
Single Copies of the 5S rRNA Inserted into 45S rDNA Intergenic Spacers in the Genomes of Nototheniidae (Perciformes, Actinopterygii)


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
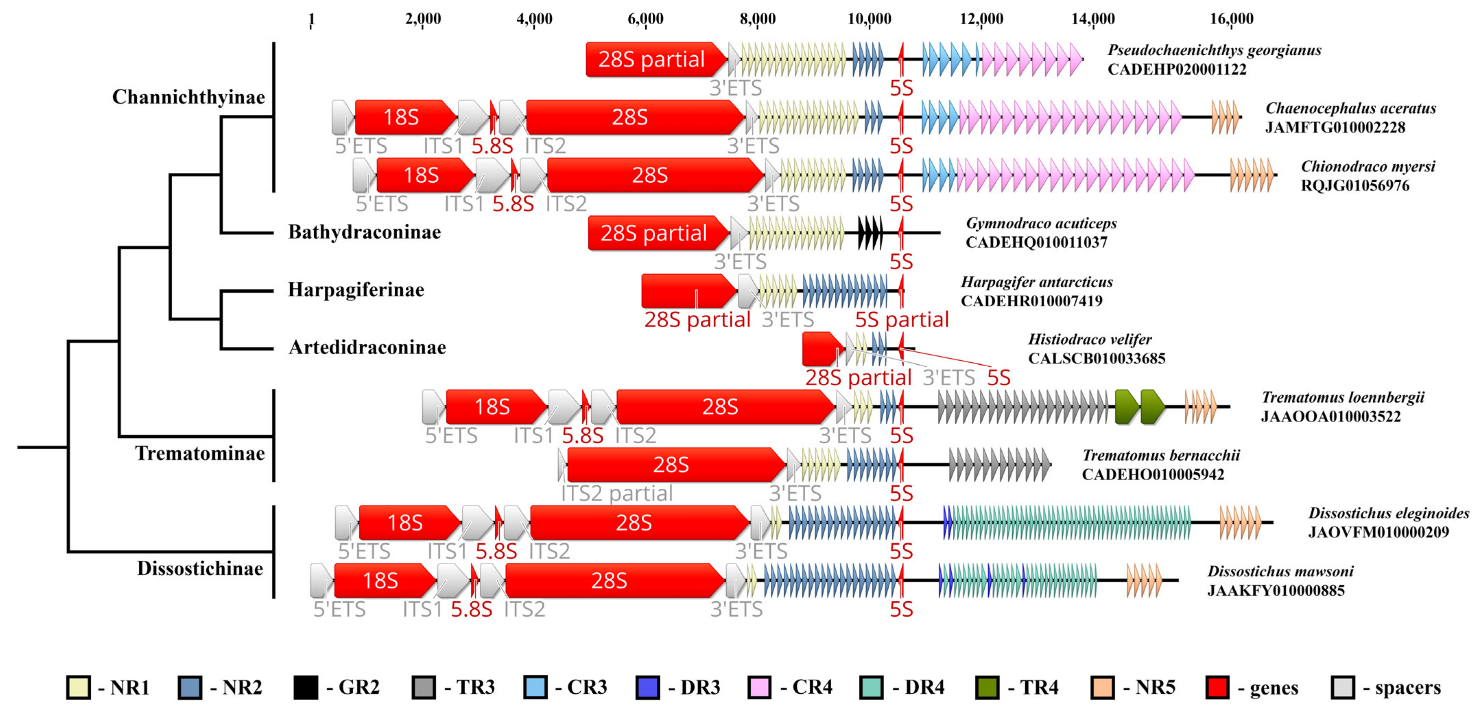
2. Results
2.1. 5S rDNA Distribution in the Genomes of Notothens
2.2. 5S rRNA Types and Their Expression
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Douet, J.; Tourmente, S. Transcription of the 5S rRNA heterochromatic genes is epigenetically controlled in Arabidopsis thaliana and Xenopus laevis. Heredity 2007, 99, 5–13. [Google Scholar] [CrossRef]
- Ciganda, M.; Williams, N. Eukaryotic 5S rRNA biogenesis. Wiley Interdiscip. Rev. RNA 2011, 2, 523–533. [Google Scholar] [CrossRef]
- Vierna, J.; Wehner, S.; Hӧner, Z.; Siederdissen, C.; Martínez-Lage, A.; Marz, M. Systematic analysis and evolution of 5S ribsomal DNA in metazoans. Heredity 2013, 111, 410–421. [Google Scholar] [CrossRef]
- Symonová, R. Integrative rDNAomics–importance of the oldest repetitive fraction of the eukaryote genome. Genes 2019, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, A.V.; Entelis, N.S.; Krasheninnikov, I.A.; Martin, R.; Tarassov, I.A. Specific features of 5S rRNA structure–Its interactions with macromolecules and possible functions. Biochemistry 2008, 73, 1418–1437. [Google Scholar] [CrossRef]
- Caburet, S.; Conti, C.; Schurra, C.; Lebofsky, R.; Edelstein, S.J.; Bensimon, A. Human ribosomal RNA gene arrays display a broad range of palindromic structures. Genome Res. 2005, 15, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Stults, D.M.; Killen, M.V.; Pierce, H.H.; Pierce, A.J. Genomic architecture and inheritance of human ribosomal RNA gene clusters. Genome Res. 2008, 18, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, J.G.; Branco, A.T.; Yu, S.; Lemos, B. Ribosomal DNA copy number is coupled with gene expression variation and mitochondrial abundance in humans. Nat. Commun. 2014, 11, 4850. [Google Scholar] [CrossRef]
- Gibbons, J.G.; Branco, A.T.; Godinho, S.A.; Yu, S.; Lemos, B. Concerted copy number variation balances ribosomal DNA dosage in human and mouse genomes. Proc. Natl. Acad. Sci. USA 2015, 112, 2485–2490. [Google Scholar] [CrossRef]
- Hori, Y.; Shimamoto, A.; Kobayashi, T. The human ribosomal DNA array is composed of highly homogenised tandem clusters. Genome Res. 2021, 31, 1971–1982. [Google Scholar] [CrossRef]
- McStay, B.; Grummt, I. The epigenetics of rRNA genes: From molecular to chromosome biology. Annu. Rev. Cell Dev. Biol. 2008, 24, 131–157. [Google Scholar] [CrossRef]
- Zillner, K.; Komatsu, J.; Kalepu, R.; Bensimon, A.; Németh, A. Active human nucleolar organizer regions are interspersed with inactive rDNA repeats in normal and tumor cells. Epigenomics 2015, 7, 363–378. [Google Scholar] [CrossRef]
- Vincent, W.S.; Halvorson, H.O.; Chen, H.-R.; Shin, D. A comparison of ribosomal gene amplification in uni- and multinucleolate oocytes. Exp. Cell Res. 1969, 57, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Raikova, E.V. Evolution of the nucleolar apparatus during oogenesis in Acipenseridae. J. Embryol. Exp. Morphol. 1976, 35, 667–687. [Google Scholar] [CrossRef] [PubMed]
- Locati, M.D.; Pagano, J.F.B.; Ensink, W.A.; van Olst, M.; van Leeuwen, S.; Nehrdich, U.; Zhu, K.; Spaink, H.P.; Girard, G.; Rauwerda, H.; et al. Linking maternal and somatic 5S rRNA types with different sequence-specific non-LTR retrotransposons. RNA 2017, 23, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.G. Differential synthesis of the genes for rRNA during amphibian oogenesis. Proc. Natl. Acad. Sci. USA 1968, 60, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Macgregor, H.C. Nucleolar DNA in oocytes of Xenopus laevis. J. Cell Sci. 1968, 3, 437–444. [Google Scholar] [CrossRef]
- Wegnez, M.; Monier, R.; Denis, H. Sequence heterogeneity of 5S RNA in Xenopus laevis. FEBS Lett. 1972, 25, 13–20. [Google Scholar] [CrossRef]
- Peterson, R.C.; Doering, J.L.; Brown, D.D. Characterization of two Xenopus somatic 5S DNAs and one minor oocyte-specific 5S DNA. Cell 1980, 20, 131–141. [Google Scholar] [CrossRef]
- Wormington, W.M.; Brown, D.D. Onset of 5S RNA gene regulation during Xenopus embryogenesis. Dev. Biol. 1983, 99, 248–257. [Google Scholar] [CrossRef]
- Macgregor, H.C. Ways of amplifying ribosomal genes. In The Nucleolus; Jordan, E.G., Cullis, C.A., Eds.; Cambridge University Press: Cambridge, UK, 1982; pp. 129–151. [Google Scholar]
- Callebaut, M.; van Nassauw, L.; Harrisson, F. Comparison between oogenesis and related ovarian structures in a reptile, Pseudemys scripta elegans (turtle) and in a bird Coturnix coturnix japonica (quail). Reprod. Nutr. Dev. 1997, 37, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Davidian, A.; Koshel, E.; Dyomin, A.; Galkina, S.; Saifitdinova, A.; Gaginskaya, E. On some structural and evolutionary aspects of rDNA amplification in oogenesis of Trachemys scripta turtles. Cell Tissue Res. 2021, 383, 853–864. [Google Scholar] [CrossRef]
- Uribe, M.C.A.; Guillette, L.J. Oogenesis and ovarian histology of the American alligator Alligator mississippiensis. J. Morphol. 2000, 245, 225–240. [Google Scholar] [CrossRef]
- Moore, B.C.; Uribe-Aranza, M.C.; Boggs, A.S.P.; Guillette, L.J., Jr. Developmental morphology of the neonatal alligator (Aligator mississippiensis) ovary. J. Morphol. 2008, 269, 302–312. [Google Scholar] [CrossRef]
- Ford, P.J.; Southern, E.M. Different sequences for 5S RNA in kidney cells and ovaries of Xenopus laevis. Nat. New Biol. 1973, 241, 7–12. [Google Scholar] [CrossRef]
- Long, E.O.; Dawid, I.B. Repeated genes in eukaryotes. Annu. Rev. Biochem. 1980, 49, 727–764. [Google Scholar] [CrossRef] [PubMed]
- Korn, L.J. Transcription of Xenopus 5S ribosomal RNA genes. Nature 1982, 295, 101–105. [Google Scholar] [CrossRef]
- Dennis, H.; Wegnez, M. Biochemical research on oogenesis: Oocytes of Xenopus laevis synthesize but do not accumulate 5S RNA of somatic type. Dev. Biol. 1977, 58, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.E.; Price, J.; Korn, L.J. Chromosomal mapping of Xenopus 5S genes: Somatic-type versus oocyte-type. Nucleic Acids Res. 1983, 11, 2313–2323. [Google Scholar] [CrossRef]
- Wormington, W.M.; Schlissel, M.; Brown, D.D. Developmental regulation of Xenopus 5S RNA genes. Cold Spring Harb. Symp. Quant. Biol. 1982, 47 Pt 2, 879–884. [Google Scholar] [CrossRef]
- Guinta, D.R.; Yun Tso, J.; Narayanswami, S.; Hamkalo, B.A.; Korn, L.J. Early replication and expression of oocyte-type 5S RNA genes in a Xenopus somatic cell line carrying a translocation. Proc. Natl. Acad. Sci. USA 1986, 83, 5150–5154. [Google Scholar] [CrossRef]
- Dennis, H.; Wegnez, M. Oocytes and liver cells of the teleost fish Tinca tinca contain different kinds of 5S rRNA. Devl. Biol. 1977, 59, 228–236. [Google Scholar]
- Mashkova, T.D.; Serenkova, T.I.; Mazo, A.M.; Avdonina, T.A.; Timofeyeva, M.Y.; Kisselev, L.L. The primary structure of oocyte and somatic SS rRNAs from the loach Misgurnus fossilis. Nucleic Acids Res. 1981, 9, 2141–2151. [Google Scholar] [CrossRef]
- Martins, C.; Wasko, A.P. Organization and evolution of 5S ribosomal DNA in the fish genome. In Focus on Genome Research; Williams, C.R., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2004; pp. 335–363. [Google Scholar]
- Martins, C.; Galetti, P.M., Jr. Organization of 5S rDNA in species of the fish Leporinus: Two different genomic locations are characterized by distinct nontranscribed spacers. Genome 2001, 44, 903–910. [Google Scholar] [CrossRef]
- Martins, C.; Wasko, A.P.; Oliveira, C.; Porto-Foresti, F.; Parise-Maltempi, P.P.; Wright, J.M.; Foresti, F. Dynamics of 5S rDNA in the tilapia (Oreochromis niloticus) genome: Repeat units, inverted sequences, pseudogenes and chromosome loci. Cytogenet. Genome Res. 2002, 98, 78–85. [Google Scholar] [CrossRef]
- Davidian, A.G.; Dyomin, A.G.; Galkina, S.A.; Makarova, N.E.; Dmitriev, S.E.; Gaginskaya, E.R. 45S rDNA repeats of turtles and crocodiles harbor a functional 5S rRNA gene specifically expressed in oocytes. Mol. Biol. Evol. 2022, 39, msab324. [Google Scholar] [CrossRef]
- Almeida-Toledo, L.; Ozouf-Costaz, C.; Foresti, F.; Bonillo, C.; Porto-Foresti, F.; Daniel-Silva, M. Conservation of the 5S-bearing chromosome pair and co-localization with major rDNA clusters in five species of Astyanax (Pisces, Characidae). Cytogenet. Genome Res. 2002, 97, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Fontana, F.; Lanfredi, M.; Congiu, L.; Leis, M.; Chicca, M.; Rossi, R. Chromosomal mapping of 18S–28S and 5S rRNA genes by two-colour fluorescent in situ hybridization in six sturgeon species. Genome 2003, 46, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Pisano, E.; Ghigliotti, L. Ribosomal genes in notothenioid fishes: Focus on the chromosomal organisation. Mar Genom. 2009, 2, 75–80. [Google Scholar] [CrossRef]
- Near, T.J.; Dornburg, A.; Kuhn, K.L.; Eastman, J.T.; Pennington, J.N.; Patarnello, T.; Zane, L.; Fernández, D.A.; Jones, C.D. Ancient climate change, antifreeze, and the evolutionary diversification of Antarctic fishes. Proc. Natl. Acad. Sci. USA 2012, 109, 3434–3439. [Google Scholar] [CrossRef] [PubMed]
- Duhamel, G.; Hulley, P.-A.; Causse, R.; Koubbi, P.; Vacchi, M.; Pruvost, P. Biogeographic patterns of fish. In Biogeographic atlas of the Southern Ocean; De Broyer, C., Koubbi, P., Griffiths, H.J., Raymond, B., Udekem d’Acoz, C., Van de Putte, A.P., Danis, B., David, B., Grant, S., Gutt, J., et al., Eds.; Scentific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 328–362. [Google Scholar]
- Auvinet, J.; Graça, P.; Dettai, A.; Amores, A.; Postlethwait, J.H.; Detrich, H.W.; Ozouf-Costaz, C.; Coriton, O.; Higuet, D. Multiple independent chromosomal fusions accompanied the radiation of the Antarctic teleost genus Trematomus (Notothenioidei: Nototheniidae). BMC Evol. Biol. 2020, 20, 39. [Google Scholar] [CrossRef]
- Wolffe, A.P. The role of transcription factors, chromatin structure and DNA replication in 5S RNA gene regulation. J. Cell Sci. 1994, 107, 2055–2063. [Google Scholar] [CrossRef] [PubMed]
- Andronescu, M.; Condon, A.; Hoos, H.H.; Mathews, D.H.; Murphy, K.P. Efficient parameter estimation for RNA secondary structure prediction. Bioinformatics 2007, 23, i19–i28. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.T.; DeVries, A.L. Aspects of body size and gonadal histology in the Antarctic toothfish, Dissostichus mawsoni, from McMurdo Sound, Antarctica. Polar Biol. 2000, 23, 189–195. [Google Scholar] [CrossRef]
- Parker, S.J.; Grimes, P.J. Length- and age-at-spawning of Antarctic Toothfish (Dissostichus mawsoni) in the Ross Sea. CCAMLR Sci. 2010, 17, 53–73. [Google Scholar]
- Piyanova, S.; Kokorin, N. Cytomorphology of female and male gonads in Antarctic toothfish Dissosctichus mawsoni (Nototheniidae) from the Ross Sea in the summer period. J. Ichthyol. 2010, 50, 374–385. [Google Scholar] [CrossRef]
- Davidson, E.H. Gene Activity in Early Development, 3rd ed.; Academic Press: New York, NY, USA, 1986. [Google Scholar]
- Leesch, F.; Lorenzo-Orts, L.; Pribitzer, C.; Grishkovskaya, I.; Roehsner, J.; Chugunova, A.; Matzinger, M.; Roitinger, E.; Belačić, K.; Kandolf, S.; et al. A molecular network of conserved factors keeps ribosomes dormant in the egg. Nature 2023, 613, 712–720. [Google Scholar] [CrossRef]
- Shiokawa, K.; Misumi, Y.; Yamana, K. Demonstration of rRNA synthesis in pre-gastrular embryos of Xenopus laevis. Dev. Growth Differ. 1981, 23, 579–587. [Google Scholar] [CrossRef]
- Pardue, M.L.; Brown, D.D.; Birnstiel, M.L. Location of the genes for 5S ribosomal RNA in Xenopus laevis. Chromosoma 1973, 42, 191–203. [Google Scholar] [CrossRef]
- Kramer, R.A.; Philippsen, P.; Davis, R.W. Divergent transcription in the yeast ribosomal RNA coding region as shown by hybridization to separated strands and sequence analysis of cloned DNA. J. Mol. Biol. 1978, 123, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Garcia, S.; Kovarïk, A.; Leitch, A.R.; Garnatje, T. Cytogenetic features of rRNA genes across land plants: Analysis of the plant rDNA database. Plant J. 2017, 89, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Vahidi, H.; Curran, J.; Nelson, D.W.; Webster, J.M.; McClure, M.A.; Honda, B.M. Unusual sequences, homologous to 5S RNA, in ribosomal DNA repeats of the nematode Meloidogyne arenaria. J. Mol. Evol. 1988, 27, 222–227. [Google Scholar] [CrossRef]
- Drouin, G. Homogenization of 5S ribosomal genes on the noncoding strand of the rDNA units of two crustacean species. Genome 1999, 42, 150–153. [Google Scholar] [CrossRef]
- Weydmann, A.; Przyłucka, A.; Lubośny, M.; Walczyńska, K.S.; Serrão, E.A.; Pearson, G.A.; Burzyński, A. Mitochondrial genomes of the key zooplankton copepods Arctic Calanus glacialis and North Atlantic Calanus finmarchicus with the longest crustacean non-coding regions. Sci. Rep. 2017, 7, 13702. [Google Scholar] [CrossRef] [PubMed]
- McElroy, K.E.; Müller, S.; Lamatsch, D.K.; Bankers, L.; Fields, P.D.; Jalinsky, J.R.; Sharbrough, J.; Boore, J.L.; Logsdon, J.M.; Neiman, M. Asexuality associated with marked genomic expansion of tandemly repeated rRNA and histone genes. Mol. Biol. Evol. 2021, 38, msab121. [Google Scholar] [CrossRef]
- Eickbush, T.H.; Eickbush, D.G. Finely orchestrated movements: Evolution of the ribosomal RNA genes. Genetics 2007, 175, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Auvinet, J.; Graça, P.; Ghigliotti, L.; Pisano, E.; Dettaï, A.; Ozouf-Costaz, C.; Higuet, D. Insertion hot spots of DIRS1 retrotransposon and chromosomal diversifications among the Antarctic Teleosts Nototheniidae. Int. J. Mol. Sci. 2019, 20, 701. [Google Scholar] [CrossRef]
- Simões, T.R.; Kammerer, C.F.; Caldwell, M.W.; Pierce, S.E. Successive climate crises in the deep past drove the early evolution and radiation of reptiles. Sci. Adv. 2022, 8, eabq1898. [Google Scholar] [CrossRef] [PubMed]
- Tomaszkiewicz, M.; Hautecoeur, M.; Coutanceau, J.-P.; Bonillo, C.; Dettaï, A.; Mazzei, F.; Ghigliotti, L.; Pisano, E.; Couloux, A.; Chanet, B.; et al. Comparative cytogenetic studies of the Nototheniidae (Teleostei: Acanthomorpha) from the Indian (Kerguelen-Heard plateau) and Atlantic (South Georgia, South Sandwich, Falklands/Malvinas, Bouvet Islands) sectors of the Southern Ocean. In The Kerguelen Plateau Marine Ecosystem and Fisheries; Société Française d’Ichtyologie; Duhamel, G., Welsford, D., Eds.; Australian Antarctic Division: Hobart, TAS, Australia, 2011; Volume 35, pp. 109–120. [Google Scholar]
- Ghigliotti, L.; Mazzei, F.; Ozouf-Costaz, C.; Bonillo, C.; Williams, R.; Cheng, C.; Pisano, E. The two giant sister species of the Southern Ocean, Dissostichus eleginoides and Dissostichus mawsoni, differ in karyotype and chromosomal pattern of ribosomal RNA genes. Polar Biol. 2007, 30, 625–634. [Google Scholar] [CrossRef]
- Mazzei, F.; Ghigliotti, L.; Lecointre, G.; Ozouf-Costaz, C.; Coutanceau, J.; Detrich, W.; Pisano, E. Karyotypes of basal lineages in notothenioid fishes: The genus Bovichtus. Polar Biol. 2006, 29, 1071–1076. [Google Scholar] [CrossRef]
- Dyomin, A.G.; Koshel, E.I.; Kiselev, A.M.; Saifitdinova, A.F.; Galkina, S.A.; Fukagawa, T.; Gaginskaya, E. Chicken rRNA gene cluster structure. PLoS ONE 2016, 11, e0157464. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.-I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dyomin, A.; Galkina, S.; Ilina, A.; Gaginskaya, E. Single Copies of the 5S rRNA Inserted into 45S rDNA Intergenic Spacers in the Genomes of Nototheniidae (Perciformes, Actinopterygii). Int. J. Mol. Sci. 2023, 24, 7376. https://doi.org/10.3390/ijms24087376
Dyomin A, Galkina S, Ilina A, Gaginskaya E. Single Copies of the 5S rRNA Inserted into 45S rDNA Intergenic Spacers in the Genomes of Nototheniidae (Perciformes, Actinopterygii). International Journal of Molecular Sciences. 2023; 24(8):7376. https://doi.org/10.3390/ijms24087376
Chicago/Turabian StyleDyomin, Alexander, Svetlana Galkina, Arina Ilina, and Elena Gaginskaya. 2023. "Single Copies of the 5S rRNA Inserted into 45S rDNA Intergenic Spacers in the Genomes of Nototheniidae (Perciformes, Actinopterygii)" International Journal of Molecular Sciences 24, no. 8: 7376. https://doi.org/10.3390/ijms24087376
APA StyleDyomin, A., Galkina, S., Ilina, A., & Gaginskaya, E. (2023). Single Copies of the 5S rRNA Inserted into 45S rDNA Intergenic Spacers in the Genomes of Nototheniidae (Perciformes, Actinopterygii). International Journal of Molecular Sciences, 24(8), 7376. https://doi.org/10.3390/ijms24087376