrier function, ameliorated fibrosis | [223] | |
| NAFLD | p.o. in drinking water | 0.1 or 1 mg/mL | reduced intestinal permeability | [224] | |
| p.o. gavage | 2 × 0.03 or 2 × 0.3 mg | ||||
| acute liver failure | rat | p.o. in drinking water | 0.01 mg/mL | decreased intestinal damage | [225] |
| p.o. gavage | 2 × 0.03 mg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veres-Székely, A.; Szász, C.; Pap, D.; Szebeni, B.; Bokrossy, P.; Vannay, Á. Zonulin as a Potential Therapeutic Target in Microbiota-Gut-Brain Axis Disorders: Encouraging Results and Emerging Questions. Int. J. Mol. Sci. 2023, 24, 7548. https://doi.org/10.3390/ijms24087548
Veres-Székely A, Szász C, Pap D, Szebeni B, Bokrossy P, Vannay Á. Zonulin as a Potential Therapeutic Target in Microbiota-Gut-Brain Axis Disorders: Encouraging Results and Emerging Questions. International Journal of Molecular Sciences. 2023; 24(8):7548. https://doi.org/10.3390/ijms24087548
Chicago/Turabian StyleVeres-Székely, Apor, Csenge Szász, Domonkos Pap, Beáta Szebeni, Péter Bokrossy, and Ádám Vannay. 2023. "Zonulin as a Potential Therapeutic Target in Microbiota-Gut-Brain Axis Disorders: Encouraging Results and Emerging Questions" International Journal of Molecular Sciences 24, no. 8: 7548. https://doi.org/10.3390/ijms24087548
APA StyleVeres-Székely, A., Szász, C., Pap, D., Szebeni, B., Bokrossy, P., & Vannay, Á. (2023). Zonulin as a Potential Therapeutic Target in Microbiota-Gut-Brain Axis Disorders: Encouraging Results and Emerging Questions. International Journal of Molecular Sciences, 24(8), 7548. https://doi.org/10.3390/ijms24087548