
Differences in Bacterial Small RNAs in Stool Samples from Hypercholesterolemic and Normocholesterolemic Subjects

 ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Anthropometric, Clinical and Biochemical Parameters
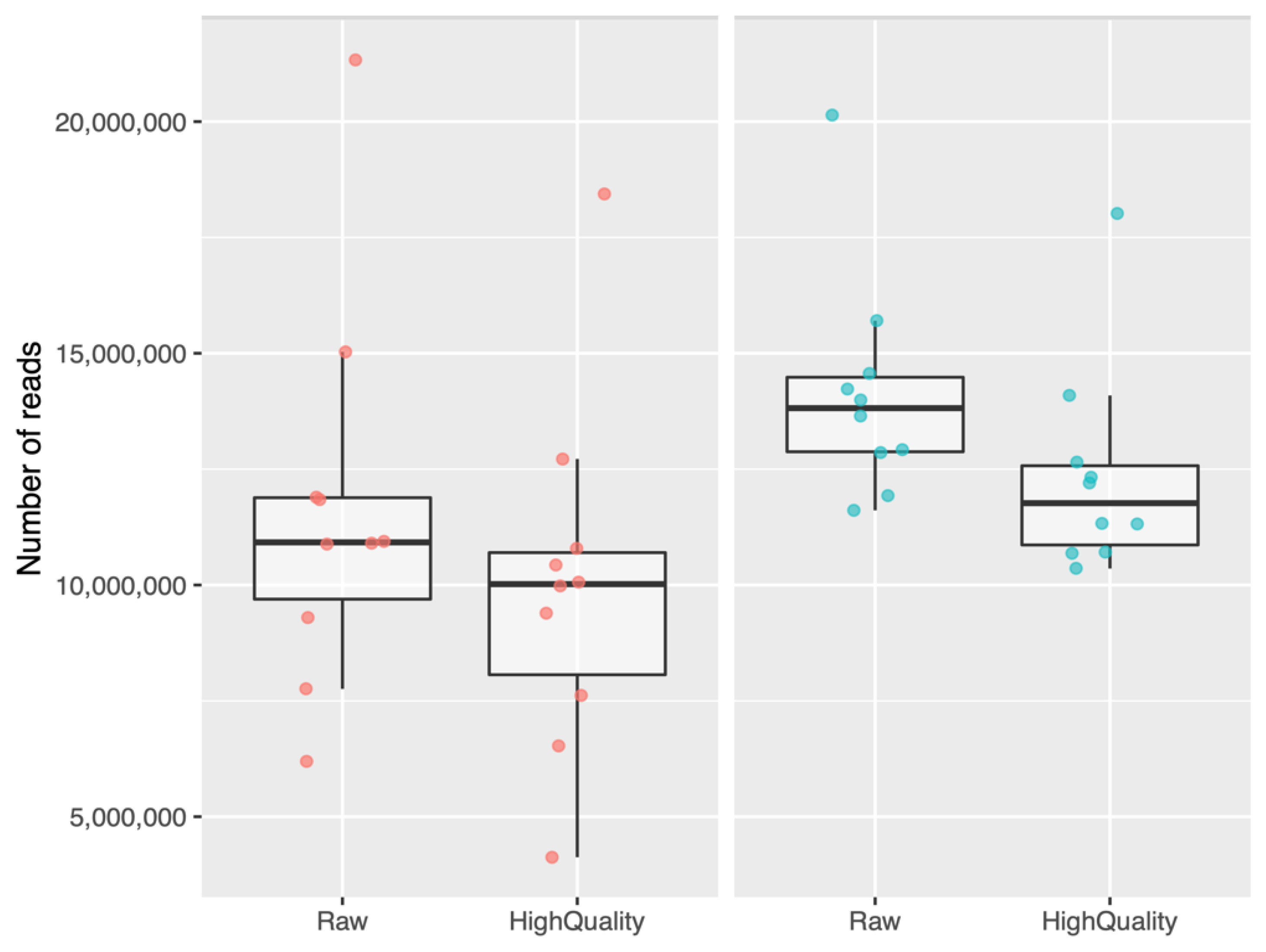
2.2. Bioinformatic Analysis of Small RNAs Sequences
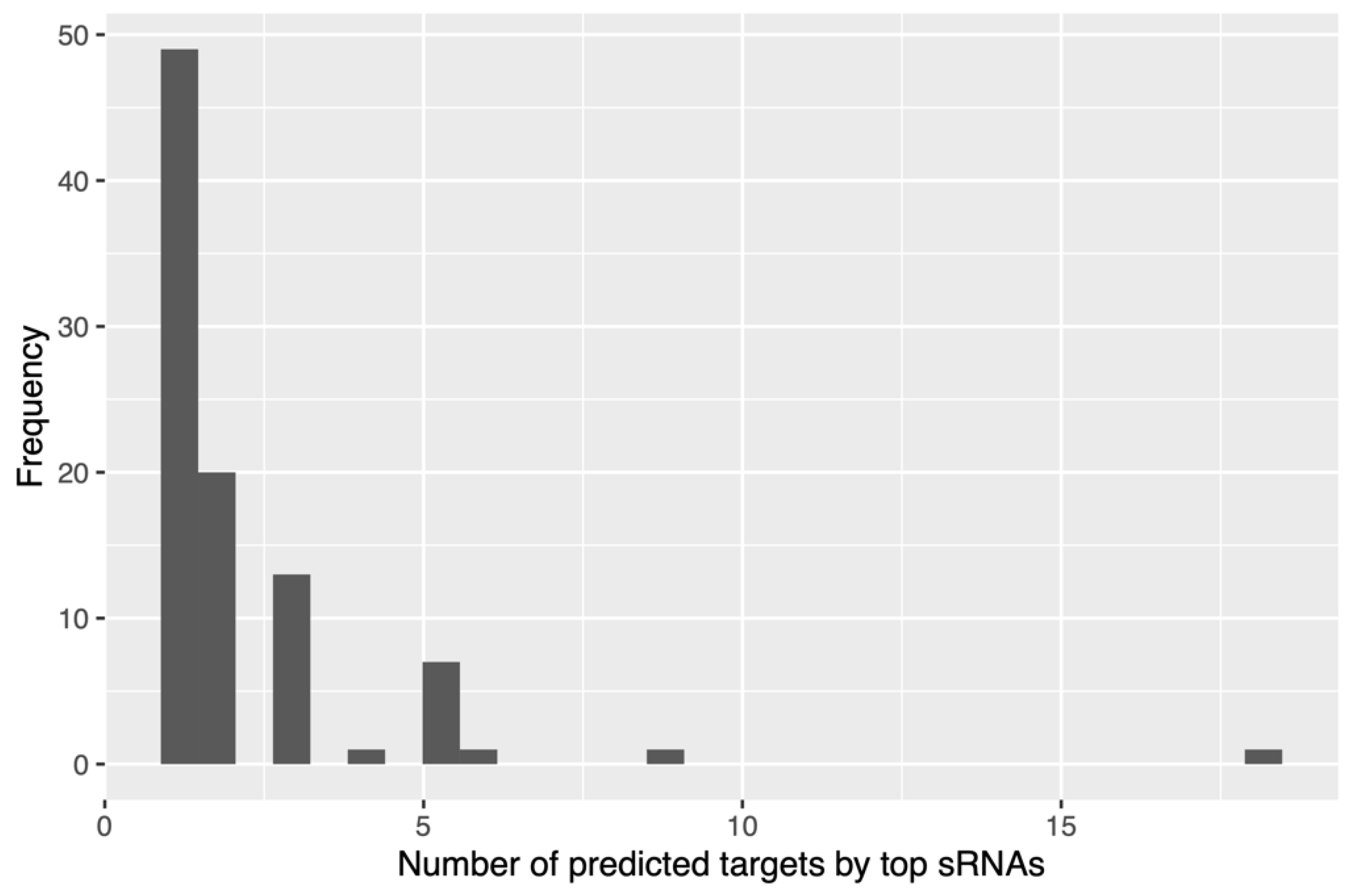
2.3. Bioinformatic Analysis of Small RNA Sequences
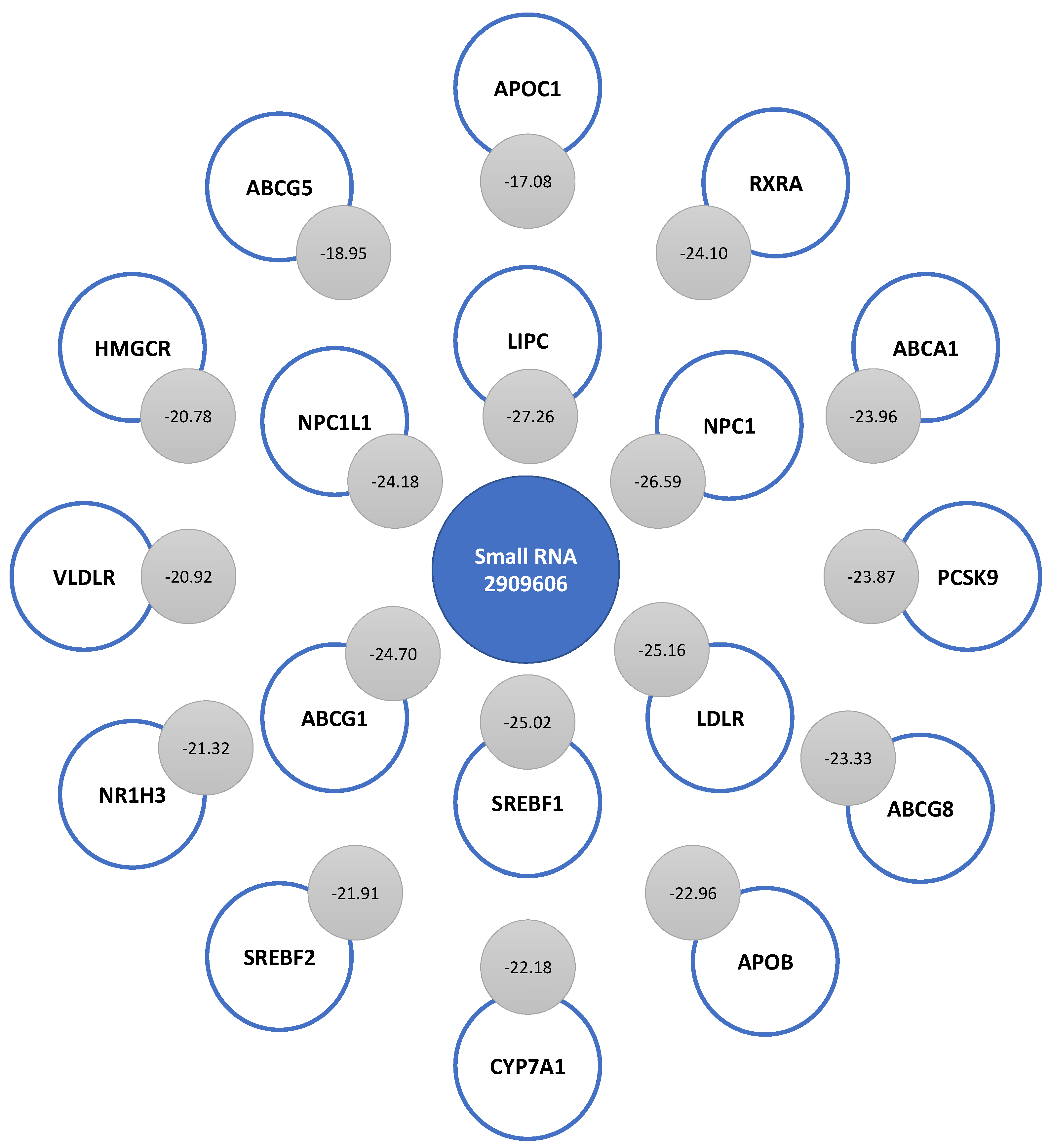
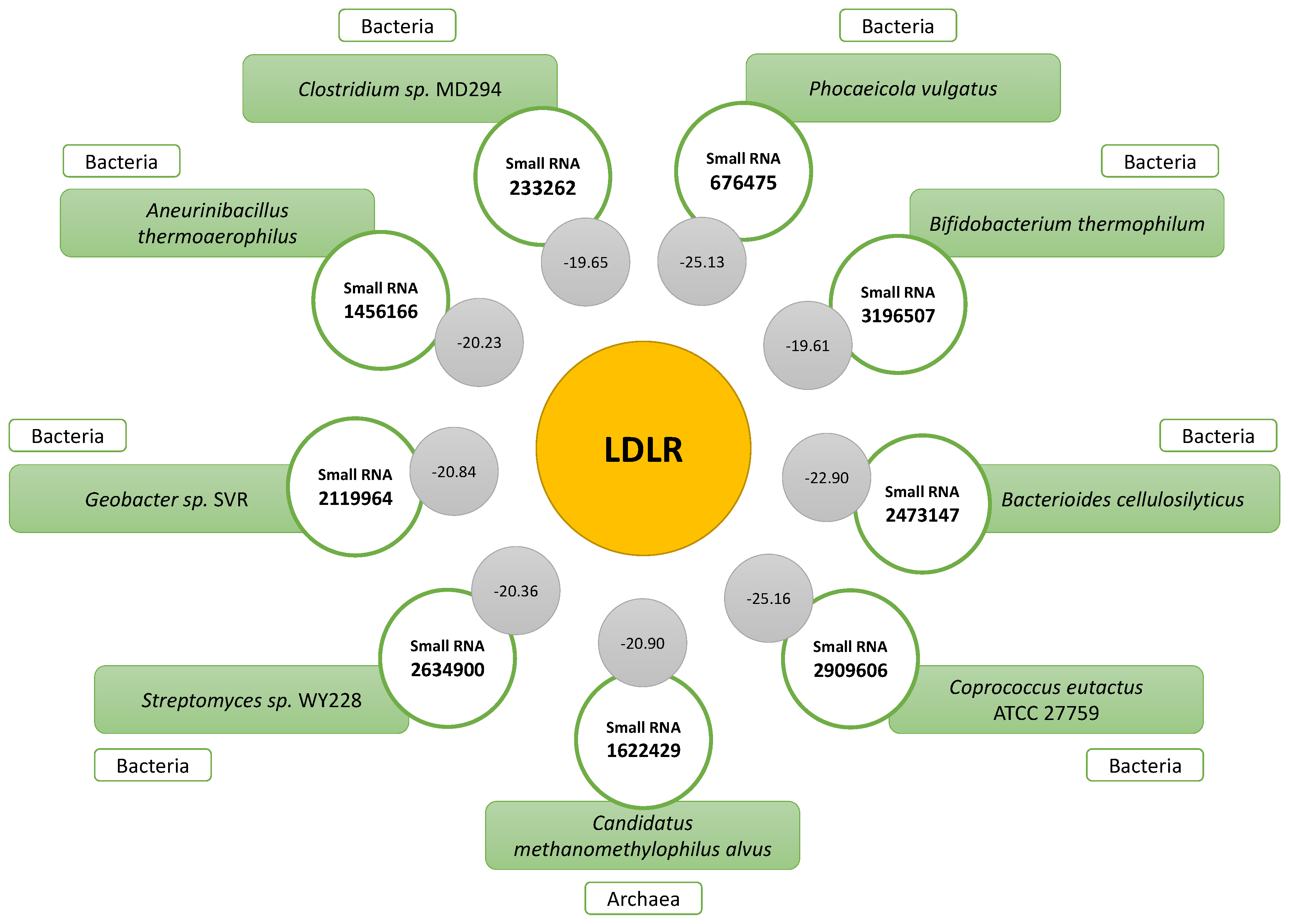
2.4. Prediction of the Secondary Structure of Small RNAs That Bind to LDLR
3. Discussion
4. Materials and Methods
4.1. Subjects and Samples
4.2. Anthropometric, Clinical, and Biochemical Parameters
4.3. RNA Extraction and Sequencing
4.4. Bioinformatic Analysis
4.5. Secondary Structure Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luo, J.; Yang, H.; Song, B.L. Mechanisms and regulation of cholesterol homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 225–245. [Google Scholar] [CrossRef]
- Schoop, V.; Martello, A.; Eden, E.R.; Höglinger, D. Cellular cholesterol and how to find it. Biochim. Et Biophys. Acta. Mol. Cell Biol. Lipids 2021, 1866, 158989. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yutuc, E.; Griffiths, W.J. Cholesterol metabolism pathways - are the intermediates more important than the products? FEBS J. 2021, 288, 3727–3745. [Google Scholar] [CrossRef] [PubMed]
- Atar, D.; Jukema, J.W.; Molemans, B.; Taub, P.R.; Goto, S.; Mach, F.; CerezoOlmos, C.; Underberg, J.; Keech, A.; Tokgözoğlu, L.; et al. New cardiovascular prevention guidelines: How to optimally manage dyslipidaemia and cardiovascular risk in 2021 in patients needing secondary prevention? Atherosclerosis 2021, 319, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Pirillo, A.; Casula, M.; Olmastroni, E.; Norata, G.D.; Catapano, A.L. Global epidemiology of dyslipidaemias. Nat. Rev. Cardiol. 2021, 18, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, A.; Likos, E.M.; Weyman, C.M.; Shukla, G.C. Regulation of cholesterol biosynthesis and lipid metabolism: A microRNA management perspective. Steroids 2021, 173, 108878. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.W.; Huang, K.; Yang, C.; Kang, C.S. Non-coding RNAs as regulators in epigenetics (Review). Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef]
- Saw, P.E.; Xu, X.; Chen, J.; Song, E.W. Non-coding RNAs: The new central dogma of cancer biology. Sci. China Life Sci. 2021, 64, 22–50. [Google Scholar] [CrossRef]
- Shi, J.; Zhou, T.; Chen, Q. Exploring the expanding universe of small RNAs. Nat. Cell Biol. 2022, 24, 415–423. [Google Scholar] [CrossRef]
- Xiong, Q.; Zhang, Y.; Li, J.; Zhu, Q. Small Non-Coding RNAs in Human Cancer. Genes 2022, 13, 2072. [Google Scholar] [CrossRef]
- Zhu, L.; Ow, D.W.; Dong, Z. Transfer RNA-derived small RNAs in plants. Sci. China. Life Sci. 2018, 61, 155–161. [Google Scholar] [CrossRef]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.S.; Zhou, R.; Rana, T.M. Gene regulation by non-coding RNAs. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 16–32. [Google Scholar] [CrossRef]
- Burgos, M.; Hurtado, A.; Jiménez, R.; Barrionuevo, F.J. Non-Coding RNAs: lncRNAs, miRNAs, and piRNAs in Sexual Development. Sex. Dev. Genet. Mol. Biol. Evol. Endocrinol. Embryol. Pathol. Sex Determ. Differ. 2021, 15, 335–350. [Google Scholar] [CrossRef] [PubMed]
- El-Mogy, M.; Lam, B.; Haj-Ahmad, T.A.; McGowan, S.; Yu, D.; Nosal, L.; Rghei, N.; Roberts, P.; Haj-Ahmad, Y. Diversity and signature of small RNA in different bodily fluids using next generation sequencing. BMC Genom. 2018, 19, 408. [Google Scholar] [CrossRef] [PubMed]
- Tan, I.L.; Coutinho de Almeida, R.; Modderman, R.; Stachurska, A.; Dekens, J.; Barisani, D.; Meijer, C.R.; Roca, M.; Martinez-Ojinaga, E.; Shamir, R.; et al. Circulating miRNAs as Potential Biomarkers for Celiac Disease Development. Front. Immunol. 2021, 12, 734763. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Wu, H.; Zhong, W.; Zhang, Q.; Yu, Z. Expression profiling and bioinformatics analysis of circulating microRNAs in patients with acute myocardial infarction. J. Clin. Lab. Anal. 2020, 34, e23099. [Google Scholar] [CrossRef]
- Poller, W.; Dimmeler, S.; Heymans, S.; Zeller, T.; Haas, J.; Karakas, M.; Leistner, D.M.; Jakob, P.; Nakagawa, S.; Blankenberg, S.; et al. Non-coding RNAs in cardiovascular diseases: Diagnostic and therapeutic perspectives. Eur. Heart J. 2018, 39, 2704–2716. [Google Scholar] [CrossRef]
- Bansal, P.; Arora, M. Small Interfering RNAs and RNA Therapeutics in Cardiovascular Diseases. Adv. Exp. Med. Biol. 2020, 1229, 369–381. [Google Scholar] [CrossRef]
- Zeng, J.; Gupta, V.K.; Jiang, Y.; Yang, B.; Gong, L.; Zhu, H. Cross-Kingdom Small RNAs Among Animals, Plants and Microbes. Cells 2019, 8, 371. [Google Scholar] [CrossRef]
- Koeppen, K.; Hampton, T.H.; Jarek, M.; Scharfe, M.; Gerber, S.A.; Mielcarz, D.W.; Demers, E.G.; Dolben, E.L.; Hammond, J.H.; Hogan, D.A.; et al. A Novel Mechanism of Host-Pathogen Interaction through sRNA in Bacterial Outer Membrane Vesicles. PLoS Pathog. 2016, 12, e1005672. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Y.; Song, Z.; Li, R.; Ruan, H.; Liu, Q.; Huang, X. sncRNAs packaged by Helicobacter pylori outer membrane vesicles attenuate IL-8 secretion in human cells. Int. J. Med. Microbiol. IJMM 2020, 310, 151356. [Google Scholar] [CrossRef]
- Zhu, L.T.; Huang, H.N.; Avellán-Llaguno, R.D.; Qin, Y.; An, X.L.; Su, J.Q.; Huang, Q.; Zhu, Y.G. Diverse functional genes harboured in extracellular vesicles from environmental and human microbiota. J. Extracell. Vesicles 2022, 11, e12292. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Guan, L.L. Noncoding RNAs: Regulatory Molecules of Host-Microbiome Crosstalk. Trends Microbiol. 2021, 29, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Ling, H. Non-coding RNAs: Therapeutic Strategies and Delivery Systems. Adv. Exp. Med. Biol. 2016, 937, 229–237. [Google Scholar] [CrossRef]
- Patel, S.; Panchasara, H.; Braddick, D.; Gohil, N.; Singh, V. Synthetic small RNAs: Current status, challenges, and opportunities. J. Cell. Biochem. 2018, 119, 9619–9639. [Google Scholar] [CrossRef]
- Rebolledo, C.; Cuevas, A.; Zambrano, T.; Acuña, J.J.; Jorquera, M.A.; Saavedra, K.; Martínez, C.; Lanas, F.; Serón, P.; Salazar, L.A.; et al. Bacterial Community Profile of the Gut Microbiota Differs between Hypercholesterolemic Subjects and Controls. BioMed Res. Int. 2017, 2017, 8127814. [Google Scholar] [CrossRef] [PubMed]
- Kappel, B.A.; Federici, M. Gut microbiome and cardiometabolic risk. Rev. Endocr. Metab. Disord. 2019, 20, 399–406. [Google Scholar] [CrossRef]
- Kim, M.; Huda, M.N.; Bennett, B.J. Sequence meets function-microbiota and cardiovascular disease. Cardiovasc. Res. 2022, 118, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Fassarella, M.; Blaak, E.E.; Penders, J.; Nauta, A.; Smidt, H.; Zoetendal, E.G. Gut microbiome stability and resilience: Elucidating the response to perturbations in order to modulate gut health. Gut 2021, 70, 595–605. [Google Scholar] [CrossRef]
- Sasaki, K.; Sasaki, D.; Hannya, A.; Tsubota, J.; Kondo, A. In vitro human colonic microbiota utilises D-β-hydroxybutyrate to increase butyrogenesis. Sci. Rep. 2020, 10, 8516. [Google Scholar] [CrossRef] [PubMed]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef] [PubMed]
- Chénard, T.; Malick, M.; Dubé, J.; Massé, E. The influence of blood on the human gut microbiome. BMC Microbiol. 2020, 20, 44. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Zhuang, X.; Zhuo, S.; Zhu, Y.; Hu, S.; Zhao, M.; Tang, C.; Zhang, Z.; Li, X.; Ma, R.; et al. Dietary inflammatory potential mediated gut microbiota and metabolite alterations in Crohn’s disease: A fire-new perspective. Clin. Nutr. 2022, 41, 1260–1271. [Google Scholar] [CrossRef]
- Petrov, V.A.; Saltykova, I.V.; Zhukova, I.A.; Alifirova, V.M.; Zhukova, N.G.; Dorofeeva, Y.B.; Tyakht, A.V.; Kovarsky, B.A.; Alekseev, D.G.; Kostryukova, E.S.; et al. Analysis of Gut Microbiota in Patients with Parkinson’s Disease. Bull. Exp. Biol. Med. 2017, 162, 734–737. [Google Scholar] [CrossRef]
- Malinen, E.; Krogius-Kurikka, L.; Lyra, A.; Nikkilä, J.; Jääskeläinen, A.; Rinttilä, T.; Vilpponen-Salmela, T.; von Wright, A.J.; Palva, A. Association of symptoms with gastrointestinal microbiota in irritable bowel syndrome. World J. Gastroenterol. 2010, 16, 4532–4540. [Google Scholar] [CrossRef]
- Chen, P.; Zhou, J.; Wan, Y.; Liu, H.; Li, Y.; Liu, Z.; Wang, H.; Lei, J.; Zhao, K.; Zhang, Y.; et al. A Cas12a ortholog with stringent PAM recognition followed by low off-target editing rates for genome editing. Genome Biol. 2020, 21, 78. [Google Scholar] [CrossRef] [PubMed]
- Citrin, K.M.; Fernández-Hernando, C.; Suárez, Y. MicroRNA regulation of cholesterol metabolism. Ann. N. Y. Acad. Sci. 2021, 1495, 55–77. [Google Scholar] [CrossRef]
- Touyama, M.; Jin, J.S.; Kibe, R.; Hayashi, H.; Benno, Y. Quantification of Blautia wexlerae and Blautia luti in human faeces by real-time PCR using specific primers. Benef. Microbes 2015, 6, 583–590. [Google Scholar] [CrossRef]
- Benítez-Páez, A.; Gómez Del Pugar, E.M.; López-Almela, I.; Moya-Pérez, Á.; Codoñer-Franch, P.; Sanz, Y. Depletion of Blautia Species in the Microbiota of Obese Children Relates to Intestinal Inflammation and Metabolic Phenotype Worsening. mSystems 2020, 5, e00857-19. [Google Scholar] [CrossRef]
- Hosomi, K.; Saito, M.; Park, J.; Murakami, H.; Shibata, N.; Ando, M.; Nagatake, T.; Konishi, K.; Ohno, H.; Tanisawa, K.; et al. Oral administration of Blautia wexlerae ameliorates obesity and type 2 diabetes via metabolic remodeling of the gut microbiota. Nat. Commun. 2022, 13, 4477. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Yu, X.; Liu, Y.; Sun, L.; Chen, P.; Ding, Q.; Gao, Y.; Zhang, X.; Yu, M.; Liu, Y.; et al. The Baseline Gut Microbiota Directs Dieting-Induced Weight Loss Trajectories. Gastroenterology 2021, 160, 2029–2042.e16. [Google Scholar] [CrossRef]
- Khan, S.A.; Naz, A.; Qamar Masood, M.; Shah, R. Meta-Analysis of Inclisiran for the Treatment of Hypercholesterolemia. Am. J. Cardiol. 2020, 134, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Merćep, I.; Friščić, N.; Strikić, D.; Reiner, Ž. Advantages and Disadvantages of Inclisiran: A Small Interfering Ribonucleic Acid Molecule Targeting PCSK9-A Narrative Review. Cardiovasc. Ther. 2022, 2022, 8129513. [Google Scholar] [CrossRef]
- Tarallo, S.; Ferrero, G.; De Filippis, F.; Francavilla, A.; Pasolli, E.; Panero, V.; Cordero, F.; Segata, N.; Grioni, S.; Pensa, R.G.; et al. Stool microRNA profiles reflect different dietary and gut microbiome patterns in healthy individuals. Gut 2022, 71, 1302–1314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef]
- Stanton, B.A. Extracellular Vesicles and Host-Pathogen Interactions: A Review of Inter-Kingdom Signaling by Small Noncoding RNA. Genes 2021, 12, 1010. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Um, J.H.; Cho, J.H.; Lee, H.J. Tiny RNAs and their voyage via extracellular vesicles: Secretion of bacterial small RNA and eukaryotic microRNA. Exp. Biol. Med. 2017, 242, 1475–1481. [Google Scholar] [CrossRef]
- Askenase, P.W. Exosome Carrier Effects; Resistance to Digestion in Phagolysosomes May Assist Transfers to Targeted Cells; II Transfers of miRNAs Are Better Analyzed via Systems Approach as They Do Not Fit Conventional Reductionist Stoichiometric Concepts. Int. J. Mol. Sci. 2022, 23, 6192. [Google Scholar] [CrossRef]
- Morales, C.; Rojas, G.; Rebolledo, C.; Rojas-Herrera, M.; Arias-Carrasco, R.; Cuadros-Orellana, S.; Maracaja-Coutinho, V.; Saavedra, K.; Leal, P.; Lanas, F.; et al. Characterization of microbial communities from gut microbiota of hypercholesterolemic and control subjects. Front. Cell. Infect. Microbiol. 2022, 12, 943609. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Babraham Institute. Babraham Bioinformatics: FastQC. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 11 January 2023).
- Moraga, C.; Sanchez, E.; Ferrarini, M.G.; Gutierrez, R.A.; Vidal, E.A.; Sagot, M.F. BrumiR: A toolkit for de novo discovery of microRNAs from sRNA-seq data. GigaScience 2022, 11, giac093. [Google Scholar] [CrossRef]
- Langmead, B.; Wilks, C.; Antonescu, V.; Charles, R. Scaling read aligners to hundreds of threads on general-purpose processors. Bioinformatics 2019, 35, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Mann, M.; Wright, P.R.; Backofen, R. IntaRNA 2.0: Enhanced and customizable prediction of RNA-RNA interactions. Nucleic Acids Res. 2017, 45, W435–W439. [Google Scholar] [CrossRef]
- Mathews, D.H.; Disney, M.D.; Childs, J.L.; Schroeder, S.J.; Zuker, M.; Turner, D.H. Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure. Proc. Natl. Acad. Sci. USA 2004, 101, 7287–7292. [Google Scholar] [CrossRef]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuböck, R.; Hofacker, I.L. The Vienna RNA websuite. Nucleic Acids Res. 2008, 36, W70–W74. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Normocholesterolemic (n = 30) | Hypercholesterolemic (n = 27) | p |
|---|---|---|---|
| Age (years) | 60.07 ± 10.27 | 60.83 ± 5.00 | 0.525 |
| Height (m) | 1.64 ± 0.02 | 1.58 ± 0.02 | 0.068 |
| Weight (kg) | 79.34 ± 2.61 | 64.48 ± 1.83 | <0.0001 * |
| BMI 1 (kg/m2) | 29.72 ± 4.20 | 25.83 ± 3.21 | 0.004 * |
| SBP 2 (mmHg) | 128.00 ± 16.77 | 123.00 ± 10.19 | 0.765 |
| DBP 3 (mmHg) | 81.29 ± 13.97 | 80.42 ± 8.93 | 0.970 |
| Glucose (mmol/L) | 5.88 ± 0.79 | 5.79 ± 0.72 | 0.688 |
| Triglycerides (mmol/L) | 1.73 ± 0.97 | 2.26 ± 0.93 | 0.003 * |
| Total Cholesterol (mmol/L) | 4.40 ± 0.53 | 7.55 ± 0.73 | <0.0001 * |
| HDL 4-Cholesterol (mmol/L) | 1.51 ± 0.31 | 1.58 ± 0.40 | 0.827 |
| LDL 5-Cholesterol (mmol/L) | 2.24 ± 0.41 | 5.23 ± 0.74 | <0.0001 * |
| NCBI Code (mRNA) | Molecule Name | Abbreviation |
|---|---|---|
| XM_011518339.3 | ATP binding cassette subfamily A member 1 | ABCA1 |
| XM_011529806.1 | ATP binding cassette subfamily G member 1 | ABCG1 |
| XM_011533024.2 | ATP binding cassette subfamily G member 5 | ABCG5 |
| XM_011533029.2 | ATP binding cassette subfamily G member 8 | ABCG8 |
| NM_000039.3 | Apolipoprotein A1 | APOA1 |
| NM_000384.3 | Apolipoprotein B | APOB |
| NM_001321065.2 | Apolipoprotein C1 | APOC1 |
| NM_001302688.2 | Apolipoprotein E | APOE |
| NM_000780.4 | Cytochrome P450 family 7 subfamily A member 1 | CYP7A1 |
| XM_011543357.1 | 3-hydroxy-3-methylglutaryl-CoA reductase | HMGCR |
| NM_000527.5 | Low density lipoprotein receptor | LDLR |
| XM_005254374.4 | Lipase C | LIPC |
| XM_005258277.1 | NPC intracellular cholesterol transporter 1 | NPC1 |
| XM_011515326.3 | NPC1 like intracellular cholesterol transporter 1 | NPC1L1 |
| NM_005693.4 | Nuclear receptor subfamily 1 group H member 3 | NR1H3 |
| NM_174936.4 | Proprotein convertase subtilisin/kexin type 9 | PCSK9 |
| NM_002957.6 | Retinoid X receptor alpha | RXRA |
| NM_003383.5 | Very low-density lipoprotein receptor | VLDLR |
| XM_024450893.1 | Sterol regulatory element binding transcription factor 1 | SREBF1 |
| XM_011530347.2 | Sterol regulatory element binding transcription factor 2 | SREBF2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales, C.; Arias-Carrasco, R.; Maracaja-Coutinho, V.; Seron, P.; Lanas, F.; Salazar, L.A.; Saavedra, N. Differences in Bacterial Small RNAs in Stool Samples from Hypercholesterolemic and Normocholesterolemic Subjects. Int. J. Mol. Sci. 2023, 24, 7213. https://doi.org/10.3390/ijms24087213
Morales C, Arias-Carrasco R, Maracaja-Coutinho V, Seron P, Lanas F, Salazar LA, Saavedra N. Differences in Bacterial Small RNAs in Stool Samples from Hypercholesterolemic and Normocholesterolemic Subjects. International Journal of Molecular Sciences. 2023; 24(8):7213. https://doi.org/10.3390/ijms24087213
Chicago/Turabian StyleMorales, Cristian, Raul Arias-Carrasco, Vinicius Maracaja-Coutinho, Pamela Seron, Fernando Lanas, Luis A. Salazar, and Nicolás Saavedra. 2023. "Differences in Bacterial Small RNAs in Stool Samples from Hypercholesterolemic and Normocholesterolemic Subjects" International Journal of Molecular Sciences 24, no. 8: 7213. https://doi.org/10.3390/ijms24087213
APA StyleMorales, C., Arias-Carrasco, R., Maracaja-Coutinho, V., Seron, P., Lanas, F., Salazar, L. A., & Saavedra, N. (2023). Differences in Bacterial Small RNAs in Stool Samples from Hypercholesterolemic and Normocholesterolemic Subjects. International Journal of Molecular Sciences, 24(8), 7213. https://doi.org/10.3390/ijms24087213