
Cannabis Use and Endocannabinoid Receptor Genes: A Pilot Study on Their Interaction on Brain Activity in First-Episode Psychosis
 , ,
, ,
Abstract
1. Introduction
Aims of the Study
2. Results
2.1. Sample Description
2.2. Neuroimaging Association Analyses
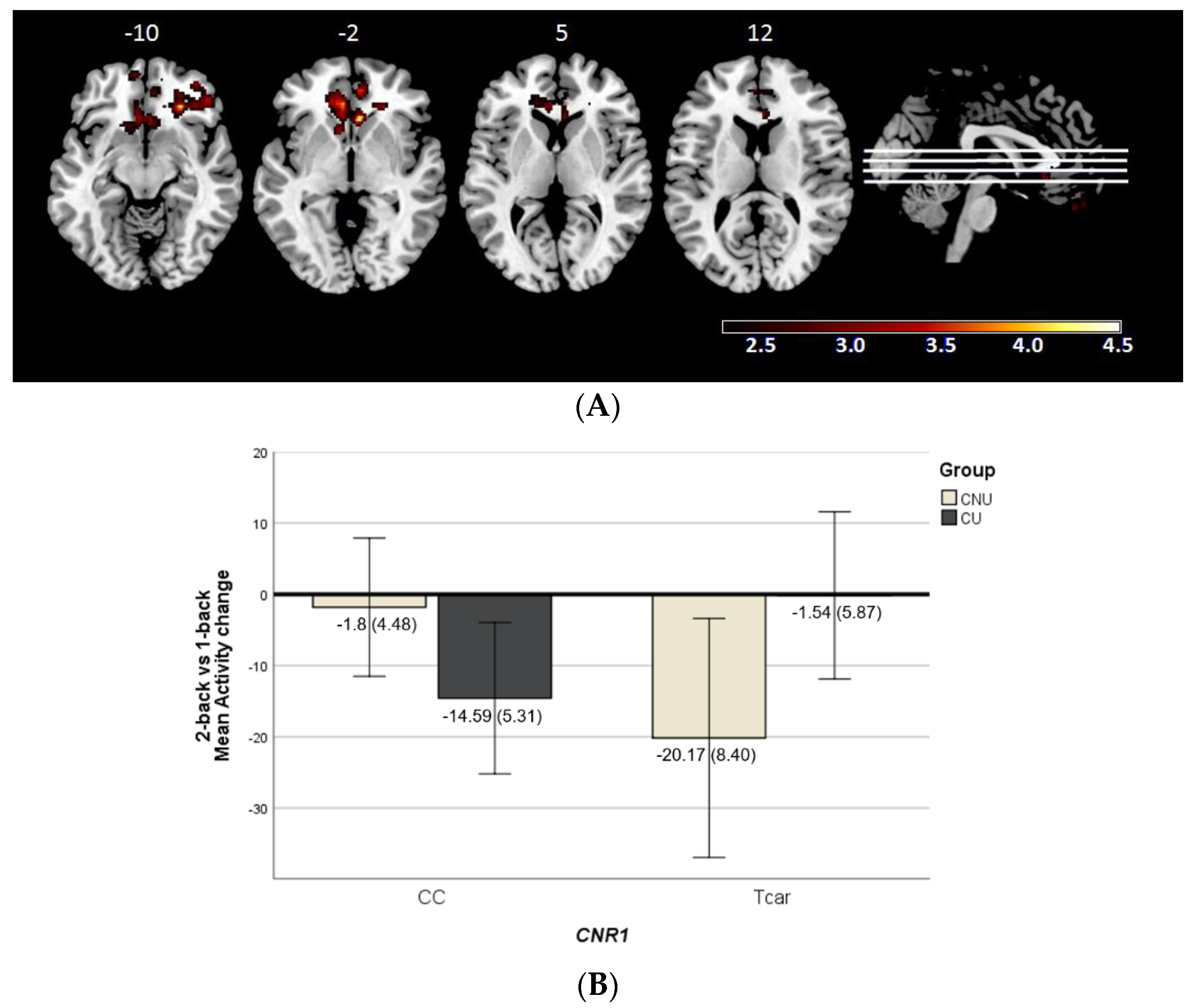
2.2.1. Cannabis Use × CNR1 Variability Interaction Effect on Brain Activity
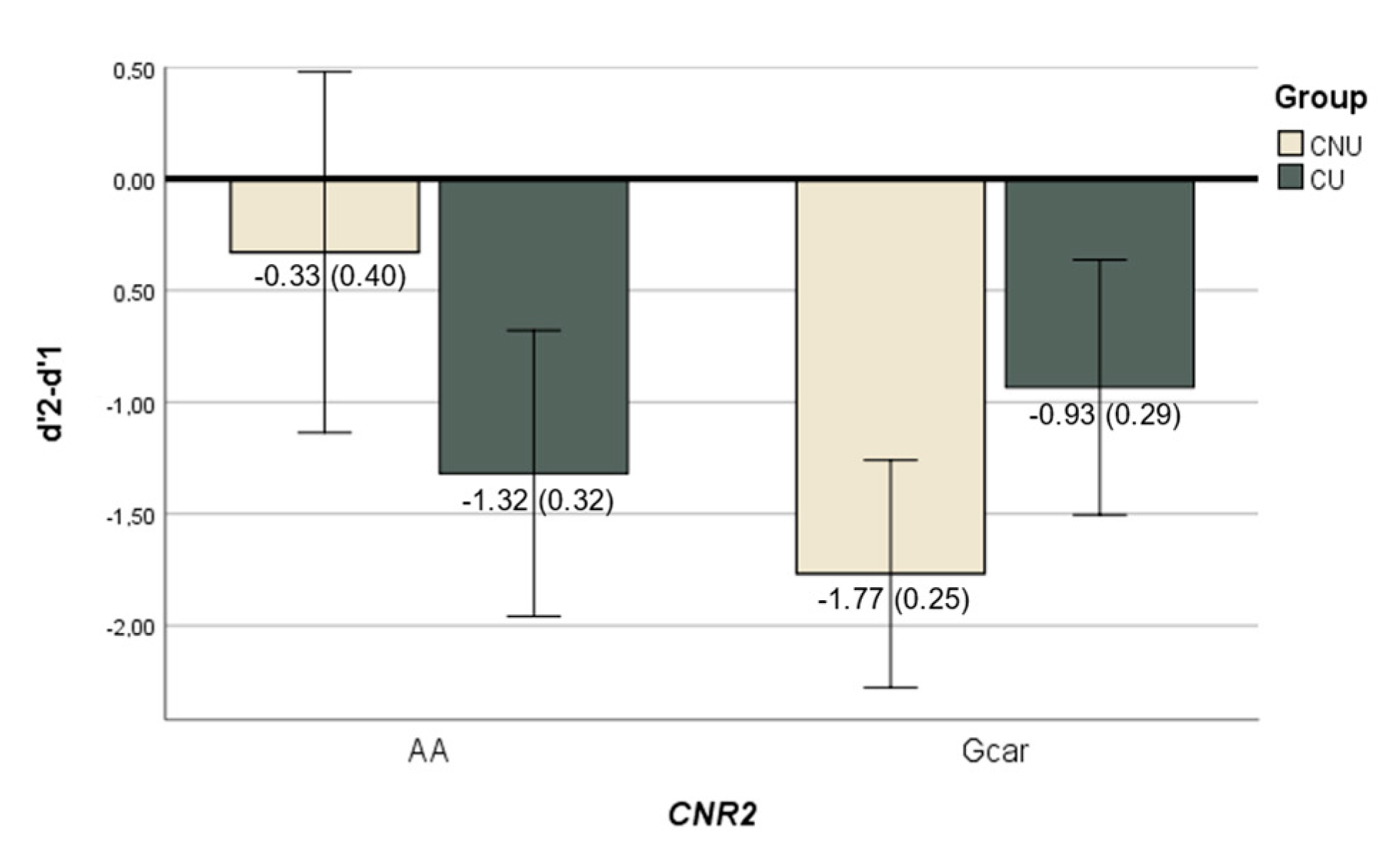
2.2.2. Cannabis Use × CNR2 Variability Interaction Effect on Brain Function
2.3. N-Back Behavioural Performance Analyses
3. Discussion
4. Materials and Methods
4.1. Participants and Study Design
4.2. Cannabis Use
4.3. fMRI Data Acquisition and fMRI Task Description
4.3.1. Acquisition Parameters
4.3.2. N-Back Task
4.3.3. N-Back Performance
4.4. Genotyping
4.5. Statistical Analyses
4.5.1. Clinical and Demographics Data Analyses
4.5.2. Neuroimaging Association Analyses
4.5.3. Behavioural Performance Association Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hilker, R.; Helenius, D.; Fagerlund, B.; Skytthe, A.; Christensen, K.; Werge, T.M.; Nordentoft, M.; Glenthøj, B. Heritability of Schizophrenia and Schizophrenia Spectrum Based on the Nationwide Danish Twin Register. Biol. Psychiatry 2018, 83, 492–498. [Google Scholar] [CrossRef]
- Sullivan, P.F.; Kendler, K.S.; Neale, M.C. Schizophrenia as a Complex Trait Evidence From a Meta-analysis of Twin Studies. Arch. Gen. Psychiatry 2003, 60, 1187–1192. [Google Scholar] [CrossRef]
- The International Schizophrenia Consortium. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 2009, 460, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Trubetskoy, V.; Pardiñas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.-Y.; Dennison, C.A.; Hall, L.S.; et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef]
- Cross-Disorder Group of the Psychiatric Genomics Consortium. Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat. Genet. 2013, 45, 984–994. [Google Scholar] [CrossRef]
- Laurens, K.R.; Luo, L.; Matheson, S.L.; Carr, V.J.; Raudino, A.; Harris, F.; Green, M.J. Common or distinct pathways to psychosis? A systematic review of evidence from prospective studies for developmental risk factors and antecedents of the schizophrenia spectrum disorders and affective psychoses. BMC Psychiatry 2015, 15, 205. [Google Scholar] [CrossRef] [PubMed]
- Tost, H.; Meyer-Lindenberg, A. Puzzling over schizophrenia: Schizophrenia, social environment and the brain. Nat. Med. 2012, 18, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Häfner, H. From Onset and Prodromal Stage to a Life-Long Course of Schizophrenia and Its Symptom Dimensions: How Sex, Age, and Other Risk Factors Influence Incidence and Course of Illness. Psychiatry J. 2019, 2019, 9804836. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.; Englund, A.; Abi-Dargham, A.; Lewis, D.; Di Forti, M.; Davies, C.; Sherif, M.; McGuire, P.; D’Souza, D. Cannabis-associated psychosis: Neural substrate and clinical impact. Neuropharmacology 2017, 124, 89–104. [Google Scholar] [CrossRef]
- Marconi, A.; Di Forti, M.; Lewis, C.; Murray, R.; Vassos, E. Meta-analysis of the Association between the Level of Cannabis Use and Risk of Psychosis. Schizophr. Bull. 2016, 42, 1262–1269. [Google Scholar] [CrossRef]
- Di Forti, M.; Sallis, H.; Allegri, F.; Trotta, A.; Ferraro, L.; Stilo, S.A.; Marconi, A.; La Cascia, C.; Marques, T.R.; Pariante, C.; et al. Daily Use, Especially of High-Potency Cannabis, Drives the Earlier Onset of Psychosis in Cannabis Users. Schizophr. Bull. 2013, 40, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- González-Pinto, A.; Alberich, S.; Barbeito, S.; Gutierrez, M.; Vega, P.; Ibáñez, B.; Haidar, M.K.; Vieta, E.; Arango, C. Cannabis and First-Episode Psychosis: Different Long-term Outcomes Depending on Continued or Discontinued Use. Schizophr. Bull. 2011, 37, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Mazzoncini, R.; Donoghue, K.; Hart, J.; Morgan, C.; Doody, G.A.; Dazzan, P.; Jones, P.B.; Morgan, K.; Murray, R.M.; Fearon, P. Illicit substance use and its correlates in first episode psychosis. Acta Psychiatr. Scand. 2010, 121, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Marino, L.; Scodes, J.; Richkin, T.; Alves-Bradford, J.-M.; Nossel, I.; Wall, M.; Dixon, L. Persistent cannabis use among young adults with early psychosis receiving coordinated specialty care in the United States. Schizophr. Res. 2020, 222, 274–282. [Google Scholar] [CrossRef]
- Pardo, M.; Matalí, J.L.; Sivoli, J.; Regina, V.-B.; Butjosa, A.; Dolz, M.; Sánchez, B.; Barajas, A.; Del Cacho, N.; Baños, I.; et al. Early onset psychosis and cannabis use: Prevalence, clinical presentation and influence of daily use. Asian J. Psychiatry 2021, 62, 102714. [Google Scholar] [CrossRef]
- Melis, M.; Muntoni, A.L.M.; Pistis, M.M. Endocannabinoids and the Processing of Value-Related Signals. Front. Pharmacol. 2012, 3, 7. [Google Scholar] [CrossRef]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; De Costa, B.R.; Rice, K.C. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci USA 1990, 87, 1932–1936. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Hernández, M.L.; Ramos, J.A. Cannabinoid-Dopamine Interaction in the Pathophysiology and Treatment of CNS Disorders. CNS Neurosci. Ther. 2010, 16, e72–e91. [Google Scholar] [CrossRef]
- Melis, M.; Pistis, M.; Perra, S.; Muntoni, A.L.; Pillolla, G.; Gessa, G.L. Endocannabinoids Mediate Presynaptic Inhibition of Glutamatergic Transmission in Rat Ventral Tegmental Area Dopamine Neurons through Activation of CB1 Receptors. J. Neurosci. 2004, 24, 53–62. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Perchuk, A.; Meozzi, P.A.; Myers, L.; Mora, Z.; Tagliaferro, P.; Gardner, E.; et al. Discovery of the Presence and Functional Expression of Cannabinoid CB2 Receptors in Brain. Ann. N. Y. Acad. Sci. 2006, 1074, 514–536. [Google Scholar] [CrossRef]
- Liu, Q.-R.; Pan, C.-H.; Hishimoto, A.; Li, C.-Y.; Xi, Z.-X.; Llorente-Berzal, A.; Viveros, M.-P.; Ishiguro, H.; Arinami, T.; Onaivi, E.S.; et al. Species differences in cannabinoid receptor 2 (CNR2gene): Identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes Brain Behav. 2009, 8, 519–530. [Google Scholar] [CrossRef]
- Jordan, C.J.; Xi, Z.-X. Progress in brain cannabinoid CB2 receptor research: From genes to behavior. Neurosci. Biobehav. Rev. 2019, 98, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-J.; Gao, M.; Gao, F.-F.; Su, Q.-X.; Wu, J. Brain cannabinoid receptor 2: Expression, function and modulation. Acta Pharmacol. Sin. 2017, 38, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bi, G.-H.; Li, X.; Li, J.; Qu, H.; Zhang, S.-J.; Li, C.-Y.; Onaivi, E.S.; Gardner, E.L.; Xi, Z.-X.; et al. Species Differences in Cannabinoid Receptor 2 and Receptor Responses to Cocaine Self-Administration in Mice and Rats. Neuropsychopharmacology 2014, 40, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Schroll, H.; Vitay, J.; Hamker, F.H. Working memory and response selection: A computational account of interactions among cortico-basalganglio-thalamic loops. Neural Netw. 2012, 26, 59–74. [Google Scholar] [CrossRef]
- Lee, J.; Park, S. Working Memory Impairments in Schizophrenia: A Meta-Analysis. J. Abnorm. Psychol. 2005, 114, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Mesholam-Gately, R.I.; Giuliano, A.J.; Goff, K.P.; Faraone, S.V.; Seidman, L.J. Neurocognition in first-episode schizophrenia: A meta-analytic review. Neuropsychology 2009, 23, 315–336. [Google Scholar] [CrossRef]
- Zhornitsky, S.; Pelletier, J.; Assaf, R.; Giroux, S.; Li, C.-S.R.; Potvin, S. Acute effects of partial CB1 receptor agonists on cognition—A meta-analysis of human studies. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 104, 110063. [Google Scholar] [CrossRef]
- Jacobsen, L.K.; Mencl, W.E.; Westerveld, M.; Pugh, K.R. Impact of Cannabis Use on Brain Function in Adolescents. Ann. N. Y. Acad. Sci. 2004, 1021, 384–390. [Google Scholar] [CrossRef]
- Harvey, M.A.; Sellman, J.D.; Porter, R.J.; Frampton, C.M. The relationship between non-acute adolescent cannabis use and cognition. Drug Alcohol Rev. 2007, 26, 309–319. [Google Scholar] [CrossRef]
- Schoeler, T.; Petros, N.; Di Forti, M.; Klamerus, E.; Foglia, E.; Ajnakina, O.; Gayer-Anderson, C.; Colizzi, M.; Quattrone, D.; Behlke, I.; et al. Effects of continuation, frequency, and type of cannabis use on relapse in the first 2 years after onset of psychosis: An observational study. Lancet Psychiatry 2016, 3, 947–953. [Google Scholar] [CrossRef]
- Bogaty, S.E.; Lee, R.S.; Hickie, I.B.; Hermens, D.F. Meta-analysis of neurocognition in young psychosis patients with current cannabis use. J. Psychiatr. Res. 2018, 99, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Løberg, E.-M. Cannabis use and cognition in schizophrenia. Front. Hum. Neurosci. 2009, 3, 53. [Google Scholar] [CrossRef]
- Yucel, M.; Bora, E.; Lubman, D.I.; Solowij, N.; Brewer, W.J.; Cotton, S.M.; Conus, P.; Takagi, M.J.; Fornito, A.; Wood, S.J.; et al. The Impact of Cannabis Use on Cognitive Functioning in Patients with Schizophrenia: A Meta-analysis of Existing Findings and New Data in a First-Episode Sample. Schizophr. Bull. 2012, 38, 316–330. [Google Scholar] [CrossRef] [PubMed]
- Mata, I.; Rodríguez-Sánchez, J.M.; Pelayo-Terán, J.M.; Pérez-Iglesias, R.; González-Blanch, C.; Ramírez-Bonilla, M.; Martínez-García, O.; Vázquez-Barquero, J.L.; Crespo-Facorro, B. Cannabis abuse is associated with decision-making impairment among first-episode patients with schizophrenia-spectrum psychosis. Psychol. Med. 2008, 38, 1257–1266. [Google Scholar] [CrossRef]
- Kanayama, G.; Rogowska, J.; Pope, H.G.; Gruber, S.A.; Yurgelun-Todd, D.A. Spatial working memory in heavy cannabis users: A functional magnetic resonance imaging study. Psychopharmacology 2004, 176, 239–247. [Google Scholar] [CrossRef]
- Smith, A.M.; Longo, C.A.; Fried, P.A.; Hogan, M.J.; Cameron, I. Effects of marijuana on visuospatial working memory: An fMRI study in young adults. Psychopharmacology 2010, 210, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Schweinsburg, A.D.; Schweinsburg, B.C.; Medina, K.L.; McQueeny, T.; Brown, S.A.; Tapert, S.F. The Influence of Recency of Use on fMRI Response During Spatial Working Memory in Adolescent Marijuana Users. J. Psychoact. Drugs 2010, 42, 401–412. [Google Scholar] [CrossRef]
- Jager, G.; Block, R.I.; Luijten, M.; Ramsey, N.F. Cannabis Use and Memory Brain Function in Adolescent Boys: A Cross-Sectional Multicenter Functional Magnetic Resonance Imaging Study. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Potvin, S.; Bourque, J.; Durand, M.; Lipp, O.; Lalonde, P.; Stip, E.; Grignon, S.; Mendrek, A. The Neural Correlates of Mental Rotation Abilities in Cannabis-Abusing Patients with Schizophrenia: An fMRI Study. Schizophr. Res. Treat. 2013, 2013, 543842. [Google Scholar] [CrossRef]
- Bourque, J.; Mendrek, A.; Durand, M.; Lakis, N.; Lipp, O.; Stip, E.; Lalonde, P.; Grignon, S.; Potvin, S. Cannabis abuse is associated with better emotional memory in schizophrenia: A functional magnetic resonance imaging study. Psychiatry Res. 2013, 214, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Løberg, E.-M.; Nygård, M.; Berle, J.; Johnsen, E.; Kroken, R.A.; Jørgensen, H.A.; Hugdahl, K. An fMRI Study of Neuronal Activation in Schizophrenia Patients with and without Previous Cannabis Use. Front. Psychiatry 2012, 3, 94. [Google Scholar] [CrossRef] [PubMed]
- Cunha, P.J.; Rosa, P.G.P.; Ayres, A.D.M.; Duran, F.L.; Santos, L.C.; Scazufca, M.; Menezes, P.R.; dos Santos, B.; Murray, R.M.; Crippa, J.A.S.; et al. Cannabis use, cognition and brain structure in first-episode psychosis. Schizophr. Res. 2013, 147, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Pelayo-Teran, J.M.; Suarez-Pinilla, P.; Chadi, N.; Crespo-Facorro, B. Gene-Environment Interactions Underlying the Effect of Cannabis in First Episode Psychosis. Curr. Pharm. Des. 2012, 18, 5024–5035. [Google Scholar] [CrossRef]
- Bioque, M.; Mas, S.; Costanzo, M.C.; Cabrera, B.; Lobo, A.; González-Pinto, A.; Rodriguez-Toscano, E.; Corripio, I.; Vieta, E.; Baeza, I.; et al. Gene-environment interaction between an endocannabinoid system genetic polymorphism and cannabis use in first episode of psychosis. Eur. Neuropsychopharmacol. 2019, 29, 786–794. [Google Scholar] [CrossRef]
- Ujike, H.; Takaki, M.; Nakata, K.; Tanaka, Y.; Takeda, T.; Kodama, M.; Fujiwara, Y.; Sakai, A.; Kuroda, S. CNR1, central cannabinoid receptor gene, associated with susceptibility to hebephrenic schizophrenia. Mol. Psychiatry 2002, 7, 515–518. [Google Scholar] [CrossRef]
- Martínez-Gras, I.; Hoenicka, J.; Ponce, G.; Rodríguez-Jiménez, R.; Jiménez-Arriero, M.A.; Perez-Hernandez, E.; Ampuero, I.; Ramos-Atance, J.A.; Palomo, T.; Rubio, G. (AAT)n repeat in the cannabinoid receptor gene, CNR1: Association with schizophrenia in a Spanish population. Eur. Arch. Psychiatry Clin. Neurosci. 2006, 256, 437–441. [Google Scholar] [CrossRef]
- Ho, B.-C.; Wassink, T.H.; Ziebell, S.; Andreasen, N.C. Cannabinoid receptor 1 gene polymorphisms and marijuana misuse interactions on white matter and cognitive deficits in schizophrenia. Schizophr. Res. 2011, 128, 66–75. [Google Scholar] [CrossRef]
- Kuzman, M.R.; Kuharic, D.B.; Ganoci, L.; Makaric, P.; Kekin, I.; Gajsak, L.R.; Prpic, N.; Bozina, T.; Bajić, Z.; Bozina, N. Association of CNR1 genotypes with changes in neurocognitive performance after eighteen-month treatment in patients with first-episode psychosis. Eur. Psychiatry 2019, 61, 88–96. [Google Scholar] [CrossRef]
- Suárez-Pinilla, P.; Roiz-Santiañez, R.; de la Foz, V.O.-G.; Guest, P.C.; Ayesa-Arriola, R.; Córdova-Palomera, A.; Tordesillas-Gutierrez, D.; Crespo-Facorro, B. Brain structural and clinical changes after first episode psychosis: Focus on cannabinoid receptor 1 polymorphisms. Psychiatry Res. 2015, 233, 112–119. [Google Scholar] [CrossRef]
- Gouvêa, E.S.; Filho, A.F.S.; Ota, V.K.; Mrad, V.; Gadelha, A.; Bressan, R.A.; Cordeiro, Q.; Belangero, S.I. The role of the CNR1 gene in schizophrenia: A systematic review including unpublished data. Revista Brasileira de Psiquiatria 2017, 39, 160–171. [Google Scholar] [CrossRef]
- Onwuameze, O.E.; Nam, K.W.; Epping, E.A.; Wassink, T.H.; Ziebell, S.; Andreasen, N.C.; Ho, B.-C. MAPK14 and CNR1 gene variant interactions: Effects on brain volume deficits in schizophrenia patients with marijuana misuse. Psychol. Med. 2013, 43, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Schacht, J.P.; Hutchison, K.E.; Filbey, F.M. Associations between Cannabinoid Receptor-1 (CNR1) Variation and Hippocampus and Amygdala Volumes in Heavy Cannabis Users. Neuropsychopharmacology 2012, 37, 2368–2376. [Google Scholar] [CrossRef]
- Colizzi, M.; Iyegbe, C.; Powell, J.; Ursini, G.; Porcelli, A.; Bonvino, A.; Taurisano, P.; Romano, R.; Masellis, R.; Blasi, G.; et al. Interaction between Functional Genetic Variation of DRD2 and Cannabis Use on Risk of Psychosis. Schizophr. Bull. 2015, 41, 1171–1182. [Google Scholar] [CrossRef]
- Taurisano, P.; Antonucci, L.A.; Fazio, L.; Rampino, A.; Romano, R.; Porcelli, A.; Masellis, R.; Colizzi, M.; Quarto, T.; Torretta, S.; et al. Prefrontal activity during working memory is modulated by the interaction of variation in CB1 and COX2 coding genes and correlates with frequency of cannabis use. Cortex 2016, 81, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Taurisano, P.; Pergola, G.; Monda, A.; Antonucci, L.A.; Di Carlo, P.; Piarulli, F.; Passiatore, R.; Papalino, M.; Romano, R.; Monaco, A.; et al. The interaction between cannabis use and a CB1-related polygenic co-expression index modulates dorsolateral prefrontal activity during working memory processing. Brain Imaging Behav. 2021, 15, 288–299. [Google Scholar] [CrossRef]
- Quickfall, J.; Crockford, D. Brain Neuroimaging in Cannabis Use: A Review. J. Neuropsychiatry Clin. Neurosci. 2006, 18, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.N.; Knutson, B. The Reward Circuit: Linking Primate Anatomy and Human Imaging. Neuropsychopharmacology 2010, 35, 4–26. [Google Scholar] [CrossRef]
- Oldham, S.; Murawski, C.; Fornito, A.; Youssef, G.; Yücel, M.; Lorenzetti, V. The anticipation and outcome phases of reward and loss processing: A neuroimaging meta-analysis of the monetary incentive delay task. Hum. Brain Mapp. 2018, 39, 3398–3418. [Google Scholar] [CrossRef]
- Curran, H.V.; Freeman, T.; Mokrysz, C.; Lewis, D.; Morgan, C.J.A.; Parsons, L.H. Keep off the grass? Cannabis, cognition and addiction. Nat. Rev. Neurosci. 2016, 17, 293–306. [Google Scholar] [CrossRef]
- Ishiguro, H.; Horiuchi, Y.; Ishikawa, M.; Koga, M.; Imai, K.; Suzuki, Y.; Morikawa, M.; Inada, T.; Watanabe, Y.; Takahashi, M.; et al. Brain Cannabinoid CB2 Receptor in Schizophrenia. Biol. Psychiatry 2010, 67, 974–982. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.R.; Madularu, D.; Ortiz, R.J.; Athanassiou, M.; Knudsen, A.; Alkislar, I.; Cai, X.; Kulkarni, P.P.; Cushing, B.S.; Ferris, C.F. Changes in brain structure and function following chronic exposure to inhaled vaporised cannabis during periadolescence in female and male mice: A multimodal MRI study. Addict. Biol. 2022, 27, e13169. [Google Scholar] [CrossRef] [PubMed]
- Spindle, T.R.; Kuwabara, H.; Eversole, A.; Nandi, A.; Vandrey, R.; Antoine, D.G.; Umbricht, A.; Guarda, A.S.; Wong, D.F.; Weerts, E.M. Brain imaging of cannabinoid type I (CB 1) receptors in women with cannabis use disorder and male and female healthy controls. Addict. Biol. 2021, 26, e13061. [Google Scholar] [CrossRef]
- Basavarajappa, B.S.; Subbanna, S. Molecular Insights into Epigenetics and Cannabinoid Receptors. Biomolecules 2022, 12, 1560. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Li, C.; Jaffe, A.E.; Shin, J.H.; Deep-Soboslay, A.; Yamin, R.; Weinberger, D.R.; Hyde, T.M.; Kleinman, J.E. Cannabinoid receptor CNR1 expression and DNA methylation in human prefrontal cortex, hippocampus and caudate in brain development and schizophrenia. Transl. Psychiatry 2020, 10, 158. [Google Scholar] [CrossRef]
- Kebir, O.; the ICAAR team; Chaumette, B.; Rivollier, F.; Miozzo, F.; Perreault, L.P.L.; Barhdadi, A.; Provost, S.; Plaze, M.; Bourgin, J.; et al. Methylomic changes during conversion to psychosis. Mol. Psychiatry 2017, 22, 512–518. [Google Scholar] [CrossRef]
- Kay, S.R.; Flszbeln, A.; Qpjer, L.A. The Positive and Negative Syndrome Scale (PANSS) for Schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef]
- Wallwork, R.; Fortgang, R.; Hashimoto, R.; Weinberger, D.; Dickinson, D. Searching for a consensus five-factor model of the Positive and Negative Syndrome Scale for schizophrenia. Schizophr. Res. 2012, 137, 246–250. [Google Scholar] [CrossRef]
- Gomar, J.J.; Ortiz-Gil, J.; McKenna, P.J.; Salvador, R.; Sans-Sansa, B.; Sarró, S.; Guerrero, A.; Pomarol-Clotet, E. Validation of the Word Accentuation Test (TAP) as a means of estimating premorbid IQ in Spanish speakers. Schizophr. Res. 2011, 128, 175–176. [Google Scholar] [CrossRef]
- Gevins, A.; Cutillo, B. Spatiotemporal dynamics of component processes in human working memory. Electroencephalogr. Clin. Neurophysiol. 1993, 87, 128–143. [Google Scholar] [CrossRef]
- Green, D.M.; Swets, J.A. Signal Detection Theory and Psychophysics; John Wiley: Oxford, UK, 1966. [Google Scholar]
- Egli, T.; Coynel, D.; Spalek, K.; Fastenrath, M.; Freytag, V.; Heck, A.; Loos, E.; Auschra, B.; Papassotiropoulos, A.; De Quervain, D.J.-F.; et al. Identification of Two Distinct Working Memory-Related Brain Networks in Healthy Young Adults. eNeuro 2018, 5, ENEURO.0222-17.2018. [Google Scholar] [CrossRef]
- Hill, S.Y.; Sharma, V.; Jones, B.L. Lifetime use of cannabis from longitudinal assessments, cannabinoid receptor (CNR1) variation, and reduced volume of the right anterior cingulate. Psychiatry Res. Neuroimaging 2016, 255, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Squassina, A.; Congiu, D.; Chillotti, C.; Niola, P.; Galderisi, S.; Pistis, M.; Del Zompo, M. Investigation of endocannabinoid system genes suggests association between peroxisome proliferator activator receptor-α gene (PPARA) and schizophrenia. Eur. Neuropsychopharmacol. 2013, 23, 749–759. [Google Scholar] [CrossRef]
- Hamdani, N.; Tabeze, J.-P.; Ramoz, N.; Ades, J.; Hamon, M.; Sarfati, Y.; Boni, C.; Gorwood, P. The CNR1 gene as a pharmacogenetic factor for antipsychotics rather than a susceptibility gene for schizophrenia. Eur. Neuropsychopharmacol. 2008, 18, 34–40. [Google Scholar] [CrossRef]
- Gerra, M.C.; Jayanthi, S.; Manfredini, M.; Walther, D.; Schroeder, J.; Phillips, K.A.; Cadet, J.L.; Donnini, C. Gene variants and educational attainment in cannabis use: Mediating role of DNA methylation. Transl. Psychiatry 2018, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.J.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 2004, 23 (Suppl. 1), S208–S219. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cannabis Non-Users (CNU) | Cannabis Users (CU) | CNU vs. CU Comparison p-Value | |
|---|---|---|---|
| N | 20 | 20 | - |
| Age (years) | 26.19 (7.72) | 25.94 (5.31) | 0.87 |
| Sex (male/female) | 15/5 | 16/4 | 0.71 |
| Premorbid IQ (TAP a) | 97.24 (11.29) | 98.78 (7.95) | 0.64 |
| PANSS scale | |||
| PANSS positive | 15.90 (5.86) | 18.45 (4.80) | 0.14 |
| PANSS negative | 16.20 (8.46) | 16.95 (8.34) | 0.78 |
| PANSS general | 30.25 (7.60) | 33.85 (8.92) | 0.18 |
| GAF b | 52.11 (11.11) | 48.74 (10.94) | 0.36 |
| CPZ c equivalents (mg/day) | 292.65 (145.16) | 301.20 (181.88) | 0.64 |
| Gene (Polymorphism) | Genotypes | Cannabis Non-Users (CNU) | Cannabis Users (CU) | CNU vs. CU Comparison χ2 Test, p-Value |
|---|---|---|---|---|
| CNR1 (rs1049353) | CC | 14 (70%) | 12 (60%) | 0.440, 0.741 |
| Tcar | 6 (30%) | 8 (40%) | ||
| CNR2 (rs2501431) | AA | 6 (30%) | 9 (45%) | 0.960, 0.257 |
| Gcar | 14 (70%) | 11 (55%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oscoz-Irurozqui, M.; Almodóvar-Payá, C.; Guardiola-Ripoll, M.; Guerrero-Pedraza, A.; Hostalet, N.; Salvador, R.; Carrión, M.I.; Maristany, T.; Pomarol-Clotet, E.; Fatjó-Vilas, M. Cannabis Use and Endocannabinoid Receptor Genes: A Pilot Study on Their Interaction on Brain Activity in First-Episode Psychosis. Int. J. Mol. Sci. 2023, 24, 7501. https://doi.org/10.3390/ijms24087501
Oscoz-Irurozqui M, Almodóvar-Payá C, Guardiola-Ripoll M, Guerrero-Pedraza A, Hostalet N, Salvador R, Carrión MI, Maristany T, Pomarol-Clotet E, Fatjó-Vilas M. Cannabis Use and Endocannabinoid Receptor Genes: A Pilot Study on Their Interaction on Brain Activity in First-Episode Psychosis. International Journal of Molecular Sciences. 2023; 24(8):7501. https://doi.org/10.3390/ijms24087501
Chicago/Turabian StyleOscoz-Irurozqui, Maitane, Carmen Almodóvar-Payá, Maria Guardiola-Ripoll, Amalia Guerrero-Pedraza, Noemí Hostalet, Raymond Salvador, Maria Isabel Carrión, Teresa Maristany, Edith Pomarol-Clotet, and Mar Fatjó-Vilas. 2023. "Cannabis Use and Endocannabinoid Receptor Genes: A Pilot Study on Their Interaction on Brain Activity in First-Episode Psychosis" International Journal of Molecular Sciences 24, no. 8: 7501. https://doi.org/10.3390/ijms24087501
APA StyleOscoz-Irurozqui, M., Almodóvar-Payá, C., Guardiola-Ripoll, M., Guerrero-Pedraza, A., Hostalet, N., Salvador, R., Carrión, M. I., Maristany, T., Pomarol-Clotet, E., & Fatjó-Vilas, M. (2023). Cannabis Use and Endocannabinoid Receptor Genes: A Pilot Study on Their Interaction on Brain Activity in First-Episode Psychosis. International Journal of Molecular Sciences, 24(8), 7501. https://doi.org/10.3390/ijms24087501