
Patients with Chronic Spinal Cord Injury and a Long Period of Evolution Exhibit an Altered Cytokine Production by CD4 and CD8 T Cell Populations

 ,
,  , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Results
2.1. Patients Demographics
2.2. Patients with Chronic Spinal Cord Injury and a Long Period of Evolution Exhibit an Altered Cytokine Production by Total, Naïve, Effector, and Central/Effector Memory CD4+ Cells
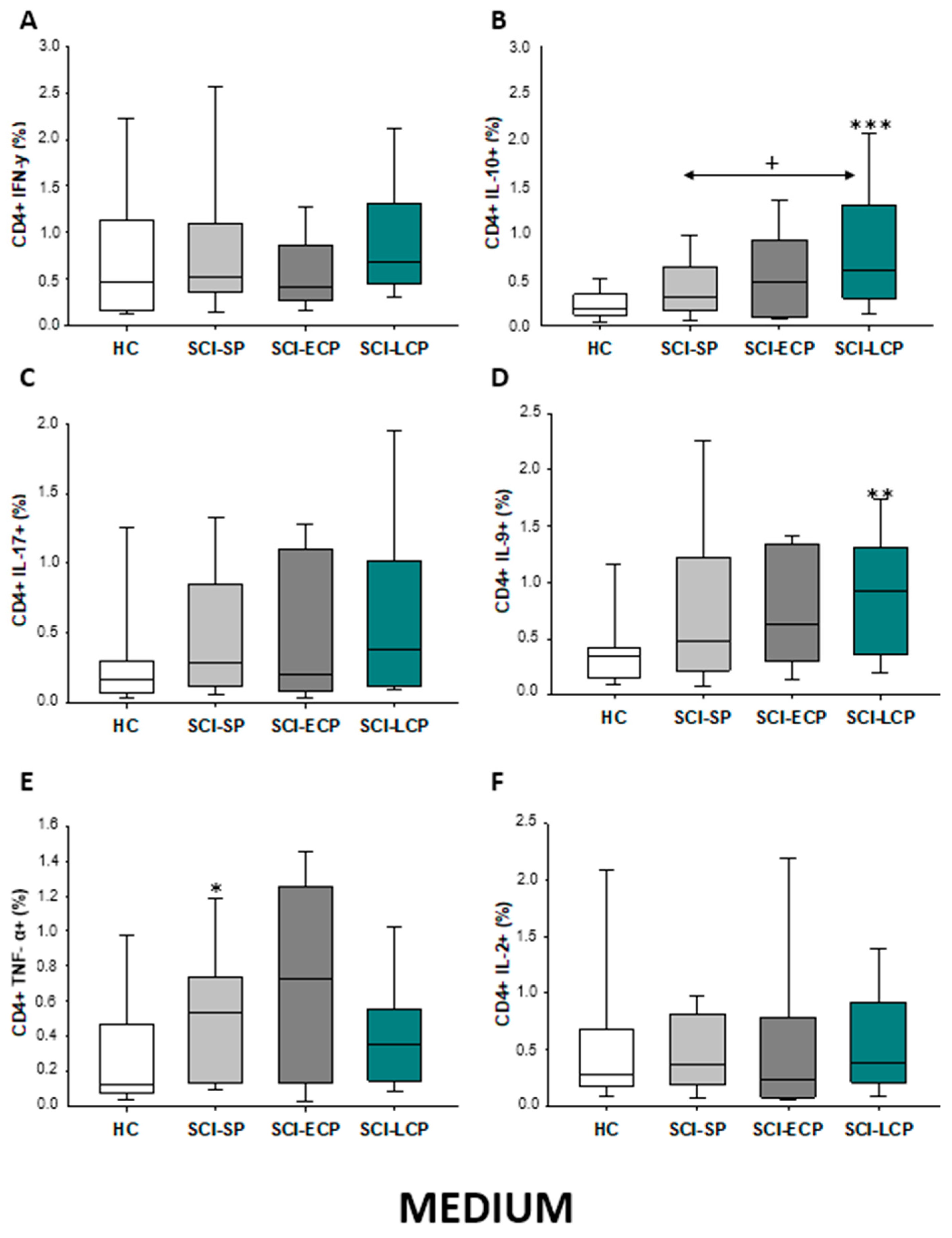
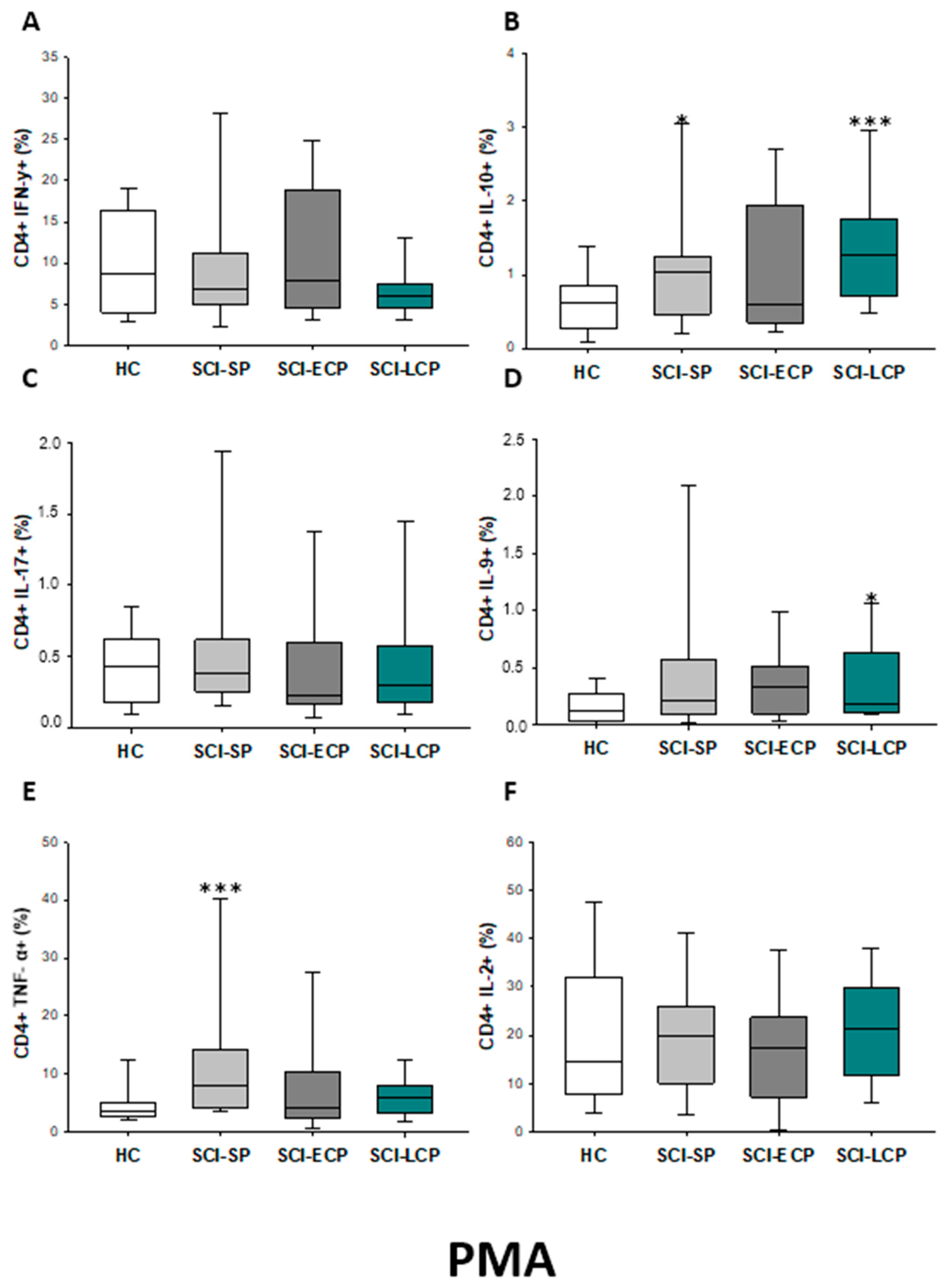
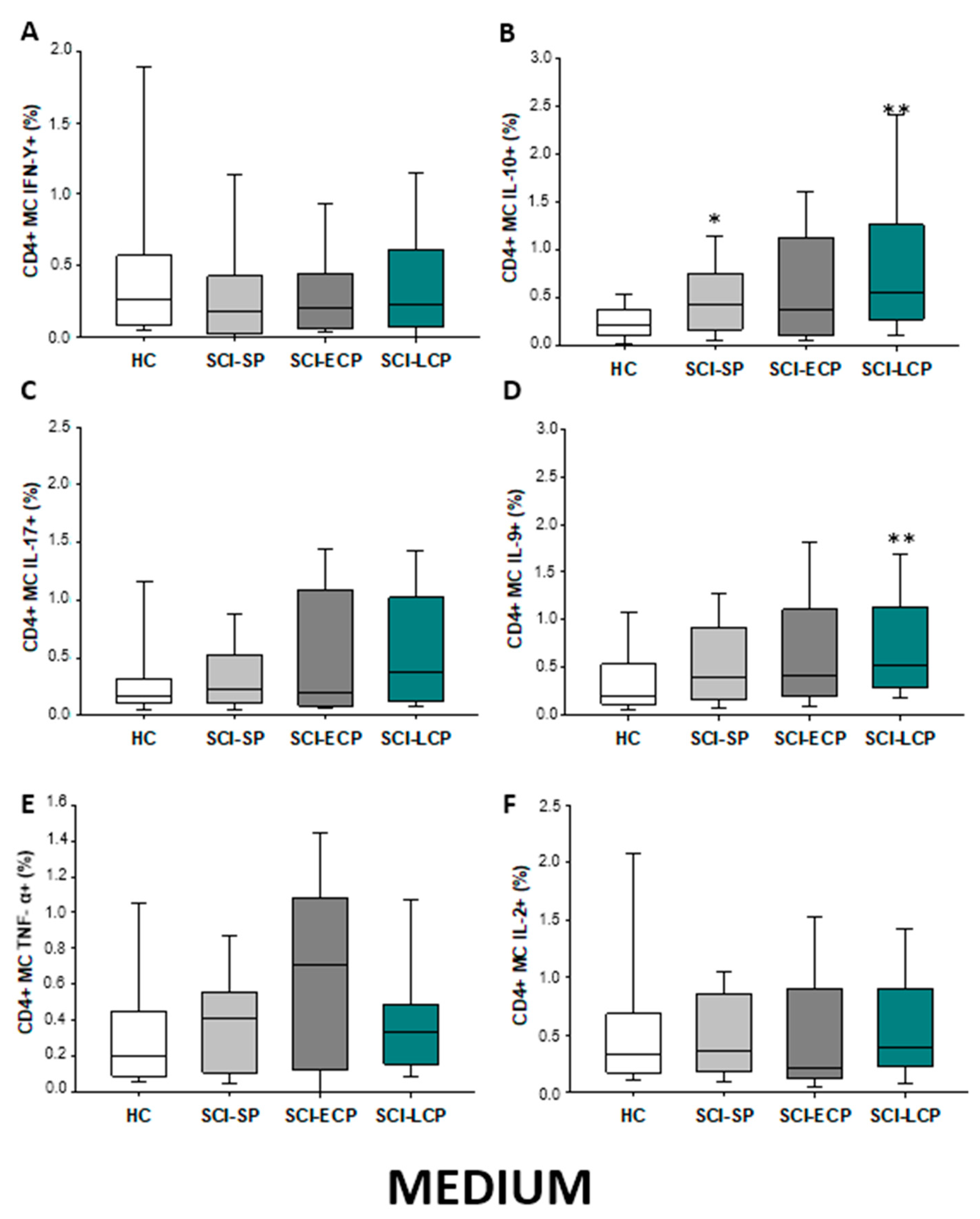
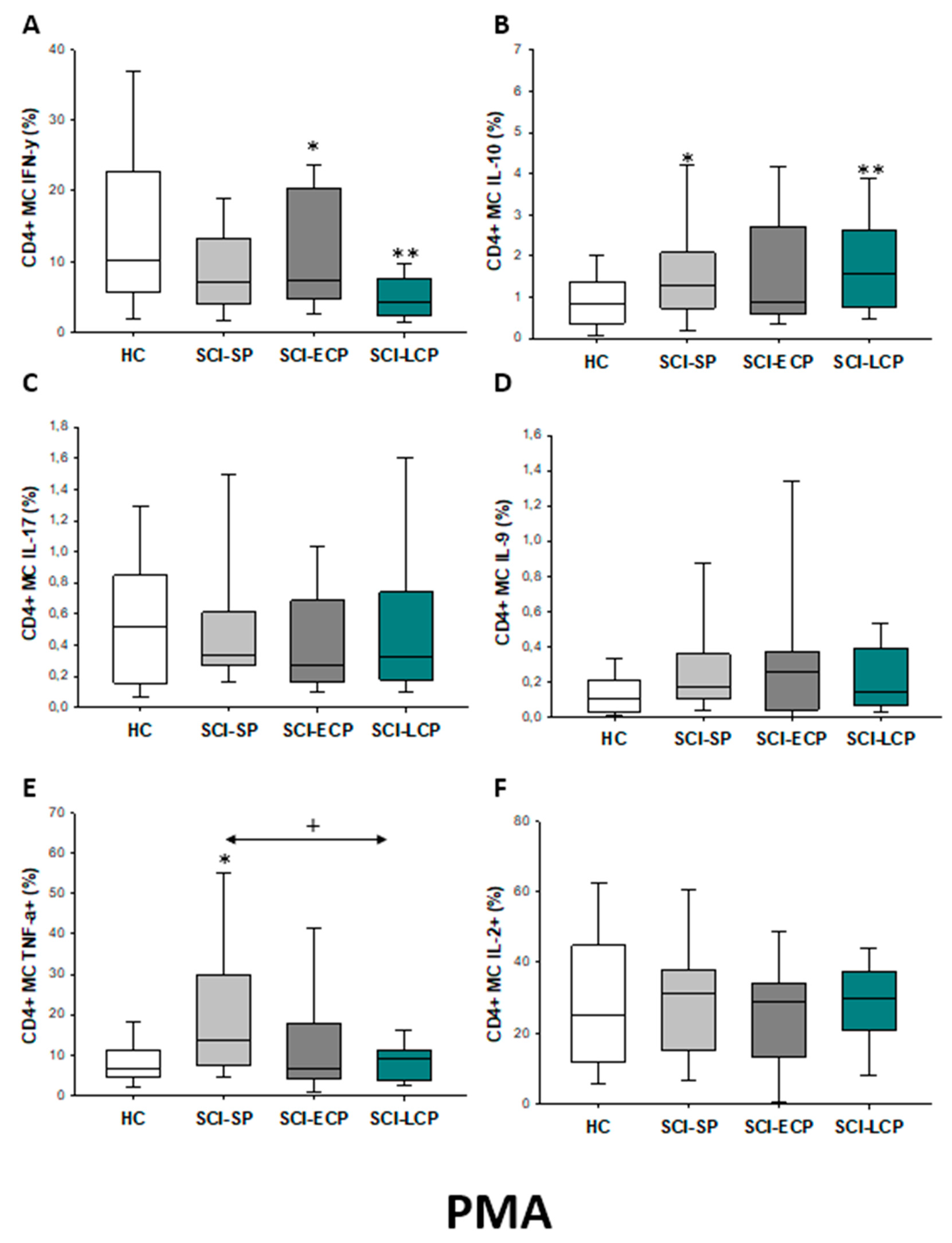
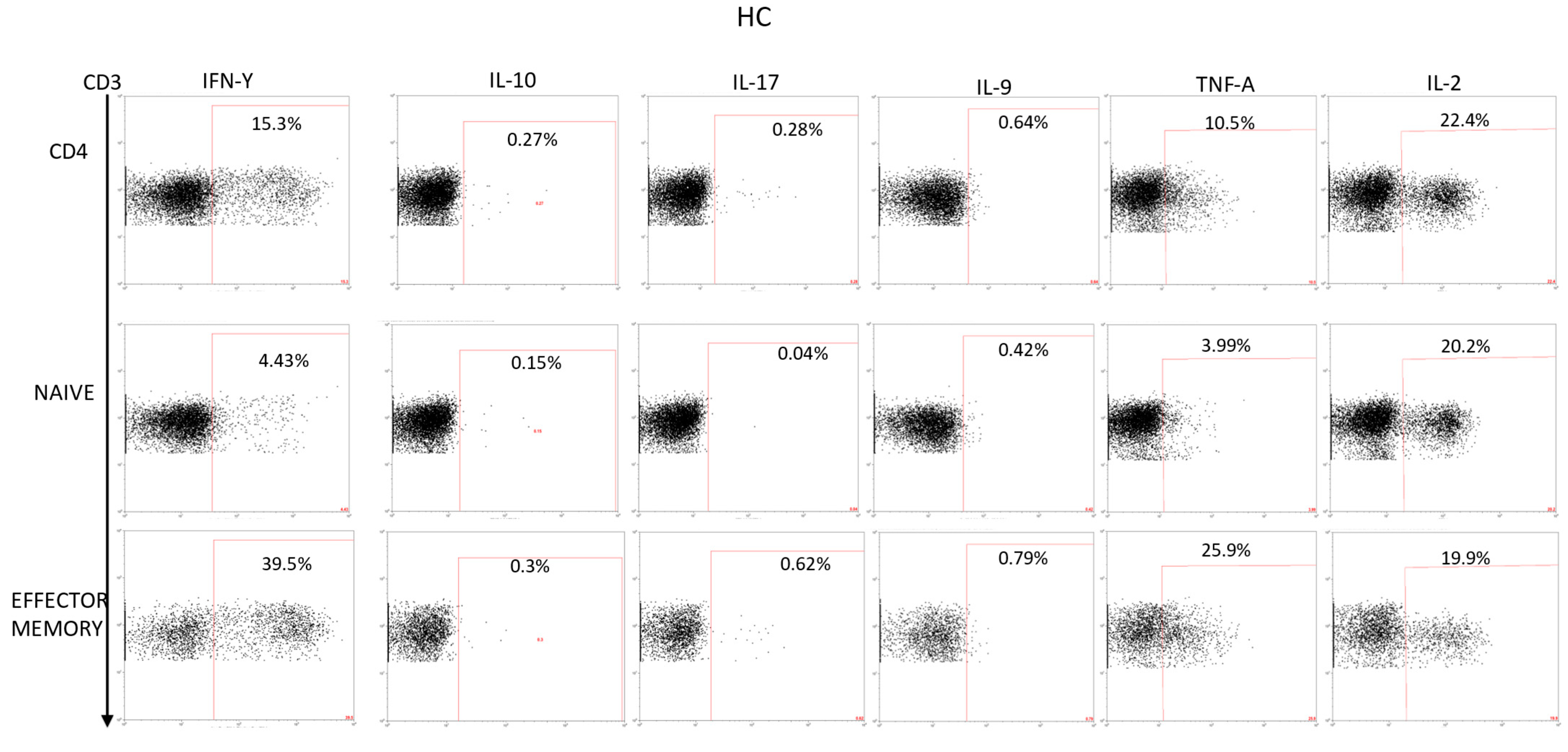
2.2.1. Cytokine Expression in CD4 Lymphocytes
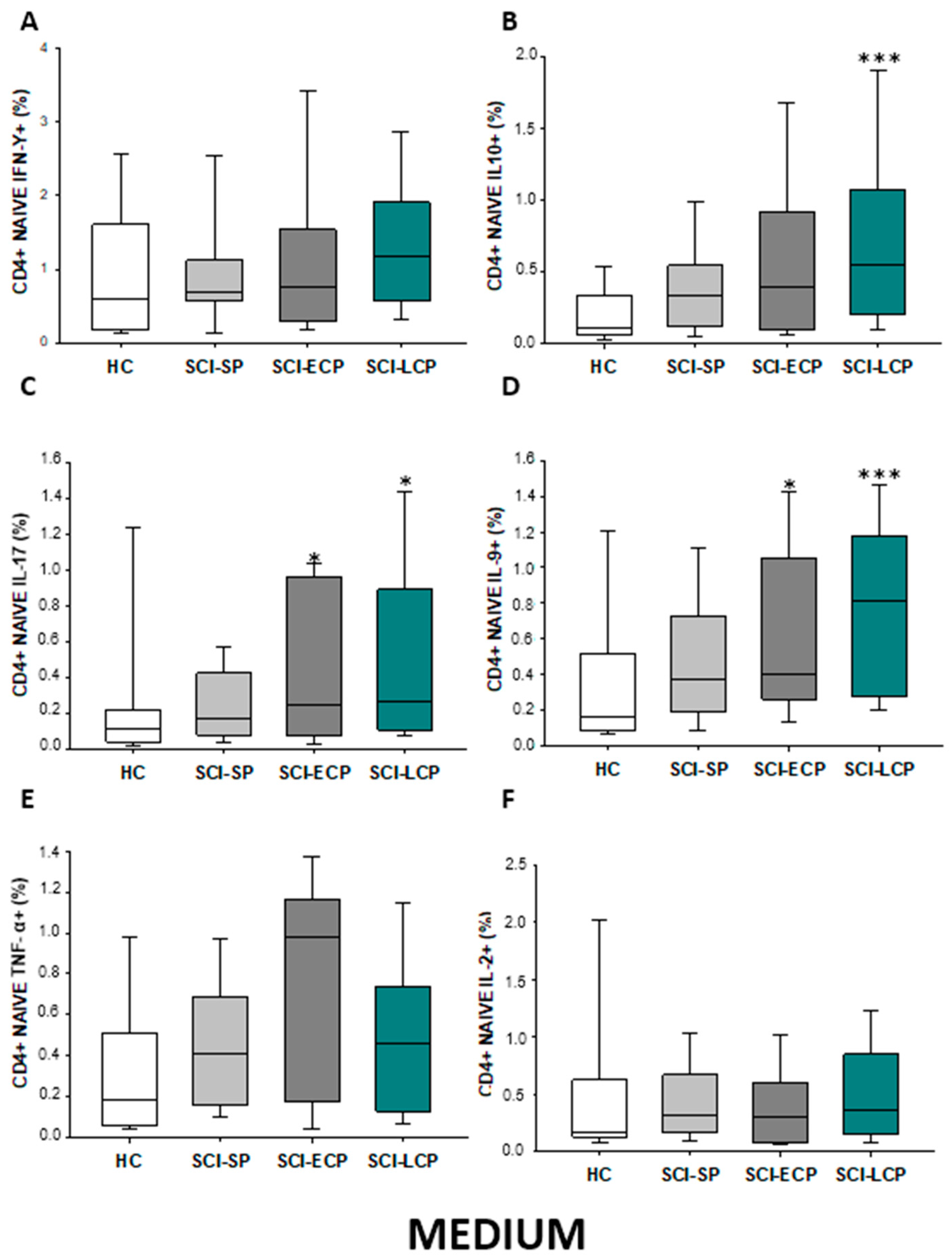
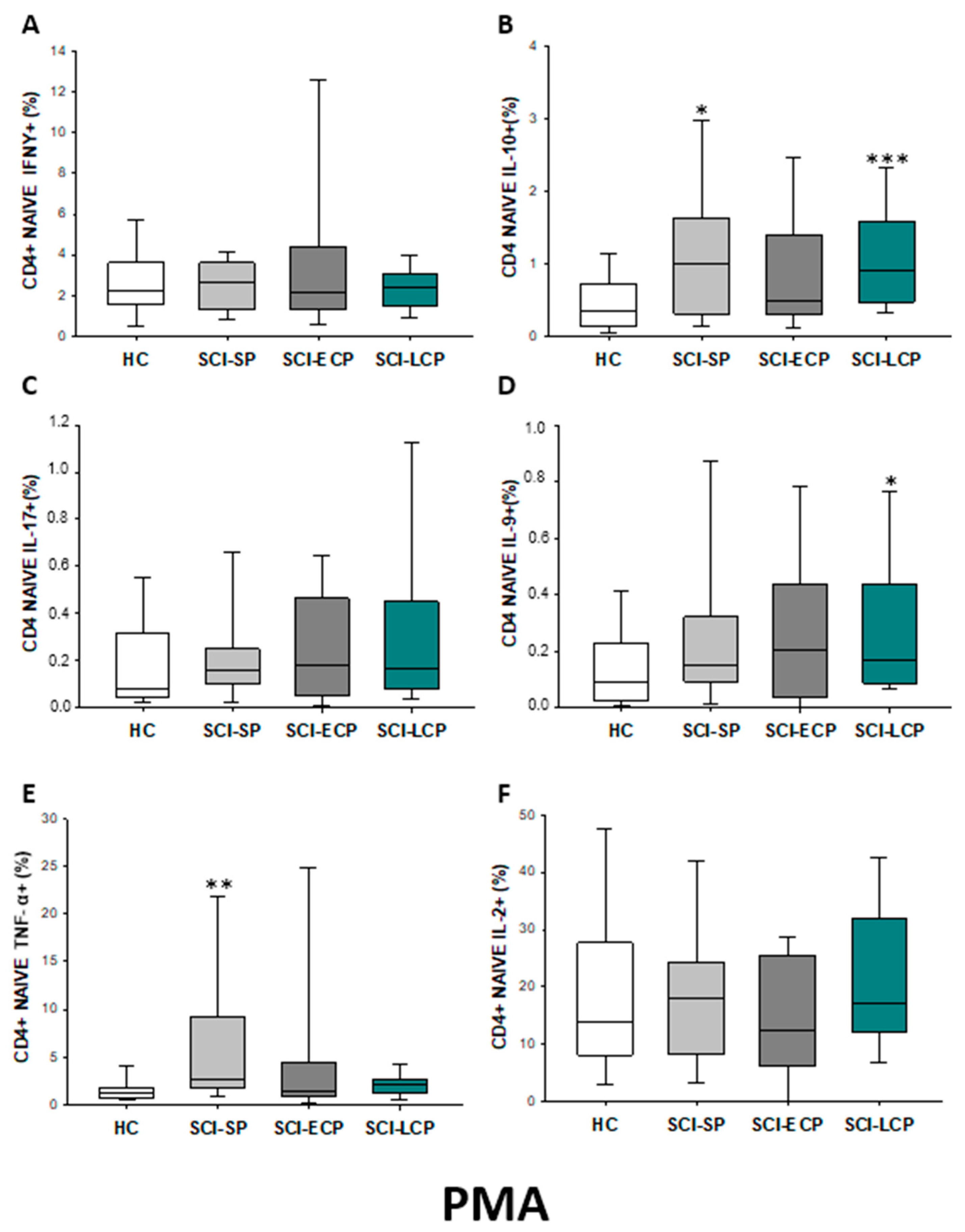
2.2.2. Cytokine Expression in CD4 Naïve Lymphocytes
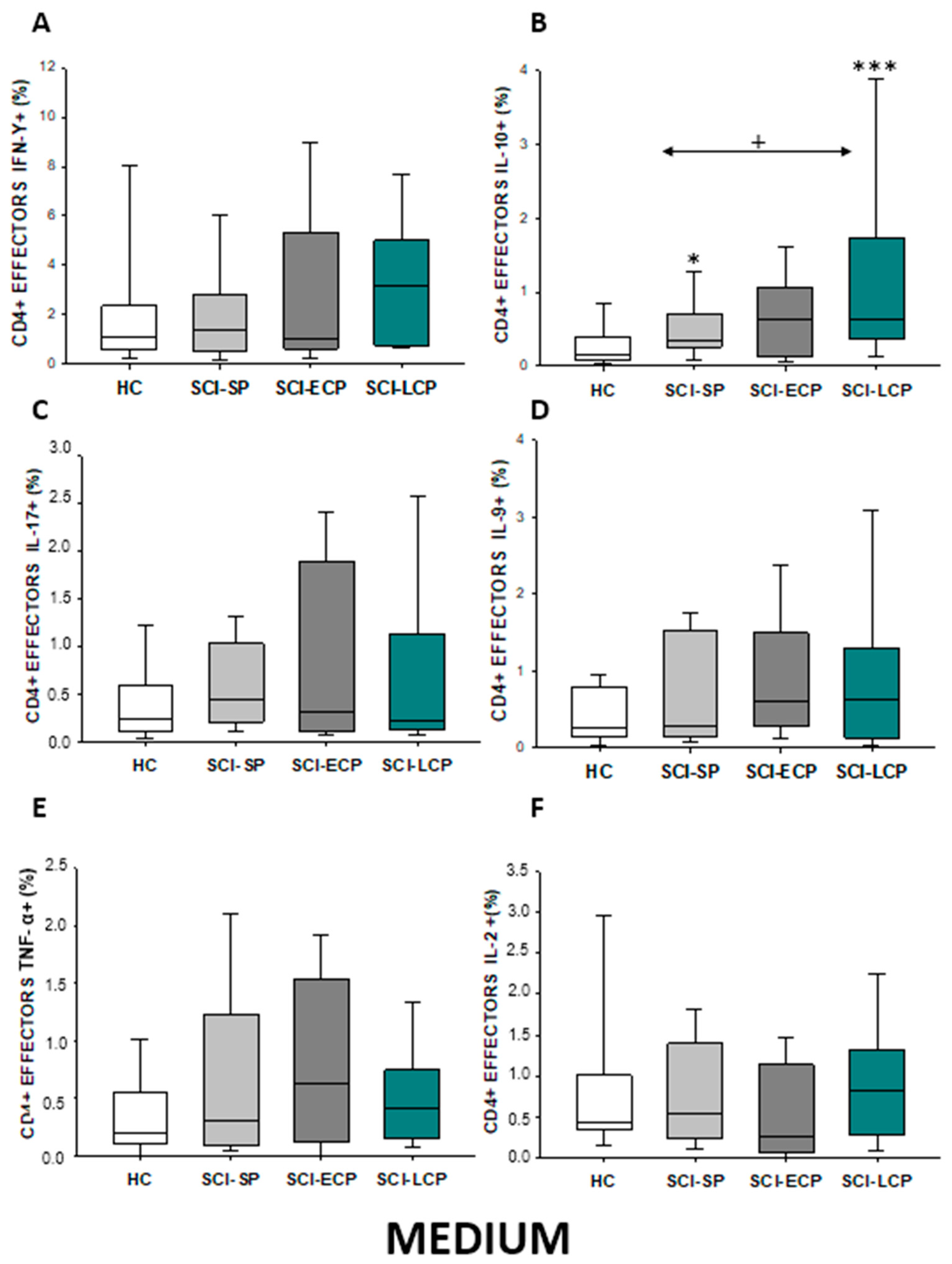
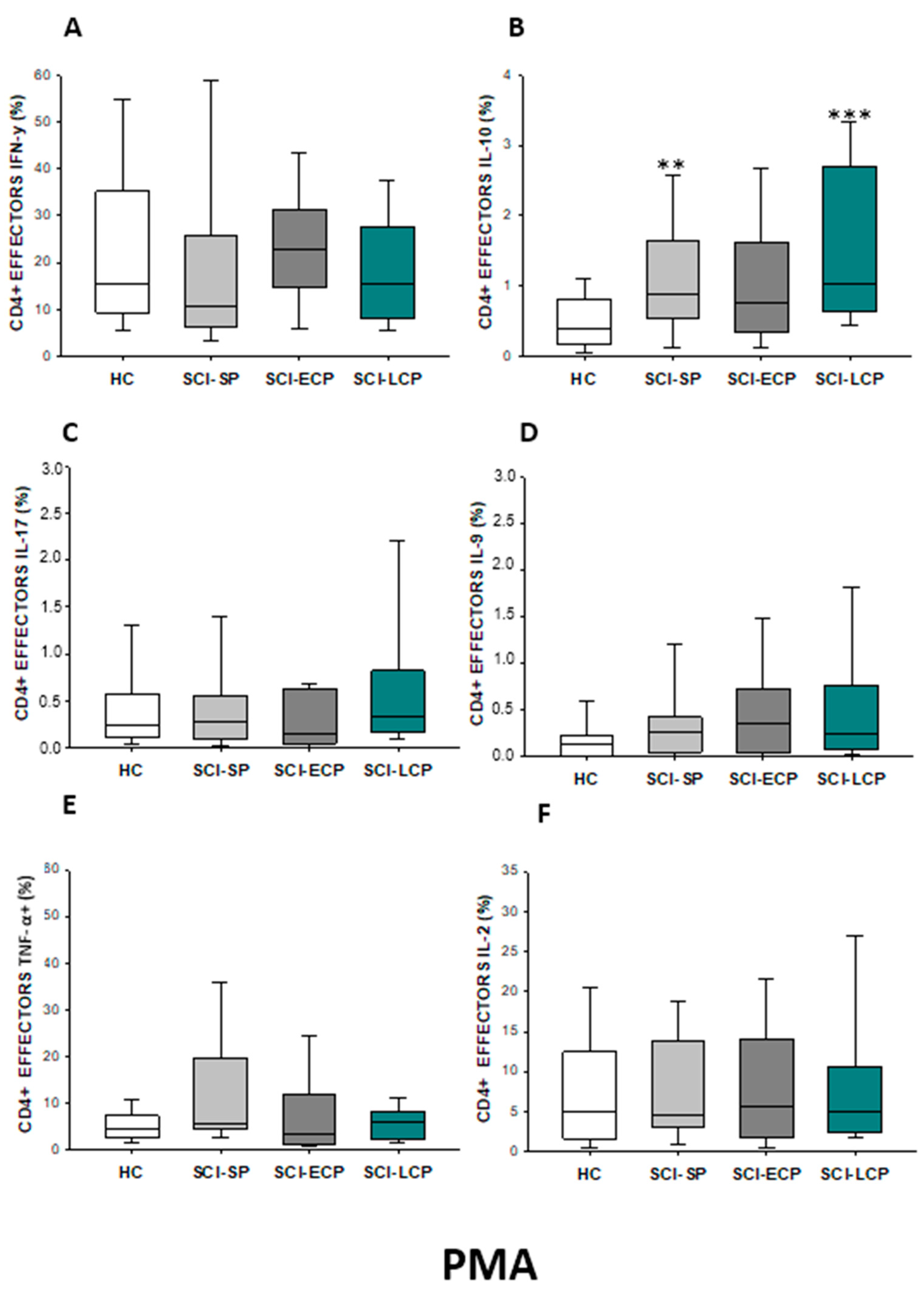
2.2.3. Cytokine Expression in CD4 Effector Lymphocytes
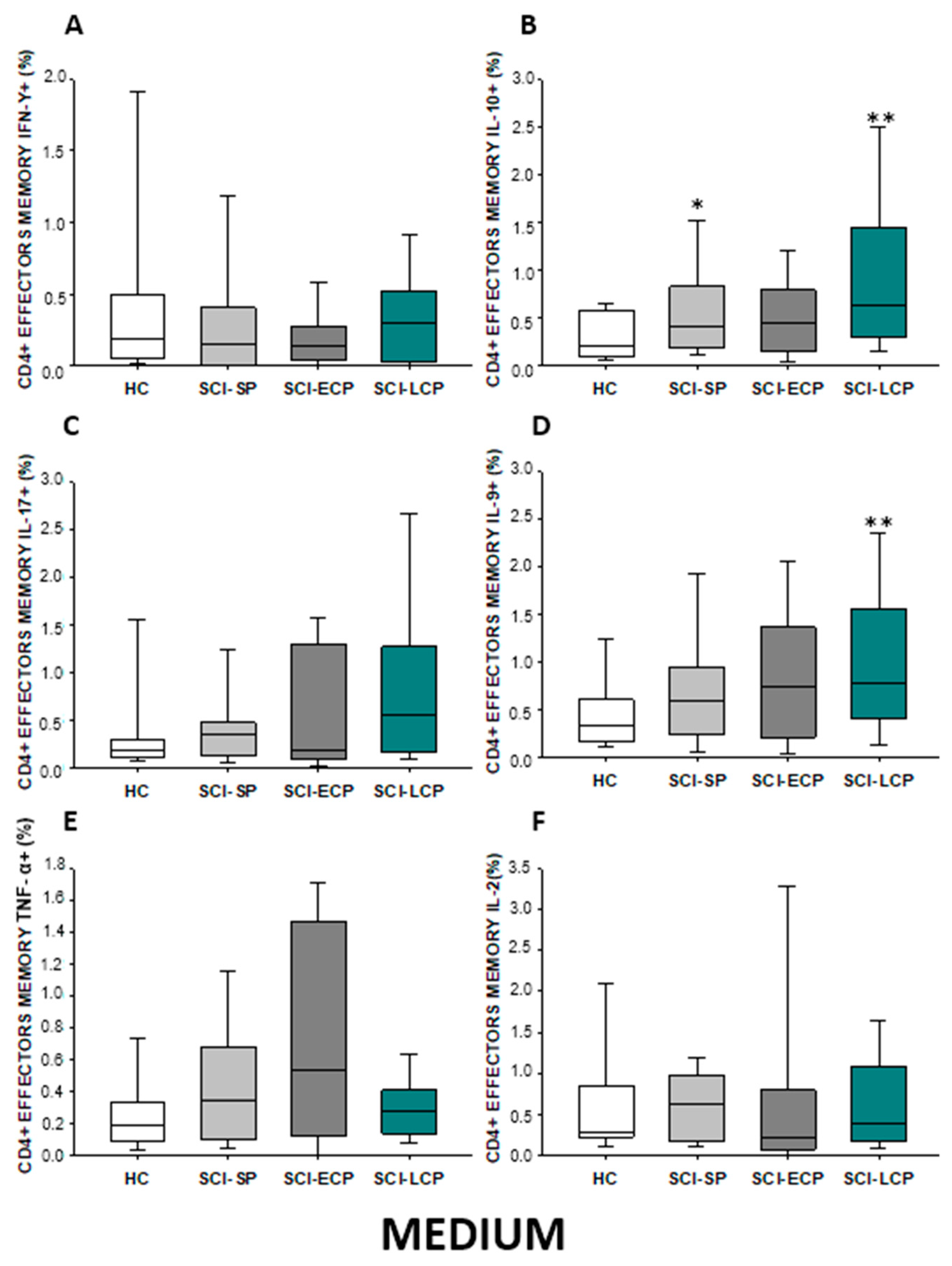
2.2.4. Cytokine Expression in CD4 Effector Memory Lymphocytes
2.2.5. Cytokine Expression in CD4 Central Memory Lymphocytes
2.3. Patients with Chronic Spinal Cord Injury Display a Distinct Cytokine Production by Circulating Total, Naïve, Effector, and Central/Effector Memory CD8 Cells Depending on the Years of Evolution
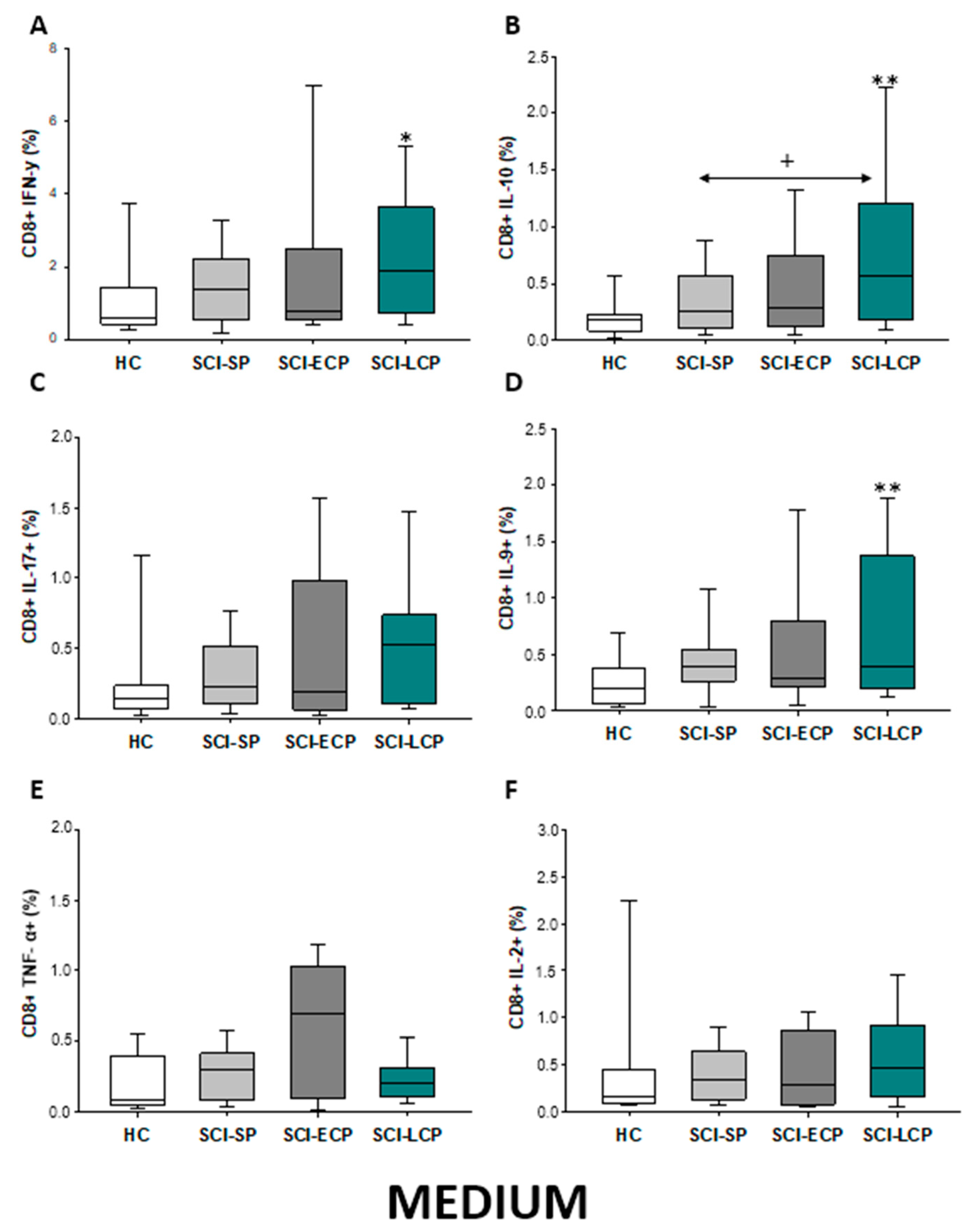
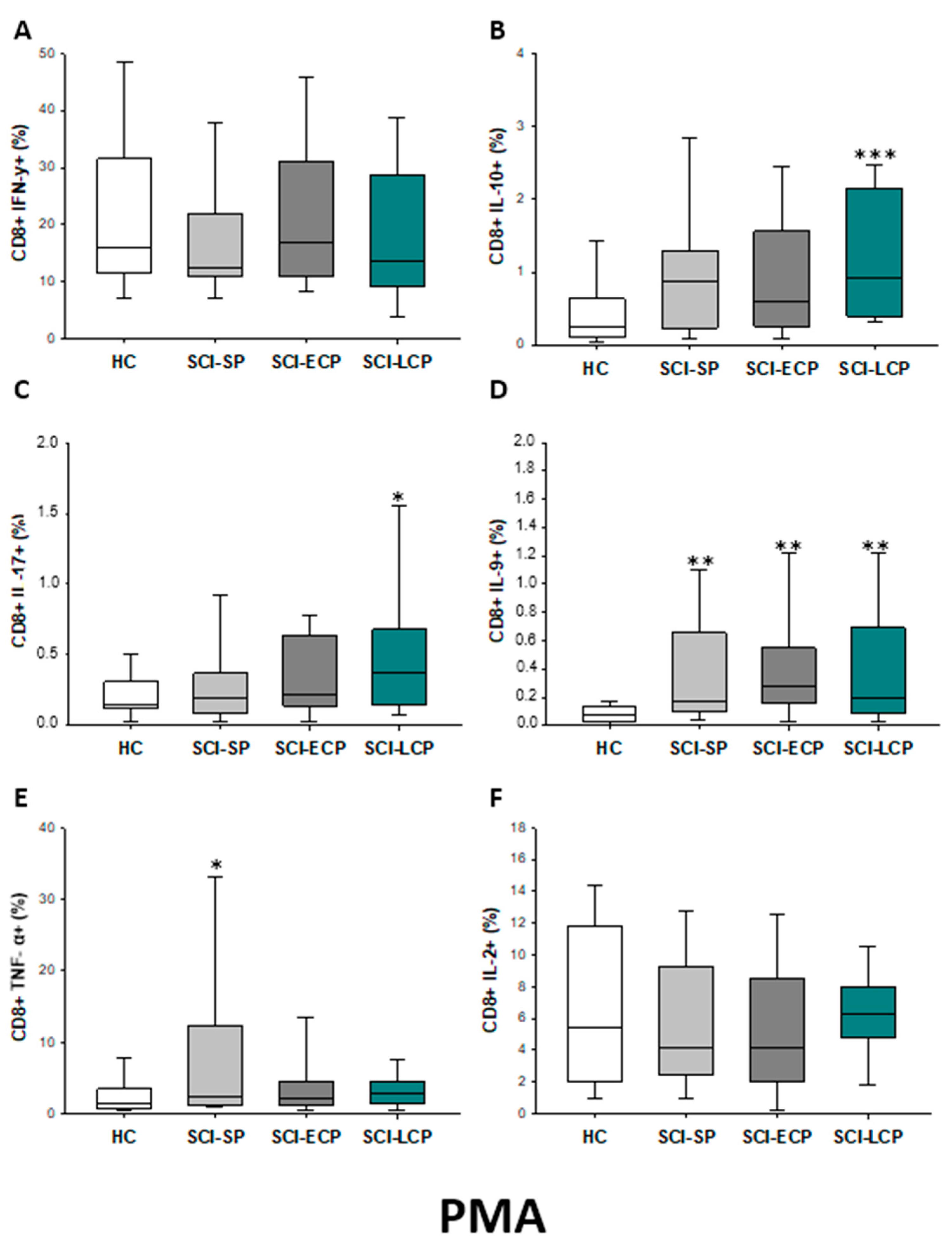
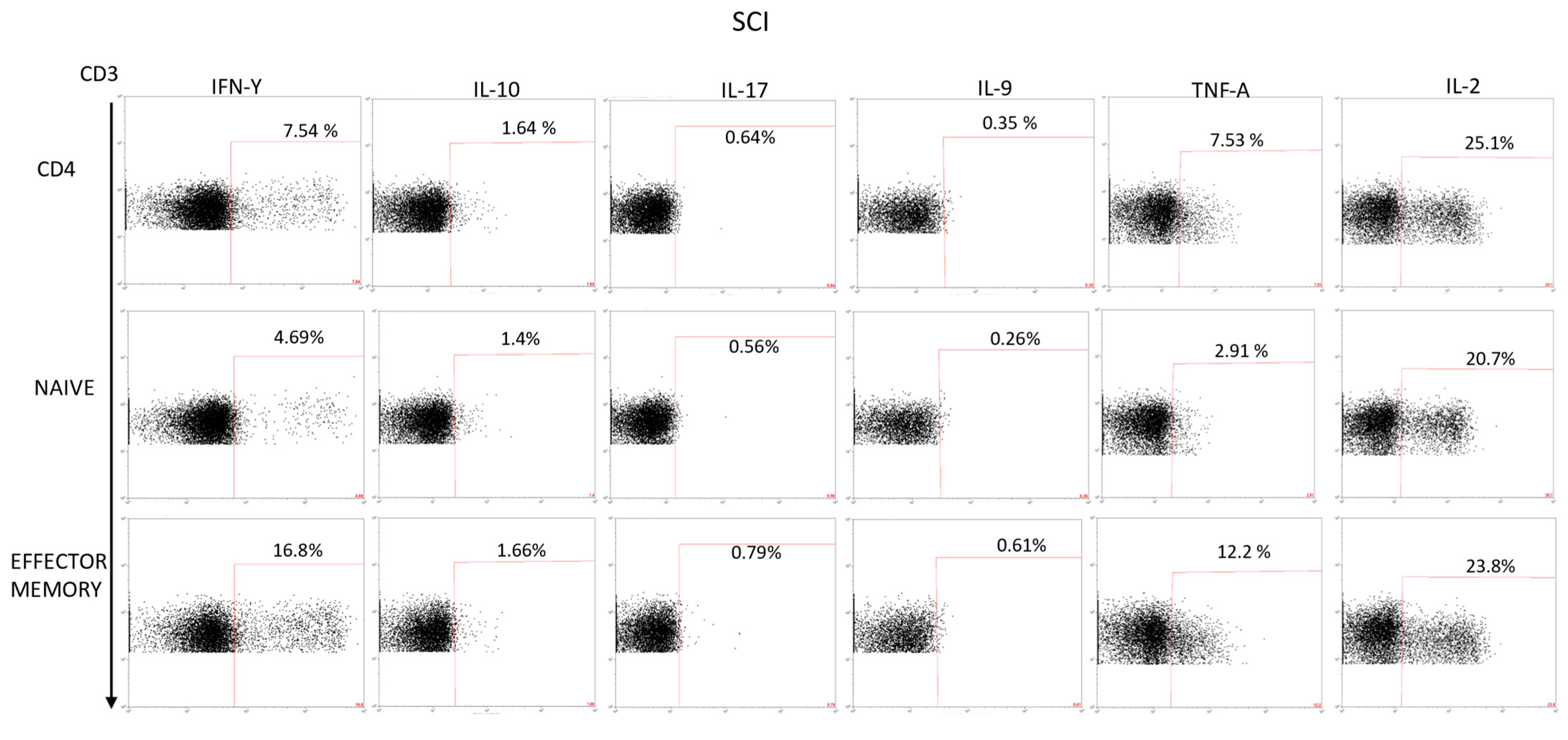
2.3.1. Cytokine Expression in CD8 Lymphocytes
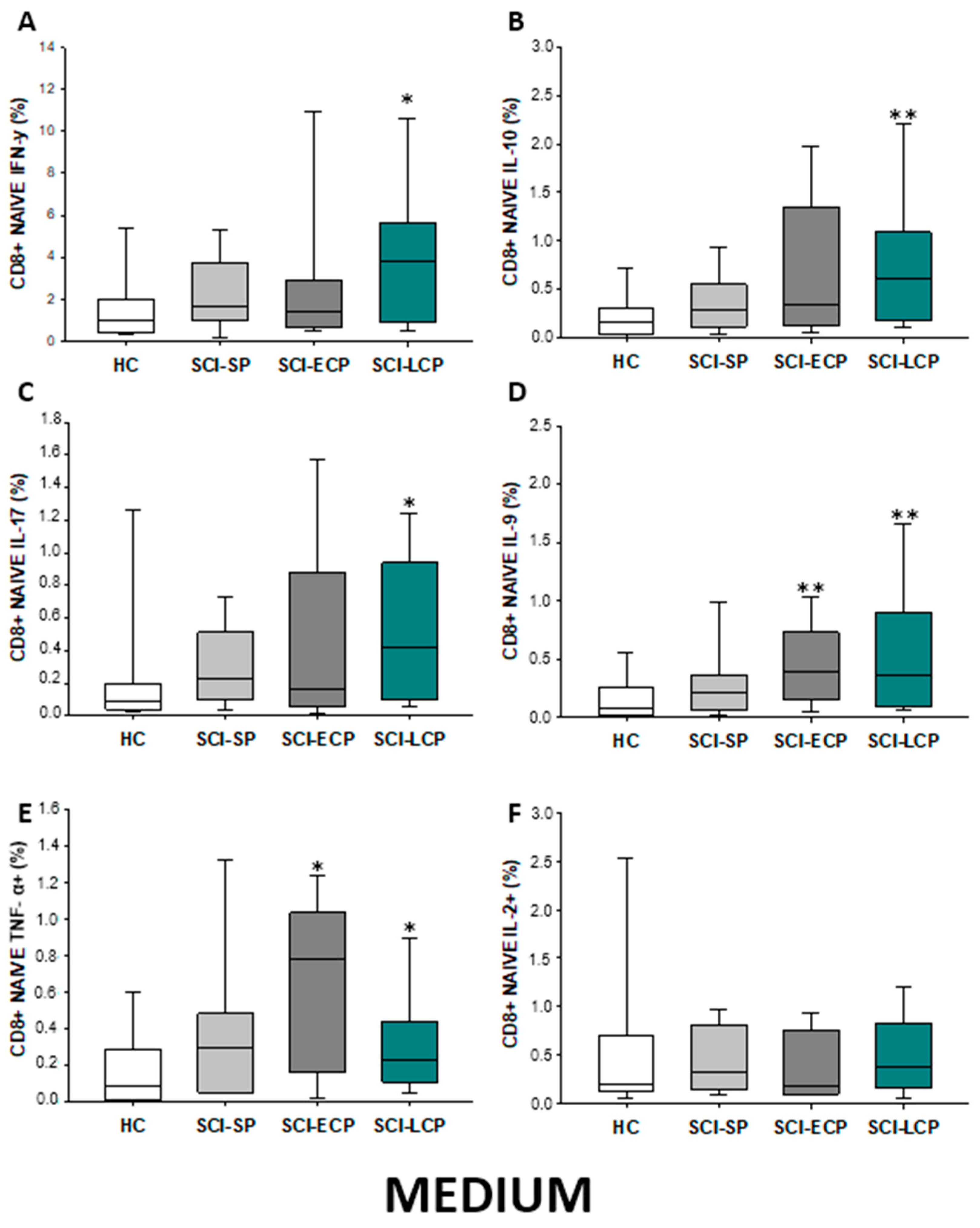
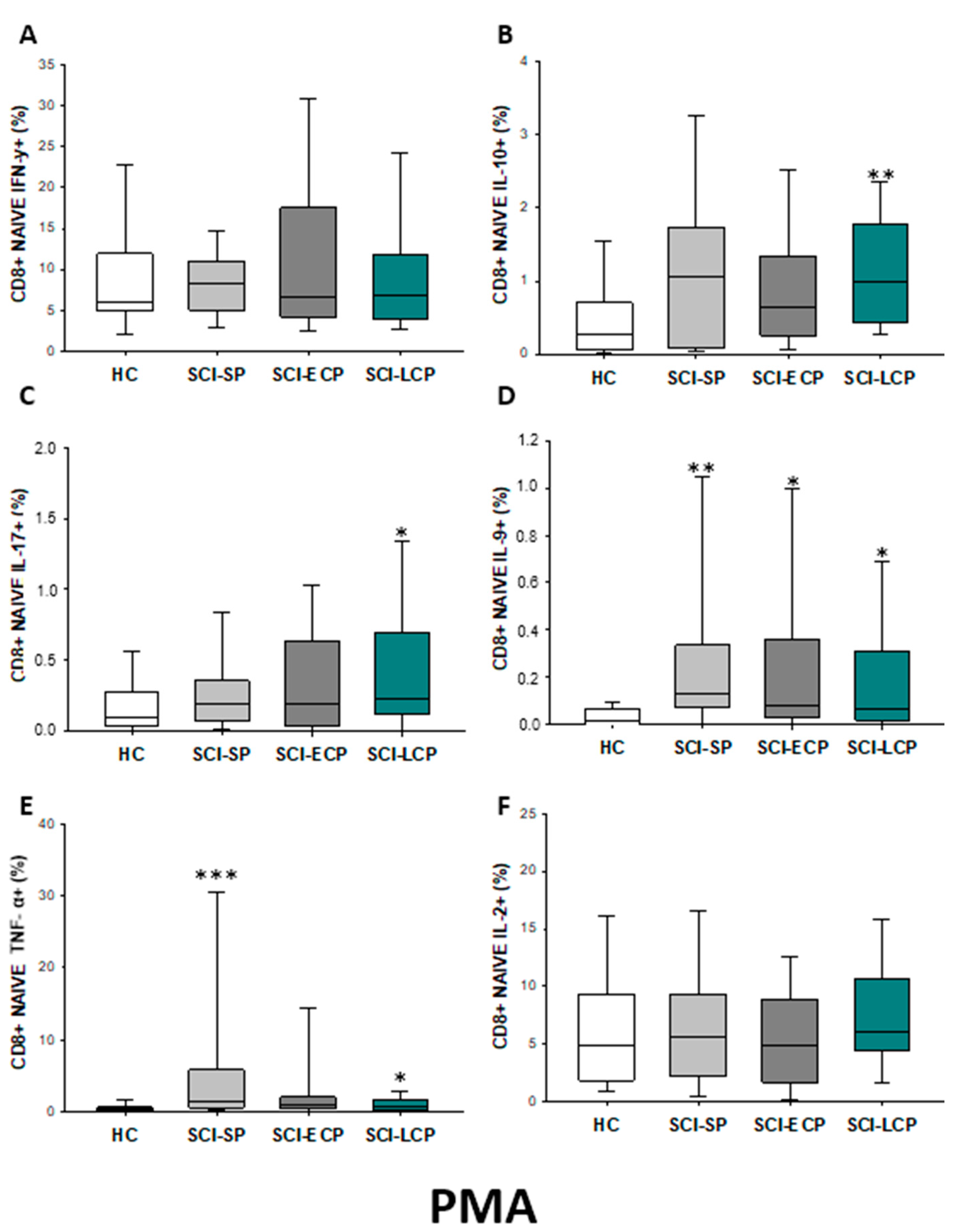
2.3.2. Cytokine Expression in CD8 Naïve Lymphocytes
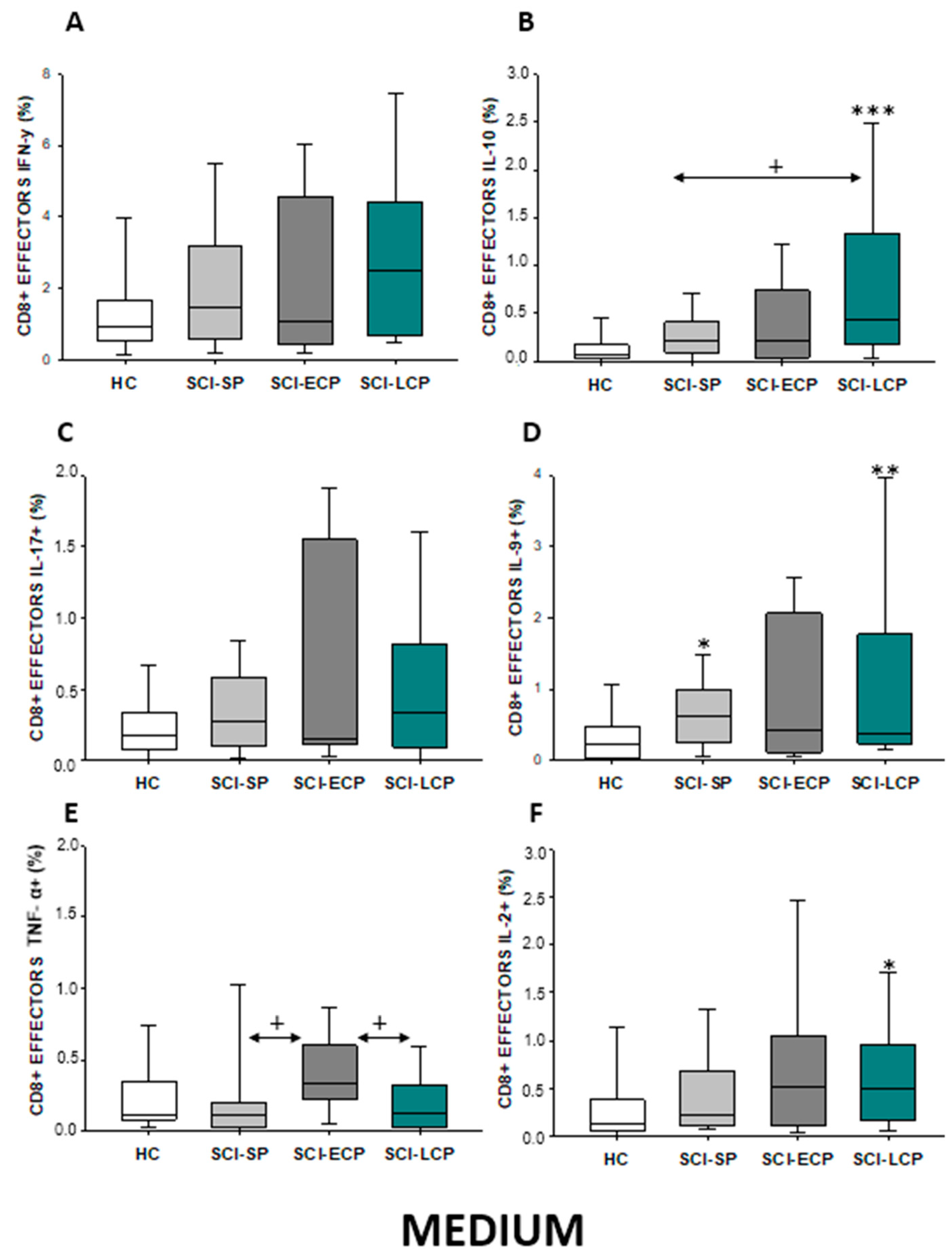
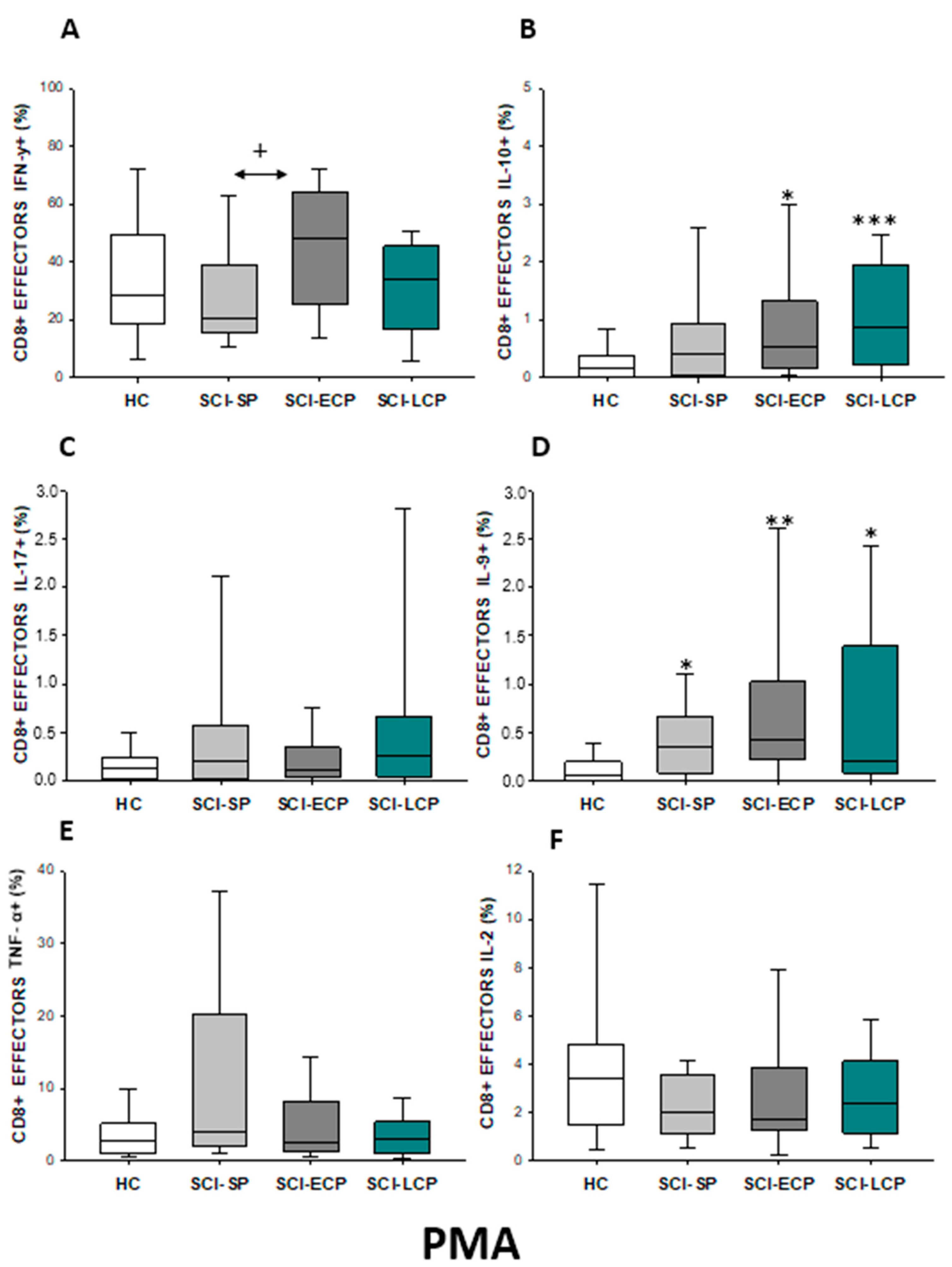
2.3.3. Cytokine Expression in CD8 Effector Lymphocytes
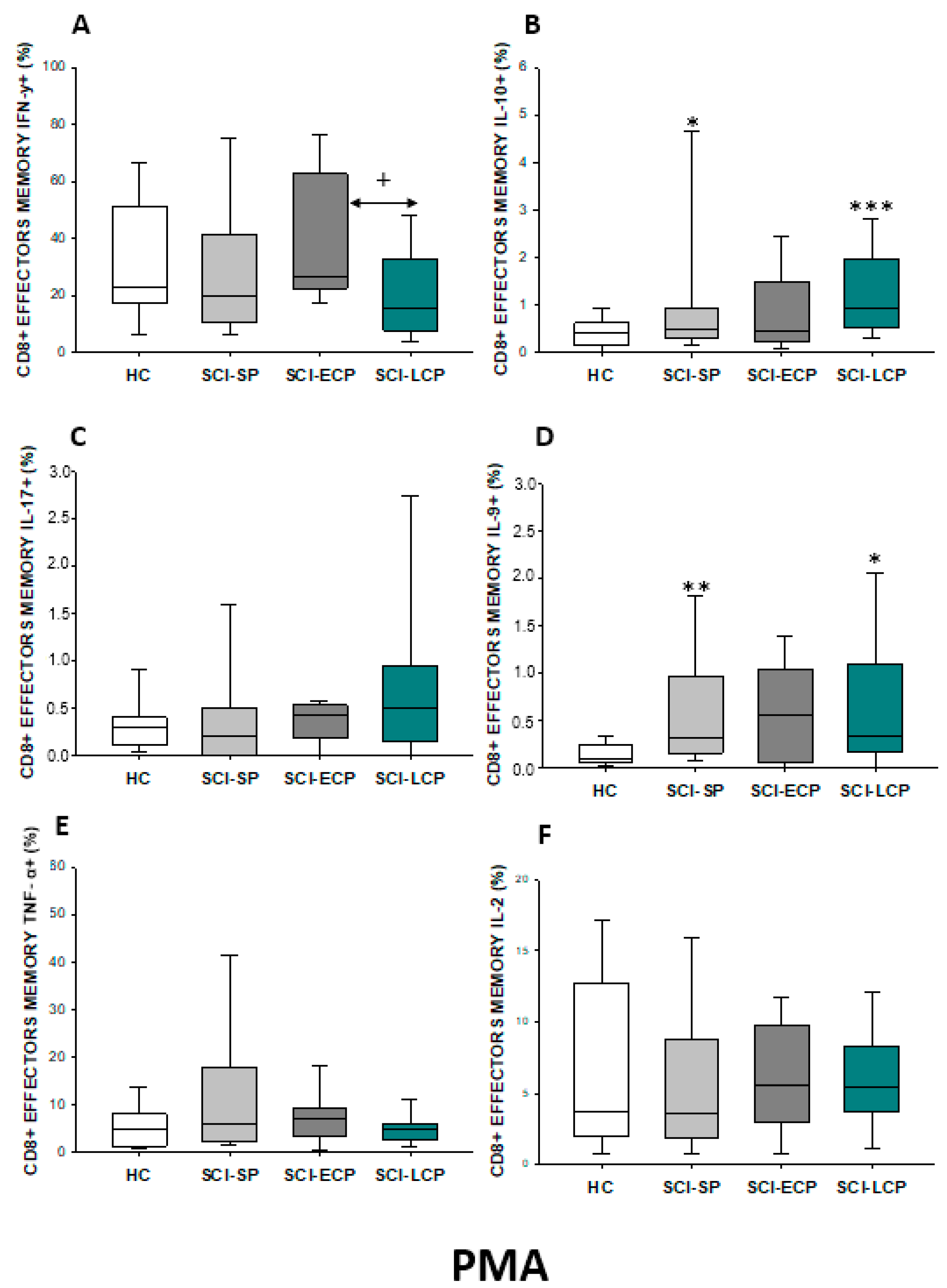
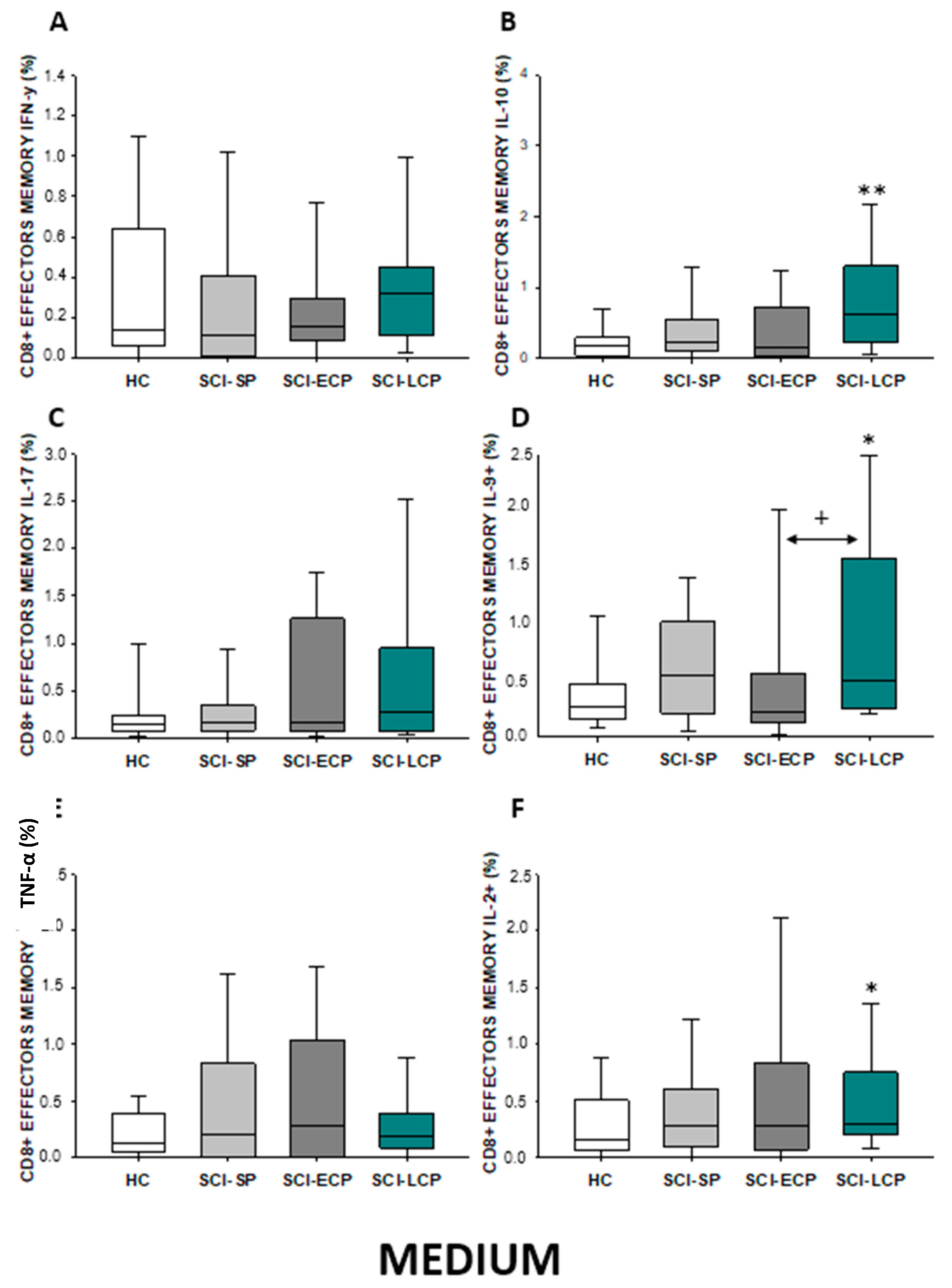
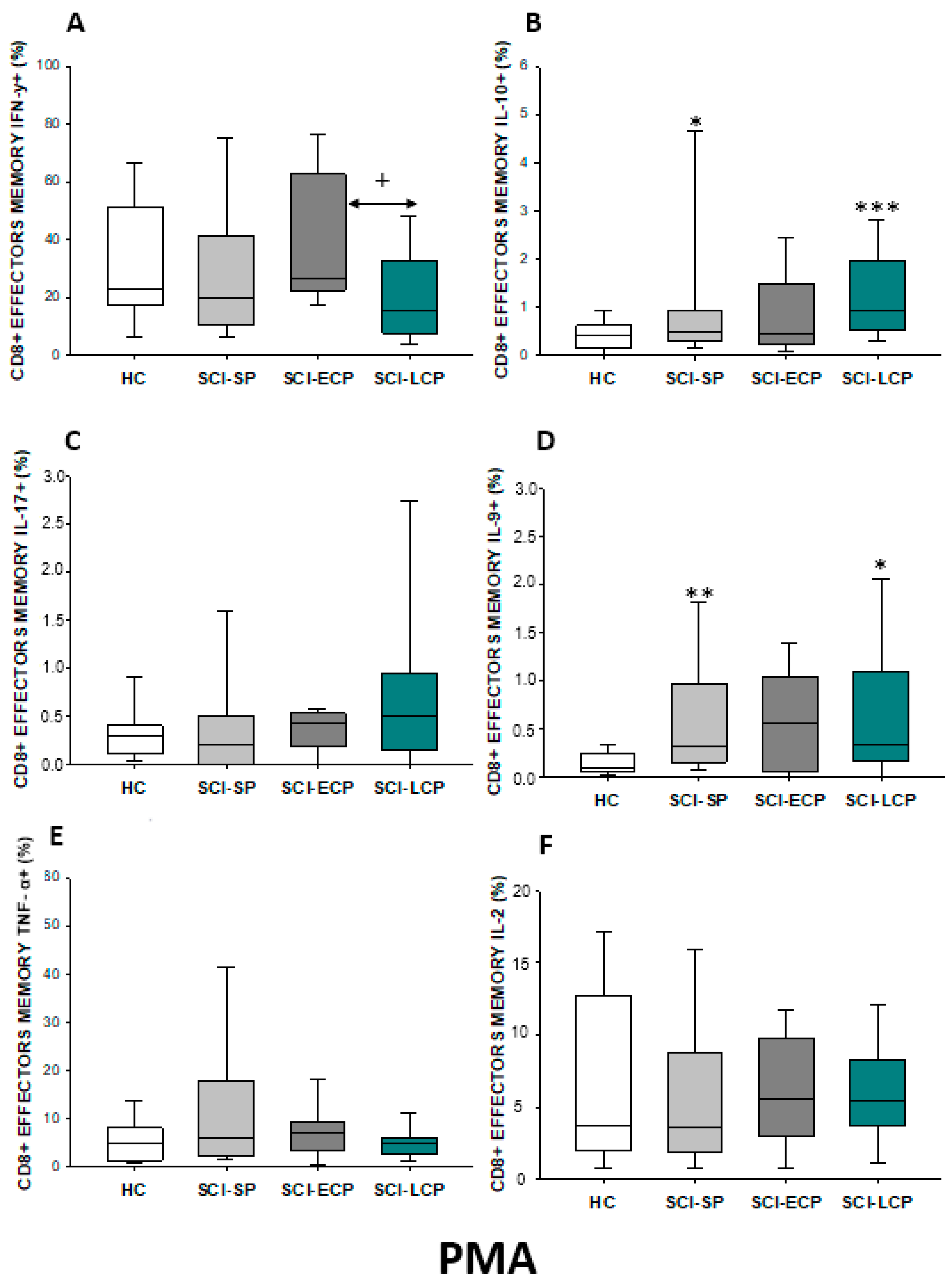
2.3.4. Cytokine Expression in CD8 Effector Memory Lymphocytes
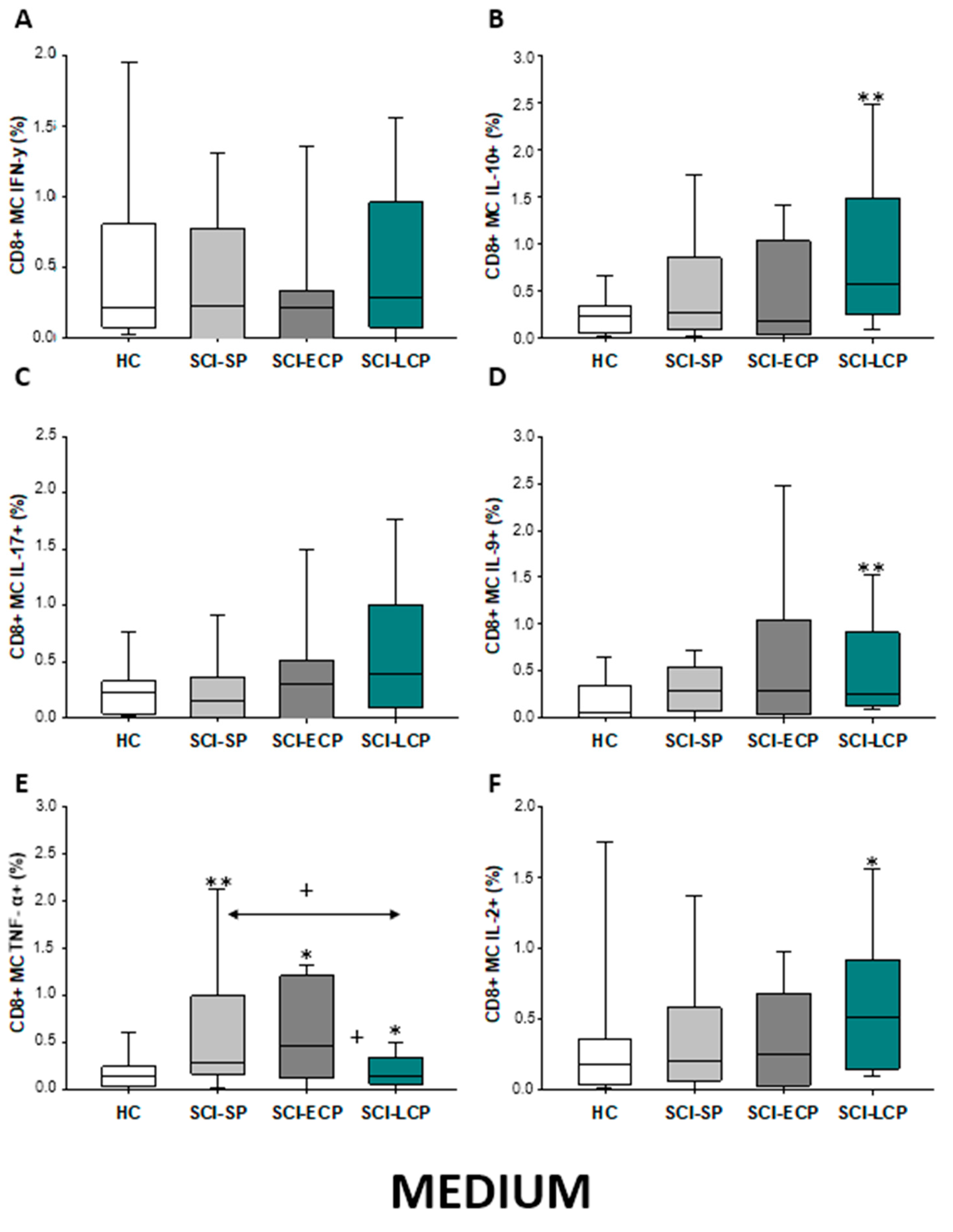
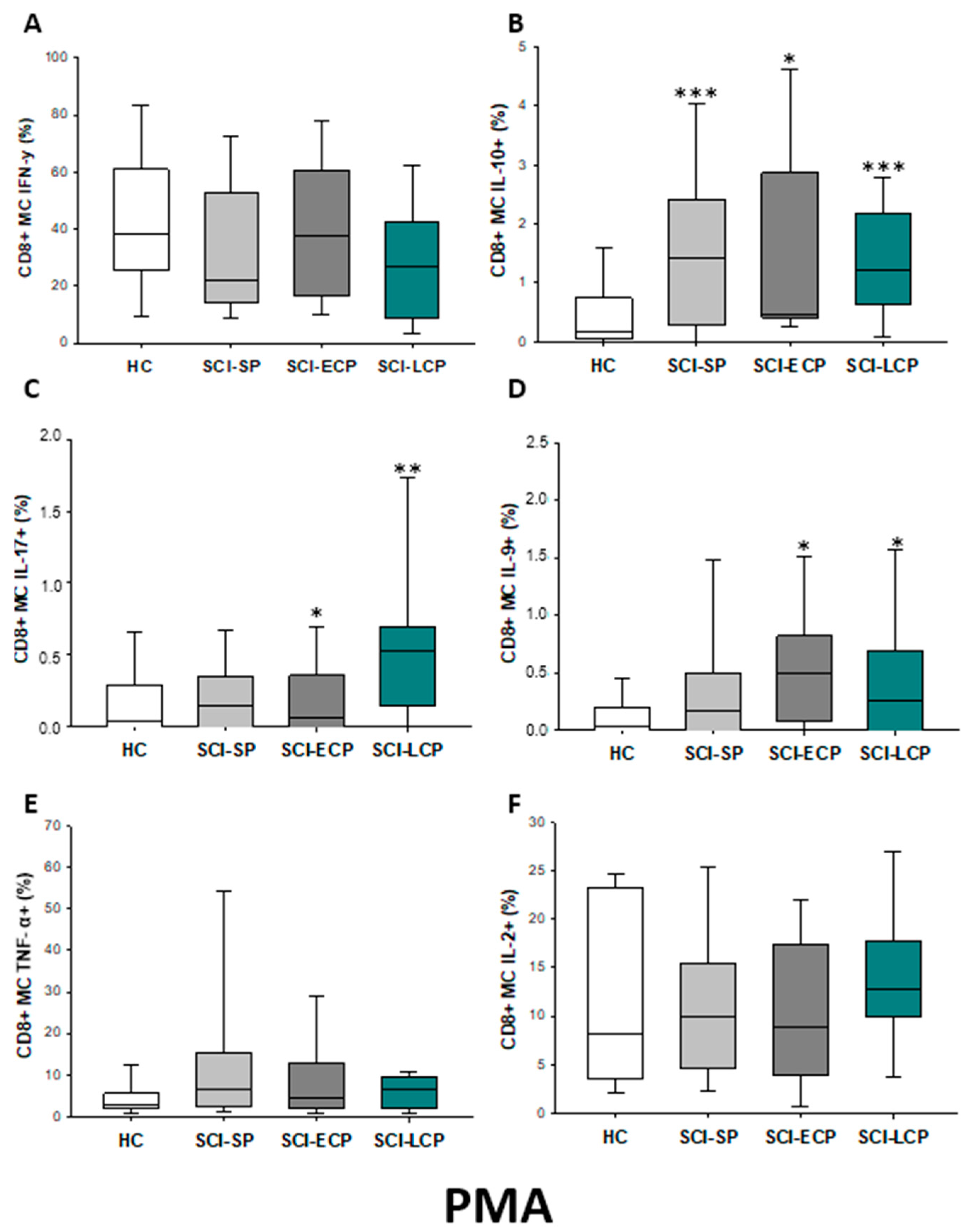
2.3.5. Cytokine Expression in CD8 Central Memory Lymphocytes
3. Discussion
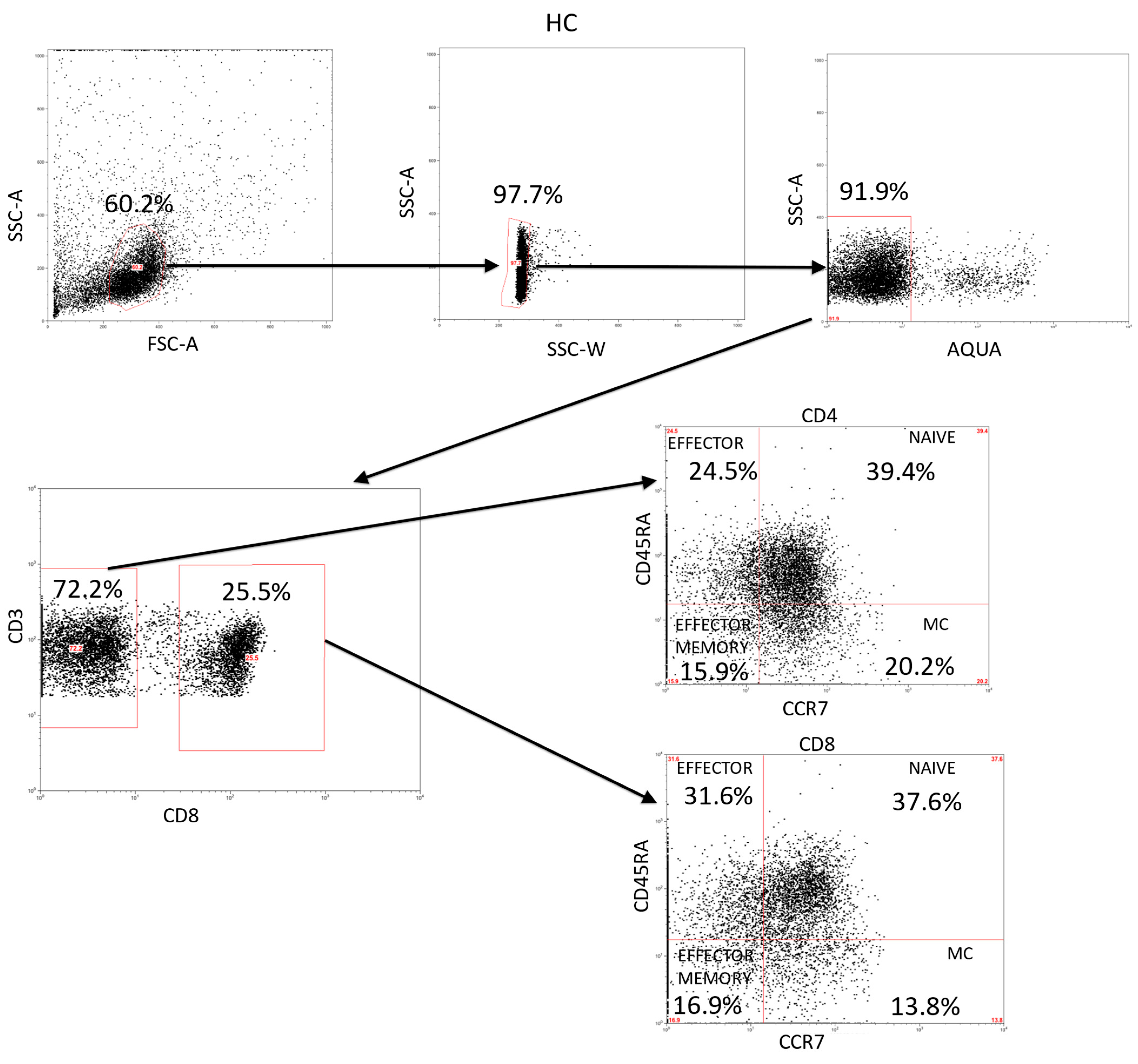
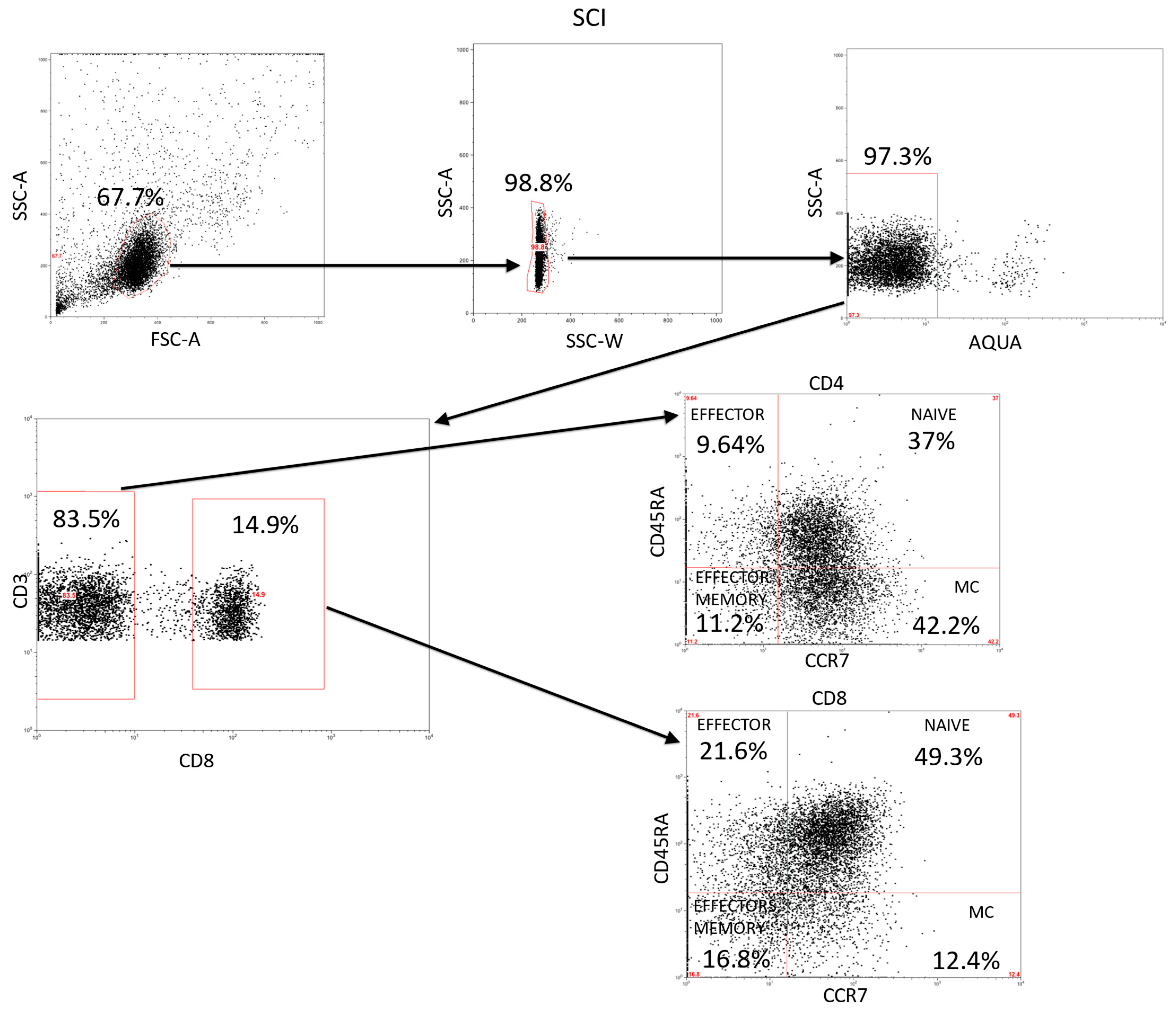
4. Materials and Methods
4.1. Study Design
4.2. Isolation of Peripheral Blood Mononuclear Cells
4.3. Immunophenotype and Intracellular Cytokines Studies
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

Appendix B

Appendix C

Appendix D

References
- Ali, Z.S.; Whitmore, R.G. Spinal Cord Injuries. In Surgical Intensive Care Medicine, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 181–193. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, Y.; Vogel, L.C.; DeVivo, M.J. Causes of Spinal Cord Injury. Top. Spinal Cord Inj. Rehabil. 2013, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Rowland, J.W.; Hawryluk, G.W.J.; Kwon, B.; Fehlings, M.G. Current status of acute spinal cord injury pathophysiology and emerging therapies: Promise on the horizon. Neurosurg. Focus 2008, 25, E2. [Google Scholar] [CrossRef]
- Anjum, A.; Yazid, M.; Daud, M.F.; Idris, J.; Hwei Ng, A.; Naicker, A.S.; Ismail, O.; Kumar, R.A.; Lokanathan, Y. Spinal Cord Injury: Pathophysiology, Multimolecular Interactions, and Underlying Recovery Mechanisms. Int. J. Mol. Sci. 2020, 21, 7533. [Google Scholar] [CrossRef] [PubMed]
- Ong, B.; Wilson, J.R.; Henzel, M.K. Management of the Patient with Chronic Spinal Cord Injury. Med. Clin. N. Am. 2020, 104, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Krause, J.S. Estimation of indirect costs based on employment and earnings changes after spinal cord injury: An observational study. Spinal Cord 2020, 58, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Malekzadeh, H.; Golpayegani, M.; Ghodsi, Z.; Sadeghi-Naini, M.; Asgardoon, M.; Baigi, V.; Vaccaro, A.R.; Rahimi-Movaghar, V. Direct Cost of Illness for Spinal Cord Injury: A Systematic Review. Glob. Spine J. 2022, 12, 1267–1281. [Google Scholar] [CrossRef]
- Kasinathan, N.; Vanathi, M.B.; Subrahmanyam, V.M.; Rao, J.V. A review on response of immune system in spinal cord injury and therapeutic agents useful in treatment. Curr. Pharm. Biotechnol. 2015, 16, 26–34. [Google Scholar] [CrossRef]
- Rodgers, K.A.; Kigerl, K.A.; Schwab, J.M.; Popovich, P.G. Immune dysfunction after spinal cord injury—A review of autonomic and neuroendocrine mechanisms. Curr. Opin. Pharmacol. 2022, 64, 102230. [Google Scholar] [CrossRef]
- Schwab, J.M.; Zhang, Y.; Kopp, M.A.; Brommer, B.; Popovich, P.G. The paradox of chronic neuroinflammation, systemic immune suppression, autoimmunity after traumatic chronic spinal cord injury. Exp. Neurol. 2014, 258, 121. [Google Scholar] [CrossRef]
- Alves, E.D.S.; Lemos, V.D.A.; da Silva, F.R.; Lira, F.S.; dos Santos, R.V.T.; Rosa, J.P.P.; Caperuto, E.; Tufik, S.; de Mello, M.T. Low-Grade Inflammation and Spinal Cord Injury: Exercise as Therapy? Mediat. Inflamm. 2013, 2013, 1–7. [Google Scholar] [CrossRef]
- Ankeny, D.P.; Lucin, K.M.; Sanders, V.M.; McGaughy, V.M.; Popovich, P.G. Spinal cord injury triggers systemic autoimmunity: Evidence for chronic B lymphocyte activation and lupus-like autoantibody synthesis. J. Neurochem. 2006, 99, 1073–1087. [Google Scholar] [CrossRef]
- Pavlicek, D.; Krebs, J.; Capossela, S.; Bertolo, A.; Engelhardt, B.; Pannek, J.; Stoyanov, J. Immunosenescence in persons with spinal cord injury in relation to urinary tract infections -a cross-sectional study-. Immun. Ageing 2017, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Guest, J.; Datta, N.; Jimsheleishvili, G.; Gater, D.R. Pathophysiology, Classification and Comorbidities after Traumatic Spinal Cord Injury. J. Pers. Med. 2022, 12, 1126. [Google Scholar] [CrossRef]
- Cardenas, D.D.; Hoffman, J.M.; Kirshblum, S.; McKinley, W. Etiology and incidence of rehospitalization after traumatic spinal cord injury: A multicenter analysis. Arch. Phys. Med. Rehabil. 2004, 85, 1757–1763. [Google Scholar] [CrossRef]
- Laginha, I.; Kopp, M.A.; Druschel, C.; Schaser, K.-D.; Brommer, B.; Hellmann, R.C.; Watzlawick, R.; Ossami-Saidi, R.-R.; Prüss, H.; Failli, V.; et al. Natural Killer (NK) Cell Functionality after human Spinal Cord Injury (SCI): Protocol of a prospective, longitudinal study. BMC Neurol. 2016, 16, 1–8. [Google Scholar] [CrossRef]
- Telegin, G.; Chernov, A.; Belogurov, A.; Balmasova, I.; Konovalov, N.; Gabibov, A. Cytokine Profile as a Marker of Cell Damage and Immune Dysfunction after Spinal Cord Injury. In Connectivity and Functional Specialization in the Brain; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Diaz, D.; Lopez-Dolado, E.; Haro, S.; Monserrat, J.; Martinez-Alonso, C.; Balomeros, D.; Albillos, A.; Alvarez-Mon, M. Systemic Inflammation and the Breakdown of Intestinal Homeostasis Are Key Events in Chronic Spinal Cord Injury Patients. Int. J. Mol. Sci. 2021, 22, 744. [Google Scholar] [CrossRef] [PubMed]
- Hellenbrand, D.J.; Quinn, C.M.; Piper, Z.J.; Morehouse, C.N.; Fixel, J.A.; Hanna, A.S. Inflammation after spinal cord injury: A review of the critical timeline of signaling cues and cellular infiltration. J. Neuroinflamm. 2021, 18, 1–16. [Google Scholar] [CrossRef]
- Fabbri, M.; Smart, C.; Pardi, R. T lymphocytes. Int. J. Biochem. Cell Biol. 2003, 35, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Sauls, R.S.; McCausland, C.; Taylor, B.N. Histology, T-Cell Lymphocyte; StatPearls: Tampa, FL, USA, 2022. [Google Scholar]
- Dong, C. Cytokine Regulation and Function in T Cells. Annu. Rev. Immunol. 2021, 39, 51–76. [Google Scholar] [CrossRef]
- Monahan, R.; Stein, A.B.; Gibbs, K.; Bank, M.; Bloom, O. Circulating T cell subsets are altered in individuals with chronic spinal cord injury. Immunol. Res. 2015, 63, 3–10. [Google Scholar] [CrossRef]
- Norden, D.M.; Qatanani, A.; Bethea, J.R.; Jiang, J. Chronic spinal cord injury impairs primary CD8 T cell antiviral immunity but does not affect generation or function of memory CD8 T cells. Exp. Neurol. 2019, 317, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Zha, J.; Smith, A.; Andreansky, S.; Bracchi-Ricard, V.; Bethea, J.R. Chronic thoracic spinal cord injury impairs CD8+ T-cell function by up-regulating programmed cell death-1 expression. J. Neuroinflamm. 2014, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Raposo, C.; Graubardt, N.; Cohen, M.; Eitan, C.; London, A.; Berkutzki, T.; Schwartz, M. CNS Repair Requires Both Effector and Regulatory T Cells with Distinct Temporal and Spatial Profiles. J. Neurosci. 2014, 34, 10141–10155. [Google Scholar] [CrossRef]
- Ogurcov, S.; Shulman, I.; Garanina, E.; Sabirov, D.; Baichurina, I.; Kuznetcov, M.; Masgutova, G.; Kostennikov, A.; Rizvanov, A.; James, V.; et al. Blood Serum Cytokines in Patients with Subacute Spinal Cord Injury: A Pilot Study to Search for Biomarkers of Injury Severity. Brain Sci. 2021, 11, 322. [Google Scholar] [CrossRef]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic spinal cord injury: An overview of pathophysiology, models and acute injury mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef]
- Garcia-Arguello, L.Y.; O’Horo, J.C.; Farrell, A.; Blakney, R.; Sohail, M.R.; Evans, C.T.; Safdar, N. Infections in the spinal cord-injured population: A systematic review. Spinal Cord 2017, 55, 526–534. [Google Scholar] [CrossRef]
- Jeffries, M.A.; Tom, V.J. Peripheral Immune Dysfunction: A Problem of Central Importance after Spinal Cord Injury. Biology 2021, 10, 928. [Google Scholar] [CrossRef]
- Ibarra, A.; Jimenez, A.M.; Cortes, C.A.F.; Correa, D. Influence of the intensity, level and phase of spinal cord injury on the proliferation of T cells and T-cell-dependent antibody reactions in rats. Spinal Cord 2006, 45, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Grütz, G.; Warszawska, K.; Kirsch, S.; Witte, E.; Wolk, K.; Geginat, J. Biology of interleukin-10. Cytokine Growth Factor Rev. 2010, 21, 331–344. [Google Scholar] [CrossRef]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Rojas, J.M.; Avia, M.; Martín, V.; Sevilla, N. IL-10: A Multifunctional Cytokine in Viral Infections. J. Immunol. Res. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef]
- Chakraborty, S.; Kubatzky, K.F.; Mitra, D.K. An Update on Interleukin-9: From Its Cellular Source and Signal Transduction to Its Role in Immunopathogenesis. Int. J. Mol. Sci. 2019, 20, 2113. [Google Scholar] [CrossRef] [PubMed]
- Bank, M.; Stein, A.; Sison, C.; Glazer, A.; Jassal, N.; McCarthy, D.; Shatzer, M.; Hahn, B.; Chugh, R.; Davies, P.; et al. Elevated Circulating Levels of the Pro-Inflammatory Cytokine Macrophage Migration Inhibitory Factor in Individuals With Acute Spinal Cord Injury. Arch. Phys. Med. Rehabil. 2015, 96, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Kalliolias, G.; Ivashkiv, L.B. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef]
- Chi, L.-Y.; Yu, J.; Zhu, H.; Li, X.-G.; Zhu, S.-G.; Kindy, M.S. The dual role of tumor necrosis factor-alpha in the pathophysiology of spinal cord injury. Neurosci. Lett. 2008, 438, 174–179. [Google Scholar] [CrossRef]
- Hayes, K.; Hull, T.; Delaney, G.; Potter, P.; Sequeira, K.; Campbell, K.; Popovich, P. Elevated Serum Titers of Proinflammatory Cytokines and CNS Autoantibodies in Patients with Chronic Spinal Cord Injury. J. Neurotrauma 2004, 19, 753–761. [Google Scholar] [CrossRef]
- Davies, A.L.; Hayes, K.C.; Dekaban, G.A. Clinical Correlates of Elevated Serum Concentrations of Cytokines and Autoantibodies in Patients With Spinal Cord Injury. Arch. Phys. Med. Rehabil. 2007, 88, 1384–1393. [Google Scholar] [CrossRef]
- Becher, B.; Spath, S.; Goverman, J. Cytokine networks in neuroinflammation. Nat. Rev. Immunol. 2016, 17, 49–59. [Google Scholar] [CrossRef]
- Zong, S.; Zeng, G.; Fang, Y.; Peng, J.; Tao, Y.; Li, K.; Zhao, J. The Role of IL-17 Promotes Spinal Cord Neuroinflammation via Activation of the Transcription Factor STAT3 after Spinal Cord Injury in the Rat. Mediat. Inflamm. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Miyajima, H.; Itokazu, T.; Tanabe, S.; Yamashita, T. Interleukin-17A regulates ependymal cell proliferation and functional recovery after spinal cord injury in mice. Cell Death Dis. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Roberts, T.T.; Leonard, G.R.; Cepela, D.J. Classifications In Brief: American Spinal Injury Association (ASIA) Impairment Scale. Clin. Orthop. Relat. Res. 2017, 475, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Biering-Sørensen, F.; Charlifue, S.; DeVivo, M.; Noonan, V.; Post, M.; Stripling, T.; Wing, P. International Spinal Cord Injury Data Sets. Spinal Cord 2006, 44, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Kirshblum, S.C.; Burns, S.P.; Biering-Sørensen, F.; Donovan, W.; Graves, D.E.; Jha, A.; Johansen, M.; Jones, L.; Krassioukov, A.; Mulcahey, M.J.; et al. International standards for neurological classification of spinal cord injury (Revised 2011). J. Spinal Cord Med. 2011, 34, 535–546. [Google Scholar] [CrossRef] [PubMed]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | HC (n = 38) | SCI (n = 105) | SCI-SP (n = 31) | SCI-ECP (n = 32) | SCI-LCP (n = 42) |
|---|---|---|---|---|---|
| Age (years) | 25.41 ± 7.99 | 36.33 ± 13.24 | 29.24 ± 14.40 | 36.81 ± 12.26 | 40.28 ± 17.65 |
| Sex (men/women) | 42.10%/57.90% | 74.28%/25.72% | 90.32%/9.68% | 77.78%/22.22% | 61.70%/38.30% |
| Time of injury (years) | 13.24 ± 9.47 | 2.30 ± 1.54 | 10.11 ± 2.55 | 22.26 ± 5.33 | |
| ASIA | |||||
| A | 46.67% | 41.93% | 55.55% | 44.68% | |
| B | 16.19% | 3.70% | 3.70% | 21.28% | |
| C | 16.19% | 18.51% | 18.52% | 19.15% | |
| D | 20.95% | 22.22% | 22.22% | 14.89% | |
| Injury level | |||||
| C1–C4 | 23.80% | 22.22% | 22.22% | 14.89% | |
| C5–C8 | 20.00% | 18.51% | 18.52% | 25.53% | |
| T1–T6 | 26.27% | 25.92% | 25.93% | 29.79% | |
| T7–T12 | 20.95% | 29.62% | 29.63% | 14.89% | |
| L1–L6 | 8.57% | 3.70% | 3.70% | 14.89% |
| CD4 Lymphocytes | CD4 Naïve | CD4 Effector | CD4 Effector Memory | CD4 Central Memory | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Medium | PMA | Medium | PMA | Medium | PMA | Medium | PMA | Medium | PMA | |
| IFN-γ | __________ | __________ | __________ | __________ | __________ | __________ | __________ | __________ | __________ | ↓ in SCI-LCP and SCI-ECP versus HC |
| IL-10 | ↑ in SCI-LCP when compared to HC and SCI-SP | ↑ in SCI-LCP and SCI-SP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP and SCI-SP when compared to HC | ↑ in SCI-LCP when compared to HC and SCI-SP (also ↑ relative to HC) | ↑ in SCI-LCP and SCI-SP when compared to HC | ↑ in SCI-LCP and SCI-SP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP and SCI-SP when compared to HC | ↑ in SCI-LCP and SCI-SP when compared to HC |
| IL-17 | __________ | __________ | ↑ in SCI-LCP and SCI-ECP when compared to HC | __________ | __________ | __________ | __________ | __________ | __________ | __________ |
| IL-9 | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP and SCI-ECP when compared to HC | ↑ in SCI-LCP when compared to HC | __________ | __________ | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP and SCI-ECP when compared to HC | ↑ in SCI-LCP when compared to HC | __________ |
| TNF-α | ↑ in SCI-SP when compared to HC | ↑ in SCI-SP when compared to HC | __________ | ↑ in SCI-SP when compared to HC | __________ | __________ | __________ | __________ | __________ | ↑ in SCI-SP when compared to HC and with SCI-LCP |
| IL-2 | __________ | __________ | __________ | __________ | __________ | __________ | __________ | __________ | __________ | __________ |
| CD8 Lymphocytes | CD8 Naïve | CD8 Effector | CD8 Effector Memory | CD8 Central Memory | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Medium | PMA | Medium | PMA | Medium | PMA | Medium | PMA | Medium | PMA | |
| IFN-γ | ↑ in SCI-LCP when compared to HC | __________ | ↑ in SCI-LCP when compared to HC | __________ | __________ | ↑ in SCI-ECP when compared to SCI-SP | __________ | ↓ in SCI-LCP when compared to SCI-ECP | __________ | __________ |
| IL-10 | ↑ in SCI-LCP when compared to HC and SCI-SP | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP when compared to HC and SCI-SP | ↑ in SCI-LCP and SCI-ECP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP and SCI-SP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP, SCI-ECP, and SCI-SP when compared to HC |
| IL-17 | __________ | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP when compared to HC | __________ | __________ | __________ | __________ | __________ | ↑ in SCI-LCP and SCI-ECP when compared to HC |
| IL-9 | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP, SCI-ECP, and SCI-SP when compared to HC | ↑ in SCI-LCP and SCI-ECP when compared to HC | ↑ in SCI-LCP, SCI-ECP, and SCI-SP when compared to HC | ↑ in SCI-LCP and SCI-SP when compared to HC | ↑ in SCI-LCP, SCI-ECP, and SCI-SP when compared to HC | ↑ in SCI-LCP when compared to HC and SCI-ECP | ↑ in SCI-LCP and SCI-SP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-LCP and SCI-ECP when compared to HC |
| TNF-α | __________ | ↑ in SCI-SP when compared to HC | ↑ in SCI-LCP and SCI-ECP when compared to HC | ↑ in SCI-LCP when compared to HC | ↑ in SCI-ECP when compared to SCI-SP and SCI-LCP | __________ | __________ | __________ | ↑ SCI-ECP and SCI-SP when compared to HC and SCI-LCP (also ↑ relative to HC) | ↑ in SCI-SP when compared to HC and with SCI-LCP |
| IL-2 | __________ | __________ | __________ | __________ | ↑ in SCI-LCP when compared to HC | __________ | ↑ in SCI-LCP when compared to HC | __________ | ↑ in SCI-LCP when compared to HC | __________ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Girón, S.H.; Gómez-Lahoz, A.M.; Sanz, J.M.; Fraile-Martínez, O.; Jiménez, D.J.; Garcia-Montero, C.; de Leon-Oliva, D.; Ortega, M.A.; Atienza-Perez, M.; Diaz, D.; et al. Patients with Chronic Spinal Cord Injury and a Long Period of Evolution Exhibit an Altered Cytokine Production by CD4 and CD8 T Cell Populations. Int. J. Mol. Sci. 2023, 24, 7048. https://doi.org/10.3390/ijms24087048
Girón SH, Gómez-Lahoz AM, Sanz JM, Fraile-Martínez O, Jiménez DJ, Garcia-Montero C, de Leon-Oliva D, Ortega MA, Atienza-Perez M, Diaz D, et al. Patients with Chronic Spinal Cord Injury and a Long Period of Evolution Exhibit an Altered Cytokine Production by CD4 and CD8 T Cell Populations. International Journal of Molecular Sciences. 2023; 24(8):7048. https://doi.org/10.3390/ijms24087048
Chicago/Turabian StyleGirón, Sergio Haro, Ana M. Gómez-Lahoz, Jorge Monserrat Sanz, Oscar Fraile-Martínez, Diego J. Jiménez, Cielo Garcia-Montero, Diego de Leon-Oliva, Miguel A. Ortega, Mar Atienza-Perez, David Diaz, and et al. 2023. "Patients with Chronic Spinal Cord Injury and a Long Period of Evolution Exhibit an Altered Cytokine Production by CD4 and CD8 T Cell Populations" International Journal of Molecular Sciences 24, no. 8: 7048. https://doi.org/10.3390/ijms24087048
APA StyleGirón, S. H., Gómez-Lahoz, A. M., Sanz, J. M., Fraile-Martínez, O., Jiménez, D. J., Garcia-Montero, C., de Leon-Oliva, D., Ortega, M. A., Atienza-Perez, M., Diaz, D., Lopez-Dolado, E., & Álvarez-Mon, M. (2023). Patients with Chronic Spinal Cord Injury and a Long Period of Evolution Exhibit an Altered Cytokine Production by CD4 and CD8 T Cell Populations. International Journal of Molecular Sciences, 24(8), 7048. https://doi.org/10.3390/ijms24087048