
Overexpression of Pericentromeric HSAT2 DNA Increases Expression of EMT Markers in Human Epithelial Cancer Cell Lines

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Chromosomal Localization of Transcribed TR Satellite DNA
2.2. Designing a Plasmid Overexpressing HSAT2
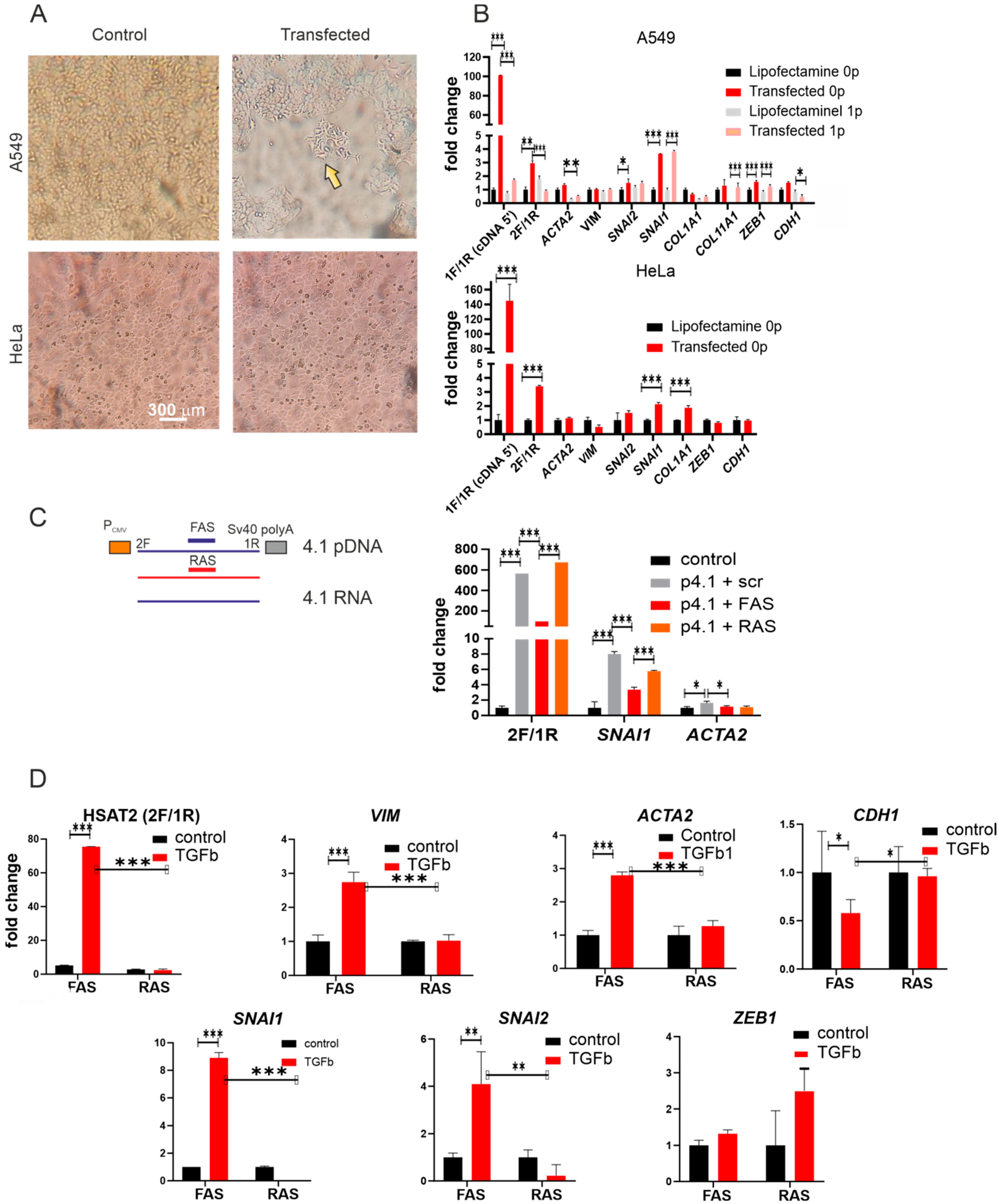
2.3. Influence of the Overexpression on Epithelial–Mesenchymal Transition
2.4. Knockdown of the HSAT2 Transcript Decreased TGFβ1-Induced Upregulation of EMT Gene Transcription
3. Discussion
3.1. Genomic Mapping of the HSAT2 Transcript
3.2. The Influence of HSAT2 Transcription on EMT
4. Materials and Methods
4.1. Cell Cultures
4.2. Oligonucleotides
4.3. Computational Analysis
4.4. Cloning
4.5. Transfection
4.6. PCR and qPCR
4.7. Fluorescent In Situ Hybridization (FISH)
4.8. Confocal Microscopy
4.9. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richard, G.-F.; Kerrest, A.; Dujon, B. Comparative Genomics and Molecular Dynamics of DNA Repeats in Eukaryotes. Microbiol. Mol. Biol. Rev. 2008, 72, 686–727. [Google Scholar] [CrossRef] [PubMed]
- Corneo, G.; Ginelli, E.; Polli, E. A Satellite DNA Isolated from Human Tissues. J. Mol. Biol. 1967, 23, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Prosser, J.; Frommer, M.; Paul, C.; Vincent, P.C. Sequence Relationships of Three Human Satellite DNAs. J. Mol. Biol. 1986, 187, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Jeanpierre, M. Human satellites 2 and 3. Ann. Genet. 1994, 37, 163–171. [Google Scholar]
- Miga, K.H. Centromeric Satellite DNAs: Hidden Sequence Variation in the Human Population. Genes 2019, 10, 352. [Google Scholar] [CrossRef]
- Valgardsdottir, R.; Chiodi, I.; Giordano, M.; Cobianchi, F.; Riva, S.; Biamonti, G. Structural and functional characterization of noncoding repetitive RNAs transcribed in stressed human cells. Mol. Biol. Cell 2005, 16, 2597–2604. [Google Scholar] [CrossRef]
- Altemose, N. A classical revival: Human satellite DNAs enter the genomics era. Semin. Cell Dev. Biol. 2022, 128, 2–14. [Google Scholar] [CrossRef]
- Podgornaya, O.; Dey, R.; Lobov, I.B.; Enukashvili, N. Human satellite 3 (HS3) binding protein from the nuclear matrix: Isolation and binding properties. Biochim. Biophys. Acta 2000, 1497, 204–214. [Google Scholar] [CrossRef]
- McGowan-Jordan, J.; Hastings, R.; Moore, S. (Eds.) ISCN 2020; S. Karger AG: Basel, Switzerland, 2020; ISBN 978-3-318-06706-4. [Google Scholar]
- Probst, A.V.; Okamoto, I.; Casanova, M.; El Marjou, F.; Le Baccon, P.; Almouzni, G. A Strand-Specific Burst in Transcription of Pericentric Satellites Is Required for Chromocenter Formation and Early Mouse Development. Dev. Cell 2010, 19, 625–638. [Google Scholar] [CrossRef]
- Solovei, I.V.; Joffe, B.I.; Gaginskaya, E.R.; Macgregor, H.C. Transcription on lampbrush chromosomes of a centromerically localized highly repeated DNA in pigeon (Columba) relates to sequence arrangement. Chromosom. Res. 1996, 4, 588–603. [Google Scholar] [CrossRef]
- Rizzi, N.; Denegri, M.; Chiodi, I.; Corioni, M.; Valgardsdottir, R.; Cobianchi, F.; Riva, S.; Biamonti, G. Transcriptional activation of a constitutive heterochromatic domain of the human genome in response to heat shock. Mol. Biol. Cell 2004, 15, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Enukashvily, N.I.; Donev, R.; Waisertreiger, I.S.-R.; Podgornaya, O.I. Human chromosome 1 satellite 3 DNA is decondensed, demethylated and transcribed in senescent cells and in A431 epithelial carcinoma cells. Cytogenet. Genome Res. 2007, 118, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Enukashvily, N.I.; Malashicheva, A.B.; Waisertreiger, I.S.-R. Satellite DNA spatial localization and transcriptional activity in mouse embryonic E-14 and IOUD2 stem cells. Cytogenet. Genome Res. 2009, 124, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.L.; Byron, M.; Carone, D.M.; Whitfield, T.W.; Pouliot, G.P.; Fischer, A.; Jones, P.; Lawrence, J.B. Demethylated HSATII DNA and HSATII RNA Foci Sequester PRC1 and MeCP2 into Cancer-Specific Nuclear Bodies. Cell Rep. 2017, 18, 2943–2956. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.; Metz, A.; Govin, J.; Vigneron, M.; Turner, B.M.; Khochbin, S.; Vourc’h, C. Stress-induced transcription of satellite III repeats. J. Cell Biol. 2004, 164, 25–33. [Google Scholar] [CrossRef]
- Dobrynin, M.A.; Korchagina, N.M.; Prjibelski, A.D.; Shafranskaya, D. Human pericentromeric tandemly repeated DNA is transcribed at the end of oocyte maturation and is associated with membraneless mitochondria associated structures. Sci. Rep. 2020, 10, 19634. [Google Scholar] [CrossRef]
- Dobrynin, M.A.; Korchagina, N.M.; Ponomartsev, N.V.; Podgornaya, O.I.; Enukashvily, N.I. Influence of Inactivation of Tandemly Repeated Pericentromeric DNA Transcription on the Formation of Membraneless Structures at the End of Oocyte Maturation. Russ. J. Dev. Biol. 2022, 53, 128–133. [Google Scholar] [CrossRef]
- Enukashvily, N.I.; Ponomartsev, N.V.; Ketkar, A.; Suezov, R.; Chubar, A.V.; Prjibelski, A.D.; Shafranskaya, D.D.; Elmshäuser, S.; Keber, C.U.; Stefanova, V.N.; et al. Pericentromeric satellite lncRNAs are induced in cancer-associated fibroblasts and regulate their functions in lung tumorigenesis. Cell Death Dis. 2023, 14, 19. [Google Scholar] [CrossRef]
- Liehr, T. Chromosomal Heteromorphisms and Cancer Susceptibility Revisited. Cells 2022, 11, 3239. [Google Scholar] [CrossRef]
- Nurk, S.; Koren, S.; Rhie, A.; Rautiainen, M.; Bzikadze, A.V.; Mikheenko, A.; Vollger, M.R.; Altemose, N.; Uralsky, L.; Gershman, A.; et al. The complete sequence of a human genome. Science 2022, 376, 44–53. [Google Scholar] [CrossRef]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The Human Genome Browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Ostell, J.; Pruitt, K.D.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2020, 48, D84–D86. [Google Scholar] [CrossRef] [PubMed]
- Altemose, N.; Logsdon, G.A.; Bzikadze, A.V.; Sidhwani, P.; Langley, S.A.; Caldas, G.V.; Hoyt, S.J.; Uralsky, L.; Ryabov, F.D.; Shew, C.J.; et al. Complete genomic and epigenetic maps of human centromeres. Science 2022, 376, eabl4178. [Google Scholar] [CrossRef] [PubMed]
- Ting, D.T.; Lipson, D.; Paul, S.; Brannigan, B.W.; Akhavanfard, S.; Coffman, E.J.; Contino, G.; Deshpande, V.; Iafrate, A.J.; Letovsky, S.; et al. Aberrant overexpression of satellite repeats in pancreatic and other epithelial cancers. Science 2011, 331, 593–596. [Google Scholar] [CrossRef]
- Solovyov, A.; Vabret, N.; Arora, K.S.; Snyder, A.; Funt, S.A.; Bajorin, D.F.; Rosenberg, J.E.; Bhardwaj, N.; Ting, D.T.; Greenbaum, B.D. Global Cancer Transcriptome Quantifies Repeat Element Polarization between Immunotherapy Responsive and T Cell Suppressive Classes. Cell Rep. 2018, 23, 512–521. [Google Scholar] [CrossRef]
- Enukashvily, N.I.; Semenova, N.; Chubar, A.V.; Ostromyshenskii, D.I.; Gushcha, E.A.; Gritsaev, S.; Bessmeltsev, S.S.; Rugal, V.I.; Prikhodko, E.M.; Kostroma, I.; et al. Pericentromeric Non-Coding DNA Transcription Is Associated with Niche Impairment in Patients with Ineffective or Partially Effective Multiple Myeloma Treatment. Int. J. Mol. Sci. 2022, 23, 3359. [Google Scholar] [CrossRef]
- Enukashvily, N.I.; Ponomartsev, N.V. Mammalian satellite DNA: A speaking dumb. Adv. Protein Chem. Struct. Biol. 2013, 90, 31–65. [Google Scholar] [CrossRef]
- Altemose, N.; Miga, K.H.; Maggioni, M.; Willard, H.F. Genomic Characterization of Large Heterochromatic Gaps in the Human Genome Assembly. PLoS Comput. Biol. 2014, 10, 3359. [Google Scholar] [CrossRef]
- Khudiakov, A.A.; Smolina, N.A.; Perepelina, K.I.; Malashicheva, A.B.; Kostareva, A.A. Extracellular MicroRNAs and Mitochondrial DNA as Potential Biomarkers of Arrhythmogenic Cardiomyopathy. Biochemistry 2019, 84, 272–282. [Google Scholar] [CrossRef]
- Lecce, L.; Xu, Y.; V’Gangula, B.; Chandel, N.; Pothula, V.; Caudrillier, A.; Santini, M.P.; D’Escamard, V.; Ceholski, D.K.; Gorski, P.A.; et al. Histone deacetylase 9 promotes endothelial-mesenchymal transition and an unfavorable atherosclerotic plaque phenotype. J. Clin. Investig. 2021, 131, 1–18. [Google Scholar] [CrossRef]
- Cassier, E.; Gallay, N.; Bourquard, T.; Claeysen, S.; Bockaert, J.; Crépieux, P.; Poupon, A.; Reiter, E.; Marin, P.; Vandermoere, F. Phosphorylation of β-arrestin2 at Thr383 by MEK underlies β-arrestin-dependent activation of Erk1/2 by GPCRs. Elife 2017, 6, e23777. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2014, 13, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Takeyama, Y.; Sato, M.; Horio, M.; Hase, T.; Yoshida, K.; Yokoyama, T.; Nakashima, H.; Hashimoto, N.; Sekido, Y.; Gazdar, A.F.; et al. Knockdown of ZEB1, a master epithelial-to-mesenchymal transition (EMT) gene, suppresses anchorage-independent cell growth of lung cancer cells. Cancer Lett. 2010, 296, 216–224. [Google Scholar] [CrossRef]
- Kit, S. Equilibrium sedimentation in density gradients of DNA preparations from animal tissues. J. Mol. Biol. 1961, 3, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.; Louzada, S.; Gama-Carvalho, M.; Chaves, R. Genomic tackling of human satellite dna: Breaking barriers through time. Int. J. Mol. Sci. 2021, 22, 4707. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Identifying centromeric satellites with DNA-brnn. Bioinformatics 2019, 35, 4408–4410. [Google Scholar] [CrossRef] [PubMed]
- Nonin-Lecomte, S.; Leroy, J.L. Structure of a C-rich strand fragment of the human centromeric satellite III: A pH-dependent intercalation topology. J. Mol. Biol. 2001, 309, 491–506. [Google Scholar] [CrossRef]
- Valgardsdottir, R.; Chiodi, I.; Giordano, M.; Rossi, A.; Bazzini, S.; Ghigna, C.; Riva, S.; Biamonti, G. Transcription of Satellite III non-coding RNAs is a general stress response in human cells. Nucleic Acids Res. 2008, 36, 423–434. [Google Scholar] [CrossRef]
- Bai, X.; Huang, W.; Zhang, C.; Niu, J.; Ding, W. Discovery of a regulatory motif for human satellite DNA transcription in response to batf2 overexpression. Anticancer. Res. 2016, 36, 1203–1210. [Google Scholar]
- Ugarković, Đ.; Sermek, A.; Ljubić, S.; Feliciello, I. Satellite DNAs in Health and Disease. Genes 2022, 13, 1154. [Google Scholar] [CrossRef]
- Lopes, M.; Louzada, S.; Ferreira, D.; Veríssimo, G.; Eleutério, D.; Carvalho, M.G.; Chaves, R. Human Satellite 1A analysis provides evidence of pericentromeric transcription. BMC Biol. 2023, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Enukashvily, N.I.; Dobrynin, M.A.; Chubar, A.V. RNA-Seeded Membraneless Bodies: Role of Tandemly Repeated RNA; Elsevier Ltd.: Amsterdam, The Netherlands, 2021; pp. 151–193. [Google Scholar]
- Puppo, I.L.; Saifitdinova, A.F.; Tonyan, Z.N. The Role of Satellite DNA in Causing Structural Rearrangements in Human Karyotype. Russ. J. Genet. 2020, 56, 41–47. [Google Scholar] [CrossRef]
- Zhu, Q.; Hoong, N.; Aslanian, A.; Hara, T.; Benner, C.; Heinz, S.; Miga, K.H.; Ke, E.; Verma, S.; Soroczynski, J.; et al. Heterochromatin-Encoded Satellite RNAs Induce Breast Cancer. Mol. Cell 2018, 70, 842–853.e7. [Google Scholar] [CrossRef] [PubMed]
- Giordano, M.; Infantino, L.; Biggiogera, M.; Montecucco, A.; Biamonti, G. Heat Shock A ff ects Mitotic Segregation of Human Chromosomes Bound to Stress-Induced Satellite III RNAs. Int. J. Mol. Sci. 2020, 21, 2812. [Google Scholar] [CrossRef] [PubMed]
- Landers, C.C.; Rabeler, C.A.; Ferrari, E.K.; D’Alessandro, L.R.; Kang, D.D.; Malisa, J.; Bashir, S.M.; Carone, D.M. Ectopic expression of pericentric HSATII RNA results in nuclear RNA accumulation, MeCP2 recruitment, and cell division defects. Chromosoma 2021, 130, 75–90. [Google Scholar] [CrossRef]
- Bersani, F.; Lee, E.; Kharchenko, P.V.; Xu, A.W.; Liu, M.; Xega, K.; MacKenzie, O.C.; Brannigan, B.W.; Wittner, B.S.; Jung, H.; et al. Pericentromeric satellite repeat expansions through RNA-derived DNA intermediates in cancer. Proc. Natl. Acad. Sci. USA 2015, 112, 15148–15153. [Google Scholar] [CrossRef]
- Porter, R.L.; Sun, S.; Flores, M.N.; Berzolla, E.; You, E.; Phillips, I.E.; Neelima, K.C.; Desai, N.; Tai, E.C.; Szabolcs, A.; et al. Satellite repeat RNA expression in epithelial ovarian cancer associates with a tumor-immunosuppressive phenotype. J. Clin. Investig. 2022, 132, 1–16. [Google Scholar] [CrossRef]
- Yang, X.; Qin, C.; Zhao, B.; Li, T.; Wang, Y.; Li, Z.; Li, T.; Wang, W. Long Noncoding RNA and Circular RNA: Two Rising Stars in Regulating Epithelial-Mesenchymal Transition of Pancreatic Cancer. Front. Oncol. 2022, 12, 1–13. [Google Scholar] [CrossRef]
- Savagner, P. Leaving the neighborhood: Molecular mechanisms involved during epithelial-mesenchymal transition. BioEssays 2001, 23, 912–923. [Google Scholar] [CrossRef]
- Vleminckx, K.; Vakaet, L.; Mareel, M.; Fiers, W.; van Roy, F. Genetic manipulation of E-cadherin expression by epithelial tumor cells reveals an invasion suppressor role. Cell 1991, 66, 107–119. [Google Scholar] [CrossRef]
- Guaita, S.; Puig, I.; Francí, C.; Garrido, M.; Domínguez, D.; Batlle, E.; Sancho, E.; Dedhar, S.; De Herreros, A.G.; Baulida, J. Snail induction of epithelial to mesenchymal transition in tumor cells is accompanied by MUC1 repression and ZEB1 expression. J. Biol. Chem. 2002, 277, 39209–39216. [Google Scholar] [CrossRef] [PubMed]
- You, E.; Zou, L.; Danaher, P.; Phillips, I.E.; Raabe, M.J.; Patel, B.; Pankaj, A.; North, K.; Kim, S.; Kim, Y.; et al. Abstract C038: Repeat RNA dysregulation of cellular states in the pancreatic cancer microenvironment. Cancer Res. 2022, 82, C038. [Google Scholar] [CrossRef]
- Verstappe, J.; Berx, G. A role for partial epithelial-to-mesenchymal transition in enabling stemness in homeostasis and cancer. Semin. Cancer Biol. 2023, 90, 132949. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Lennox, K.A.; Behlke, M.A. Chemical modification and design of anti-miRNA oligonucleotides. Gene Ther. 2011, 18, 1111–1120. [Google Scholar] [CrossRef]
- Adachi, H.; Hengesbach, M.; Yu, Y.T.; Morais, P. From antisense RNA to RNA modification: Therapeutic potential of RNA-based technologies. Biomedicines 2021, 9, 1–26. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosome | Total Number of Hits | CEN Satellite DNA Annotation * | Number of Hits | Strand | Chromosome Band |
|---|---|---|---|---|---|
| chr1 | 7876 | hsat2_1_1(B) | 3 | − | 1q12 |
| hsat2_1_3(A1,B) | 411 | + | 1q12 | ||
| hsat2_1_4(B,A1,A2) | 23 | + | 1q12 | ||
| hsat2_1_5(B) | 2 | + | 1q12 | ||
| hsat2_1_6(A2,A1,B) | 7437 | − | 1q12 | ||
| chr2 | 118 | hsat2_2_2(B) | 9 | +/− | 2p11.2 |
| hsat2_2_3(B) | 5 | +/− | 2p11.2 | ||
| hsat2_2_4(B) | 10 | +/− | 2p11.2 | ||
| hsat2_2_5(B) | 76 | +/− | 2p11.2 | ||
| hsat2_2_6(B) | 8 | + | 2p11.2 | ||
| hsat2_2_7(B) | 10 | − | 2q11.2 | ||
| chr7 | 83 | hsat2_7_5(B) | 10 | +/− | 7p11.2 |
| hsat2_7_6(B) | 6 | + | 7p11.2 | ||
| hsat2_7_7(B) | 6 | − | 7p11.2 | ||
| hsat2_7_10(B) | 7 | + | 7p11.2 | ||
| hsat2_7_12(B) | 2 | − | 7p11.2 | ||
| hsat2_7_13(B) | 14 | + | 7p11.2 | ||
| hsat2_7_15(B) | 3 | − | 7p11.2 | ||
| hsat2_7_16(B) | 6 | − | 7p11.2 | ||
| hsat2_7_17(B) | 18 | + | 7p11.2 | ||
| hsat2_7_18(B) | 10 | − | 7p11.2 | ||
| hsat2_7_19(B) | 1 | + | 7q11.21 | ||
| chr9 | 10 | hsat2_9_1(B) | 10 | + | 9p11.2 |
| chr10 | 273 | hsat2_10_1(B) | 6 | − | 10p11.21 |
| hsat2_10_2(A1,A2) | 9 | + | 10q11.21 | ||
| hsat2_10_3(A1,A2) | 247 | − | 10q11.21 | ||
| hsat2_10_4(B) | 1 | − | 10q11.21 | ||
| chr16 | 3833 | hsat2_16_3(B) | 1 | +/− | 16p11.2 |
| hsat2_16_4(B) | 4 | + | 16p11.2 | ||
| hsat2_16_5(B) | 10 | − | 16q11.2 | ||
| hsat2_16_6(B) | 4 | − | 16q11.2 | ||
| hsat2_16_7(B) | 9 | +/− | 16q11.2 | ||
| hsat2_16_8(B) | 2 | − | 16q11.2 | ||
| hsat2_16_9(B) | 12 | − | 16q11.2 | ||
| hsat2_16_10(B) | 1 | − | 16q11.2 | ||
| hsat2_16_11(B) | 11 | − | 16q11.2 | ||
| hsat2_16_12(B) | 12 | − | 16q11.2 | ||
| hsat2_16_13(B) | 2 | − | 16q11.2 | ||
| hsat2_16_15(B) | 3765 | +/− | 16q11.2 | ||
| chr17 | 31 | hsat2_17_1(B) | 2 | + | 17p11.2 |
| hsat2_17_2(B) | 29 | + | 17p11.2 | ||
| chr22 | 48 | hsat2_22_2(B) | 4 | + | 22q11.21 |
| hsat2_22_3(B) | 17 | + | 22q11.21 | ||
| hsat2_22_4(B) | 18 | + | 22q11.21 | ||
| hsat2_22_5(B) | 8 | − | 22q11.21 | ||
| chrY | 1 | ct_Y_48 | 1 | + | q11.21 |
| Total | 12,262 |
| Oligonucleotide | Sequence | Target | Reference | Application |
|---|---|---|---|---|
| 2F | CGATTCTGTTCGGTGATTCC | BK063198 OP912407.2 OP912406.2 OP912405.2 OP912404.2 OP912403.2 | Own design | qPCR |
| 1R | TGAAATCCAATATGATCATCATCGAA | qPCR | ||
| 2R | AATGGATGGACTCATCATCG | With 2F—5′ end of OP912406.2 | Own design | qPCR |
| 1F | CGTTTCCTTTCGATGGCGTT | With 1R—3′ end of OP912406.2 | Own design | qPCR |
| S1 | AGTCCATTCAATGATTCCATTCCAGT | HSAT2 transcript *** X60726.1 | [13] | qPCR |
| AS1 | AATCATCATCCAACGGAAGCTAATG | qPCR | ||
| SnailF | CTCTTTCCTCGTCAGGAAGC | SNAI1 NM_005985.4 | [30] | qPCR |
| SnailR | GGCTGCTGGAAGGTAAACTC | qPCR | ||
| SlugF | ATGAGGAATCTGGCTGCTGT | SNAI2 NM_003068.5 | [30] | qPCR |
| SlugR | CAGGAGAAAATGCCTTTGGA | qPCR | ||
| Zeb1F | GTCAGCCCTGCAGTCCAAGAACCAC | ZEB1 NM_001128128.3 | [19] | qPCR |
| Zeb1R | CCGCATTTTCTTTTTGGGCGGTGTA | qPCR | ||
| VimF | GGCTCAGATTCAGGAACAGC | VIM NM_003380.5 | [31] | qPCR |
| VimR | AGCCTCAGAGAGGTCAGCAA | qPCR | ||
| Col11A1F | GACTATCCCCTCTTCAGAACTGTTAAC | COL11A1 NM_001854.4 | [19] | qPCR |
| Col11A1R | CTTCTATCAAGTGGTTTCGTGGTTT | qPCR | ||
| Col1A1F | TCAGCAAGAACCCCAAGGACAAGAGG | COL1A1 NM_000088.4 | qPCR | |
| Col1A1R | AGGAAGGTCAGCTGGATGGCCACAT | qPCR | ||
| eCadF | GCTGGACCGAGAGAGTTTCC | CDH1 NM_004360.3 | Own design | qPCR |
| eCadR | CGACGTTAGCCTCGTTCTCA | qPCR | ||
| AgeI-2F * | GCTACCGGTCGATTCTGTTCGGTGATTCC | OP912406.2 | Cloning into an expression vector | |
| SalI-1R * | CGTCGACTGCTGAAATCCAATATGATCATCATCGAA | |||
| GAPDHF | AGGTCGGAGTCAACGGATTT | NM_002046.7 | Own design | Reference gene for qPCR |
| GAPDHR | TTCCCGTTCTCAGCCTTGAC | |||
| FAS ** | meAmeAmeUmeGlnGlnAlnAlnUlnClnGlnAlnAlnUlnGlnGmeAmeAmeUmeC | OP912407.2 OP912406.2 OP912405.2 OP912404.2 OP912403.2 X60726.1 | Own design | HSAT2 2F/1R cloned transcript knockdown |
| RAS ** | meGmeAmeUmeUlnClnClnAlnUlnUlnClnGlnAlnUlnUlnCmeCmeAmeUmeU | Own design | HSAT2 2F/1R cloned transcript’s reverse strand knockdown | |
| Scr | meAmeAmeUmeUmeCmeUmeCmeCmeGmeAmeAmeCmeGmeUmeGmeUmeCmeAmeCmeGmeU | Scramble sequence for knockdown | [32] | A control sequence in knockdown experiments |
| Tr-3′ | Cy3—GATGGCGTTTCCATTCGATTCCATTCGATGTTGATTCCATTTGTTTCCATTGGATGATGATTCCGTTCGTGTCCATTCGA | BK063198, OP912407.2 OP912406.2 OP912405.2 OP912404.2 OP912403.2 | Own design | RNA-FISH |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponomartsev, N.; Zilov, D.; Gushcha, E.; Travina, A.; Sergeev, A.; Enukashvily, N. Overexpression of Pericentromeric HSAT2 DNA Increases Expression of EMT Markers in Human Epithelial Cancer Cell Lines. Int. J. Mol. Sci. 2023, 24, 6918. https://doi.org/10.3390/ijms24086918
Ponomartsev N, Zilov D, Gushcha E, Travina A, Sergeev A, Enukashvily N. Overexpression of Pericentromeric HSAT2 DNA Increases Expression of EMT Markers in Human Epithelial Cancer Cell Lines. International Journal of Molecular Sciences. 2023; 24(8):6918. https://doi.org/10.3390/ijms24086918
Chicago/Turabian StylePonomartsev, Nikita, Danil Zilov, Ekaterina Gushcha, Alexandra Travina, Alexander Sergeev, and Natella Enukashvily. 2023. "Overexpression of Pericentromeric HSAT2 DNA Increases Expression of EMT Markers in Human Epithelial Cancer Cell Lines" International Journal of Molecular Sciences 24, no. 8: 6918. https://doi.org/10.3390/ijms24086918
APA StylePonomartsev, N., Zilov, D., Gushcha, E., Travina, A., Sergeev, A., & Enukashvily, N. (2023). Overexpression of Pericentromeric HSAT2 DNA Increases Expression of EMT Markers in Human Epithelial Cancer Cell Lines. International Journal of Molecular Sciences, 24(8), 6918. https://doi.org/10.3390/ijms24086918